Abstract
Protected areas are intended to achieve the long-term conservation of nature, but not all such areas are equal in their effectiveness because of their varying regulation of human activities. In Costa Rica, we assessed mammal and bird species presence and relative abundance in three protected areas in the northern Talamanca Mountains. In this humid tropical forest area, we placed camera traps in an adjacent national park, forest reserve, and indigenous territories, each with a different mix of human activities. In 10,120 trap nights, we obtained 6181 independent photos of mostly mammals (34 species other than humans) and birds (34 species). Species with greater abundance or only occurrence in the national park were mammals and birds commonly hunted outside of the park, large carnivores rarely documented in other areas, and poachers. Species found more often outside of the park were medium-sized mammals, some birds, and domestic mammals. We conclude that even in the same ecological area, varying regulations related to type of protected area have significant effects on some mammal and bird species abundances and occurrences, and thus need to be considered when assessing the overall effectiveness of protection as a conservation strategy.
1. Introduction
The International Union for Conservation of Nature [1] defines a protected area as a “clearly defined geographical space, recognized, dedicated and managed, through legal or other effective means, to achieve the long-term conservation of nature with associated ecosystem services and cultural values.” Thus, protected areas are considered important for maintaining biodiversity and the integrity of the ecosystems [2,3]. Large natural areas such as national parks and reserves are essential for conserving wildlife populations [4,5]; large mammals, especially top predators and large herbivores, are often able to thrive in these protected areas because they are intended to provide shelter from human persecution and anthropogenic habitat changes [6]. There are, however, several categories of protected areas, each of which have different restrictions on human activities [7,8]. Right now, in Costa Rica and other countries, some protected areas are no longer sufficient to provide protection to large mammals [9], though establishing or maintaining landscape connectivity between multiple protected areas can mitigate inadequate protection in a single area [10,11,12].
Costa Rica has nine official types of protected areas [8,13] as well as indigenous territories that are often considered a kind of protected area [14], each of which has different restrictions on anthropogenic activities. For example, national parks are “areas [intended] to protect outstanding natural and scenic areas of national and international significance for scientific, educational, and recreational use. They are relatively large natural areas not materially altered by human activity where extractive resources use is not allowed” [14]. Forest reserves are forests in which the main function is the production of timber and those forest lands that by nature are especially suitable for that purpose [15]; more and less-restricted human activities are allowed than in national parks. The aim for indigenous territories is “conservation of cultures and their environments and the protection of life systems in these communities and the way natural resources are used” [14]; they are autonomous, there are location-specific regulations (e.g., hunting is allowed only for the indigenous population), and land can be used in any way without the need for governmental permission [16].
Costa Rica has designated about 32% of its territory as some sort of protected area. Many of these areas encompass humid ecosystems [17] where herbivores play an important role as seed dispersers and thus predators not only affect prey populations, but also shape patterns of plant distribution and diversity [18,19,20]. The population density of tropical forest vertebrates largely depends on climatic factors such as elevation, floristic composition, and net primary productivity sources. Human disturbance (e.g., hunting pressure and land-use change) also affects the density and distribution of vertebrate species [6,21], and thus variation in such disturbance among different types of protected areas can result in variation in the density and distribution of vegetation [7] and wildlife [22].
Here, we report the results of an assessment of mammal and bird species presence and relative abundance in three protected areas in the northern Talamanca Mountains of Costa Rica. In this area of humid tropical forest, we placed camera traps in an adjacent National Park, Forest Reserve, and Indigenous Territories, each with a varying mix of human activities. We anticipated that, in this area of similar basic ecological conditions, the effects of varying levels of protection would result in higher diversity and abundance of mammals and birds in the areas with more protection [3], and a change in species presence in some areas as the result of hunting by humans of certain species [23], followed by an ecological cascade effect (e.g., mesopredator release; [24]).
