Wildlife Diversity and Relative Abundance Among a Variety of Adjacent Protected Areas in the Northern Talamanca Mountains of Costa Rica


Abstract
1. Introduction
2. Methods
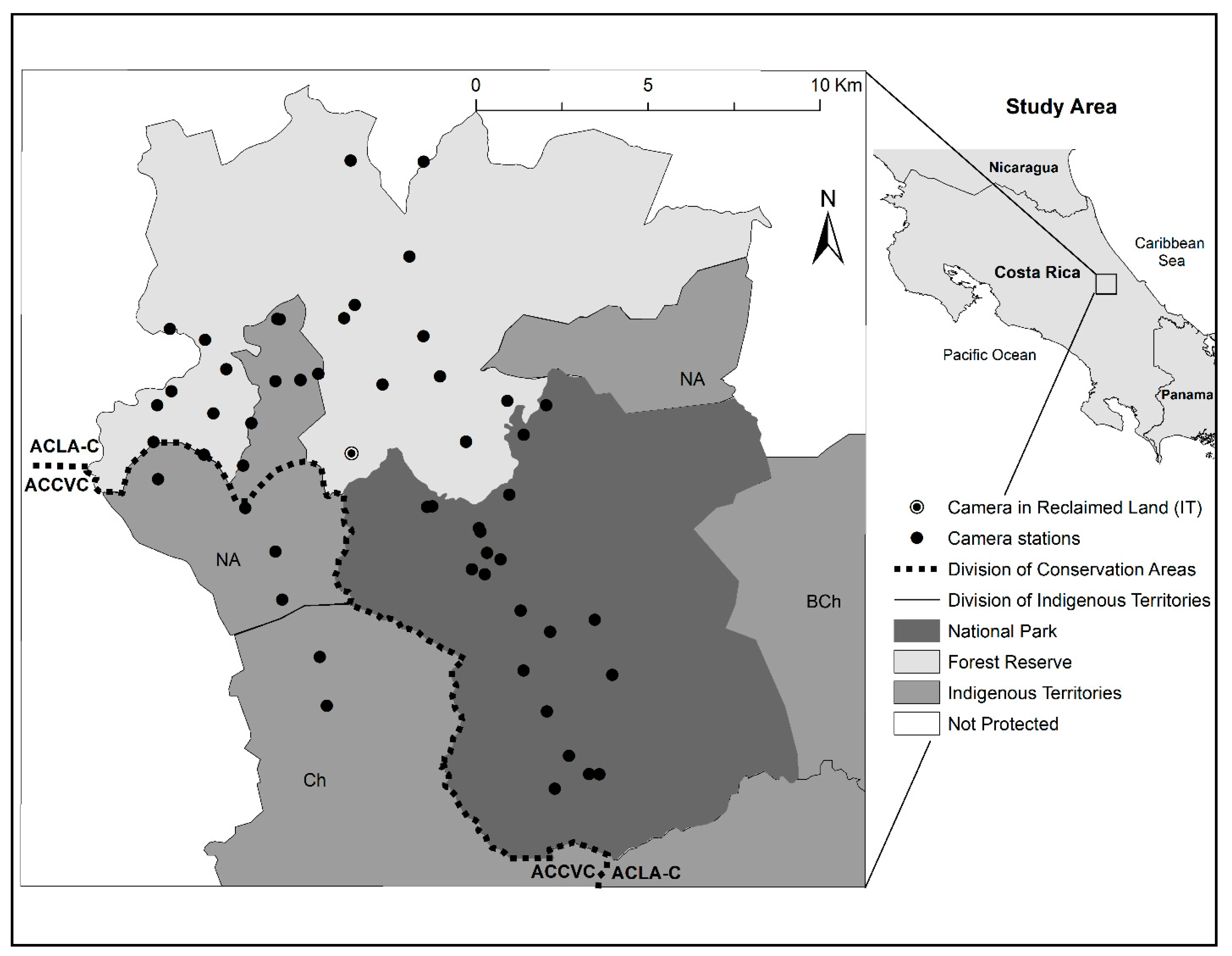
2.1. Study Area
2.2. Camera Deployment and Data Collection
2.3. Analysis
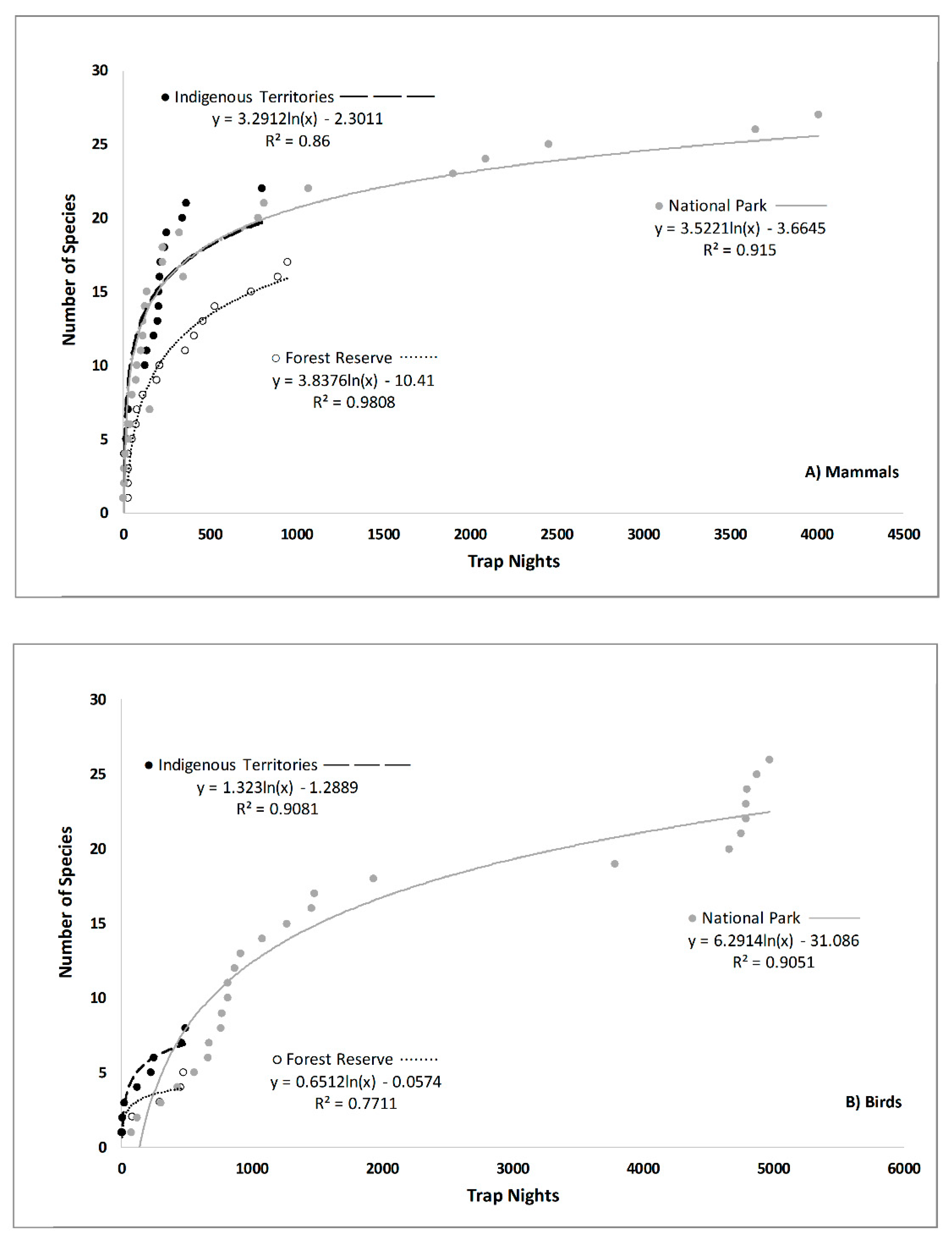
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dudley, N. Guidelines for Applying Protected Area Management Categories; Dudley, N., Ed.; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N. Effectiveness of terrestrial protected areas in reducing habitat lost and population declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Gray, C.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [PubMed]
- Bruner, A.G.; Gullison, R.E.; Rice, R.E.; da Fonseca, G.A.B. Effectiveness of parks in protecting tropical biodiversity. Science 2001, 291, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Peres, C.A. Why we need megareserves in Amazonia. Conserv. Biol. 2005, 19, 728–733. [Google Scholar] [CrossRef]
- Galetti, M.; Giacomini, H.C.; Bueno, R.S.; Bernardo, C.S.S.; Marques, R.M.; Bovendorp, R.S.; Steffler, C.E.; Rubim, P.; Gobbo, S.K.; Donatti, C.I.; et al. Priority areas for the conservation of Atlantic forest large mammals. Biol. Conserv. 2009, 142, 1229–1241. [Google Scholar] [CrossRef]
- Ferraro, P.J.; Hanauer, M.M.; Miteva, D.A.; Canavire-Bacarreza, G.J.; Pattanayak, S.K.; Sims, K.R.E. More strictly protected areas are not necessarily more protective evidence from Bolivia, Costa Rica Indonesia, and Thailand. Environ. Rest. Lett. 2013, 8, 025011. [Google Scholar] [CrossRef]
- Sistema Nacional de Áreas de Conservación. Available online: http://www.sinac.go.cr/EN-US/asp/Pages/default.aspx (accessed on 27 February 2020).
