Current State and Drivers of Arable Plant Diversity in Conventionally Managed Farmland in Northwest Germany

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
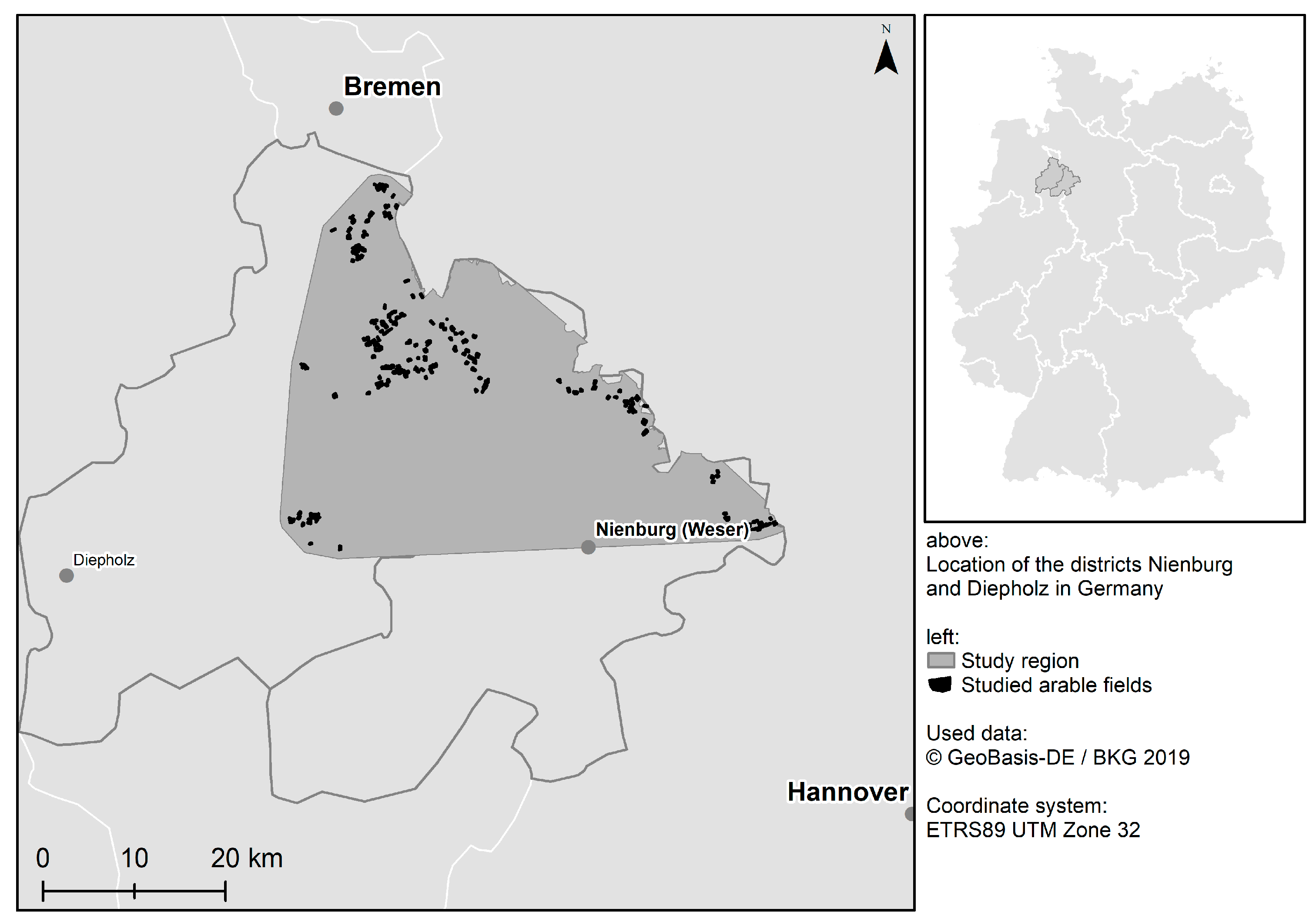
2.1. Study Region
2.2. Vegetation Survey
2.3. Abiotic and Management Data
2.4. Statistical Analyses
3. Results
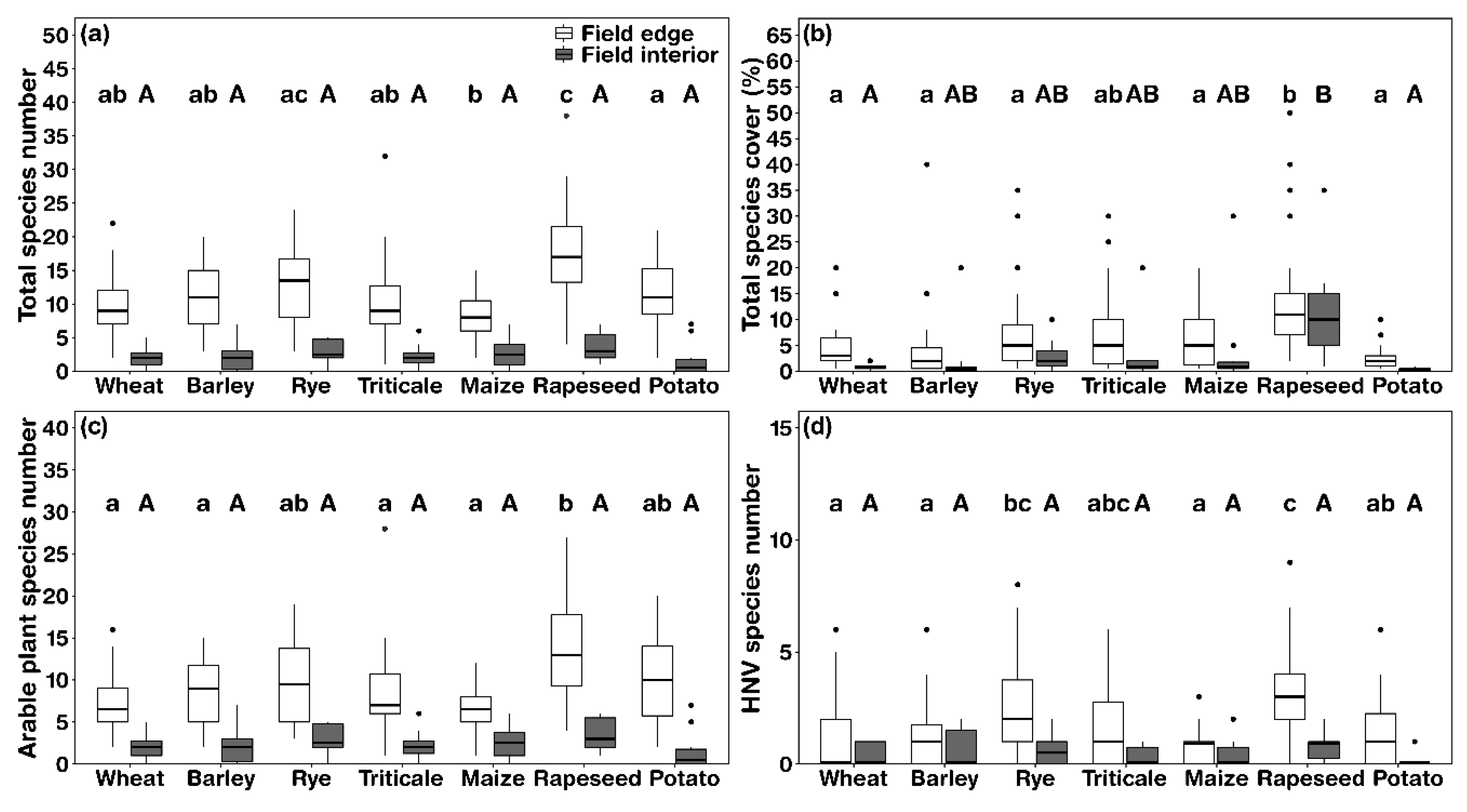
3.1. Patterns of Plant Species Richness and Cover
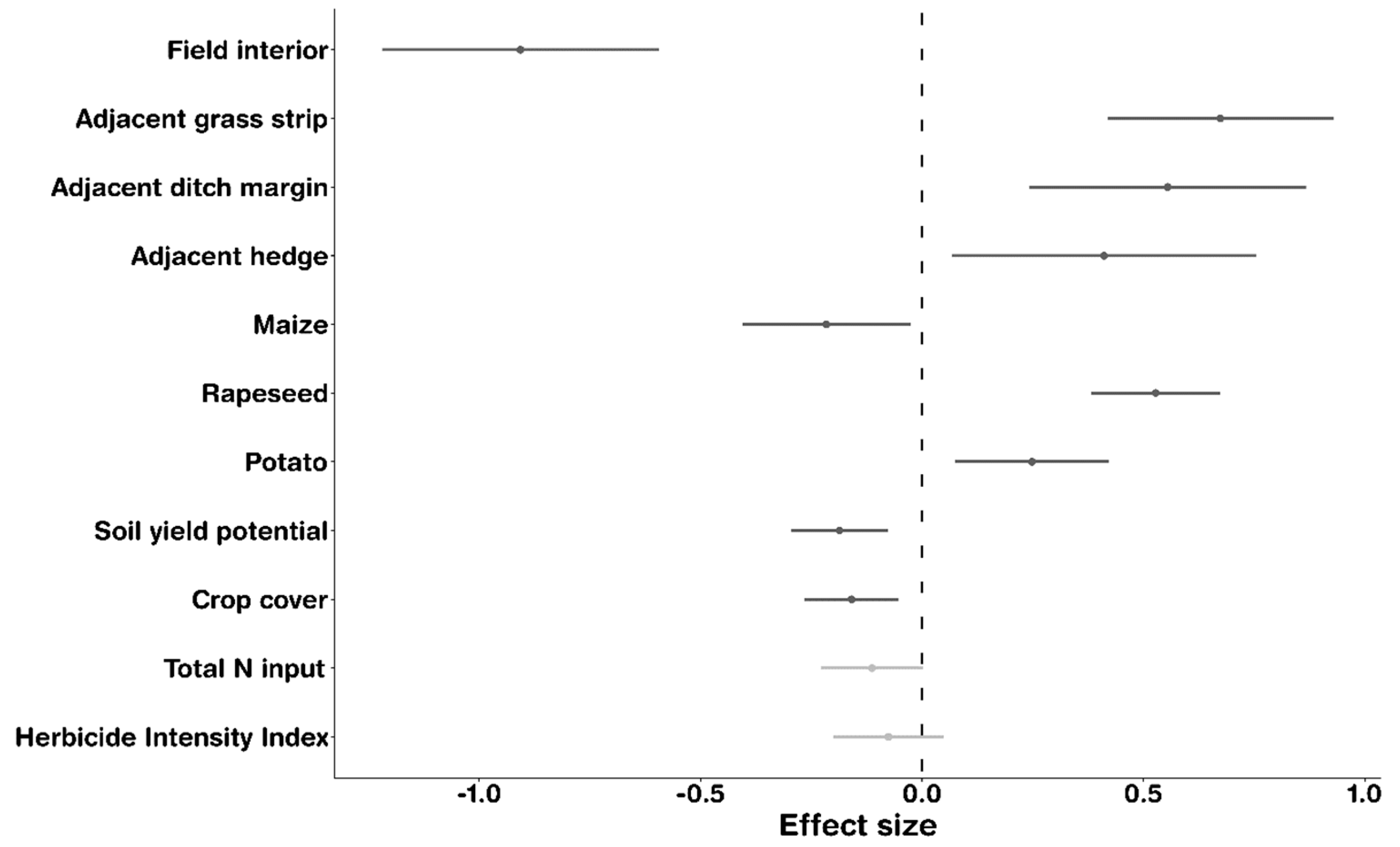
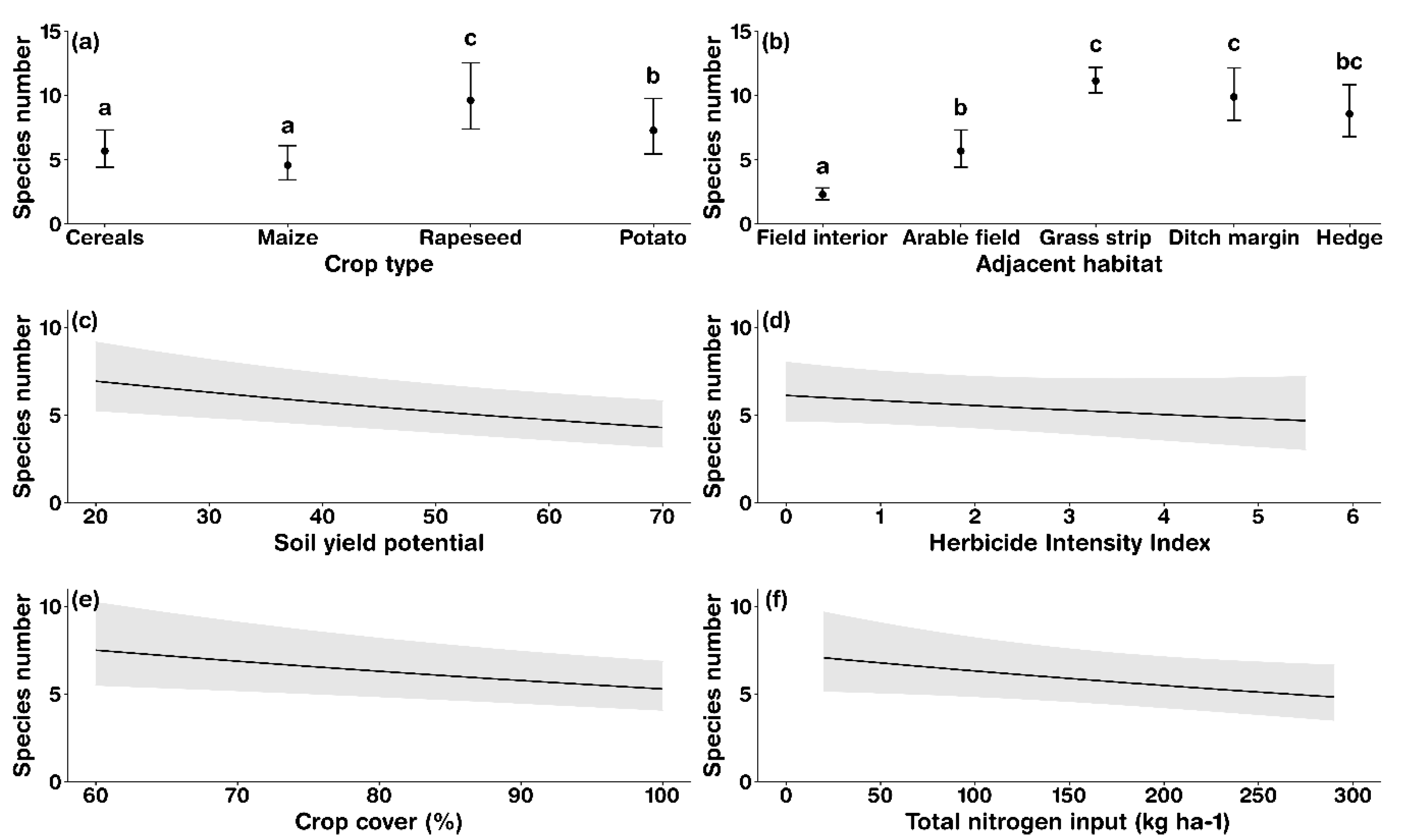
3.2. Factors Determining Arable Plant Species Richness
3.3. Variation in Community Composition
4. Discussion
4.1. Current State of Phytodiversity in Conventionally Managed Farmland
4.2. The Importance of the Field Edge
4.3. Composition of the Recent Arable Vegetation and the Role of Field Neighborhood
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Storkey, J.; Meyer, S.; Still, K.S.; Leuschner, C. The impact of agricultural intensification and land-use change on the European arable flora. Proc. R. Soc. B Biol. Sci. 2012, 279, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; van Bommel, F.P.J. Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric. Ecosyst. Environ. 2006, 116, 189–196. [Google Scholar] [CrossRef]