2. Methods
2.1. Study Area
The Barbilla Sector in the northern Talamanca Mountains of Costa Rica is in the Volcánica Central-Talamanca Biological Corridor between Limón and Cartago Provinces [13] and includes three different kinds of protected areas, each one with a specific category of management; they include a National Park (NP), a Forest Reserve (FR), and Indigenous Territories (IT; Figure 1). The main objective of the 120-km2 Barbilla National Park (BNP) is the conservation of the tropical humid forest that provides a large proportion of water production along the Caribbean slope of the Talamanca Mountain Range [25]. Precipitation in the national park and the surrounding areas occurs throughout the year, with relatively more rain during November–December and less in March, and averages ~4000 mm annually [26]. In the Pacuare River Forest Reserve (PRFR) to the north of the Barbilla National Park, an area of virgin forest was logged in the mid 1970s by means of 15 km of new unpaved roads, which subsequently allowed for additional roads and settlements in the area [14]. Currently, the Forest Reserve has some grasslands for cattle, forest plantations, human settlements, and eco-lodges. We also sampled in adjacent indigenous territories including the Nairi Awari Indigenous Territory and the Chirripó Indigenous Territory to the west and northwest of the national park. In both these areas, there is a population of 6814 Cabécar, the second largest indigenous group in Costa Rica [24,25,26,27,28,29]. There are relatively few settlements, and the Cabécar mainly hunt, plant bananas, cassavas and grains, and also raise pigs, chickens, and cows.
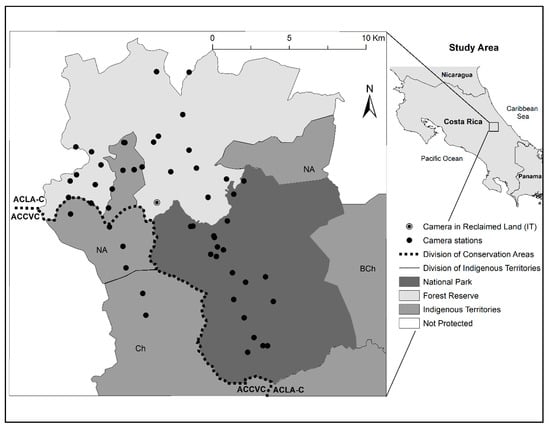
Figure 1.
Location of camera stations within Barbilla National Park (dark gray), Indigenous Territories (medium gray; Nairi Awari [NA], Chirripó [Ch], and Bajo Chirripó [BCh]), and the Pacuare River Forest Reserve (light gray) in the Conservation Area Cordillera Volcánica Central [ACCVC] and Conservation Area La Amistad-Caribe [ACLA-C] in the northern Talamanca Mountains of Costa Rica.
Quantitatively, we calculated landscape characteristics (using geographic information system data from ArcMap 10.2.2; ESRI 2014) of areas within 2 km of all camera traps set inside each of three adjacent protected areas in the northern Talamanca Mountains of Costa Rica (Table 1). Elevations in each of the areas are generally similar, though on average the Forest Reserve is at lower elevations. There are no roads within the national park, and almost three times as many in the Forest Reserve compared to the indigenous territories, and as a result of logging and clearing, the Forest Reserve was least forested. The composition of protected area type near each of our camera categories varied because cameras were often located near borders of the protected areas (Figure 1); a number of Forest Reserve cameras were also near unprotected areas.

Table 1.
Landscape characteristics of areas within 2 km of all camera traps set inside each of three adjacent protected areas in the northern Talamanca Mountains of Costa Rica.
2.2. Camera Deployment and Data Collection
We placed cameras (Bushnell Trophy Cam®) on two different trail types to maximize the trap success because wildlife is known to use both types [30,31]. Human trails were trails regularly used by tourists, researchers, rangers, and local people and were 2–3 m in width. Animal paths were trails mostly used by wildlife and were narrower (1½–2 m) than human trails. Of the 55 camera traps we deployed to maximize the captures, 26 were on human trails (NP = 7, FR = 7, IT = 12) and 29 were on the animal paths (NP = 14, FR = 10, IT = 5); two of 17 camera stations in the IT were moved to within 100 m of where the camera was located the previous year. Another camera location on the map appeared as if it was in FR (Figure 1), but was actually in reclaimed indigenous territory land that had not yet been updated in Atlas Digital.
