Effects of Temperature Rise on Multi-Taxa Distributions in Mountain Ecosystems


Abstract
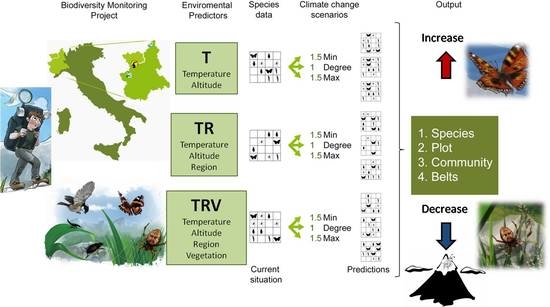
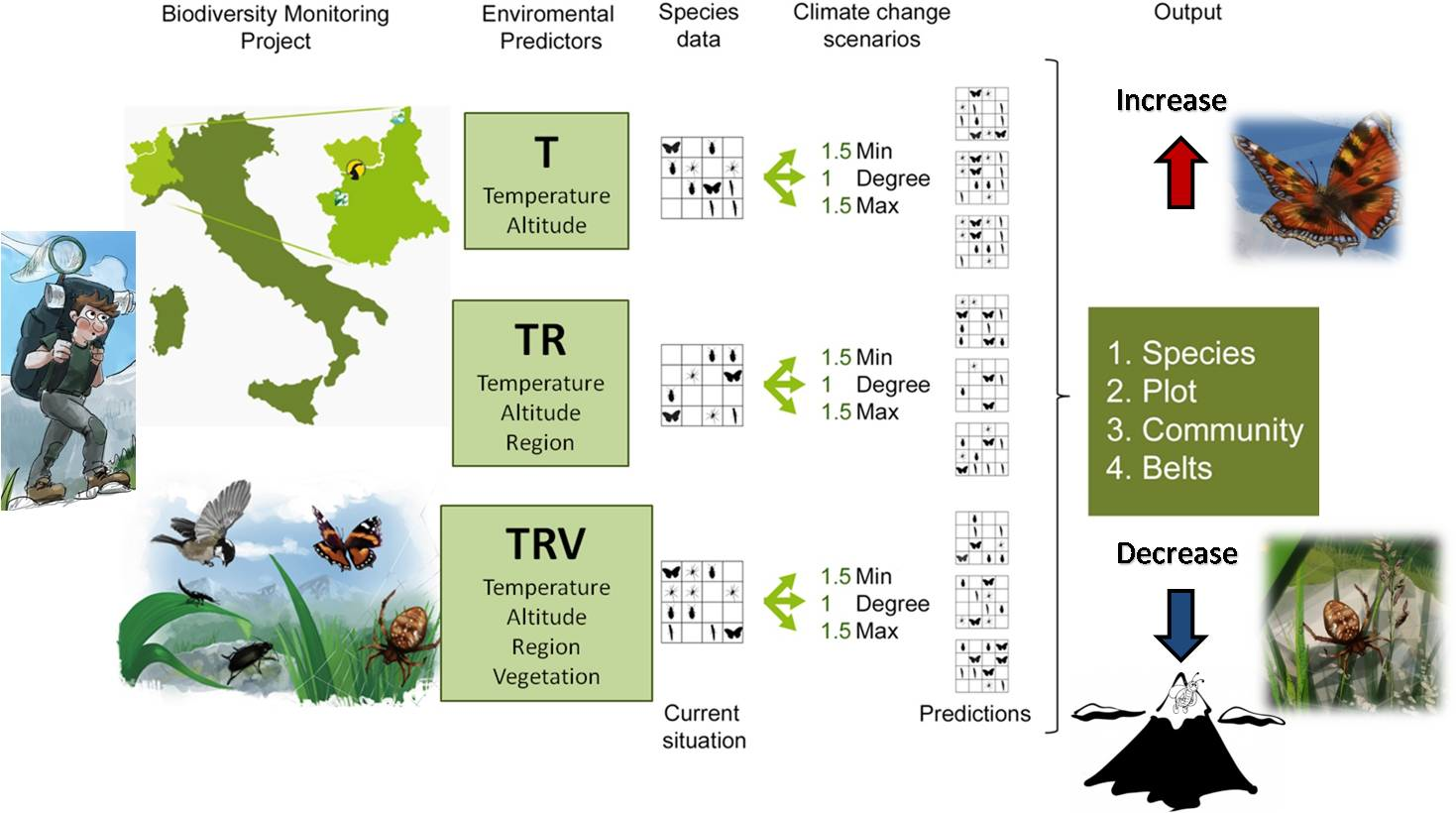
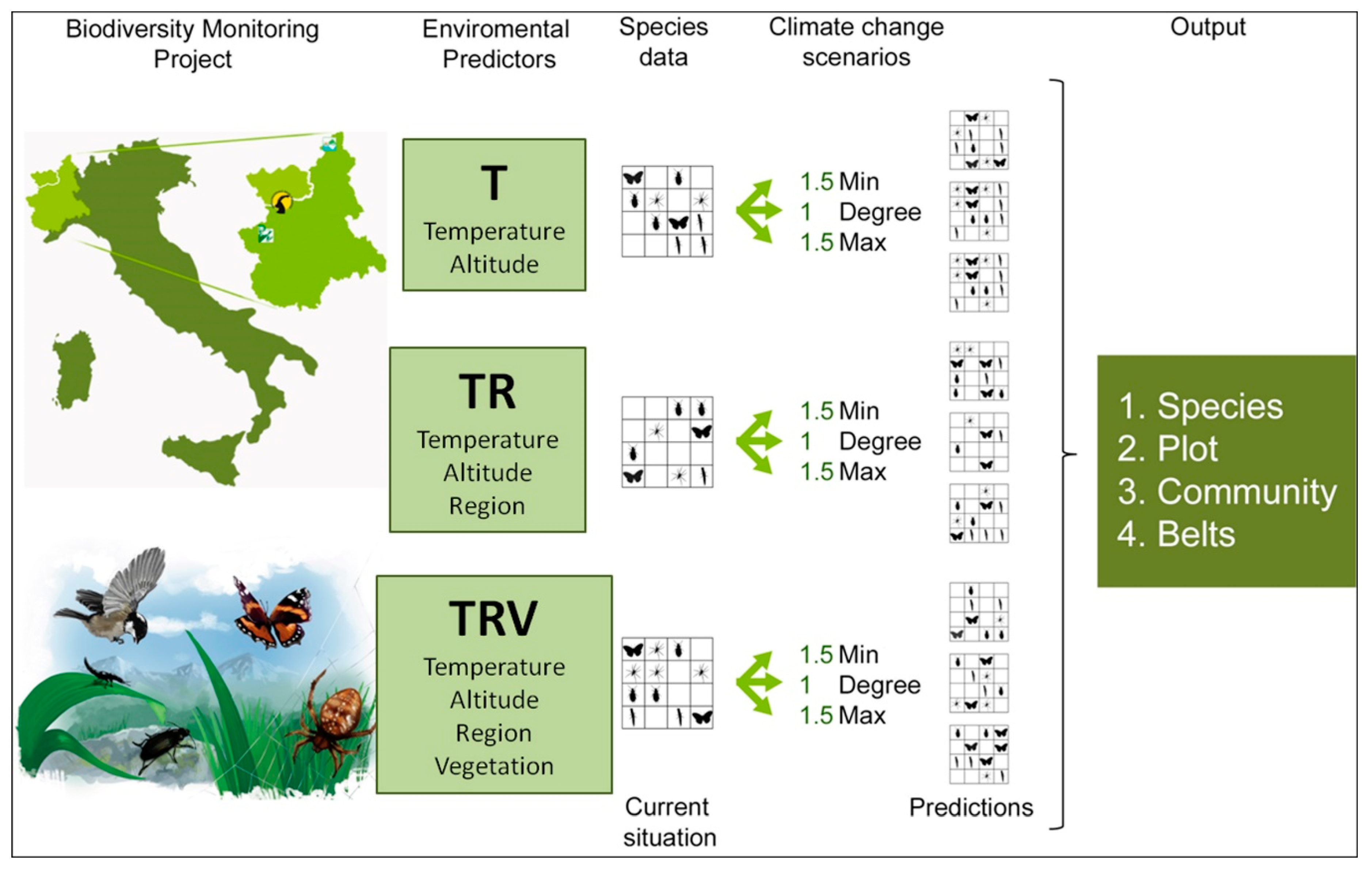
1. Introduction
2. Materials and Methods
2.1. Biodiversity Inventory in the Northwestern Italian Alps: Data Sources
2.2. Model Simulation: Current Conditions and Temperature Change Scenarios
- Temperature (T), which considers only temperature-derived variables (seasonal mean, maximum, minimum temperature, and standard deviation, in °C) and altitude to model species distribution;
- Temperature+Region (TR), which considers temperature-derived variables, altitude, and geographical location;
- Temperature+Region+Vegetation Structure (TRV), which considers temperature-derived variables, altitude, geographical location, and vegetation cover.
- 1Degree (d), in which minimum, mean, and maximum temperature are all equally increased by 1 °C;