- Gabriel, D.; Sait, S.M.; Kunin, W.E.; Benton, T.G. Food production vs. biodiversity: Comparing organic and conventional agriculture. J. Appl. Ecol. 2013, 50, 355–364. [Google Scholar] [CrossRef]
- Meyer, S.; Wesche, K.; Krause, B.; Leuschner, C. Dramatic losses of specialist arable plants in Central Germany since the 1950s/60s—A cross-regional analysis. Divers. Distrib. 2013, 19, 1175–1187. [Google Scholar] [CrossRef]
- Cirujeda, A.; Aibar, J.; Zaragoza, C. Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007. Agron. Sustain. Dev. 2011, 31, 675–688. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Dessaint, F.; Reboud, X. Arable weed decline in Northern France: Crop edges as refugia for weed conservation? Biol. Conserv. 2009, 142, 238–243. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats: Vegetation Ecology of Central Europe, Volume II; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-43046-1. [Google Scholar]
- Gholamhoseini, M.; AghaAlikhani, M.; Mirlatifi, S.M.; Sanavy, S.A.M.M. Weeds—Friend or foe? Increasing forage yield and decreasing nitrate leaching on a corn forage farm infested by redroot pigweed. Agric. Ecosyst. Environ. 2013, 179, 151–162. [Google Scholar] [CrossRef]
- Hawes, C.; Haughton, A.J.; Osborne, J.L.; Roy, D.B.; Clark, S.J.; Perry, J.N.; Rothery, P.; Bohan, D.A.; Brooks, D.R.; Champion, G.T.; et al. Responses of plants and invertebrate trophic groups to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 1899–1913. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Duraiappah, A.K.; Naeem, S.; Agardy, T.; Ash, N.J.; Cooper, H.D.; Diaz, S.; Faith, D.P.; Mace, G.; McNeely, J.A.; Mooney, H.A.; et al. Millennium Ecosystem Assessment. In Ecosystems and Human Well-being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005; p. 100. [Google Scholar]
- Fried, G.; Norton, L.R.