At each deployment location (camera station), an unbaited camera was placed ~0.5 m above the ground, 2–4 m from the center of the trail, and with an unobstructed view of the trail. Cameras were active 24 h per day and when activated they recorded a 30-s video with a minimum of 1 min between consecutive videos (2013–2016), or a series of three still photos taken two seconds apart (all other years). Cameras were set in the national park year-round during 2013 through 2016, in the Forest Reserve during April–May 2009 and September–December 2012, and in the Indigenous Territories during April–May 2009 and September–December 2011. During 2009 and 2011, cameras were checked every month to change batteries and collect files because the cameras were not as technologically advanced as in the recent years, when we checked every three months to change the batteries.
During all years of study, we lost some cameras because some stopped working due to high humidity, some were damaged by jaguars or vandalized by people, and at least 10 were stolen. For each camera that was deployed, we determined the number of trap nights; trap nights were the total days a camera worked.
2.3. Analysis
Photos or videos were considered an independent record of a species if they were: (1) taken at least 30 min apart (e.g., a series of three photos of the same species taken in consecutive seconds = 1 photo event); (2) consecutive photos of the same species could be identified as different individuals (spots, scars, horns/antlers, sex) and not part of the same group (e.g., 15 min apart, going in opposite directions = two photo events); or (3) photos of the same species separated by photos of a different species (e.g., species 1, followed 2 min later by a species 2, followed 5 min later by species 1 = one species with two photo events and another species with one photo event). Mammal and bird species were identified and named using local field guides [32,33]. Photographs of humans were classified as: (1) Research-Protection: researchers and park rangers; (2) Local persons who live, work, or transit an area without hunting equipment or carrying killed wild animals; (3) Poacher: persons with hunting/fishing equipment (e.g., rifle, blowgun, harpoon), or carrying killed wild animals; (4) Tourist, hikers or persons with photo equipment; and (5) Unclassified people: persons that could not be classified as one of the above.
We recognize that our samples were relatively small and did not justify extensive statistical analysis. However, descriptive presentations of the data and simple statistical comparisons were justified and both could still reveal and suggest important distributional differences. Thus, for a basic assessment of species diversity, we tallied up the total number of species for the two seasons (September–April and May–August) in the National Park (we only sampled in one season, September–April, in the IT and FR), and assessed the differences between them. Then, we tested for differences during the September–April season between the three protected area types. To assess possible total species diversity differences, we plotted accumulation curves by counting how many traps nights were necessary to find a new species in each area, and by season in the national park. For assessing differences in relative species abundances among areas during the same season, we calculated relative abundance indices (RAI; no. of independent photos/100 trap nights [34], and used Chi-squared tests (p < 0.001) to compare species-specific differences in the RAI rates among areas and between seasons for the national park.
3. Results
A total of 55 different camera stations in the three areas were monitored for a total of 10,120 trap nights (Table 2). Though the number of camera stations was fairly similar in each area, most effort focused on the national park (8450), with much less in the Forest Reserve (796), and the Indigenous Territories (874). From this effort, we obtained 6181 independent photos, mostly of mammal (including two domestic) and bird species. Agoutis (scientific names listed in the Supplementary Material; total n = 2548 independent photos) and great tinamous (n = 778 photos) were by far the most commonly photographed mammal and bird species, respectively, in all areas (Supplementary Figure S1). Three mammal species and 11 bird species were photographed only once.
Table 2.
Summary data for camera-trapping efforts during 2009–2016 in three adjacent protected areas in the northern Talamanca Mountains of Costa Rica.