- 1.5Min (min), in which minimum temperature is increased by 1.5 °C, mean temperature by 1 °C, maximum temperature by 0.5 °C;
- 1.5Max (max), in which minimum temperature is increased by 0.5 °C, mean temperature by 1 °C, maximum temperature by 1.5 °C.
2.3. Analysis of Model Outputs
2.3.1. Species Distribution
2.3.2. Species Richness
2.3.3. Community Composition
3. Results
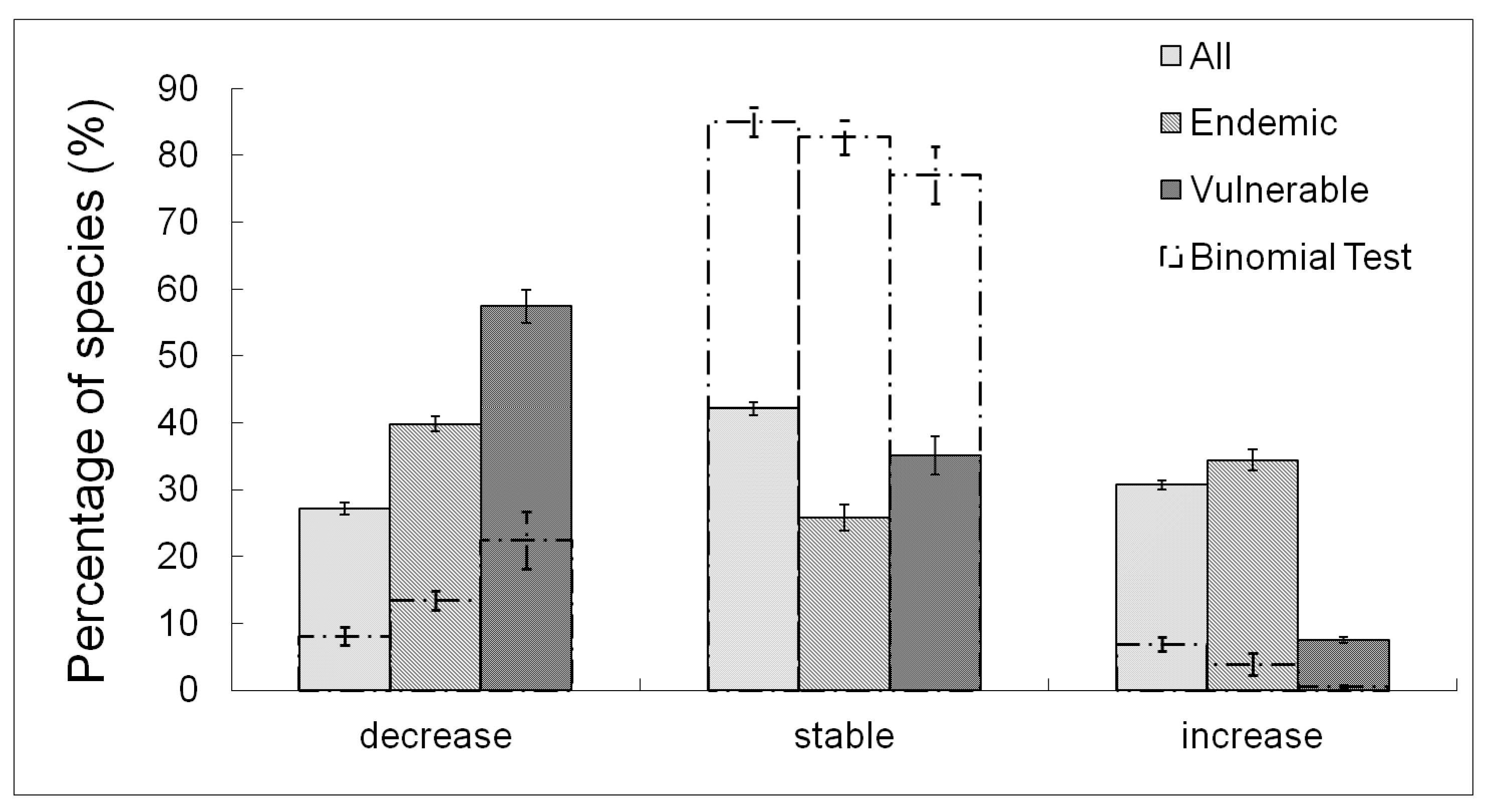
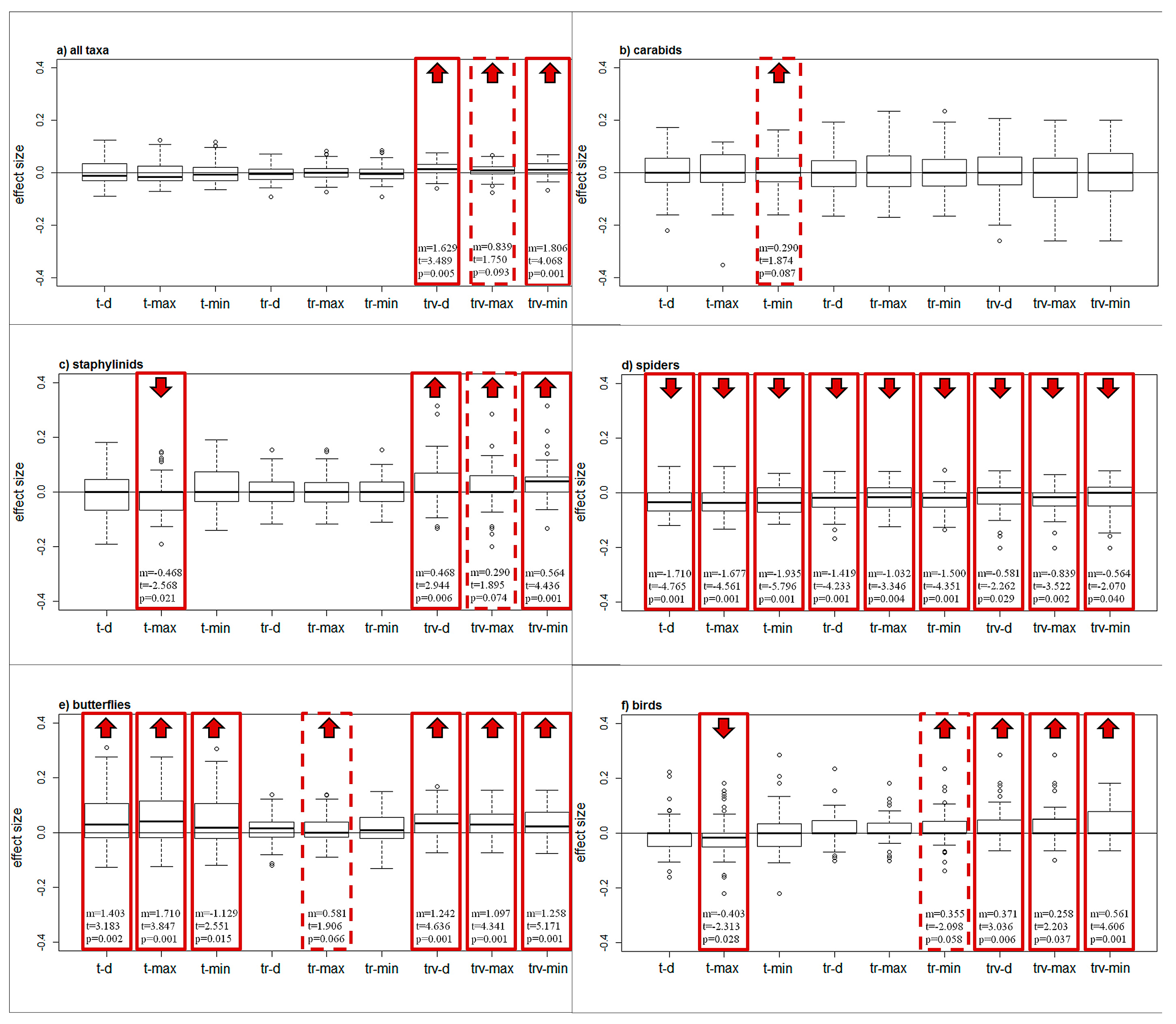
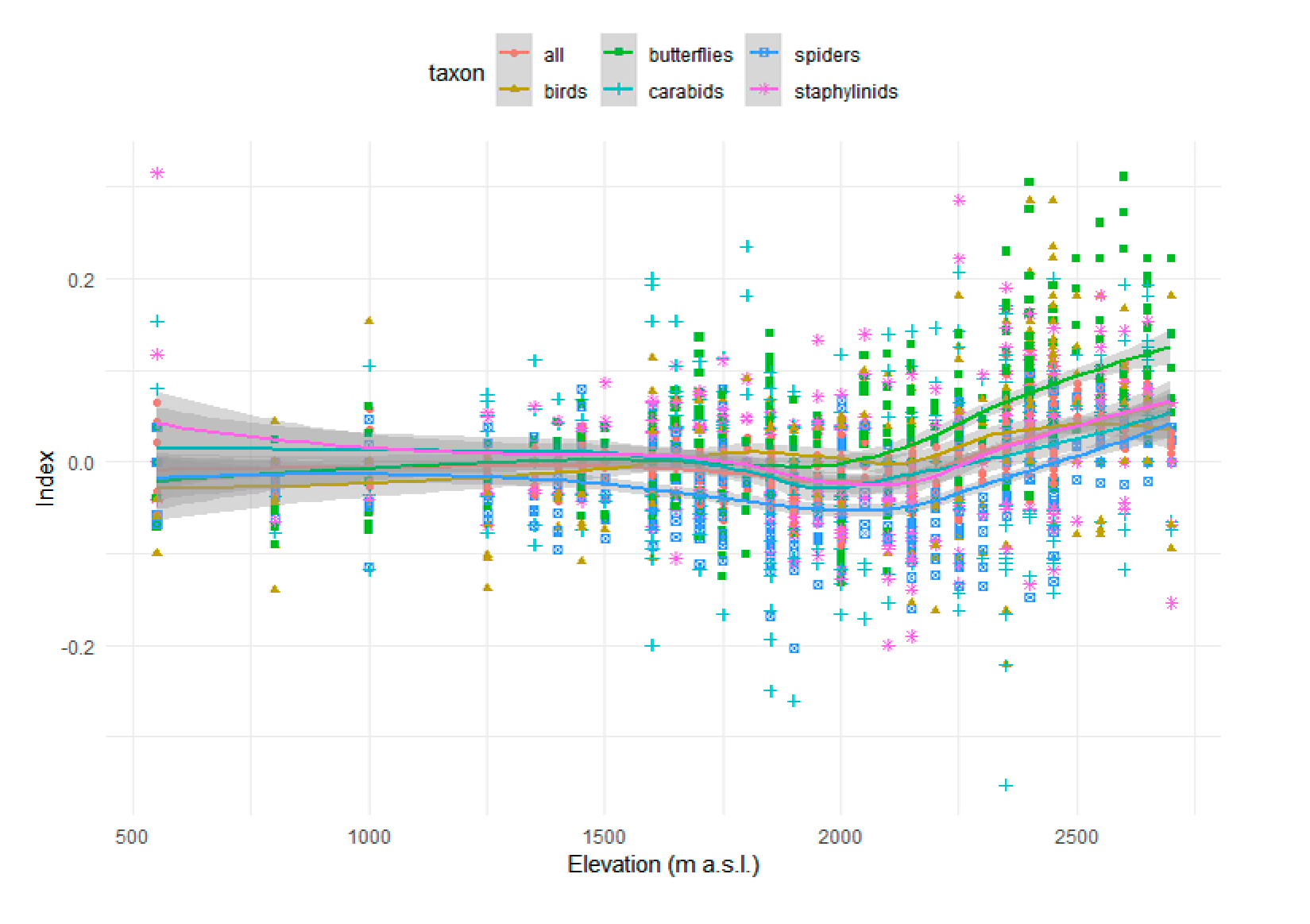
3.1. Species Distribution
3.2. Species Richness
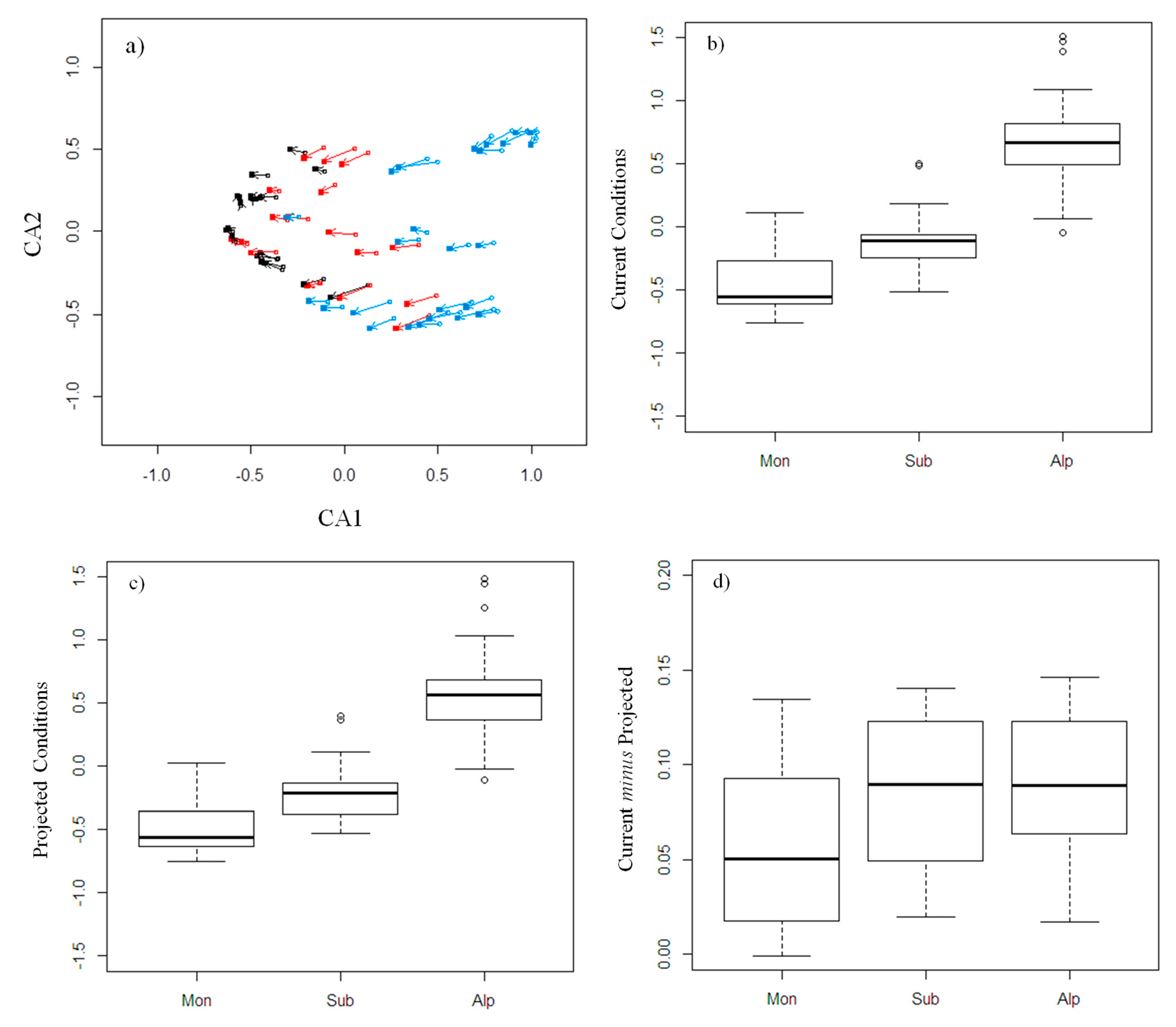
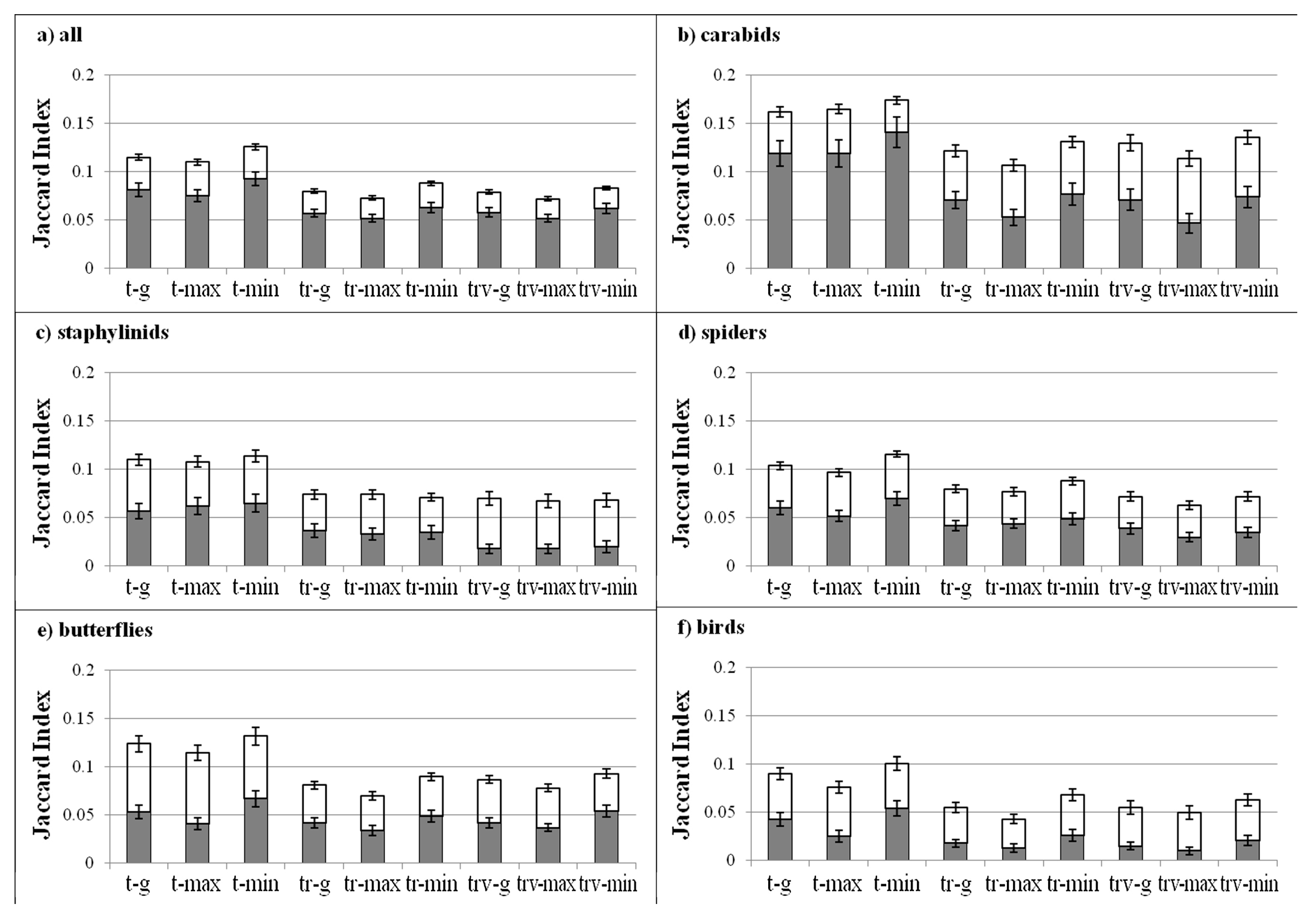
3.3. Community Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Settele, J.; Scholes, R.; Betts, R.A.; Bunn, S.; Leadley, P.; Nepstad, D.; Overpeck, J.T.; Taboada, M.A.; Fischlin, A.; Moreno, J.M.; et al. Terrestrial and inland water systems. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, J.K., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 271–359. [Google Scholar]
