Abstract
In the last decades, pollinators have drastically declined as a consequence of anthropogenic activities that have local and global impacts. The food industry has been expanding intensive agriculture crops, many of them dependent on animal pollination, but simultaneously reducing native pollinator habitats. Chile is a good example of this situation. Chile is becoming an agro-alimentary powerhouse in Latin America, where intensive agriculture expansion is performed at the expense of natural lands, posing a major threat to biodiversity. Here, we discussed the drivers responsible for the decline of pollinators (including habitat loss, pesticides, invasive species, and climate change) and its synergistic effects. This is particularly critical considering that Chile is a hotspot of endemic bee species locally adapted to specific habitats (e.g., Mediterranean-type ecosystems). However, there is a lack of data and monitoring programs that can provide evidence of their conservation status and contribution to crop yields. Based on our analysis, we identified information gaps to be filled and key threats to be addressed to reconcile crop production and biodiversity conservation. Addressing the local context is fundamental to undertake management and conservation actions with global impact.
1. Introduction
The pollination crisis is a global problem. During the last decades, pollinators have drastically decreased as a result of major biodiversity loss drivers, namely land-use change, invasive species, overharvesting, pollution, and climate change [1,2]. From those five drivers, climate change operates at a global scale, but the remaining four operate at local scales, varying in time and space, with potential consequences at larger scales. Therefore, biodiversity change results from the combination of those drivers operating at different scales. Anthropogenic disturbances alter not only species occurrence and abundance, but their interactions as well. Thus, the loss of key ecological interactions (such as pollination and seed dispersal) has more insidious effects on biodiversity and ecosystem functioning than losing some particular species [3]. To address the current global pollination crisis, we also need to consider local anthropogenic impacts causing its decline and the associated local drivers.
Pollination services are crucial for food security and represent an ecosystem service worth USD 1 billion worldwide [4]. Although approximately 75% of crop species worldwide depend on animal pollination to some degree [5], the dominant conventional agriculture threatens pollinator conservation due to chemical contamination, habitat disturbance and loss, and the introduction of exotic and invasive species. Both agricultural production and biodiversity conservation are not mutually exclusive. On the contrary, biodiversity and food production can both benefit when including ecological processes and the ecosystem perspective in agricultural management [6,7]. Examples of these perspectives are agroecology [8,9], ecological intensification [10], and alternative agricultural schemes (e.g., biodynamic agriculture, natural farming, permaculture).
In this context, Chile is a good example of increasing dependence on animal-mediated crop pollination (Figure 1) while being subject to the drivers of Global Change with little regulation to protect its biodiversity [11]. As a food-producing country oriented toward export markets, Chile is uniquely positioned to contribute to a growing body of knowledge on pollination services. Chile exports pollination-dependent fruits and vegetables to many countries, mainly the USA, the EU, and China. Those exports represent an annual profit of ca. USD 3500 million (data from [12]). Fruit orchards alone cover about 112,725 ha [13], mostly established by replacing native vegetation [14]. Nonetheless, such large-scale production is often carried out at the expense of its biodiversity—a nefarious trade-off. A multidisciplinary approach to analyzing and dealing with the pollination crisis using Chile as a study case to address the local drivers of this crisis will also allow us a better understanding of the global pollinator crisis. Accordingly, this general review aims to assess the status of Chilean pollinators in relation to food production and elucidate the main threats to their long-term conservation and knowledge gaps that need to be prioritized to safeguard pollination services, based on a local perspective.
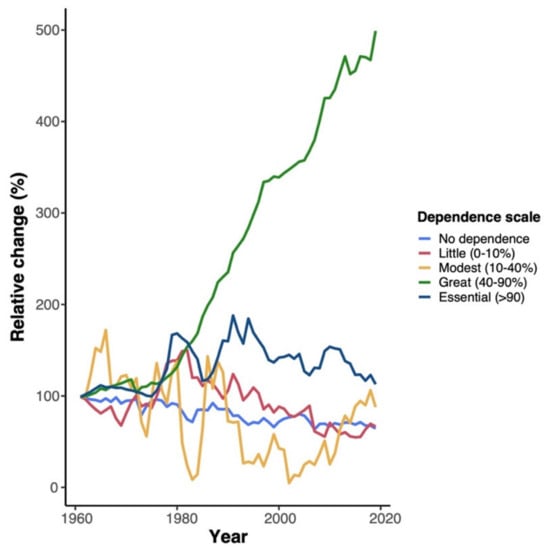
Figure 1.
Relative change (expressed as %) in pollinator-dependent crops in Chile between 1960 and 2020. Crop area was retrieved from FAOSTATS [15] and classified in five pollinator dependency categories following Klein et al. [5].
2. Pollinators in Agricultural Landscapes
Productive lands are not biological deserts. Many agricultural systems can harbor a significant proportion of the original species that inhabited the area before changing land use from natural to productive [16,17]. Both local and landscape effects have significant consequences on pollinators: landscape heterogeneity, non-crop habitat availability, local and landscape food resources, nesting habitats, and agricultural schemes, among others [18,19]. However, to optimize local and landscape management for pollinator promotion, the ecological requirements of these species must be known first. In Chile, although general knowledge of the local and landscape effects on pollinators are known based on global studies, specific ecological requirements at the species level (e.g., nesting habitat, plant resources, pollinator networks, dispersal distances) are scarce. However, a large dataset of 160 bee species (36% of bee species estimated to exist in Chile), including ecological features, was recently published by López-Aliste et al. [20]. The orchard agroindustry in Chile imports queens of the buff-tailed bumblebee Bombus terrestris [21], ignoring the pollination efficiency of this species and the real contribution to crop yield compared to the surrounding local pollinating species, such as Bombus dahlbomii.
On the other hand, massive and extensive agricultural practices can lead to biodiversity simplification as a result of land-use change [22,23], biotic homogenization (i.e., large monospecific crops), and the proliferation of invasive species (weeds, introduced pollinators). As a result of such biodiversity simplification, native pollinator populations are declining along with pollination services.
2.1. Native Pollinators
The services provided by native pollinators are not limited to native plant populations but are also relevant for crop plants. There is consensus that wild pollinators can contribute in different ways to improve pollination efficiency in crops (e.g., higher visitation rate, increase in pollen transfer, among others), and interfere with the behavior of foraging by honeybees, improving total pollination performance [24,25,26]. Moreover, it has been observed that an increase in pollinators’ diversity can favor fruit quantity and the quality of the harvests, favoring the final production and, therefore, the economic return for the producer (e.g., [27,28]).
Unfortunately, the contribution of native pollinators to crop production has been scarcely explored in Chile, especially of non-bee species. Studies on crop pollination by native insects have been performed in greenhouse tomatoes [29], avocados [30,31], and fava beans [32]. However, non-crop species pollination has also given some insights into native pollinators’ performance [33,34,35]. For example, Rego et al. [35] reported syrphids as the most frequent pollinators and those that carried the highest amount of pollen grains of Gomortega keule (an endangered tree from Central Chile), both in native and low-intensity agriculture systems, revealing the importance of dipteran pollinators to the fructification and persistence of this native endangered tree species, even in managed landscapes. Similarly, Murúa et al. [34,36] have shown the pivotal role of native oil-collecting bees for plant reproduction and the maintenance of reproductive barriers in two sympatric oil-secreting Calceolaria species inhabiting the high Andes Mountain range in Chile.
The potential importance for agriculture of the native bees in Chile is enormous, with 464 bee species described (Figure 2), of which 70% are endemic [37] and mainly solitary [20]. Thus, it is relevant to research basic biology and ecology of native solitary bee species (e.g., diet breadth and nesting biology) to promote research programs that allow the selection and of domestic species that may provide pollination service demanded by the agriculture practice. In Chile, 132 syrphid species have been described [38]. Other relevant pollinator species for agriculture, belonging to other dipteran families, have been rarely studied. Plant–pollinator interactions have been even less studied. Although a large proportion of native bees are restricted to the Mediterranean region, a recent review identified a total of 69 studies on plant–pollinator interactions in this region [39]. In this review, flower visitors were described for 162 genera belonging to 58 families and 8 orders, and the topics addressed varied from plant traits effects on pollination to community and network assessments. However, none of these studies have addressed the contribution of native pollinators to crop yields. A study conducted further south in the Temperate Rainforest in southern Chile identified plant visitors associated with 26 plant species. The most represented group was Diptera with 75 species, followed by Coleoptera with 56 species and Hymenoptera with 30 species [40]. The same study identified four bird species and two Lepidoptera species visiting flowers as well. Despite of this species richness, we did not find studies addressing pollinator contribution to crops in that region.