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Pinke, G.; Karácsony, P.; Czúcz, B.; Botta-Dukát, Z.; Lengyel, A. The influence of environment, management and site context on species composition of summer arable weed vegetation in Hungary. Appl. Veg. Sci. 2012, 15, 136–144. [Google Scholar] [CrossRef]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot. Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef]
- Seifert, C.; Leuschner, C.; Meyer, S.; Culmsee, H. Inter-relationships between crop type, management intensity and light transmissivity in annual crop systems and their effect on farmland plant diversity. Agric. Ecosyst. Environ. 2014, 195, 173–182. [Google Scholar] [CrossRef]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- Maréchal, P.-Y.; Henriet, F.; Vancutsem, F.; Bodson, B. Ecological review of black-grass (Alopecurus myosuroides Huds.) propagation abilities in relationship with herbicide resistance. Biotechnol. Agron. Soc. Environ. 2012, 16, 103–113. [Google Scholar]
- Egan, J.F.; Graham, I.M.; Mortensen, D.A. A comparison of the herbicide tolerances of rare and common plants in an agricultural landscape: Herbicide tolerances of rare and common plants. Environ. Toxicol. Chem. 2014, 33, 696–702. [Google Scholar] [CrossRef]
- Meyer, S.; Bergmeier, E.; Becker, T.; Wesche, K.; Krause, B.; Leuschner, C. Detecting long-term losses at the plant community level—Arable fields in Germany revisited. Appl. Veg. Sci. 2015, 18, 432–442. [Google Scholar] [CrossRef]
- Hofmeister, H.; Garve, E. Lebensraum Acker; Reprint der 2. neubearbeiteten Auflage; Verlag Kessel: Remagen, Germany, 2006; ISBN 978-3-935638-61-6. [Google Scholar]
- Hüppe, J.; Hofmeister, H. Syntaxonomische Fassung und Übersicht über die Ackerunkrautgesellschaften der Bundesrepublik Deutschland. Ber. Reinh.-Tüxen-Ges. 1990, 2, 57–77. [Google Scholar]
- Kaussmann, B.; Kudocke, J. Die ökologisch-soziologischen Artengruppen der Ackerunkrautvegetation für den Norden der DDR. Feddes Repert. 1973, 84, 589–605. [Google Scholar] [CrossRef]
- Statistical Office of the European Union (EUROSTAT) Main Farmland Use by NUTS 2 Regions & Organic Crop Area by Agricultural Production Methods and Crops. Available online: https://ec.europa.eu/eurostat/web/agriculture/data/database (accessed on 24 October 2018).