- Kujala, H.; Burgman, M.A.; Moilanen, A. Treatment of uncertainty in conservation under climate change. Conserv. Lett. 2013, 6, 73–85. [Google Scholar] [CrossRef]
- Reside, A.E.; Butt, N.; Adams, V.M. Adapting systematic conservation planning for climate change. Biodivers. Conserv. 2018, 27, 1–29. [Google Scholar] [CrossRef]
- Lloret, F. Trade-offs in high mountain conservation. In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J., Aniz, M., Eds.; Advances in Global Change Research, 62; Springer: Berlin, Germany, 2017; pp. 37–59. [Google Scholar]
- Dirnböck, T.; Essel, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Huber, U.; Reasoner, M.; Bugmann, H. Global Change and Mountain Regions: An Overview of Current Knowledge; Advances Global Change Research; Springer: Berlin, Germany, 2005. [Google Scholar]
- Diaz, H.F.; Grosejeanan, M.; Graumlich, L. Climate variability and change in high elevation regions: Past, present and future. Clim. Chang. 2003, 59, 1–4. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.; Erread, M.P.; Martínez-Rica, J.P. Exposure of global mountain systems to climate warming during the 21st Century. Glob. Environ. Chang. 2007, 17, 420–428. [Google Scholar] [CrossRef]
- Grabherr, G.; Gottfried, M.; Pauli, H. Global change effects on alpine plant diversity. In Biodiversity Hotspots. Distribution and Protection of Conservation Priority Areas; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin, Germany, 2011; pp. 529–536. [Google Scholar]
- Schmitt, T. Biogeographical and evolutionary importance of the European high mountain systems. Front. Zool. 2009, 6, 1–9. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Reiter, K.; Klettner, C.; Grabherr, G. Signals of range expansions and contractions of vascular plants in the high Alps: Observations (1994–2004) at the GLORIA master site Schrankogel, Tyrol, Austria. Glob. Chang. Biol. 2007, 13, 147–156. [Google Scholar] [CrossRef]
- Beniston, M. Mountain weather and climate: A general overview and a focus on climatic change in the Alps. Hydrobiologia 2006, 562, 3–16. [Google Scholar] [CrossRef]
- Calmanti, S.; Motta, L.; Turco, M.; Provenzale, A. Impact of climate variability on Alpine glaciers in northwestern Italy. Int. J. Climatol. 2007, 27, 2041–2053. [Google Scholar] [CrossRef]
- Keiler, M.; Knight, J.; Harrison, S. Climate change and geomorphological hazards in the eastern European Alps. Philos. Trans. R. Soc. A 2010, 368, 2461–2479. [Google Scholar] [CrossRef] [PubMed]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraer, M.; Cáceres, B.; Forsythe, N.; Fowler, H.J.; Greenwood, G.; Ziaur Rahman Hashmi, M.; Liu, X.D.; et al. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Special Report on the Ocean and Cryosphere in a Changing Climate. Available online: https://www.ipcc.ch/srocc/ (accessed on 25 March 2020).