- Pringle, R.M. Upgrading protected areas to conserve wild biodiversity. Nature 2017, 546, 91–99. [Google Scholar] [CrossRef]
- Soulé, M.; Noss, R. Rewilding and biodiversity: Complementary goals for continental conservation. Wild Earth 1998, 8, 18–28. [Google Scholar]
- Di Minin, E.; Hunter, L.T.B.; Balme, G.A.; Smith, R.J.; Goodman, P.S.; Slotow, R. Creating larger and better-connected protected areas enhances the persistence of big game species in the Maputaland Pondoland-Albany biodiversity hotspot. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Castilho, C.S.; Hackbart, V.C.S.; Pivello, V.R.; dos Santos, R.F. Evaluating landscape connectivity for Puma concolor and Panthera onca among Atlantic forest protected areas. Environ. Manage. 2015, 55, 1377–1389. [Google Scholar] [CrossRef]
- Atlas digital de Costa Rica 2014. E. Ortiz-Malavasi. Instituto Tecnológico de Costa Rica (ITCR), Laboratorio de Sistemas de Información Geográfica, Escuela de Ingeniería Forestal, ITCR. Cartago, CR. Available online: https://repositoriotec.tec.ac.cr/handle/2238/6749 (accessed on 28 March 2020).
- Hedström, I. Talamanca Indómita: Relato-guía de campo del Parque Nacional Barbilla, Costa Rica (Untamed Talamanca: Chronicle—Fieldguide to Barbilla Nacional Park, Costa Rica), 1st ed.; Fundación Nairi: San José, Costa Rica, 2006; p. 428. [Google Scholar]
- Food and Agriculture Organization. Evaluación de los recursos forestales mundiales 2010, Informe nacional Costa Rica. Roma, Italia. Available online: https://www.sirefor.go.cr/pdfs/publicaciones/2010_FAO_Informe_Evaluacion_Recursos_Forestales_Mundiales_Costa_Rica_FRA_2010.pdf (accessed on 28 February 2020).
- Ley Indígena 1977. Art 6, Law No. 6172. Available online: http://www.conai.go.cr/documentos.html (accessed on 27 February 2020).
- Huston, M.A. Biological diversity: The coexistence of species on changing landscapes; Cambridge University Press: Cambridge, UK; p. 681.
- Terborgh, J.; Lopez, L.; Nunez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecological meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef] [PubMed]
- Galetti, M.; Donatti, C.I.; Pires, A.S.; Guimarães, P.R.; Jordano, P. Seed survival and dispersal of an endemic Atlantic forest palm: The combined effects of defaunation and forest fragmentation. Bot. J. Linn. Soc. 2006, 151, 141–149. [Google Scholar] [CrossRef]
- Stoner, C.; Caro, T.; Mduma, S.; Mlingwa, C.; Sabuni, G.; Borner, M.; Schelten, C. Changes in large herbivore populations across large areas of Tanzania. Afr. J. Ecol. 2007, 45, 202–215. [Google Scholar] [CrossRef]
- Peres, C.A.; Palacios, E. Basin-wide effects of game harvest on vertebrate population densities in Amazonian forests: Implications for animal-mediated seed dispersal. Biotropica 2007, 39, 304–315. [Google Scholar] [CrossRef]
- Carrillo, E.; Wong, G.; Cuarón, A.D. Monitoring Mammal Populations in Costa Rican Protected Areas under Different Hunting Restrictions. Conserv. Biol. 2000, 14, 1580–1591. [Google Scholar] [CrossRef]
- Abrahams, M.I.; Peres, C.A.; Costa, H.C.M. Measuring local depletion of terrestrial game vertebrates by central-place hunters in rural Amazonia. PLoS ONE 2017, 12, 1–25. [Google Scholar] [CrossRef]
- Crooks, K.R.; Soulé, M.E. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 1999, 400, 463–466. [Google Scholar] [CrossRef]
- Sistema Nacional de Áreas de Conservación. Available online: http://www.sinac.go.cr/ES/buscador/Paginas/default.aspx (accesed on 12 October 2017).