Figure 2.
Examples of common bee species in Chile (Photos: Luis Flores-Prado).
Plant–pollinator interactions are usually disturbed when exotic plant species are introduced. In Chile and other Latin American countries, pollinator networks are characterized by having many exotic species [41]. Not only are managed pollinators exotic but also other wild pollinators (e.g., Bombus ruderatus, Xylocopa augusti, X. splendidula, Eristalinus taeniops). On the other hand, almost all pollinator-dependent crops in Chile are exotic (some exceptions are Ugni molinae, Aristotelia chilensis, and Berberis microphylla), and a high proportion of wild plant species have also been introduced in the last centuries [42]. This dominance of exotic species generates additional complexity for the conservation of pollinators since it increases resource competition [43] and favors the arrival of novel pathogens [44] to native species.
2.2. Managed Pollinators
Crop pollination is primarily enhanced by placing honeybee (Apis mellifera) colonies within the target crop during blooming. This species has been proposed as the essential pollinator for crops and native plants [45]. Crop pollination is mediated by A. mellifera to a high percentage in the USA [46] and the European Union (see review in [47]). Thus, pollination of many crops in most parts of the world relies on honeybees, whose pollinating activity can be greater, in some plant species, than pollination provided by native bee species [48]. However, in many cases, native pollinators are more effective [26]. Several crops have also incorporated colonies of the buff-tailed bumblebee (Bombus terrestris) in the last decades, imported mainly from Europe.
Worldwide, honeybee colonies have increased at lower rates than crop pollination demand [49]. In Latin America, such an increase is very little compared to the global trend [50]. In Chile, honeybees are managed for honey production and pollination services. Their annual stocks are reported by the FAOSTATS database [15], but since 2015, new Chilean regulation has required beekeepers to register their colonies at a State agency (SAG, Spanish acronym of the Chilean Agricultural and Livestock Service), which monitors their numbers based on these registries (Supplementary Figure S1). Comparing both sources, we found important inconsistencies between them. Between the years 2016–2019, FAO [15] reports between 445,788 and 452,377 colonies annually, while SAG [51] records range between 554,463 and 688,871. According to surveys based on interviews with beekeepers, honeybee declines have been reported to be 19.7 (95% confidence interval of 15.6–23.9) and 5.9 (95% confidence interval of 2.9–8.8) each year in 2014–2015 and 2015–2016, respectively [50]. It is difficult to provide explanations for these temporal inconsistencies.
In addition to honeybees, buff-tailed bumblebee (Bombus terrestris) has been increasingly adopted by farmers. Since 1997, this species has been imported to Chile every year to date (Figure 3). Although it has currently invaded a great portion of the country, including the whole distribution area of the only native bumblebee of central and southern Chile (Bombus dahlbomii), it is expected to continue its expansion [52]. Other managed pollinators are rare to find, such as the fly Lucilia sericata, used to pollinate some crops in central Chile.
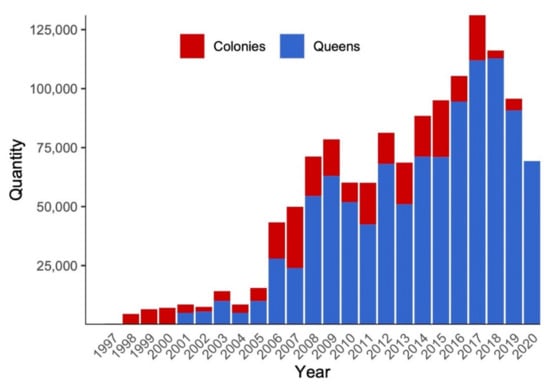
Figure 3.
Bombus terrestris colonies and queens imported to Chile from 1997 to 2020 [51].
2.3. Crop Dependence on Pollination
Most of the food we consume depends on biotic pollination [5]. In Chile, when analyzing FAOSTAT [15] crop area data by classifying crops according to their animal pollination dependency [5], we found that 27% of the total crop area in 2019 was cultivated with crops that depend on some degree on animal pollination. Of these, canola, cherry, and apple are the crops with the highest amount of harvested area with 15, 12, and 10%, respectively (Supplementary Table S1). However, pollinator-dependent crops have dramatically increased in the last decades, while non-dependent crops have decreased (Figure 1). In particular, crops with “great” pollination dependency (i.e., in the absence of pollen vectors, yields decrease between 40 and 90%) have continuously increased since the 1980s, from 46,230 ha in 1980 to 174,300 ha in 2019. In this group, the crop species with the highest increase are apples, avocados, and cherries (Supplementary Figure S1). Other relevant pollinator-dependent crops in this category are blueberries, plums, peaches, and nectarines.
There is no unified spatial crop species database in Chile that allows us to conduct an exhaustive analysis of crop pollination dependency in a spatially explicit manner. However, data regarding fruit crops are available by alternative monitoring schemes. We used the last Agricultural Census data from 2007 (the last available dataset of its kind) to consider all possible pollinator-dependent crops in Chile, classifying crops dependency on animal pollination based on Klein et al. [5]. This database aggregates spatial crop data in polygon areas. Therefore, we filtered these with a land-use database generated by Zhao et al. [53] to detect crop areas and other land uses. The demand for crop pollination is concentrated in the Central Valley area of central Chile (Figure 4). This is consistent with the concentration of pollinator-dependent crops extracted for each administrative region according to ODEPA-CIREN [54]. This database integrates the most recent available crop data (mostly from 2018 to 2020) in which central Chile shows the highest concentration of these crops, while the area of natural forest (i.e., native forest or mixed forest), shrubland, and pastures, is relatively lower (Supplementary Figure S2). This region is exposed to high anthropogenic pressure concentrating most of the Chilean population, partly explaining lower amounts of protected areas for conservation. Although subject to climate change disturbances, climate conditions in the area are optimal for the several Mediterranean and temperate crops. The vast majority of crop production in Chile is under a conventional management scheme. Organic crops are more concentrated in fruit production, reaching approximately 3% of fruit crop area in 2017 [55]. Chilean farmland is currently dominated by export-oriented large farms (>100 ha) that occupy almost 89% of the area, while small farms (<20 ha), although large in number (73% of total farms), occupy less than 4% of this area [56]. The non-crop area is scarce except for sites located next to the mountain areas (Figure 4). In addition, the pressure of exotic tree species plantations, such as Pinus radiata and Eucalyptus globulus, is high in several areas. These forest plantations consist almost exclusively of exotic monocultures with intensive management, which generates high soil erosion, suppresses other plants and harvests through clear-cutting. Therefore, these areas are probably not highly suitable habitats for pollinators. Areas with highly dependent animal pollinated crops and little suitable pollinator habitat are likely the areas with higher risks for yields limited by pollination.
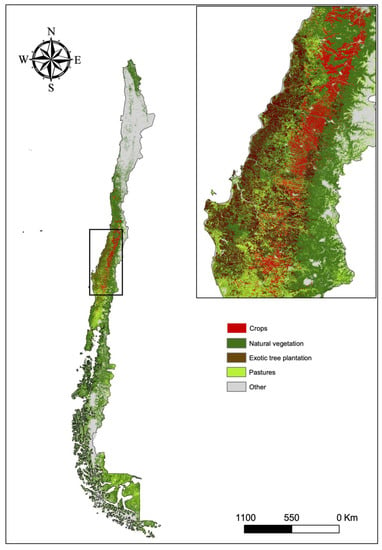
Figure 4.