Seasonal species accumulation curves in the national park were very similar for mammals (Supplementary Figure S2), but were perhaps lower for birds during the May through August period when migratory species may have left for northern breeding ranges. Our sampling comparisons among areas during the 8-month September–April season (Figure 2) suggested that, for mammals, there may be fewer species in the Forest Reserve. Our samples for birds outside of the National Park were too small for meaningful comparison, but two periods of rapid species accumulations occurred over the course of the multiple year sampling in the park.
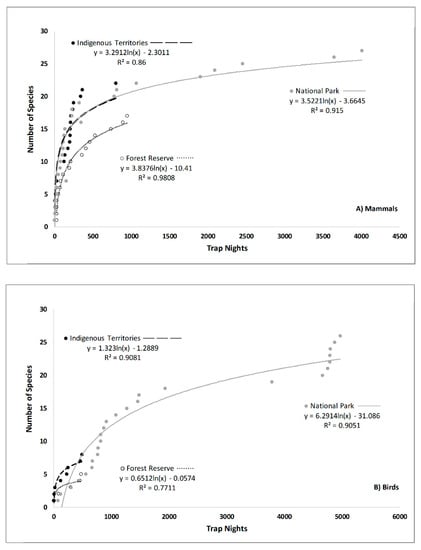
Figure 2.
Trend lines and correlations for the number of camera trap nights (effort) versus total cumulative number of (A) mammal and (B) bird species photographed (diversity) in the National Park (solid line, solid circle), Indigenous Territories (dotted line, gray circle), and Forest Reserve (dash line, open circle) during September–April in the northern Talamanca Mountains of Costa Rica.
Not counting humans, we documented 34 species of mammals including an unidentified bat, rat, and mouse, and two domestic species (dog and pig; Supplementary Table S1). We also documented 34 species of birds including unidentified nightjar and hummingbird (Supplementary Table S1).
Seasonal differences in species abundance in the National Park were limited to a few species (Table 3). Agoutis and nine-banded armadillos were photographed more often during May–August, and great tinamous were photographed more often during September–April.
Table 3.
Areas (and seasons) with highest photo rates (no. of independent photos/100 trap nights) of mammal and bird species, including humans, detected by camera-trapping efforts during 2009–2016 in three adjacent protected areas in the northern Talamanca Mountains of Costa Rica. Statistical differences (p < 0.001) between seasons for BNP are indicated in italics; statistical differences among the three protected areas are identified in bold.
Statistically different photo rates among protected areas were identified for 23 species (Table 3). More ocelots, pumas, jaguars, pacas, agoutis, and great tinamous were recorded in the National Park; agouti abundance was also high in Indigenous Territories compared to Forest Reserve. In Indigenous Territories, photo rates were also higher for five other wild and two domestic mammals (dogs and pigs), and five bird species. In the Forest Reserve, white-nosed coatis and northern raccoons were more common, as was the chestnut-backed antbird; unknown rats and nine-banded armadillos were also more common in Forest Reserve than in the National Park. Poachers were only recorded in the National Park, and local people were most often photographed in the Indigenous Territories. We also noted that even though sampling effort was substantially greater in the National Park, commonly hunted species including white-lipped peccaries and tapirs (among a total of nine mammal species), and slaty-breasted tinamou, crested guan, black guan, and great curassow (among a total of 24 bird species) were photographed only in the National Park, although statistical differences in rates among areas were not identified.
4. Discussion
We recorded only 30 identified bird species of more than 225 previously recorded in BNP and its surroundings [35] and, as expected, most of those were ground-dwelling birds [33]. The seven bird species in Table 3 are species associated with mature forest, and thus are vulnerable to changes in the ecosystem. Many more mammal species occur in the area than we recorded [36], even though most of those not documented are bats, small rodents, and primates or other arboreal species.