- Bernal, L.; García, P. Viabilidad de realización de un proyecto de turismo rural comunitario en las comunidades indígenas de Nairi-Awari. Unpublished work, 2007. [Google Scholar]
- Sáenz-Bolaños, C.; Montalvo, V.; Fuller, T.K.; Carrillo, E. Records of black jaguars at Parque Nacional Barbilla. CatNews 2015, 62, 38–39. [Google Scholar]
- Instituto Nacional de Estadística y Censos [Costa Rica] (INEC). X Censo Nacional de Población y IV de vivienda: Territorios indígenas/Instituto Nacional de Estadística y Censos, 1st ed.; INEC: San José, Costa Rica, 2013; p. 56. [Google Scholar]
- Ministerio de Planificación Nacional y Política Económica (MIDEPLAN). Análisis de desarrollo: Población Indígena en Cifras. Unpublished work, 2015. [Google Scholar]
- Blake, J.G.; Mosquera, D. Camera trapping on and off trails in lowland forest of eastern Ecuador: Does location matter? Mastozool. Neotrop. 2014, 21, 17–26. [Google Scholar]
- Blake, J.G.; Mosquera, D.; Loiselle, B.A.; Romo, D.; Swing, K. Effects of human traffic on use of trails by mammals in lowland forest of eastern Ecuador. Neodiversity 2017, 3, 57–64. [Google Scholar] [CrossRef]
- Wainwright, M. The Mammals of Costa Rica: A Natural History field Guide; Cornell University Press: New York, NY, USA, 2007. [Google Scholar]
- Garrigues, R.; Dean, R. The Birds of Costa Rica: A Field Guide, 2nd ed.; Zona Tropical Cornell University Press: New York, NY, USA, 2014. [Google Scholar]
- O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. Camera-traps in Animal Ecology: Methods and Analyses, 1st ed.; O’Connell, A.F., Nichols, J.D., Karanth, K.U., Eds.; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Hedström, I. Preliminary check list of observed bird species within Tapir River Private Wild Life Reserve. Unpublished work, 2011. [Google Scholar]
- Alvarado, R.; Escobar, B.; Ramos, J.; Sagastume, V. Anfibios, aves y mamíferos del Parque Nacional Barbilla y una propuesta de indicadores para evaluar su integridad ecológica; Alvarado, R., Ed.; ICOMVIS-Universidad Nacional de Costa Rica: Heredia, Costa Rica, 2017. [Google Scholar]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Influences of Anthropogenic Resources on Raccoon (Procyon lotor) Movements and Spatial Distribution. J. Mammal. 2004, 85, 483–490. [Google Scholar] [CrossRef]
- Schulte-Hostedde, A.I.; Mazal, Z.; Jardine, C.M.; Gagnon, J. Enhanced access to anthropogenic food waste is related to hyperglycemia in raccoons (Procyon lotor). Conserv. Physiol. 2018, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hass, C.C.; Valenzuela, D. Anti-predator benefits of group living in white-nosed coatis (Nasua narica). Behav. Ecol. Sociobiol. 2002, 51, 570–578. [Google Scholar] [CrossRef]
- Borge, C.; Martínez, J. El Pago por Servicios Ambientales en Territorios Indígenas de Costa Rica. PES Learning Paper 2009-1S. (Spanish). Payments for Environmental Services (PES) learning paper. DC World Bank Group: Washington, DC, USA, 2009. Available online: http://documents.worldbank.org/curated/en/440201468261580966/El-pago-por-servicios-ambientales-en-territorios-ind-237-genas-de-Costa-Rica (accessed on 28 February 2020).