- Poussin, C.; Guigoz, Y.; Palazzi, E.; Terzago, S.; Chatenoux, B.; Giuliani, G. Snow cover evolution in the Gran Paradiso National Park, Italian Alps, using the Earth Observation data cube. Data 2019, 4, 138. [Google Scholar] [CrossRef]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Payne, D.; Spehn, E.M.; Snethlage, M.; Fischer, M. Opportunities for research on mountain biodiversity under global change. Curr. Opin. Environ. Sustain. 2017, 29, 40–47. [Google Scholar] [CrossRef]
- Theurillat, J.P.; Guisan, A. Potential impact of climate change on vegetation in the European Alps: A review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Lesica, P.; McCune, B. Decline of arctic-alpine plants at the southern margin of their range following a decade of climatic warming. J. Veg. Sci. 2004, 15, 679–690. [Google Scholar] [CrossRef]
- Winkler, D.; Lubetkin, K.C.; Carrell, A.A.; Jabis, M.D.; Yang, Y.; Kueppers, L.M. Responses of alpine plant communities to climate warming. In Ecosystem Consequences of Soil Warming; Mohan, J.E., Ed.; Academic Press: London, UK, 2019; pp. 297–346. [Google Scholar]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Wilson, R.J.; Gutiérrez, D.; Gutiérrez, D.M.; Agudo, R.; Monserrat, V.J. Changes to the elevation limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef]
- Gobbi, M.; Fontaneto, D.; de Bernardi, F. Influence of climate changes on animal communities in space and time: The case of spider assemblages along an alpine glacier foreland. Glob. Chang. Biol. 2006, 12, 1985–1992. [Google Scholar] [CrossRef]
- Buntgen, U.; Greuter, L.; Bollman, K.; Jenny, H.; Liebhold, A.; Galvan, J.D.; Stenseth, N.C.; Andrew, C.; Mysterud, A. Elevational range shifts in four mountain ungulate species from the Swiss Alps. Ecosphere 2017, 8, e01761. [Google Scholar] [CrossRef]
- Menéndez, R.; Megías, A.G.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species richness changes lag behind climate change. Proc. R. Soc. B 2006, 273, 1465–1470. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.V.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef]
- Nooten, S.S.; Andrew, N.R. Transplant experiments—A powerful method to study climate change impacts. In Invertebrates and Global Climate Change; Johnson, S., Jones, H., Eds.; Wiley-Blackwell: Oxford, UK, 2017; pp. 46–67. [Google Scholar]
- Palmer, G.; Hill, J.K. Using historical data for studying range changes. In Invertebrates and Global Climate Change; Johnson, S., Jones, H., Eds.; Wiley-Blackwell: Oxford, UK, 2017; pp. 9–29. [Google Scholar]
- Ferrier, S.; Guisan, A. Spatial modelling of biodiversity at the community level. J. Appl. Ecol. 2006, 43, 393–404. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, L.R. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2012, 14, 484–492. [Google Scholar] [CrossRef]
- Dunlop, M. Biodiversity: Strategy conservation. Nat. Clim. Chang. 2013, 3, 1019–1020. [Google Scholar] [CrossRef]
- Heller, N.E.; Zavaleta, E.S. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Catalan, J.; Ninot, J.; Aniz, M. The High Mountain Conservation in a Changing World; Catalan, J., Ninot, J., Aniz, M., Eds.; Advances in Global Change Research, 62; Springer: Berlin, Germany, 2017; pp. 3–36. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models—With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef]
- Lavergne, S.; Mouquet, N.; Thuiller, W.; Ronce, O. Biodiversity and climate change: Integrating evolutionary and ecological responses of species and communities. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 321–350. [Google Scholar] [CrossRef]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef]
- Cotto, O.; Wessely, J.; Georges, D.; Klonner, G.; Schmid, M.; Dullinger, S.; Thuiller, W.; Guillaume, F. A dynamic eco-evolutionary model predicts slow response of alpine plants to climate warming. Nat. Commun. 2017, 8, 15399. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond predictions: Biodiversity conservation in a changing climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Arribas, P.; Abellán, P.; Velasco, J.; Millán, A.; Sánchez-Fernández, D. Conservation of insects in the face of global climate change. In Invertebrates and Global Climate Change; Johnson, S., Jones, H., Eds.; Wiley-Blackwell: Oxford, UK, 2017; pp. 349–367. [Google Scholar]
- Scherrer, D.; Massy, S.; Meier, S.; Vittoz, P.; Guisan, A. Assessing and predicting shifts in mountain forest composition across 25years of climate change. Divers. Distrib. 2017, 23, 517–528. [Google Scholar] [CrossRef]
- Khalyani, A.H.; Gould, W.A.; Falkowski, M.J.; Muscarella, R.; Uriarte, M.; Yousef, F. Climate change increases potential plant species richness on Puerto Rican uplands. Clim. Chang. 2019, 156, 15–30. [Google Scholar] [CrossRef]
- Viterbi, R.; Cerrato, C.; Bassano, B.; Bionda, R.; von Hardenberg, A.; Provenzale, A.; Bogliani, G. Patterns of biodiversity in the northwestern Italian Alps: A multi-taxa approach. Com. Ecol. 2013, 14, 18–30. [Google Scholar] [CrossRef]
- Cerrato, C.; Rocchia, E.; Brunetti, M.; Bionda, R.; Bassano, B.; Provenzale, A.; Bonelli, S.; Viterbi, R. Butterfly distribution along altitudinal gradients: Temporal changes over a short time period. Nat. Conserv. 2019, 34, 91–118. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; le Lay, G.; Piedallu, C.; Albert, C.H.; et al. 21st century climate change threatens mountain flora unequally across Europe. Glob. Chang. Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Thuiller, W.; Pollock, L.J.; Gueguen, M.; Münkemüller, T. From species distributions to meta-communities. Ecol. Lett. 2015, 18, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Mouquet, N.; Lagadeuc, Y.; Devictor, V.; Doyen, L.; Duputié, A.; Eveillard, D.; Faure, D.; Garnier, E.; Gimenez, O.; Huneman, P.; et al. Review: Predictive ecology in a changing world. J. Appl. Ecol. 2015, 52, 1293–1310. [Google Scholar] [CrossRef]
- Perrigo, A.; Hoorn, C.; Antonelli, A. Why mountains matter for biodiversity. J Biogeogr. 2020, 47, 315–325. [Google Scholar] [CrossRef]