General land cover map for Chile [53]. Land Use/Land Cover was classified into crops, natural vegetation (forests or shrubland), exotic tree plantations, pastures (natural or managed), and other (wetlands, water, impervious surface, bare soil, snow, ice). The inset highlights the crop area (in red) in central Chile.
The exponential increase in pollination-dependent crops along with pollinator declines stimulate the increasing demand for managed pollinators, which consist almost exclusively of Apis mellifera and imported Bombus terrestris (detailed in Section 2.1).
3. Drivers of Pollination Decline
During the last few decades, A. mellifera colonies have been affected by the so-called Colony Collapse Disorder (CCD hereafter) syndrome. This phenomenon has affected bee colonies on different continents [57]. CCD is disturbing because no single cause has emerged as a conclusive explanation. CCD involves the interaction between pathogens and other stress factors, as well as the sub-lethal effect of pesticide residues and climate change [58,59,60]. Moreover, it has been proposed that some native bees have experienced a decline in population size and species richness in several regions of the world during the last few decades [61,62]. This decline seems to be mainly related to human disturbances causing habitat loss and fragmentation, pesticide exposure, simplifying biodiversity through intensive agriculture, and facilitating the spread of invasive species via multiple introductions [58,63]. Those effects are enhanced by the impact of climate change [64]. In Latin America, change in land use associated with intensive agriculture and the constant introduction of managed pollinators (that rapidly turn into invasive species) are the most important causes of local bee decline [65,66].
3.1. Habitat Loss
The Chilean agriculture industry has rapidly expanded at the expense of natural habitats. A broad analysis for each administrative Region in Chile reveals that Regions with higher pollinator-dependent crops have less native forest and State-protected areas than other regions where agriculture is incipient (Supplementary Figure S2), revealing not only the impact of agriculture expansion but also the low commitment of the Chilean State to protect the remnants of vegetation and species diversity present in those regions.
There is no exhaustive analysis of native forest loss in Chile. However, Miranda et al. [67] integrated all available information based on previous studies that covered 36.5% of the Chilean Biodiversity Hotspot located in Central and Southern Chile, and revealed that between 1973 and 2011, 19% of native forest (782,120 ha) was lost and converted to shrubland, exotic tree plantations, and agriculture/pastureland. This land use and cover change was higher in Central Chile, between 35.5° S and 40.0° S, where native forest was mostly replaced by exotic tree plantations. Consequently, the remaining natural habitats have undergone an attrition process (following the definition of Lindenmayer and Fischer [68]), in which habitat fragments become smaller, fewer, and more isolated. Extensive agriculture encroaches over the landscape, reducing natural habitat cover and leaving the extant remnants surrounded by a heterogeneous matrix that increases edge effects [69]. Thus, many wild pollinators are lost due to the destruction of natural habitats, while others visit or inhabit croplands [70]. Habitat loss and fragmentation processes directly and indirectly impact native pollinator species that have a beneficial role in productive lands, and reduce the diversity and abundance of other animal species that act as natural enemies and control crop pests [71,72,73]. This inevitably leads to increasing the use of pesticides (see Section 3.2 below) and managed pollinators that may become invasive (see Section 3.3 below), enhancing biodiversity loss problems and negatively impacting crop yields [74].
3.2. Pesticides
Extensive monospecific crop fields are more vulnerable to being attacked by different kinds of pests (e.g., herbivorous insects), which reduce crop yields and cause important economic losses [75]. As natural enemies become less diverse and abundant due to the loss of natural habitats near crop fields, pesticides are a common practice to control crop pests. Such chemical substances effectively control pest insects but do not discriminate pests from non-pest insects (e.g., pollinators). The indiscriminate application of pesticides with little regulation from the local authorities is common in Latin America [76]. As crop pests become resistant to chemical control, farmers use higher concentrations and apply pesticides more frequently. Particularly, neonicotinoid pesticides are widely used due to their high effectiveness [2] and are known to have sub-lethal effects on pollinators [77,78,79]. Because of this, on several occasions, pesticides effects have been withdrawn since they do not exert a noticeable effect on bee abundance, but they can compromise pollination effectiveness due to the neurological and physiological damage caused by those chemical agents. Affected bees become disoriented, losing track of floral resources and the hives [80], altering their reproduction and longevity [81,82]. Moreover, recent studies have revealed important indirect effects of different pesticides (including fungicides, insecticides, and herbicides) on pollinators’ health through their impacts on gut microbiota [83,84]. It has been observed that a decrease in bacterial diversity reduces the immune response against pathogens but also affects the metabolism and degradation of carbohydrates and sugar [85]. Recently, Henríquez-Piskulich et al. [86] described 44 pesticides used in Chile but banned by the European Union, all producing damage to bees. Although Chilean law demands that the farmers must notify the agricultural authority before using pesticides, this is only required when pesticides are applied near honeybee hives (Act N°158/2015). In addition, farms are required to keep records on pesticides applications, but these data are not centralized by any state agency and are thus unreachable for the general public. In this sense, Chile must to coordinate studies that document the direct impacts of pesticides on native pollinators and the indirect impacts on fruit production, and to reinforce laws that reduce the use of harmful pesticides.
3.3. Invasive Species
As native pollinators decline, farmers supplement crops with managed pollinators, which are exotic species intentionally introduced for this purpose. The honeybee Apis mellifera and the bumblebee Bombus terrestris are the most common managed species introduced for crop pollination in Chile. A recent study in Central Chile found an influence of honeybee abundance interaction with patch area on native bee visitation rates, but no effects of honeybee abundance on native bee species richness [87]. In southern Chile, a long-term monitoring of a pollinator assemblage for 10 years suggests that honeybee abundance oscillates with B. dahlbomii (a native bumblebee), and that its abundance negatively affects the abundances of less frequent insect visitors of the studied tree, Eucryphia cordifolia [88]. Whereas honeybee introduction has a long history, and this species is now well-integrated into most plant–pollinator networks [89], bumblebee introduction is more recent. Bombus terrestris was introduced in Chile in 1997 and was intended for greenhouse pollination only. However, a few years later, the Chilean authorities allowed its use in the open field, leading to a rapid invasion process across southern South America [65]. Although Argentina has banned B. terrestris introduction, this country was severely affected by the rapid expansion of this invasive bumblebee from Chile, and its populations are constantly supplemented by new introductions [21].
The rapid expansion of B. terrestris was followed by a replacement of the dominant native pollinators in plant–pollinator networks [90] and a consequent biological homogenization [41]. In addition, this species has rapidly replaced the native bumblebee Bombus dahlbomii [43] as a consequence of the horizontal transmission of alien parasites [91,92]. Bombus terrestris is a super-generalist species and, for that reason, is frequently used for crop pollination [93]. When crops are blooming, managed pollinators concentrate their feeding activities within the crop fields. However, when flowering is over, there is a spillover effect over neighboring natural areas [94], facilitating the invasion process. The consequences of this are noticeable in the structure of plant–pollinator networks [95], having cascade effects across the invaded communities. In addition, B. terrestris might act as an inefficient pollinator of the native flora, attributable to their nectar robbing behavior [96,97] and poor pollen carrier capacity [98]. A less pervasive effect of B. terrestris is that it may promote plant invasion because it actively visits exotic plants [99], favoring their reproduction and reinforcing the spread process [100], which recursively might increase weeds and invasive pollinator availability for crops. Moreover, in high mountain environments, B. terrestris has been observed to disrupt prezygotic reproductive barriers between native plant species by interspecific pollen exchange, favoring the development of infertile hybrids [36]. Undoubtedly, these complex interactions need to be revisited to determine the causes and consequences of B. terrestris invasion and stop its spread.