- Hunter, P. The human impact on biological diversity. How species adapt to urban challenges sheds light on evolution and provides clues about conservation. EMBO Rep. 2007, 8, 316–318. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| National Park | Forest Reserve | Indigenous Territories | |
|---|---|---|---|
| Elevation (m) | 729 (295–1281) a | 421 (285–630) | 665 (303–1083) |
| Road density (km/km2) | 0.0 | 0.29 | 0.10 |
| Distance to nearest road (km) | 0.0 | 1.38 (0.10–3.30) | 2.79(1.49–4.36) |
| Vegetation cover type (%) | |||
| Primary forest | 93 | 83 | 92 |
| Secondary forest | 5 | 0 | 1 |
| Forest plantation | 0 | 1 | 0 |
| Non-forested b | 2 | 16 | 7 |
| Protected area composition (%) | |||
| National Park | 85 | 5 | 4 |
| Forest Reserve | 10 | 70 | 34 |
| Indigenous Territories | 4 | 12 | 59 |
| Not Protected | 0 | 13 | 3 |
| No. of Species Detected | ||||||
|---|---|---|---|---|---|---|
| Season | Protected Area | No. of Camera Stations | No. of Trap Nights | Mammal a | Bird | Total |
| May–August | National Park | 19 | 2630 | 29 | 17 | 46 |
| September–April | National Park | 21 | 5820 | 30 | 26 | 56 |
| Forest Reserve | 17 | 796 | 19 | 5 | 24 | |
| Indigenous Territories | 17 | 874 | 24 | 8 | 32 | |
| Both | All | 55 | 10,120 | 34 | 34 | 68 |
| May–August | September–April | ||||
|---|---|---|---|---|---|
| Area with Highest Photo Rate | Species a | National Park (NP) (19/2630) b | National Park (NP) (21/5820) | Forest Reserve (FR) (17/796) | Indigenous Territories (IT) (17/874) |
| National | Agouti c | 37.98 | 21.94 | 13.19 | 19.11 |
| Park | Paca | 1.44 | 3.21 | 0.25 | 1.03 |
| Ocelot | 3.95 | 4.35 | 2.01 | 2.29 | |
| Puma | 1.14 | 1.53 | 0 | 0 | |
| Jaguar | 1.06 | 1.29 | 0 | 0.11 | |
| Great Tinamou d | 7.15 | 9.54 | 1.38 | 2.75 | |
| Human (Poacher e) | 2.51 | 2.41 | 0 | 0 | |
| Indigenous | Gray Four-eyed Opossum | 0 | 0 | 0 | 0.80 |
| Territory | Nine-banded Armadillo f | 2.28 | 1.27 | 49 | 3.09 |
| Tamandua | 0.08 | 0.15 | 1.13 | 0.57 | |
| Red-tailed Squirrel | 1.41 | 1.75 | 063 | 3.78 | |
| Unknown rat g | 148 | 1.53 | 4.65 | 5.95 | |
| Domestic dog | 1.48 | 0.84 | 1.01 | 2.4 | |
| Domestic pig | 0.34 | 0.15 | 0.13 | 5.26 | |
| Human (Local h) | 1.98 | 1.61 | 0.50 | 5.61 | |
| Black-earned Wood-Quail | 0 | 0 | 0 | 0.23 | |
| Olive-backed Quail-Dove | 0.61 | 0.57 | 0.13 | 1.72 | |
| Spotted Antbird | 0 | 0.02 | 0 | 0.34 | |
| Streak-chested Antpitta | 0 | 0 | 0 | 0.23 | |
| Wood Thrush | 0 | 0.07 | 0.13 | 2.97 | |
| Forest | White-nosed Coati | 0.57 | 0.43 | 2.26 | 0.34 |
| Reserve | Northern Raccoon | 0 | 0.03 | 0.38 | 0 |
| Chestnut-backed Antbird | 0 | 0 | 0.25 | 0 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáenz-Bolaños, C.; Fuller, T.K.; Carrillo J., E. Wildlife Diversity and Relative Abundance Among a Variety of Adjacent Protected Areas in the Northern Talamanca Mountains of Costa Rica. Diversity 2020, 12, 134. https://doi.org/10.3390/d12040134
Sáenz-Bolaños C, Fuller TK, Carrillo J. E. Wildlife Diversity and Relative Abundance Among a Variety of Adjacent Protected Areas in the Northern Talamanca Mountains of Costa Rica. Diversity. 2020; 12(4):134. https://doi.org/10.3390/d12040134
Chicago/Turabian StyleSáenz-Bolaños, Carolina, Todd K. Fuller, and Eduardo Carrillo J. 2020. "Wildlife Diversity and Relative Abundance Among a Variety of Adjacent Protected Areas in the Northern Talamanca Mountains of Costa Rica" Diversity 12, no. 4: 134. https://doi.org/10.3390/d12040134
APA StyleSáenz-Bolaños, C., Fuller, T. K., & Carrillo J., E. (2020). Wildlife Diversity and Relative Abundance Among a Variety of Adjacent Protected Areas in the Northern Talamanca Mountains of Costa Rica. Diversity, 12(4), 134. https://doi.org/10.3390/d12040134