- Thuiller, W.; Münkemüller, T.; Lavergne, S.; Mouillot, D.; Mouquet, N.; Schiffers, K.; Gravel, D. A road map for integrating eco-evolutionary processes into biodiversity models. Ecol. Lett. 2013, 16, 94–105. [Google Scholar] [CrossRef]
- Boulangeat, I.; Georges, D.; Dentant, C.; Bonet, R.; Van Es, J.; Abdulhak, S.; Zimmermann, N.E.; Thuiller, W. Anticipating the spatio-temporal response of plant diversity and vegetation structure to climate and land use change in a protected area. Ecography 2014, 37, 1230–1239. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- McGeoch, M.A. The selection, testing and application of terrestrial insects as biondicators. Biol. Rev. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Kati, V.; Devillers, P.; Dufrêne, M.; Legakis, A.; Vokou, D.; Lebrun, P. Testing the value of six taxonomic groups as biodiversity indicators at a local scale. Conserv. Biol. 2004, 18, 667–675. [Google Scholar] [CrossRef]
- Sauberer, N.; Zulka, P.K.; Abensperg-Traun, M.; Berg, H.; Bieringer, G.; Milasowszky, N.; Moser, D.; Plutzar, C.; Pollheimer, M.; Storch, C.; et al. Surrogate taxa for biodiversity in agricultural landscapes of eastern Austria. Biol. Conserv. 2004, 117, 181–190. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Jackson, J.K. Terrestrial and aquatic invertebrates as bioindicators for environmental monitoring, with particular reference to mountain ecosystems. Environ. Manag. 2005, 35, 649–666. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.; Paulsen, J.; Spehn, E. A definition of mountains and their bioclimatic belts for gr global comparisons of biodiversity data. Alp. Bot. 2011, 121, 73–78. [Google Scholar] [CrossRef]
- Beniston, M. Environmental Change in Mountains and Uplands; Routledge: Abingdon-on-Thames, UK, 2000. [Google Scholar]
- Marini, L.; Bommarco, R.; Fontana, P.; Battisti, A. Disentangling effects of habitat diversity and area on orthopteran species with contrasting mobility. Biol. Conserv. 2010, 143, 2164–2171. [Google Scholar] [CrossRef]
- Oliver, T.; Roy, D.B.; Hill, J.K.; Brereton, T.; Thomas, C.D. Heterogeneous landscapes promote population stability. Ecol.Lett. 2010, 13, 473–484. [Google Scholar] [CrossRef]
- Mateo, R.G.; Mokany, K.; Guisan, A. Biodiversity models: What if unsaturation is the rule? Trends Ecol. Evol. 2012, 32, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Philips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Warren, R.; VanDerWal, J.; Price, J.; Welbergen, J.A.; Atkinson, I.; Ramirez-Villegas, J.; Osborn, T.J.; Jarvis, A.; Shoo, L.P.; Williams, S.E.; et al. Quantifying the benefit of early climate change mitigation in avoiding biodiversity loss. Nat. Clim. Chang. 2013, 3, 678–682. [Google Scholar] [CrossRef]
- Bradie, J.; Leung, B. A quantitative synthesis of the importance of variables used in MaxEnt species distribution models. J. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef]
- Williams, J.N.; Seo, C.; Thorne, J.; Nelson, J.K.; Erwin, S.; O’Brien, J.M.; Schwartz, M.W. Using species distribution models to predict new occurrences for rare plants. Divers. Distrib. 2009, 15, 565–576. [Google Scholar] [CrossRef]
- Rebelo, H.; Jones, G. Ground validation of presence-only modelling with rare species: A case study on barbastelles Barbastella barbastellus (Chiroptera: Vespertilionidae). J. App. Ecol. 2010, 47, 410–420. [Google Scholar] [CrossRef]
- Rinnhofer, L.J.; Roura-Pascual, N.; Arthofer, W.; Dejaco, T.; Thaler-Knoflach, B.; Wachter, G.A.; Christian, E.; Steiner, F.M.; Schlick-Steiner, B.C. Iterative species distribution modelling and ground validation in endemism research: An Alpine jumping bristletail example. Biodivers. Conserv. 2012, 21, 2845–2863. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Dobrowski, S.Z. A climatic basis for microrefugia: The influence of terrain on climate. Glob. Chang. Biol. 2011, 17, 1022–1035. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Nijs, I.; Lenoir, J. Incorporating microclimate into species distribution models. Ecography 2019, 42, 1267–1279. [Google Scholar] [CrossRef]
- Kulonen, A.; Imboden, R.A.; Rixen, C.; Maier, S.B.; Wipf, S. Enough space in a warmer world? Microhabitat diversity and small-scale distribution of alpine plants on mountain summits. Divers. Distrib. 2018, 24, 252–261. [Google Scholar] [CrossRef]
- Ciccarelli, N.; von Hardenberg, J.; Provenzale, A.; Ronchi, C.; Vargiu, A.; Pelosini, R. Climate variability in north-western Italy during the second half of the 20th century. Glob. Planet Chang. 2008, 63, 185–195. [Google Scholar] [CrossRef]
- Rottler, E.; Kormann, C.; Francke, T.; Bronstert, A. Elevation-dependent warming in the Swiss Alps 1981–2017: Features, forcings and feedbacks. Int. J. Climatol. 2019, 39, 2556–2568. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Global Warming of 1.5 °C; Special Report; Published online by UNEP and WMO; 2018; Available online: https://www.ipcc.ch/sr15/ (accessed on 25 March 2020).
- Turco, M.; Rosa-Cánovas, J.J.; Bedia, J.; Jerez, S.; Montávez, J.P.; Llasat, M.C.; Provenzale, A. Exacerbated fires in Mediterranean Europe due to anthropogenic warming projected with nonstationary climate-fire models. Nat. Commun. 2018, 9, 3821. [Google Scholar] [CrossRef] [PubMed]
- Terzago, S.; von Hardenberg, J.; Palazzi, E.; Provenzale, A. Snow water equivalent in the Alps as seen by gridded data sets, CMIP5 and CORDEX climate models. Cryosphere 2017, 11, 1625–1645. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Olden, J.D.; Joy, M.K.; Death, R.G. Rediscovering the species in community-wide predictive modeling. Ecol. Appl. 2006, 16, 1449–1460. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Van Turnhout, C.; Foppen, R.P.B.; Leuven, R.S.E.W.; Siepel, H.; Esselink, H. Scale-dependent homogenization: Changes in breeding bird diversity in the Netherlands over a 25-year period. Biol. Conserv. 2007, 134, 505–516. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.43.15. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 June 2019).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A. Nonparametric Statistical Methods; John Wiley & Sons: New York, NY, USA, 1973; pp. 139–146. [Google Scholar]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 10 January 2018).
