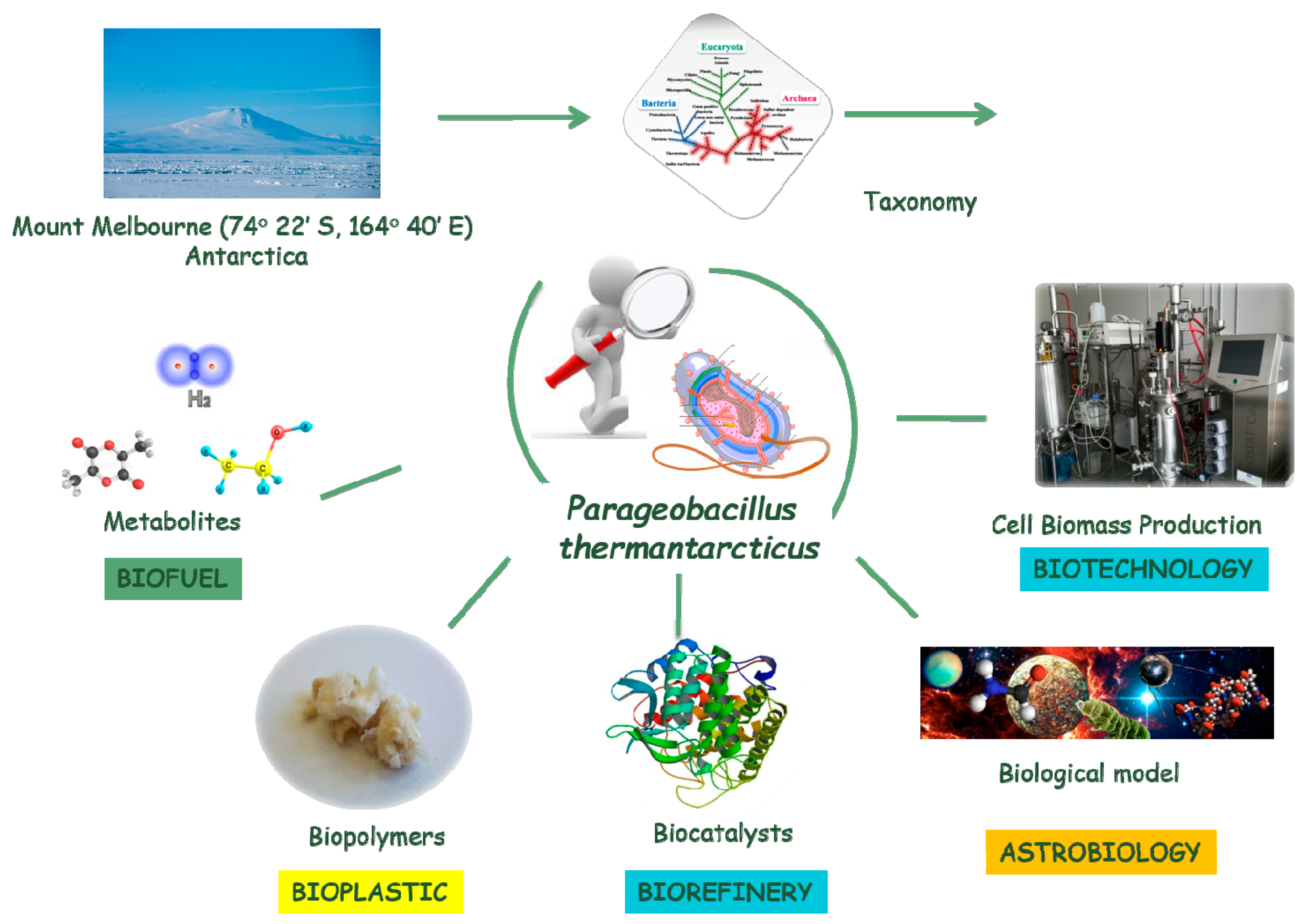
Parageobacillus thermantarcticus, an Antarctic Cell Factory: From Crop Residue Valorization by Green Chemistry to Astrobiology Studies

 ,
, 


Abstract
1. Introduction
2. Phylogenomic Re-Assessment of Bacillus thermoantarcticus to Parageobacillus thermantarcticus
3. Main Enzymes in P. thermantarcticus
3.1. Xylanase and β-Xylosidase
3.2. Xylose (Glucose) Isomerase
3.3. Protease(s)
4. Production of Xylo-Oligosaccharides as High Value-Added Molecules
5. Relevance of P. thermantarcticus to Astrobiology
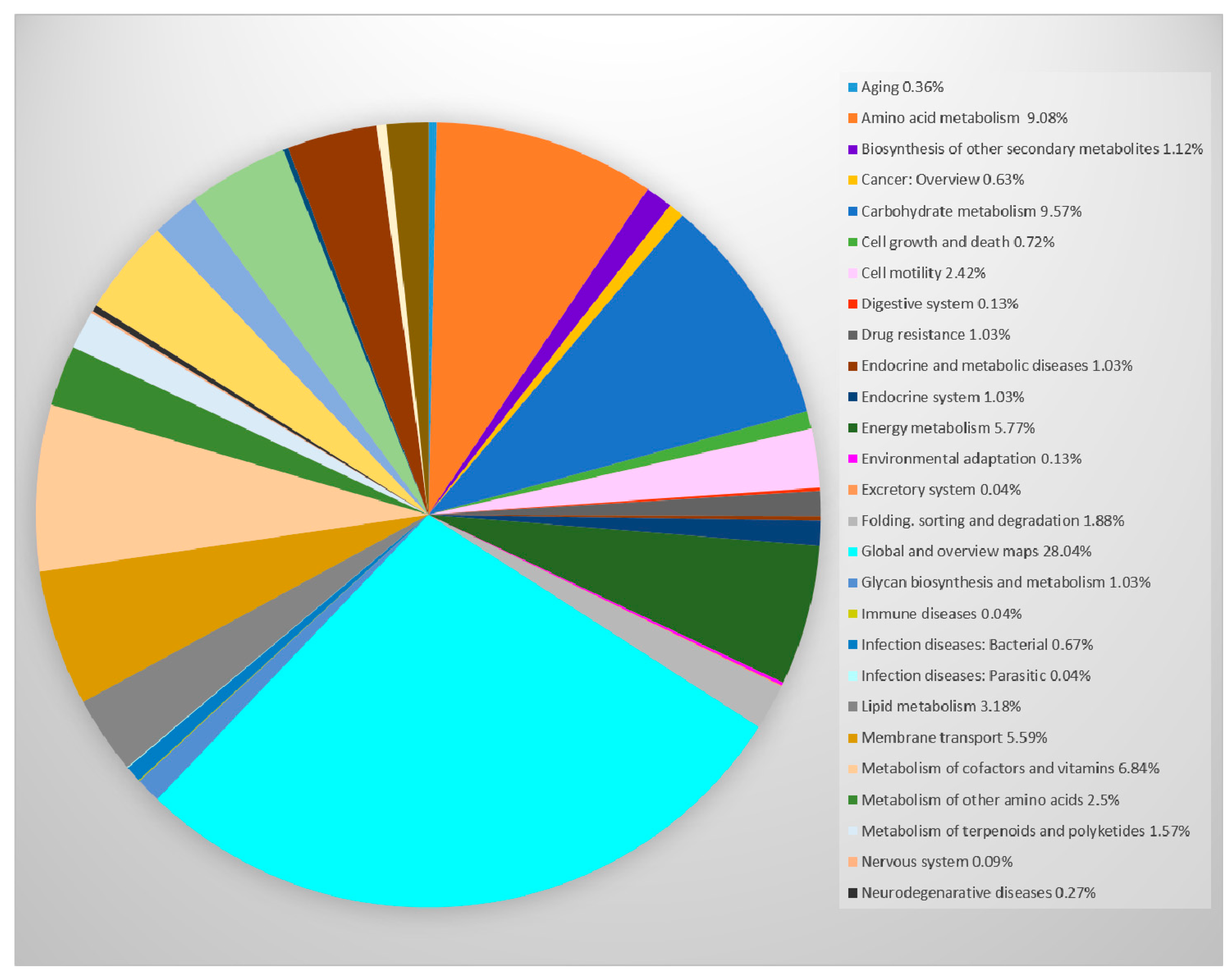
6. Genome of P. thermantarcticus
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kambourova, M.; Radchenkova, N.; Tomova, I.; Bojadjieva, I. Thermophiles as a Promising Source of Exopolysaccharides with Interesting Properties. In Biotechnology of Extremophiles; Rampelotto, P.H., Ed.; Grand Challenges in Biology and Biotechnology; Springer International Publishing: Cham, Switzerland, 2016; pp. 117–139. [Google Scholar]
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial enzyme applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Van Beilen, J.B.; Li, Z. Enzyme technology: An overview. Curr. Opin. Biotechnol. 2002, 13, 338–344. [Google Scholar] [CrossRef]