3.4. Climate Change
Chile is one of the most vulnerable countries to climate change [101]. Despite accounting for a small fraction of the worldwide greenhouse gas emissions, Chile is expected to suffer drastic consequences of climate change, primarily associated with the increase of magnitude and frequency of drought events [102,103]. This situation directly impacts crop yield and indirectly pollinators, both native and introduced, due to the reduction of floral resources and increased plant mortality. The effects of drought and water stress on flower production were recently reported for the temperate rainforests of South America [104]. However, the same pattern, associated with crop fields, has been reported in the United Kingdom [64]. Most of the agricultural production in Chile is concentrated in the central region, where drought effects are more pronounced [103]. Extensive agricultural activities consume most of the freshwater resources in central Chile at the expense of the natural habitats and local populations [105]. Nevertheless, the dramatic water shortage conditions of the last decade have compromised crop yield despite the privileged access to water resources that the productive sector has (in Chile, water resources are private and granted as perpetual concessions to particular owners [106]). Despite the high impact that climate change has on Chilean crops, we found no studies assessing its effects on animal crop pollination. The impacts of extreme drought events on a specialized hummingbird-pollinated plant species have been assessed, providing evidence of the complexity associated with climate change effects on a plant–pollinator interaction [107]. Beyond crop fields, climate change-induced drought events are responsible for the rapid deterioration of natural habitats, increasing plant stress [108] and plant mortality as well [109], generating massive mortality that affects plant communities, a process known as vegetation browning [110]. With the floral resource reduction, this rapid deterioration of the natural vegetation puts native pollinators in check, negatively impacting native plant species pollination and indirectly impacting crop pollination. A dramatic example of this situation is the relict tree Pouteria splendens, a narrow endemism of coastal Mediterranean-type ecosystem in Chile [111], which was severely impacted by a prolonged drought [112], causing a drastic reduction in the number of flowers produced and impacting the flower visitor insects (CJP unpublished data).
In addition, abrupt changes in temperatures, precipitation, and wind patterns can disrupt plant–pollinator interactions through a change in pollinator foraging activities and species abundances [113,114]. For example, in a study on different almond orchards pollinated by managed and wild bees, Pitts-Singer et al. [115] observed that some individuals do not fly at low temperatures, altering their visit frequency. Concomitant with this, they found a reduction in fruit yield in the studied years, suggesting that low temperatures and high humidity can affect pollination efficiency and production as a whole [115]. In addition, the authors suggest that these unfavorable climatic conditions can also alter the nesting process, reducing pollinator abundance inside the orchards. Indirect effects, mediated by reducing pollination services in the new dry conditions, require quantifying extinction risks in plants and pollinators.
4. Gaps and Future Challenges
Based on the presented diagnostic, we identified major gaps and key threats that need to be addressed to tackle the pollination crisis in Chile, using local actions to tackle a global problem. This section discusses some challenges for the knowledge gaps and threats that will continue to affect crop pollination.
4.1. Knowledge Gaps on Pollinators and Pollination in Chile
The absence of information is probably one of the most important gaps that affect decision-making toward effective management and conservation actions. Thus, to understand how pollination services are being affected in this changing world scenario, we need more and better data. We have identified major data gaps for Chile, which preclude us from making informed decisions and suggesting appropriate actions to tackle this crisis. Recently, a large database of wild bees has been published [20], dramatically increasing the amount of freely available information. However, there are still major information gaps to be covered, as this large database contains information on 167 out of 464 bee species described for Chile (leaving 64% of the bee species with no reported evidence). Further, if we consider that bees are a relatively well-studied group among insects, other relevant pollinator groups, such as hoverflies (Syrphidae) and butterflies, remain largely understudied. Their role in safeguarding pollination services in productive lands is rarely assessed. In addition, less explored is the contribution of vertebrate pollinators, such as hummingbirds [107], to crop pollination. For instance, Sephanoides sephaniodes and Patagona gigas in Chile, which are considered as super-generalist visitors of the native flora [116,117,118,119], frequently visit flowers in orchards (e.g., almond and citrus plants; GOC pers. obs.). Thus, they might supply pollen and could potentially be used as managed pollinators. In addition, this knowledge gap is closely related to lack of taxonomists due to limited incentive to the development of taxonomy as a discipline in Chile, which is part of a global phenomenon [120]. Currently, research funds available in Chile do not fund studies whose objective is increasing taxonomic knowledge and classification of species. This undoubtedly prevents the advance of this discipline in Chile, discourages the training of new specialists, and leaves many taxonomic groups with no taxonomist working on them.
Landscape management and ecological intensification arise as important mechanisms to increase pollination in orchards [121]. Intercropping and the use of flowering plants as “magnet species” (such plant species that attract pollinators and increase pollination of surrounding plants [122,123]) are strategies to improve pollination richness and fruit production because these plants provide animal refuge and interconnect patches [124]. However, the use of magnet species has some disadvantages. For instance, floral phenology could show a mismatch with the focal crop species, or the magnet species might dilute pollination of focal species [124]. In Chile, some agricultural enterprises, especially vineyards, use ecological intensification to give aggregate value to their products sold as “organic” in markets [125]. Unfortunately, one of the main strategies to give “organic” status to crops is to cover vineyards with exotic species [126]. This is a nefarious method because it promotes biological invasions and might cause a decline in pollinators in the long term. Unequivocally, it is necessary to include a monitoring network scheme to assess the effectiveness of the different strategies used by agricultural enterprises to manage pollination. Monitoring plans are an important but often neglected component, and the research plans derived from them are often impacted by the missing contributions of key parameters. Therefore, having appropriate monitoring schemes (i.e., standardized schemes collecting the information that is really needed) will allow us to make informed decisions to improve pollination in productive activities.
We cannot rely solely on research projects to cover that enormous information gap on pollinator diversity and distribution. Therefore, we also need to explore other initiatives, such as citizen science to gather information. Contrary to what happens in the USA or Europe, where citizen science endeavors have been conducted since a long time ago, these initiatives began in Chile about 10 years ago. In this regard, open platforms such as iNaturalist are gaining relevance, and many people (experts and non-experts) use them to record species occurrence (Figure 5). The information, gathered via the citizen science initiatives, revealed the important contribution of such activities in developing knowledge at large scales [127]. However, implementing a good citizen-science monitoring scheme for pollinators can be a complex endeavor [128]. On the one hand, most of the pollinator-monitoring endeavors are biased toward honeybees and crop pollination, while native pollinators and pollination interactions in natural areas remain largely underrepresented. On the other hand, even in countries with a long citizen science history (e.g., the USA and many European countries), the lack of taxonomic expertise can be a major shortcoming, biasing citizen-science records against native and rare species [128].
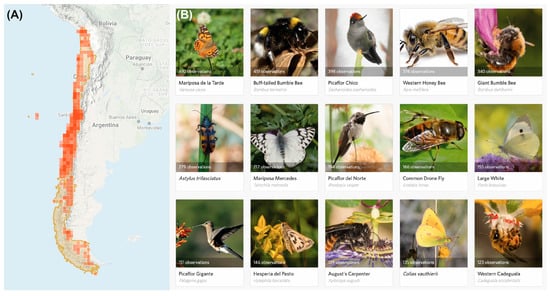
Figure 5.
(A) iNaturalist records of pollinator species in Chile (squares indicate sites with pollinator records and color intensity is proportional to the number of records in three tones corresponding to 1, 10, and 100 records); (B) the most common pollinator species recorded in iNaturalist (data from the Chilean Pollination Network project, updated 15 September 2021).
Thus, a comprehensive monitoring—that includes citizen science—will allow to assess and understand the deep sense of commitment and the great level of awareness that local stakeholders (citizens, farmers, authorities) have regarding the pollination crisis. We have synthesized those gaps in Table 1.

Table 1.
Summary of key knowledge gaps, their descriptions, and opportunities.
4.2. Future Challenges
Paradoxically, one of the main threats to pollination services and pollinators in crops is expanding productive lands. Land-use changes are rapidly homogenizing landscapes and reducing extant natural areas, compromising the long-term persistence of wild pollinators. We cannot safeguard pollination services and conserve pollinator species without guaranteeing the conservation of natural habitats and key ecological interactions. Therefore, the conservation of the extant natural habitats (particularly those from central Chile, where most agricultural activities are conducted) should be prioritized.