The differences in relative abundances of species we documented suggest that anthropogenic forces related to protected area regulation have a large impact on many species. Species with greater abundance or only occurrence in the national park on both types of trails were mammals and bird species commonly hunted outside of the park, large carnivores rarely documented in other areas, and poachers. Species found more often outside of the Park were medium-sized mammals, some birds, and domestic mammals. Local people were identified in all areas, but poachers were only documented in the National Park, perhaps because the most commonly hunted species (large birds such as great curassow, crested guan, black guan, and great tinamou, and mammals such as paca, tapir, red brocket deer, collared peccary, and white-lipped peccary) are only found there.
The prevalence of large birds, jaguars, and pumas in the National Park can be attributed to the fact that it has been largely free from human pressure, provides food (paca, agouti, and tinamou for carnivores, and a variety of seeds and small vertebrates and invertebrates for birds), and thus habitat resources necessary to fulfill required daily activities. The near absence of jaguars and pumas outside the National Park may be due to reduced prey species abundance as a result of hunting. Moreover, one big threat for big cats outside the National Park is retaliatory killing by humans when they lose a cow, pig, or other domestic animal, or even just from fear. This may also be true for ocelots, though they still appear relatively common outside of the Park.
The two mammal species that had higher abundance in the FR were white-nosed coati and raccoon, known to be associated with human settlements and the associated food resources [37,38]; in the FR it is possible for them to obtain dog and cow food, crops, etc. Additionally, their increased abundance outside of the park may be due to a mesopredator release effect [24] (i.e., the absence or low frequency of predation by felids [39]) that have been reduced by humans resulting in increased abundance of smaller competitors.
Bird species do not suffer retaliations as do big cats, but they are persecuted by poachers and indigenous people and this is likely to be why they were not detected outside the National Park. On the other hand, some species with higher RAI outside the park were species associated with mature forest and we expected to find them more in the NP than IT. However, both areas were similar in vegetation structure and thus their relative abundances may simply reflect differences in available food resources. It will be important for future studies to sample vegetation and see how its composition may influence the interpretation of our findings.
Our results showed that even though the ecosystem was similar among the protected areas we surveyed, the amount of forest cover and the influence of human activities varied (Table 1), and species abundances differed. The cover types were similar in the National Park and Indigenous Territories, whereas the forest reserve had less primary forest and some exotic forest plantations, and more human presence and activities such as roads. These differences in the PRFR compared with BNP and IT were also related to the regulations of each area. On the other hand, Cabecar indigenous associations have opted to receive payments for environmental services (PSA by the Spanish abbreviation) as income for protecting the forest [40], resulting in a substantial area of forest being protected under this scheme, and increasing habitat connectivity for wildlife.
The characteristics of each area influence the species richness and composition. The National Park was the most diverse area including very elusive and rare species listed in the IUCN red list. For example, species that require a well-protected forest such as tapir, white-lipped peccary, oncilla, jaguar, and paca are present in this area. Paradoxically, the seemingly well-protected forest shelters important game species and thus appears to currently have more poaching pressure compared with the Forest Reserve and Indigenous Territories. In the long run, the poaching pressure in the National Park is something to pay attention to in order to avoid long-term effects [41].