- Hanzlik, K.; Gerowitt, B. Methods to conduct and analyse weed surveys in arable farming: A review. Agron. Sustain. Dev. 2016, 36. [Google Scholar] [CrossRef]
- Batáry, P.; Gallé, R.; Riesch, F.; Fischer, C.; Dormann, C.F.; Mußhoff, O.; Császár, P.; Fusaro, S.; Gayer, C.; Happe, A.-K.; et al. The former Iron Curtain still drives biodiversity–profit trade-offs in German agriculture. Nat. Ecol. Evol. 2017, 1, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- von Drachenfels, O. Überarbeitung der Naturräumlichen Regionen Niedersachsens. Inform. Nat. Niedersachs. 2010, 30, 249–252. [Google Scholar]
- German Meteorological Service (DWD) Climate Data (2013–2017) via the CDC FTP Server. Available online: https://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/annual/kl/recent/ & https://www.dwd.de/DE/leistungen/klimadatendeutschland/statliste/statlex_html.html?view=nasPublication&nn=16102 (accessed on 25 October 2018).
- Federal Institute for Geosciences and Natural Resources Soil Map of the Federal Republic of Germany 1:1,000,000 (BÜK1000). Available online: https://www.bgr.bund.de/DE/Themen/Boden/Informationsgrundlagen/Bodenkundliche_Karten_Datenbanken/BUEK1000/buek1000_node.html (accessed on 26 October 2016).
- State Authority for Mining, Energy and Geology (LBEG) German Soil Assessment Map 1:5000 (BS5). Available online: https://www.lbeg.niedersachsen.de/karten_daten_publikationen/karten_daten/boden/bodenkarten/bodenschaetzungskarte_15000/bodenschaetzungskarte-von-niedersachsen-im-mastab-1--5-000-bs5-681.html (accessed on 26 October 2018).
- Wietzke, A.; Leuschner, C. Surveying the arable plant diversity of conventionally managed farmland: A comparison of methods. Environ. Monit. Assess. 2020, 192, 98. [Google Scholar] [CrossRef]
- Buttler, K.P. Florenliste von Deutschland—Gefäßpflanzen. Available online: http://www.kp-buttler.de (accessed on 13 February 2019).
- Environmental Systems Research Institute (ESRI) ArcGIS Desktop 10.6.1. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview (accessed on 26 October 2018).
- Roßberg, D.; Gutsche, V.; Enzian, S.; Wick, M. NEPTUN 2000—Erhebung von Daten zum Tatsächlichen Einsatz Chemischer Pflanzenschutzmittel im Ackerbau Deutschlands; Berichte aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft, Heft 98; SaphirVerlag: Ribbesbüttel, Germany, 2002. [Google Scholar]
- Land24 GmbH Crop Protection Manger. Available online: https://www.raiffeisen.com/pflanzenschutzmittel (accessed on 26 October 2018).
- Garve, E. Rote Liste und Florenliste der Farn- und Blütenpflanzen in Niedersachsen und Bremen, 5. Fassung vom 1.3.2004. Inform. Nat. Niedersachs. 2004, 24, 1–76. [Google Scholar]
- German Federal Agency for Nature Conservation High-Nature-Value Farmland, Germany. Available online: https://www.bfn.de/themen/monitoring/monitoring-von-landwirtschaftsflaechen-mit-hohem-naturwert.html (accessed on 19 October 2018).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 6 November 2018).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R. J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Chen, H. VennDiagram: Generate High-Resolution Venn and Euler Plots. R Package Version 1.6.20. Available online: https://CRAN.R-project.org/package=VennDiagram (accessed on 7 November 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-2. Available online: https://CRAN.R-project.org/package=vegan (accessed on 6 November 2018).