Insecticides have long been considered a major culprit in pollinator decline. A particular group of insecticides, the neonicotinoids, disperse easily through the environment and are found many kilometers from their target areas. In addition, these pesticides are absorbed in plant tissues, disrupting the pollinator physiology [129,130,131]. Newer insecticides, such as flupyradifurone and sulfoxaflor, were considered good candidates to replace neonicotinoids. However, recent studies have shown that these insecticides also have non-lethal negative effects on native pollinators [131]. In Chile, agricultural regulation bans the use of certain insecticides [132], but the chemical composition of the approved insecticides has not been tested for non-lethal effects on non-target species. This information gap is particularly concerning as most insecticides can disperse freely through the environment, creating negative synergistic and cascading effects.
Finally, to make real strides in biodiversity conservation, we need to know and understand pollinators. Education is, therefore, a key factor in slowing biodiversity loss [133]. Investing in education raises awareness of the environmental problems within the society, which is fundamental to forge positive attitudes toward biodiversity. A few projects have highlighted the importance of native pollinators, most notably the Chilean bumblebee (Bombus dahlbomii), which has had measurable success in the regions where it was implemented (e.g., “Salvemos nuestro abejorro” (Let’s save our bumblebee) initiative; the results from this initiative are available at https://salvemosnuestroabejorro.wordpress.com). Highlighting the importance of other pollinator species in this way could help to slow pollinator decline.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d13110571/s1, Table S1: Crop area harvested, Figure S1: Honeybee stocks by year, Figure S2: Regional comparison of pollination-dependent crops.
Author Contributions
Conceptualization, F.E.F., L.V., M.M.M., L.F.-P. and G.O.C.; resources, M.L.-A., C.J.-P. and C.A.; data curation, M.L.-A., F.E.F., L.V., M.M.M., L.F.-P., C.E.V. and G.O.C.; writing—original draft preparation, L.V., M.M.M., L.F.-P., G.O.C., G.M. and F.E.F.; writing—review and editing, L.V., M.M.M., L.F.-P., G.O.C., C.E.V., G.M., C.J.-P., C.A. and F.E.F.; visualization, L.V., L.F.-P. and F.E.F.; supervision, F.E.F.; project administration, F.E.F.; funding acquisition, G.O.C. and F.E.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by the Chilean Agency for Research and Development (ANID), grant number ANID/PCI/NE/S011870/1.
Acknowledgments
We are grateful to all our collaborators within the SURPASS Chile project, and particularly to Javiera Díaz, who leads the project’s communication program. Comments of two anonymous reviewers improved the manuscript. C.J.P. acknowledges the ANID 21150472 doctoral fellowship.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Biodiversity—Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES secretariat: Bonn, Germany, 2019. [Google Scholar]
- Valiente-Banuet, A.; Aizen, M.A.; Alcantara, J.M.; Arroyo, J.; Cocucci, A.; Galetti, M.; Garcia, M.B.; Garcia, D.; Gomez, J.M.; Jordano, P.; et al. Beyond species loss: The extinction of ecological interactions in a changing world. Funct. Ecol. 2015, 29, 299–307. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. P Roy. Soc. B-Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Perfecto, I.; Vandermeer, J.; Wright, A. Nature’s matrix. Linking agriculture, biodiversity conservation and food sovereignty; Routledge: London, UK, 2019. [Google Scholar]
- Altieri, M.A.; Nicholls, C. Agroecology: Challenges and opportunities for farming in the Anthropocene. Int. J. Agric. Nat. Resour. 2020, 47, 204–215. [Google Scholar]
- Altieri, M.A. Agroecology: The Science of Sustainable Agriculture, 2nd ed.; Westview Press: Boulder, CO, USA, 2015. [Google Scholar]
- Gliessman, S.R. Agroecology: The Ecology of Sustainable Food Systems, 3rd ed.; CRC Press Taylor & Francis: Boca Raton, FL, USA, 2015. [Google Scholar]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Schutz, J. Creating an integrated protected area network in Chile: A GIS assessment of ecoregion representation and the role of private protected areas. Environ. Conserv. 2018, 45, 269–277. [Google Scholar] [CrossRef]
- Pérez, R.; Valdés, A. Análisis de la Evolución y Desempeño Agroexportador de Chile, 2000–2017; Oficina de estudios y Políticas Agrarias—ODEPA, 2019. Available online: https://www.odepa.gob.cl/wp-content/uploads/2019/04/EvolucionAgroExp.pdf (accessed on 28 April 2021).
- Pefaur, J. Evolución de la Fruticultura Chilena en los Últimos 20 años; Oficina de estudios y Políticas Agrarias—ODEPA, 2020. Available online: https://bibliotecadigital.odepa.gob.cl/bitstream/handle/20.500.12650/70234/evolucionFruticulturachilena.pdf (accessed on 28 April 2021).
- Schulz, J.J.; Cayuela, L.; Echeverria, C.; Salas, J.; Benayas, J.M.R. Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Appl. Geogr. 2010, 30, 436–447. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Statistics Database (updated March 2021); Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef]
- Firbank, L.G.; Petit, S.; Smart, S.; Blain, A.; Fuller, R.J. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Philos. T. R. Soc. B 2008, 363, 777–787. [Google Scholar] [CrossRef]
- Lonsdorf, E.; Kremen, C.; Ricketts, T.; Winfree, R.; Williams, N.; Greenleaf, S. Modelling pollination services across agricultural landscapes. Ann. Bot. 2009, 103, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef] [PubMed]
- López-Aliste, M.; Flores-Prado, L.; Ruz, L.; Sepúlveda, Y.; Rodríguez, S.; Saraiva, A.M.; Fontúrbel, F.E. Wild bees of Chile: A database on taxonomy, sociality, and ecology. Ecology 2021, 102, e03377. [Google Scholar] [CrossRef] [PubMed]
- Smith-Ramirez, C.; Vieli, L.; Barahona-Segovia, R.M.; Montalva, J.; Cianferoni, F.; Ruz, L.; Fontúrbel, F.E.; Valdivia, C.E.; Medel, R.; Pauchard, A.; et al. The reasons why Chile should stop importing commercial bumblebee Bombus terrestris (Linnaeus) and to start controlling it. Gayana 2018, 82, 118–127. [Google Scholar] [CrossRef]
- Mogren, C.L.; Rand, T.A.; Fausti, S.W.; Lundgren, J.G. The effects of crop intensification on the diversity of native pollinator communities. Environ. Entomol. 2016, 45, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.a.; Steffan-Dewenter, I. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land-use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, G.P.; Buchmann, S.L. Services provided by pollinators. In Nature’s Services: Societal Dependence on Natural Ecosystems; Daily, G.C., Ed.; Island Press: Washington, DC, USA, 1997; pp. 133–150. [Google Scholar]