5. Conclusions
The distribution of mammals and birds identified in this study suggest ways to anticipate the effects of designating specific kinds of protected areas. Better protected sites and areas with enough vegetation cover to sustain them such as the Barbilla National Park seem optimal. Although Indigenous Territories had forest cover similar to the Park, the differences observed in wildlife distribution were likely due to the number of people and the activities allowed in each of them. For example, indigenous people can and do hunt in their territories, thus the lower abundance of certain species in those area. In the Forest Reserve, some species that were more common seemed related to what might be expected in more populated, or even urban, areas. Therefore, we can conclude that even in the same ecological area, varying regulations related to type of protected area have significant effects on some mammal and bird species abundances and occurrences. These effects need to be considered when assessing the overall effectiveness of a protected area for wildlife conservation.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/12/4/134/s1, Figure S1: Total number of independent photos obtained of A) mammal (including humans; *see footnotes for Figure S1) and B) bird species in a National Park, Indigenous Territories, and Forest Reserve adjacent to each other in the northern Talamanca Mountains of Costa Rica. Figure S2: Trend lines and correlations for the number of camera trap nights (effort) versus total cumulative number of A) mammal and B) bird species photographed (diversity) in the National Park during September-April (solid line, solid circle), and May-August (dash line, open circle) in the northern Talamanca Mountains of Costa Rica. Table S1: Photo rates (no. of independent photos/100 trap nights) of mammal species, including humans, detected by camera-trapping efforts during 2009-2016 in three adjacent protected areas in the northern Talamanca Mountains of Costa Rica. Significant differences (P<0.001) between seasons for the National Park are indicated in italics; differences among the three protected areas during September-April are identified in bold. Table S2: Photo rates (no. of independent photos/100 trap nights) of bird species detected by camera-trapping efforts during 2009-2016 in three adjacent protected areas in the northern Talamanca Mountains of Costa Rica. Significant differences (P<0.001) between seasons for the National Park are indicated in italics; differences among the three protected areas during September-April are identified in bold.
Author Contributions
Conceptualization, C.S.-B.; methodology, C.S.-B.; formal analysis, C.S.-B and T.K.F.; investigation, C.S.-B..; resources, C.S.-B.; writing—original draft preparation, C.S.-B and T.K.F.; writing—review and editing, C.S.-B., T.K.F. and E.C.J.; visualization, C.S.-B.; supervision, T.K.F.; project administration, C.S.-B.; funding acquisition, C.S.-B. and E.C.J. All authors have read and agreed to the published version of the manuscript.
Funding
This project was generously supported with: equipment, transportation, field assistance, and housing from Pacuare Lodge; equipment donated by Ruffing Montessori School; field assistance funding from Panthera; and per diem costs and transportation provided by Universidad Nacional de Costa Rica. A fellowship from Universidad Nacional de Costa Rica and the Consejo Nacional para Investigaciones Científicas y Tecnológicas of Ministerio de Ciencia y Tecnología supported CS-B’s PhD studies and page charges.
Acknowledgments
We wish to thank everyone who was involved in the different stages of this research including students in the field and office, rangers, field assistants, drivers, and the Pacuare Lodge. Additional thanks go to CONICIT-MICIT and the National University of Costa Rica for essential financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dudley, N. Guidelines for Applying Protected Area Management Categories; Dudley, N., Ed.; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N. Effectiveness of terrestrial protected areas in reducing habitat lost and population declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Gray, C.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [PubMed]
- Bruner, A.G.; Gullison, R.E.; Rice, R.E.; da Fonseca, G.A.B. Effectiveness of parks in protecting tropical biodiversity. Science 2001, 291, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Peres, C.A. Why we need megareserves in Amazonia. Conserv. Biol. 2005, 19, 728–733. [Google Scholar] [CrossRef]
- Galetti, M.; Giacomini, H.C.; Bueno, R.S.; Bernardo, C.S.S.; Marques, R.M.; Bovendorp, R.S.; Steffler, C.E.; Rubim, P.; Gobbo, S.K.; Donatti, C.I.; et al. Priority areas for the conservation of Atlantic forest large mammals. Biol. Conserv. 2009, 142, 1229–1241. [Google Scholar] [CrossRef]
- Ferraro, P.J.; Hanauer, M.M.; Miteva, D.A.; Canavire-Bacarreza, G.J.; Pattanayak, S.K.; Sims, K.R.E. More strictly protected areas are not necessarily more protective evidence from Bolivia, Costa Rica Indonesia, and Thailand. Environ. Rest. Lett. 2013, 8, 025011. [Google Scholar] [CrossRef]
- Sistema Nacional de Áreas de Conservación. Available online: http://www.sinac.go.cr/EN-US/asp/Pages/default.aspx (accessed on 27 February 2020).