- Gelman, A. Scaling regression inputs by dividing by two standard deviations. Stat. Med. 2008, 27, 2865–2873. [Google Scholar] [CrossRef]
- Chamorro, L.; Masalles, R.M.; Sans, F.X. Arable weed decline in Northeast Spain: Does organic farming recover functional biodiversity? Agric. Ecosyst. Environ. 2016, 223, 1–9. [Google Scholar] [CrossRef]
- Kolářová, M.; Tyšer, L.; Soukup, J. Diversity of current weed vegetation on arable land in selected areas of the Czech Republic. Plant Soil Environ. 2013, 59, 208–213. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Batáry, P.; Báldi, A.; Harnos, A. Interaction of local and landscape features in the conservation of Hungarian arable weed diversity: Weed richness in Hungarian cereal fields. Appl. Veg. Sci. 2011, 14, 40–48. [Google Scholar] [CrossRef]
- Meyer, S.; Wesche, K.; Krause, B.; Brütting, C.; Hensen, I.; Leuschner, C. Diversitätsverluste und floristischer Wandel im Ackerland seit 1950. Nat. Landsch. 2014, 89, 392–398. [Google Scholar]
- Meisel, K.; von Hübschmann, A. Veränderungen der Acker- und Grünlandvegetation im nordwestdeutschen Flachland in jüngerer Zeit. Schr. Veg. 1976, 10, 109–124. [Google Scholar]
- Preising, E.; Vahle, H.-C.; Brandes, D.; Hofmeister, H.; Tüxen, J.; Weber, H.E. Die Pflanzengesellschaften Niedersachsens—Bestandsentwicklung, Gefährdung und Schutzprobleme. Einjährige ruderale Pionier-, Tritt- und Ackerwildkraut- Gesellschaften. Nat. Landsch. Niedersachs. 1995, 20, 1–92. [Google Scholar]
- Leuschner, C.; Krause, B.; Meyer, S.; Bartels, M. Structural change in the arable land and grassland of Lower Saxony and Schleswig-Holstein since 1950. Nat. Landsch. 2014, 89, 386–391. [Google Scholar]
- Meisel, K. Über die Artenverbindung des Aphanion arvensis J. et R. Tx. 1960 im west-und nordwestdeutschen Flachland. Schr. Veg. 1967, 2, 123–133. [Google Scholar]
- Aavik, T.; Augenstein, I.; Bailey, D.; Herzog, F.; Zobel, M.; Liira, J. What is the role of local landscape structure in the vegetation composition of field boundaries? Appl. Veg. Sci. 2008, 11, 375–386. [Google Scholar] [CrossRef]