- Eeraerts, M.; Smagghe, G.; Meeus, I. Pollinator diversity, floral resources and semi-natural habitat, instead of honey bees and intensive agriculture, enhance pollination service to sweet cherry. Agr. Ecosyst. Environ. 2019, 284, 106586. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Truslove, C.; Coston, D.; Evans, R.; Moss, E.; Dodson, C.; Jenner, N.; Biesmeijer, J.; Potts, S. Pollination deficits in UK apple orchards. J. Pollinat. Ecol. 2014, 12, 9–14. [Google Scholar] [CrossRef]
- Miñarro, M.; Somoano, A.; Ventura, J. Intra-annual continuous reproduction of the apple pest Microtus lusitanicus: Implications for management. Crop. Prot. 2017, 96, 164–172. [Google Scholar] [CrossRef]
- Estay, P.; Wagner, A.; Escaff, M. Evaluation of Bombus dahlbomii (Guér.) as a pollinating agent for tomato flowers under greenhouse conditions. Agricultura Técnica 2001, 61, 113–119. [Google Scholar] [CrossRef]
- Monzón, V.H.; Avendano-Soto, P.; Araujo, R.O.; Garrido, R.; Mesquita-Neto, J.N. Avocado crops as a floral resource for native bees of Chile. Rev. Chil. Hist. Nat. 2020, 93, art5. [Google Scholar] [CrossRef]
- Muñoz, A.E.; Plantegenest, M.; Amouroux, P.; Zaviezo, T. Native flower strips increase visitation by non-bee insects to avocado flowers and promote yield. Basic Appl. Ecol. 2021, 56, 369–378. [Google Scholar] [CrossRef]
- Smith-Ramirez, C.; Rendón-Funes, A.; Barahona-Segovia, R.; Moya, W. Consequences of the high abundance of Bombus terrestris on the pollination of Vicia faba. J. Pollinat. Ecol. 2021, 29, 258–272. [Google Scholar] [CrossRef]
- Lander, T.A.; Harris, S.A.; Boshier, D.H. Flower and fruit production and insect pollination of the endangered Chilean tree, Gomortega keule in native forest, exotic pine plantation and agricultural environments. Rev. Chil. Hist. Nat. 2009, 82, 403–412. [Google Scholar] [CrossRef]
- Murúa, M.; Cisterna, J.; Rosende, B. Pollination ecology and breeding system of two Calceolaria species in Chile. Rev. Chil. Hist. Nat. 2014, 87, art7. [Google Scholar] [CrossRef]
- Rego, J.O.; Schlindwein, C.; Garrido, R.; Monzón, V.H. Low fruit set in an endangered tree: Pollination by exotic bumblebees and pollen resource for relictual native bees. Arthropod-Plant. Interact. 2021, 15, 491–501. [Google Scholar] [CrossRef]
- Murúa, M.; Espindola, A.; González, A.; Medel, R. Pollinators and crossability as reproductive isolation barriers in two sympatric oil-rewarding Calceolaria (Calceolariaceae) species. Evol. Ecol. 2017, 31, 421–434. [Google Scholar] [CrossRef]
- Montalva, J.; Ruz, L. Actualización de la lista sistemática de las abejas chilenas (Hymenoptera: Apoidea). Revista Chilena Entomología 2017, 35, 15–52. [Google Scholar]
- Barahona-Segovia, R.M.; Riera, P.; Paninao-Monsalvez, L.; Guzmán, V.V.; Henriquez-Piskulich, P. Updating the knowledge of the flower flies (Diptera: Syrphidae) from Chile: Illustrated catalog, extinction risk and biological notes. Zootaxa 2021, 4959, 1–178. [Google Scholar] [CrossRef]
- Medel, R.; González-Browne, C.; Fontúrbel, F.E. Pollination in the Chilean Mediterranean-type ecosystem: A review of current advances and pending tasks. Plant Biol. 2018, 20, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Smith-Ramirez, C.; Martinez, P.; Nuñez, M.; González, C.; Armesto, J.J. Diversity, flower visitation frequency and generalism of pollinators in temperate rain forests of Chiloe Island, Chile. Bot. J. Linn. Soc. 2005, 147, 399–416. [Google Scholar] [CrossRef]
- Aizen, M.A.; Morales, C.L.; Morales, J.M. Invasive mutualists erode native pollination webs. PLoS Biol. 2008, 6, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, N.; Sánchez, P.; Pauchard, A.; Urrutia, J.; Cavieres, L.; Marticorena, A. Plantas Invasoras del Centro-sur de Chile; Instituto de Ecología y Biodiversidad (IEB): Santiago de Chile, Chile, 2014. [Google Scholar]
- Morales, C.L.; Arbetman, M.P.; Cameron, S.A.; Aizen, M.A. Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 2013, 11, 529–534. [Google Scholar] [CrossRef]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathé, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef]
- Aebi, A.; Neumann, P. Endosymbionts and honey bee colony losses? Trends Ecol. Evol. 2011, 26, 494. [Google Scholar] [CrossRef][Green Version]
- Committee on the Status of Pollinators in North America, N.R.C. Status of Pollinators in North. Ame; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Williams, I.H. Insect Pollination and Crop Production: A European Perspective. In Pollinating Bees—The Conservation Link Between Agriculture and Nature; Kevan, P., Imperatriz-Fonseca, V.L., Eds.; Ministry of Environment: Brasília, Brazil, 2002; pp. 59–65. [Google Scholar]
- Rader, R.; Howlett, B.G.; Cunningham, S.A.; Westcott, D.A.; Newstrom-Lloyd, L.E.; Walker, M.K.; Teulon, D.A.; Edwards, W. Alternative pollinator taxa are equally efficient but not as effective as the honeybee in a mass flowering crop. J. Appl. Ecol. 2009, 46, 1080–1087. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef]
- Requier, F.; Antúnez, K.; Morales, C.L.; Aldea Sánchez, P.; Castilhos, D.; Garrido, P.M.; Giacobino, A.; Reynaldi, F.J.; Rosso Londoño, J.M.; Santos, E. Trends in beekeeping and honey bee colony losses in Latin America. J. Apic Res. 2018, 57, 657–662. [Google Scholar] [CrossRef]
- SAG. Response Letter Nº 3462 (May 26th 2021) by Request Under the Law 20.285 of Access to Public Information; Chilean Ministry of Agriculture: Santiago de Chile, Brazil, 2021.
- Fontúrbel, F.E.; Murúa, M.M.; Vieli, L. Invasion dynamics of the European bumblebee Bombus terrestris in the southern part of South America. Sci. Rep. 2021, 11, 1–7. [Google Scholar]
- Zhao, Y.Y.; Feng, D.L.; Yu, L.; Wang, X.Y.; Chen, Y.L.; Bai, Y.Q.; Hernandez, H.J.; Galleguillos, M.; Estades, C.; Biging, G.S.; et al. Detailed dynamic land cover mapping of Chile: Accuracy improvement by integrating multi-temporal data. Remote Sens. Environ. 2016, 183, 170–185. [Google Scholar] [CrossRef]
- ODEPA-CIREN. Estadísticas Productivas. 2021. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/estadisticas-productivas (accessed on 26 October 2021).
- Eguillor, P.M. Agricultura Orgánica, ODS y Cambio Climático: Mercado Internacional y Nacional. 2020. Available online: https://bibliotecadigital.odepa.gob.cl/bitstream/handle/20.500.12650/70558/Articulo%20AgriculturaOrganica.pdf (accessed on 26 October 2021).
- ODEPA-INE. VII Nacional Census of Agriculture and Forestry. 2007. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/ficha-nacional-y-regionales (accessed on 26 October 2021).