- Pringle, R.M. Upgrading protected areas to conserve wild biodiversity. Nature 2017, 546, 91–99. [Google Scholar] [CrossRef]
- Soulé, M.; Noss, R. Rewilding and biodiversity: Complementary goals for continental conservation. Wild Earth 1998, 8, 18–28. [Google Scholar]
- Di Minin, E.; Hunter, L.T.B.; Balme, G.A.; Smith, R.J.; Goodman, P.S.; Slotow, R. Creating larger and better-connected protected areas enhances the persistence of big game species in the Maputaland Pondoland-Albany biodiversity hotspot. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Castilho, C.S.; Hackbart, V.C.S.; Pivello, V.R.; dos Santos, R.F. Evaluating landscape connectivity for Puma concolor and Panthera onca among Atlantic forest protected areas. Environ. Manage. 2015, 55, 1377–1389. [Google Scholar] [CrossRef]
- Atlas digital de Costa Rica 2014. E. Ortiz-Malavasi. Instituto Tecnológico de Costa Rica (ITCR), Laboratorio de Sistemas de Información Geográfica, Escuela de Ingeniería Forestal, ITCR. Cartago, CR. Available online: https://repositoriotec.tec.ac.cr/handle/2238/6749 (accessed on 28 March 2020).
- Hedström, I. Talamanca Indómita: Relato-guía de campo del Parque Nacional Barbilla, Costa Rica (Untamed Talamanca: Chronicle—Fieldguide to Barbilla Nacional Park, Costa Rica), 1st ed.; Fundación Nairi: San José, Costa Rica, 2006; p. 428. [Google Scholar]
- Food and Agriculture Organization. Evaluación de los recursos forestales mundiales 2010, Informe nacional Costa Rica. Roma, Italia. Available online: https://www.sirefor.go.cr/pdfs/publicaciones/2010_FAO_Informe_Evaluacion_Recursos_Forestales_Mundiales_Costa_Rica_FRA_2010.pdf (accessed on 28 February 2020).
- Ley Indígena 1977. Art 6, Law No. 6172. Available online: http://www.conai.go.cr/documentos.html (accessed on 27 February 2020).
- Huston, M.A. Biological diversity: The coexistence of species on changing landscapes; Cambridge University Press: Cambridge, UK; p. 681.
- Terborgh, J.; Lopez, L.; Nunez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecological meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef] [PubMed]
- Galetti, M.; Donatti, C.I.; Pires, A.S.; Guimarães, P.R.; Jordano, P. Seed survival and dispersal of an endemic Atlantic forest palm: The combined effects of defaunation and forest fragmentation. Bot. J. Linn. Soc. 2006, 151, 141–149. [Google Scholar] [CrossRef]
- Stoner, C.; Caro, T.; Mduma, S.; Mlingwa, C.; Sabuni, G.; Borner, M.; Schelten, C. Changes in large herbivore populations across large areas of Tanzania. Afr. J. Ecol. 2007, 45, 202–215. [Google Scholar] [CrossRef]
- Peres, C.A.; Palacios, E. Basin-wide effects of game harvest on vertebrate population densities in Amazonian forests: Implications for animal-mediated seed dispersal. Biotropica 2007, 39, 304–315. [Google Scholar] [CrossRef]
- Carrillo, E.; Wong, G.; Cuarón, A.D. Monitoring Mammal Populations in Costa Rican Protected Areas under Different Hunting Restrictions. Conserv. Biol. 2000, 14, 1580–1591. [Google Scholar] [CrossRef]
- Abrahams, M.I.; Peres, C.A.; Costa, H.C.M. Measuring local depletion of terrestrial game vertebrates by central-place hunters in rural Amazonia. PLoS ONE 2017, 12, 1–25. [Google Scholar] [CrossRef]
- Crooks, K.R.; Soulé, M.E. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 1999, 400, 463–466. [Google Scholar] [CrossRef]
- Sistema Nacional de Áreas de Conservación. Available online: http://www.sinac.go.cr/ES/buscador/Paginas/default.aspx (accesed on 12 October 2017).