- Vickery, J.A.; Feber, R.E.; Fuller, R.J. Arable field margins managed for biodiversity conservation: A review of food resource provision for farmland birds. Agric. Ecosyst. Environ. 2009, 133, 1–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
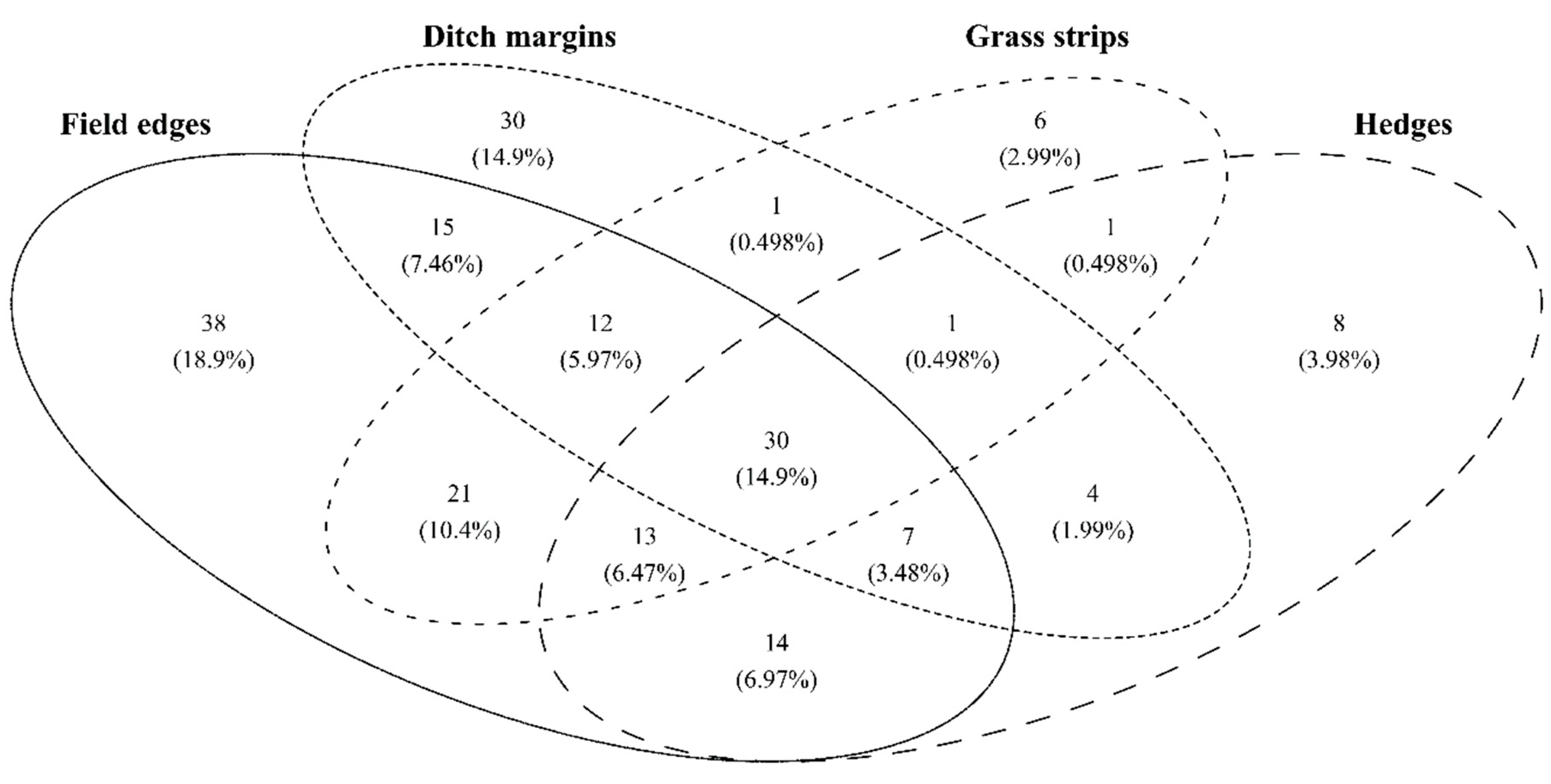{kind=link}
| Plot Type | Crop | Arable Plants | Class | Order | Alliance | Association | Total Number of Diagnostic Species |
|---|---|---|---|---|---|---|---|
| Field edge | Wheat | 6.5 | 0 | 0 | 0 | 1 | 1 |
| Field interior | Wheat | 2 | 0 | 0 | 0 | 0 | 1 |
| Field edge | Barley | 9 | 1 | 0 | 1 | 1 | 2.5 |
| Field interior | Barley | 2 | 0 | 0 | 0 | 0 | 1 |
| Field edge | Rye | 9.5 | 1 | 0 | 2 | 1 | 4 |
| Field interior | Rye | 2.5 | 1 | 0 | 0 | 0 | 1 |
| Field edge | Triticale | 7 | 0 | 0 | 0.5 | 1 | 2 |
| Field interior | Triticale | 2 | 0 | 0 | 0 | 0 | 0 |
| Field edge | Maize | 6.5 | 1 | 0 | 0 | 1 | 2 |
| Field interior | Maize | 2.5 | 0 | 0 | 0 | 1 | 1.5 |
| Field edge | Rapeseed | 13 | 2 | 0 | 1 | 2 | 5 |
| Field interior | Rapeseed | 3 | 1 | 0 | 0 | 1 | 2 |
| Field edge | Potato | 10 | 2 | 0 | 0.5 | 1 | 4 |
| Field interior | Potato | 0.5 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wietzke, A.; van Waveren, C.-S.; Bergmeier, E.; Meyer, S.; Leuschner, C. Current State and Drivers of Arable Plant Diversity in Conventionally Managed Farmland in Northwest Germany. Diversity 2020, 12, 469. https://doi.org/10.3390/d12120469
Wietzke A, van Waveren C-S, Bergmeier E, Meyer S, Leuschner C. Current State and Drivers of Arable Plant Diversity in Conventionally Managed Farmland in Northwest Germany. Diversity. 2020; 12(12):469. https://doi.org/10.3390/d12120469
Chicago/Turabian StyleWietzke, Alexander, Clara-Sophie van Waveren, Erwin Bergmeier, Stefan Meyer, and Christoph Leuschner. 2020. "Current State and Drivers of Arable Plant Diversity in Conventionally Managed Farmland in Northwest Germany" Diversity 12, no. 12: 469. https://doi.org/10.3390/d12120469
APA StyleWietzke, A., van Waveren, C.-S., Bergmeier, E., Meyer, S., & Leuschner, C. (2020). Current State and Drivers of Arable Plant Diversity in Conventionally Managed Farmland in Northwest Germany. Diversity, 12(12), 469. https://doi.org/10.3390/d12120469