- Sekercioglu, C.H.; Schneider, S.H.; Fay, J.P.; Loarie, S.R. Climate change, elevational range shifts, and bird extinctions. Conserv. Biol. 2008, 22, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Kreyling, J.; Thiel, D.; Simmnacher, K.; Willner, E.; Jentsch, A.; Beierkuhnlein, C. Geographic origin and past climatic experience influence the response to late spring frost in four common grass species in central Europe. Ecography 2012, 35, 268–275. [Google Scholar] [CrossRef]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Polato, N.R.; Gill, B.A.; Shah, A.A.; Gray, M.M.; Casner, K.L.; Barthelet, A.; Messer, P.W.; Simmons, M.P.; Guayasamin, J.M.; Encalada, A.C.; et al. Narrow thermal tolerance and low dispersal drive higher speciation in tropical mountains. Proc. Natl. Acad. Sci. USA 2018, 115, 12471–12476. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Devictor, V.; van Swaay, C.; Brereton, T.; Brotons, L.; Chamberlain, D.; Heliola, J.; Herrando, S.; Julliard, R.; Kuussaari, M.; Lindstrom, A.; et al. Differences in the climatic debts of birds and butterflies at a continental scale. Nat. Clim. Chang. 2012, 2, 121–124. [Google Scholar] [CrossRef]
- Konvička, M.; Beneš, J.; Čižek, O.; Kuras, T.; Klečkova, I. Has the currently warming climate affected populations of the mountain ringlet butterfly, Erebia epiphron (Lepidoptera: Nymphalidae), in low-elevation mountains? Eur. J. Entomol. 2016, 113, 295–301. [Google Scholar] [CrossRef]
- Roth, T.; Plattner, M.; Amrhein, V. Plants, birds and butterflies: Short-term responses of species communities to climate warming vary by taxon and with altitude. PLoS ONE 2014, 9, e82490. [Google Scholar] [CrossRef]
- Bell, J.R.; Wheater, C.P.; Cullen, W.R. The implications of grassland and heathland management for the conservation of spider communities: A review. J. Zool. 2001, 255, 377–387. [Google Scholar] [CrossRef]
- DeVito, J.; Meik, J.M.; Gerson, M.M.; Formanowicz, D.R. Physiological tolerances of three sympatric riparian wolf spiders (Araneae: Lycosidae) correspond with microhabitat distributions. Can. J. Zool. 2004, 82, 1119–1125. [Google Scholar] [CrossRef]
- Davey, C.M.; Chamberlain, D.E.; Newson, S.E.; Noble, D.G.; Johnston, A. Rise of the generalists: Evidence for climate driven homogenization in avian communities. Glob. Ecol. Biogeogr. 2012, 21, 568–578. [Google Scholar] [CrossRef]
- Savage, J.; Vellend, M. Elevational shifts, biotic homogenization and time lags in vegetation change during 40 years of climate warming. Ecography 2014, 37, 1–10. [Google Scholar] [CrossRef]
- Magurran, A.E.; Dornelas, M.; Moyes, F.; Gotelli, N.J.; McGill, B. Rapid biotic homogenization of marine fish assemblages. Nat. Commun. 2015, 6, 8405. [Google Scholar] [CrossRef]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Radeloff, V.C.; Williams, J.W.; Bateman, B.L.; Burke, K.D.; Carter, S.K.; Childress, E.S.; Cromwell, K.J.; Gratton, C.; Hasley, A.O.; Kraemer, B.M.; et al. The rise of novelty in ecosystems. Ecol. App. 2015, 25, 2051–2068. [Google Scholar] [CrossRef] [PubMed]
- Steinbauer, M.J.; Grytnes, J.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef]
- Vose, R.S.; Easterling, D.R.; Gleason, B. Maximum and minimum temperature trends for the globe. An update through 2004. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-term datasets in biodiversity research and monitoring: Assessing change in ecological communities through time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response of Different Taxa | Estimate | t-Value | p-Value |
|---|---|---|---|
| Intercept | 0.004 (0.005) | 0.926 | 0.354 |
| Birds | 0.008 (0.003) | 2.292 | 0.022 |
| Butterflies | 0.027 (0.003) | 7.778 | 0.000 |
| Carabids | −0.001 (0.003) | −0.201 | 0.841 |
| Spiders | −0.028 (0.003) | −7.965 | 0.000 |
| Staphylinids | 0.005 (0.003) | 1.420 | 0.156 |
| Differences Across Model Classes | All Taxa | Carabids | Staphylinids | Spiders | Butterflies | Birds |
|---|---|---|---|---|---|---|
| Intercept | 0.003 (0.004) | 0.012 (0.008) | 0.000 (0.007) | −0.030 (0.005) | 0.047 (0.007) | −0.009 (0.007) |
| t-value | 0.673 | 1.5 | 0.082 | −5.750 | 6.299 | −1.291 |
| p-value | 0.501 | 0.134 | 0.935 | <0.0001 | <0.0001 | 0.197 |
| TR | −0.004 (0.002) | −0.012 (0.007) | −0.000 (0.006) | 0.006 (0.003) | −0.033 (0.005) | 0.024 (0.005) |
| t-value | −1.768 | −1.629 | −0.023 | 1.729 | −6.865 | 4.723 |
| p-value | 0.078 | 0.104 | 0.981 | 0.084 | <0.0001 | <0.0001 |
| TRV | 0.009 (0.002) | −0.013 (0.007) | 0.026 (0.006) | 0.012 (0.003) | −0.014 (0.005) | 0.039 (0.005) |
| t-value | 3.622 | −1.736 | 4.429 | 3.213 | −3.009 | 7.573 |
| p-value | 0.0003 | 0.083 | <0.0001 | 0.001 | 0.003 | <0.0001 |
| F-value | 15.099 | 1.893 | 13.15 | 5.173 | 23.684 | 29.259 |
| p-value | <0.0001 | 0.152 | <0.0001 | 0.006 | <0.0001 | <0.0001 |
| Differences Across Vegetation Belts | All taxa | Carabids | Staphylinids | Spiders | Butterflies | Birds |
|---|---|---|---|---|---|---|
| Intercept | −0.007 (0.006) | 0.008 (0.012) | −0.001 (0.010) | −0.031 (0.008) | 0.007 (0.010) | −0.007 (0.011) |
| t-value | −1.076 | 0.682 | −0.139 | −3.862 | 0.661 | −0.701 |
| p-value | 0.283 | 0.496 | 0.890 | 0.0001 | 0.509 | 0.484 |
| Subalpine | −0.003 (0.009) | −0.021 (0.017) | 0.008 (0.015) | −0.007 (0.011) | −0.002 (0.015) | 0.018 (0.015) |
| t-value | −0.292 | −1.212 | 0.555 | −0.649 | −0.128 | 1.172 |
| p-value | 0.771 | 0.230 | 0.581 | 0.519 | 0.899 | 0.246 |
| Alpine | 0.029 (0.008) | 0.004 (0.016) | 0.020 (0.014) | 0.023 (0.011) | 0.062 (0.014) | 0.036 (0.014) |
| t-value | 3.503 | 0.228 | 1.505 | 2.224 | 4.428 | 2.564 |
| p-value | 0.001 | 0.821 | 0.138 | 0.030 | <0.0001 | 0.013 |
| F-value | 9.285 | 1.252 | 1.170 | 4.759 | 13.972 | 3.308 |
| p-value | 0.0003 | 0.293 | 0.317 | 0.012 | <0.0001 | 0.043 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viterbi, R.; Cerrato, C.; Bionda, R.; Provenzale, A. Effects of Temperature Rise on Multi-Taxa Distributions in Mountain Ecosystems. Diversity 2020, 12, 210. https://doi.org/10.3390/d12060210
Viterbi R, Cerrato C, Bionda R, Provenzale A. Effects of Temperature Rise on Multi-Taxa Distributions in Mountain Ecosystems. Diversity. 2020; 12(6):210. https://doi.org/10.3390/d12060210
Chicago/Turabian StyleViterbi, Ramona, Cristiana Cerrato, Radames Bionda, and Antonello Provenzale. 2020. "Effects of Temperature Rise on Multi-Taxa Distributions in Mountain Ecosystems" Diversity 12, no. 6: 210. https://doi.org/10.3390/d12060210
APA StyleViterbi, R., Cerrato, C., Bionda, R., & Provenzale, A. (2020). Effects of Temperature Rise on Multi-Taxa Distributions in Mountain Ecosystems. Diversity, 12(6), 210. https://doi.org/10.3390/d12060210