- Stokstad, E. The case of the empty hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- VanEngelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef]
- Flores, J.M.; Gámiz, V.; Jiménez-Marín, Á.; Flores-Cortés, A.; Gil-Lebrero, S.; Garrido, J.J.; Hernando, M.D. Impact of Varroa destructor and associated pathologies on the colony collapse disorder affecting honey bees. Res. Vet. Sci. 2021, 135, 85–95. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Murray, T.E.; Kuhlmann, M.; Potts, S.G. Conservation ecology of bees: Populations, species and communities. Apidologie 2009, 40, 211–236. [Google Scholar] [CrossRef]
- Brown, M.J.; Paxton, R.J. The conservation of bees: A global perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef]
- Phillips, B.B.; Shaw, R.F.; Holland, M.J.; Fry, E.L.; Bardgett, R.D.; Bullock, J.M.; Osborne, J.L. Drought reduces floral resources for pollinators. Global Change Biol. 2018, 24, 3226–3235. [Google Scholar] [CrossRef]
- Aizen, M.A.; Smith-Ramirez, C.; Morales, C.L.; Vieli, L.; Saez, A.; Barahona-Segovia, R.M.; Arbetman, M.P.; Montalva, J.; Garibaldi, L.A.; Inouye, D.W.; et al. Coordinated species importation policies are needed to reduce serious invasions globally: The case of alien bumblebees in South America. J. Appl. Ecol. 2019, 56, 100–106. [Google Scholar] [CrossRef]
- De Palma, A.; Abrahamczyk, S.; Aizen, M.A.; Albrecht, M.; Basset, Y.; Bates, A.; Blake, R.J.; Boutin, C.; Bugter, R.; Connop, S. Predicting bee community responses to land-use changes: Effects of geographic and taxonomic biases. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.; Altamirano, A.; Cayuela, L.; Lara, A.; González, M. Native forest loss in the Chilean biodiversity hotspot: Revealing the evidence. Reg. Environ. Change 2017, 17, 285–297. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Miñarro, M.; García, D.; Martínez, R. Impact of insect pollinators in agriculture: Importance and management of their biodiversity. Ecosistemas 2018, 27, 81–90. [Google Scholar] [CrossRef]
- Karp, D.S.; Mendenhall, C.D.; Sandi, R.F.; Chaumont, N.; Ehrlich, P.R.; Hadly, E.A.; Daily, G.C. Forest bolsters bird abundance, pest control and coffee yield. Ecol. Lett. 2013, 16, 1339–1347. [Google Scholar] [CrossRef]
- Rodriguez-San Pedro, A.; Chaperon, P.N.; Beitran, C.A.; Allendes, J.L.; Avila, F.I.; Grez, A.A. Influence of agricultural management on bat activity and species richness in vineyards of central Chile. J. Mammal. 2018, 99, 1495–1502. [Google Scholar] [CrossRef]
- García, D.; Miñarro, M.; Martínez-Sastre, R. Enhancing ecosystem services in apple orchards: Nest boxes increase pest control by insectivorous birds. J. Appl. Ecol. 2021, 58, 465–475. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Gemmill-Herren, B.; D’Annolfo, R.; Graeub, B.E.; Cunningham, S.A.; Breeze, T.D. Farming approaches for greater biodiversity, livelihoods, and food security. Trends Ecol. Evol. 2017, 32, 68–80. [Google Scholar] [CrossRef]
- Manosathiyadevan, M.; Bhuvaneshwari, V.; Latha, R. Impact of insects and pests in loss of crop production: A review. In Sustainable Agriculture towards Food Security; Dhanarajan, A., Ed.; Springer: Singapore, 2017; pp. 57–67. [Google Scholar]
- Casallanovo, F.; Simone, D.M.; Santos, G.S.; de Oliveira Kaminski, T.S.; Cione, A.P.; Peranginangin, N. Estimating pesticide environmental concentrations in Latin America: The importance of developing local scenarios. Integr. Environ. Assess. Manage. 2021, 17, 901–904. [Google Scholar] [CrossRef]
- Blacquiere, T.; Smagghe, G.; Van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment (vol 21, pg 973, 2012). Ecotoxicology 2012, 21, 1581. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Amado, J.; Manzi, C.; Franco, L.M.; Contecha, S.C.; Marquez, S.J.; Solano-Iguaran, J.J.; Haro, R.E.; Silva, A.X. Effects of residual doses of neonicotinoid (imidacloprid) on metabolic rate of queen honey bees Apis mellifera (Hymenoptera: Apidae). Apidologie 2020, 51, 1091–1099. [Google Scholar] [CrossRef]
- Bass, C.; Field, L.M. Neonicotinoids. Curr. Biol. 2018, 28, R772–R773. [Google Scholar] [CrossRef]
- Tasei, J.N.; Lerin, J.; Ripault, G. Sub-lethal effects of imidacloprid on bumblebees, Bombus terrestris (Hymenoptera: Apidae), during a laboratory feeding test. Pest. Manag. Sci. 2000, 56, 784–788. [Google Scholar] [CrossRef]
- Moncharmont, F.X.D.; DeCourtye, A.; Hennequet-Hantier, C.; Pons, O.; Pham-Delegue, M.H. Statistical analysis of honeybee survival after chronic exposure to insecticides. Environ. Toxicol. Chem. 2003, 22, 3088–3094. [Google Scholar] [CrossRef]
- Diaz, T.; del-Val, E.; Ayala, R.; Larsen, J. Alterations in honey bee gut microorganisms caused by Nosema spp. and pest control methods. Pest. Manag. Sci. 2019, 75, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Syromyatnikov, M.Y.; Isuwa, M.M.; Savinkova, O.V.; Derevshchikova, M.I.; Popov, V.N. The effect of pesticides on the microbiome of animals. Agriculture 2020, 10, 79. [Google Scholar] [CrossRef]
- Zhu, L.; Qi, S.; Xue, X.; Niu, X.; Wu, L. Nitenpyram disturbs gut microbiota and influences metabolic homeostasis and immunity in honey bee (Apis mellifera L.). Environ. Pollut. 2020, 258, 113671. [Google Scholar] [CrossRef]
- Henríquez-Piskulich, P.A.; Schapheer, C.; Vereecken, N.J.; Villagra, C. Agroecological strategies to safeguard insect pollinators in biodiversity hotspots: Chile as a case study. Sustainability 2021, 13, 6728. [Google Scholar] [CrossRef]
- Rodríguez, S.S.; Pérez-Giraldo, L.C.; Vergara, P.M.; Carvajal, M.A.; Alaniz, A.J. Native bees in Mediterranean semi-arid agroecosystems: Unravelling the effects of biophysical habitat, floral resource, and honeybees. Agr. Ecosyst. Environ. 2021, 307, 107188. [Google Scholar] [CrossRef]
- Smith-Ramirez, C.; Ramos-Jiliberto, R.; Valdovinos, F.S.; Martinez, P.; Castillo, J.A.; Armesto, J.J. Decadal trends in the pollinator assemblage of Eucryphia cordifolia in Chilean rainforests. Oecologia 2014, 176, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Jiliberto, R.; Albornoz, A.A.; Valdovinos, F.S.; Smith-Ramírez, C.; Arim, M.; Armesto, J.J.; Marquet, P.A. A network analysis of plant–pollinator interactions in temperate rain forests of Chiloé Island, Chile. Oecologia 2009, 160, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Medel, R.; González-Browne, C.; Salazar, D.A.; Ferrer, P.; Ehrenfeld, M. The most effective pollinator principle applies to new invasive pollinators. Biol. Lett. 2018, 30, art14. [Google Scholar] [CrossRef] [PubMed]
- Arbetman, M.P.; Meeus, I.; Morales, C.L.; Aizen, M.A.; Smagghe, G. Alien parasite hitchhikes to Patagonia on invasive bumblebee. Biol. Invasions 2013, 15, 489–494. [Google Scholar] [CrossRef]
- Arismendi, N.; Riveros, G.; Zapata, N.; Smagghe, G.; Gonzalez, C.; Vargas, M. Occurrence of bee viruses and pathogens associated with emerging infectious diseases in native and non-native bumble bees in southern Chile. Biol. Invasions 2021, 23, 1175–1189. [Google Scholar] [CrossRef]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyorgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef]
- Magrach, A.; González-Varo, J.P.; Boiffier, M.; Vilà, M.; Bartomeus, I. Honeybee spillover reshuffles pollinator diets and affects plant reproductive success. Nat. Ecol. Evol. 2017, 1, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.L.; Aizen, M.A. Invasive mutualisms and the structure of plant-pollinator interactions in the temperate forests of north-west Patagonia, Argentina. J. Ecol. 2006, 94, 171–180. [Google Scholar] [CrossRef]
- Ruz, L.; Herrera, R. Preliminary observations on foraging activities of Bombus dahlbomii and Bombus terrestris (Hym: Apidae) on native and non-native vegetation in Chile. In Proceedings of the VIII International Symposium on Pollination-Pollination: Integrator of Crops and Native Plant Systems, Mosonmagyaróvár, Hungary, 10–14 July 2000; Volume 561, pp. 165–169. [Google Scholar]