- Bernal, L.; García, P. Viabilidad de realización de un proyecto de turismo rural comunitario en las comunidades indígenas de Nairi-Awari. Unpublished work, 2007. [Google Scholar]
- Sáenz-Bolaños, C.; Montalvo, V.; Fuller, T.K.; Carrillo, E. Records of black jaguars at Parque Nacional Barbilla. CatNews 2015, 62, 38–39. [Google Scholar]
- Instituto Nacional de Estadística y Censos [Costa Rica] (INEC). X Censo Nacional de Población y IV de vivienda: Territorios indígenas/Instituto Nacional de Estadística y Censos, 1st ed.; INEC: San José, Costa Rica, 2013; p. 56. [Google Scholar]
- Ministerio de Planificación Nacional y Política Económica (MIDEPLAN). Análisis de desarrollo: Población Indígena en Cifras. Unpublished work, 2015. [Google Scholar]
- Blake, J.G.; Mosquera, D. Camera trapping on and off trails in lowland forest of eastern Ecuador: Does location matter? Mastozool. Neotrop. 2014, 21, 17–26. [Google Scholar]
- Blake, J.G.; Mosquera, D.; Loiselle, B.A.; Romo, D.; Swing, K. Effects of human traffic on use of trails by mammals in lowland forest of eastern Ecuador. Neodiversity 2017, 3, 57–64. [Google Scholar] [CrossRef]
- Wainwright, M. The Mammals of Costa Rica: A Natural History field Guide; Cornell University Press: New York, NY, USA, 2007. [Google Scholar]
- Garrigues, R.; Dean, R. The Birds of Costa Rica: A Field Guide, 2nd ed.; Zona Tropical Cornell University Press: New York, NY, USA, 2014. [Google Scholar]
- O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. Camera-traps in Animal Ecology: Methods and Analyses, 1st ed.; O’Connell, A.F., Nichols, J.D., Karanth, K.U., Eds.; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Hedström, I. Preliminary check list of observed bird species within Tapir River Private Wild Life Reserve. Unpublished work, 2011. [Google Scholar]
- Alvarado, R.; Escobar, B.; Ramos, J.; Sagastume, V. Anfibios, aves y mamíferos del Parque Nacional Barbilla y una propuesta de indicadores para evaluar su integridad ecológica; Alvarado, R., Ed.; ICOMVIS-Universidad Nacional de Costa Rica: Heredia, Costa Rica, 2017. [Google Scholar]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Influences of Anthropogenic Resources on Raccoon (Procyon lotor) Movements and Spatial Distribution. J. Mammal. 2004, 85, 483–490. [Google Scholar] [CrossRef]
- Schulte-Hostedde, A.I.; Mazal, Z.; Jardine, C.M.; Gagnon, J. Enhanced access to anthropogenic food waste is related to hyperglycemia in raccoons (Procyon lotor). Conserv. Physiol. 2018, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hass, C.C.; Valenzuela, D. Anti-predator benefits of group living in white-nosed coatis (Nasua narica). Behav. Ecol. Sociobiol. 2002, 51, 570–578. [Google Scholar] [CrossRef]
- Borge, C.; Martínez, J. El Pago por Servicios Ambientales en Territorios Indígenas de Costa Rica. PES Learning Paper 2009-1S. (Spanish). Payments for Environmental Services (PES) learning paper. DC World Bank Group: Washington, DC, USA, 2009. Available online: http://documents.worldbank.org/curated/en/440201468261580966/El-pago-por-servicios-ambientales-en-territorios-ind-237-genas-de-Costa-Rica (accessed on 28 February 2020).
- Hunter, P. The human impact on biological diversity. How species adapt to urban challenges sheds light on evolution and provides clues about conservation. EMBO Rep. 2007, 8, 316–318. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).