- Valdivia, C.E.; Carroza, J.P.; Orellana, J.I. Geographic distribution and trait-mediated causes of nectar robbing by the European bumblebee Bombus terrestris on the Patagonian shrub Fuchsia magellanica. Flora 2016, 225, 30–36. [Google Scholar] [CrossRef]
- Esterio, G.; Cares-Suarez, R.; Gonzalez-Browne, C.; Salinas, P.; Carvallo, G.; Medel, R. Assessing the impact of the invasive buff-tailed bumblebee (Bombus terrestris) on the pollination of the native Chilean herb Mimulus luteus. Arthropod-Plant. Interact. 2013, 7, 467–474. [Google Scholar] [CrossRef]
- Montalva, J.; Arroyo, M.T.K.; Ruz, L. Bombus terrestris Linnaeus (Hymenoptera: Apidae: Bombini) en Chile: Causas y consecuencias de su introducción. Rev. Jardín Botánico Chagual 2008, 6, 13–20. [Google Scholar]
- Carvallo, G.O.; Medel, R.; Navarro, L. Assessing the effects of native plants on the pollination of an exotic herb, the blueweed Echium vulgare (Boraginaceae). Arthropod-Plant. Interact. 2013, 7, 475–484. [Google Scholar] [CrossRef]
- Aldunce, P.; Araya, D.; Sapiain, R.; Ramos, I.; Lillo, G.; Urquiza, A.; Garreaud, R. Local perception of drought impacts in a changing climate: The mega-drought in central Chile. Sustainability 2017, 9, 2053. [Google Scholar] [CrossRef]
- Boisier, J.P.; Rondanelli, R.; Garreaud, R.D.; Muñoz, F. Anthropogenic and natural contributions to the Southeast Pacific precipitation decline and recent megadrought in central Chile. Geophys Res. Lett 2016, 43, 413–421. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010-2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sc. 2017, 21, 6307–6327. [Google Scholar] [CrossRef]
- Fontúrbel, F.E.; Lara, A.; Lobos, D.; Little, C. The cascade impacts of climate change could threaten key ecological interactions. Ecosphere 2018, 9, e02485. [Google Scholar] [CrossRef]
- Novoa, V.; Ahumada-Rudolph, R.; Rojas, O.; Saez, K.; de la Barrera, F.; Arumi, J.L. Understanding agricultural water footprint variability to improve water management in Chile. Sci. Total Environ. 2019, 670, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Barria, P.; Rojas, M.; Moraga, P.; Murnoz, A.; Bozkurt, D.; Alvarez-Garreton, C. Anthropocene and streamflow: Long-term perspective of streamflow variability and water rights. Elementa-Sci. Anthrop. 2019, 7, 1–15. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Robles, V.; Tamburrino, I.; Martinez-Harms, J.; Garreaud, R.D.; Jara-Arancio, P.; Pliscoff, P.; Copier, A.; Arenas, J.; Keymer, J.; et al. Extreme drought affects visitation and seed set in a plant species in the central Chilean Andes heavily dependent on hummingbird pollination. Plants 2020, 9, 1553. [Google Scholar] [CrossRef] [PubMed]
- Rais, A.; van de Kuilen, J.W.G.; Pretzsch, H. Growth reaction patterns of tree height, diameter, and volume of Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) under acute drought stress in Southern Germany. Eur. J. Forest Res. 2014, 133, 1043–1056. [Google Scholar] [CrossRef]
- Bell, D.M.; Pabst, R.J.; Shaw, D.C. Tree growth declines and mortality were associated with a parasitic plant during warm and dry climatic conditions in a temperate coniferous forest ecosystem. Glob. Chang. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; Gonzalez, M.E.; Camarero, J.J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 2020, 115, 106401. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Carrasco, B.; Orchard, C. Combining niche modelling, land-use change, and genetic information to assess the conservation status of Pouteria splendens populations in central Chile. Int. J. Ecol. 2015, 2015. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepulveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010-2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Blüthgen, N.; Klein, A.M. Functional complementarity and specialisation: The role of biodiversity in plant-pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]
- Pitts-Singer, T.L.; Artz, D.R.; Peterson, S.S.; Boyle, N.K.; Wardell, G.I. Examination of a managed pollinator strategy for almond production using Apis mellifera (Hymenoptera: Apidae) and Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2018, 47, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Aizen, M.A.; Vásquez, D.P.; Smith-Ramírez, C. Historial natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Rev. Chil. Hist. Nat. 2002, 75, 79–97. [Google Scholar] [CrossRef]
- Smith-Ramírez, C. Los picaflores y su recurso floral en el bosque templado de la isla de Chiloé, Chile [Hummingbirds and their floral resources in temperate forests of Chiloé island, Chile]. Rev. Chil. Hist. Nat. 1993, 66, 65–73. [Google Scholar]
- Hornung-Leoni, C.T.; Gonzalez-Gomez, P.L.; Troncoso, A.J. Morphology, nectar characteristics and avian pollinators in five Andean Puya species (Bromeliaceae). Acta Oecol. 2013, 51, 54–61. [Google Scholar] [CrossRef]
- Guerrero, P.C.; Antinao, C.A.; Vergara-Merino, B.; Villagra, C.A.; Carvallo, G.O. Bees may drive the reproduction of four sympatric cacti in a vanishing coastal mediterranean-type ecosystem. PeerJ 2019, 7, e7865. [Google Scholar] [CrossRef]
- Engel, M.S.; Ceríaco, L.M.; Daniel, G.M.; Dellapé, P.M.; Löbl, I.; Marinov, M.; Reis, R.E.; Young, M.T.; Dubois, A.; Agarwal, I. The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- Knight, T.M.; Ashman, T.L.; Bennett, J.M.; Burns, J.H.; Passonneau, S.; Steets, J.A. Reflections on, and visions for, the changing field of pollination ecology. Ecol. Lett. 2018, 21, 1282–1295. [Google Scholar] [CrossRef] [PubMed]
- Cuadra-Valdés, J.; Vizentin-Bugoni, J.; Fontúrbel, F.E. An exotic magnet plant alters pollinator abundance and behavior: A field test with a native mistletoe. Biol. Invasions 2021. [Google Scholar] [CrossRef]
- Johnson, S.D.; Peter, C.I.; Nilsson, L.A.; Agren, J. Pollination success in a deceptive orchid is enhanced by co-occurring rewarding magnet plants. Ecology 2003, 84, 2919–2927. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological intensification to mitigate impacts of conventional intensive land use on pollinators and pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Polit-Granja, E. Situación de las Viñas y los Vinos Orgánicos Chilenos; Oficina de Estudios y Políticas Agrarias (ODEPA): Santiago de Chile, Chile, 2005.
- Barbosa, O.; Villagra, P. Socio-ecological studies in urban and rural ecosystems in Chile. In Earth Stewardship: Linking Ecology and Ethics in Theory and Practice; Rozzi, R., Chapin, F.S., Callicott, J.B., Pickett, S.T.A., Power, M.E., Armesto, J.J., May, R.H., Eds.; Springer: Cham, Switzerland, 2015; pp. 297–311. [Google Scholar]
- Pandya, R.E. A framework for engaging diverse communities in citizen science in the US. Front. Ecol. Environ. 2012, 10, 314–317. [Google Scholar] [CrossRef]
- Giovanetti, M.; Albertazzi, S.; Flaminio, S.; Ranalli, R.; Bortolotti, L.; Quaranta, M. Pollination in agroecosystems: A review of the conceptual framework with a view to sound monitoring. Land 2021, 10, 540. [Google Scholar] [CrossRef]
- Decourtye, A.; Armengaud, C.; Renou, M.; Devillers, J.; Cluzeau, S.; Gauthier, M.; Pham-Delegue, M.H. Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pestic Biochem. Phys. 2004, 78, 83–92. [Google Scholar] [CrossRef]
- Guez, D.; Suchail, S.; Gauthier, M.; Maleszka, R.; Belzunces, L.P. Contrasting effects of imidacloprid on habituation in 7- and 8-day-old honeybees (Apis mellifera). Neurobiol. Learn. Mem. 2001, 76, 183–191. [Google Scholar] [CrossRef]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? P. Roy. Soc. B-Biol. Sci. 2020, 287, 20201265. [Google Scholar] [CrossRef] [PubMed]
- SAG. Listado de Plaguicidas de Uso Agrícola Prohibidos en Chile. Última actualización 24 de febrero de 2020. Código: F-RIS-RAI-PA-013; Servicio Agrícola y Ganadero: Santiago de Chile, Chile, 2014.
- Cordero, S.; Castano-Villa, G.J.; Fontúrbel, F.E. The best bang for the bucks: Rethinking global investment in biodiversity conservation. Sustainability 2020, 12, 9252. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).