- Zeigler, D.R. The Geobacillus paradox: Why is a thermophilic bacterial genus so prevalent on a mesophilic planet? Microbiology 2014, 160, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.A.; Flores, P.A.; Boehmwald, F.A.; Blamey, J.M. Thermophilic bacteria present in a sample from Fumarole Bay, Deception Island. Antarct. Sci. 2011, 23, 549–555. [Google Scholar] [CrossRef]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef]
- Nicolaus, B.; Lama, L.; Gambacorta, A. Thermophilic bacillus isolated from Antarctic environments. In Thermophilic Bacillus Isolates from Antarctic Environments Applications and systematics of Bacillus and Relatives; Berkeley, R., Heyndrickx, M., Logan, M., de Vos, P., Eds.; Blackwell: Oxford, UK, 2002; pp. 47–63. [Google Scholar]
- Nicolaus, B.; Lama, L.; Esposito, E.; Manca, M.C.; di Prisco, G.; Gambacorta, A. ‘‘Bacillus thermoantarcticus’’ sp. nov., from Mount Melbourne, Antarctica: A novel thermophilic species. Polar Biol. 1996, 16, 101–104. [Google Scholar]
- Lama, L.; Tramice, A.; Finore, I.; Anzelmo, G.; Calandrelli, V.; Pagnotta, E.; Tommonaro, G.; Poli, A.; Di Donato, P.; Nicolaus, B.; et al. Degradative actions of microbial xylanolytic activities on hemicelluloses from rhizome of Arundo donax. AMB Express 2014, 4, 55–64. [Google Scholar] [CrossRef]
- Finore, I.; Poli, A.; Di Donato, P.; Lama, L.; Trincone, A.; Fagnano, M.; Mori, M.; Nicolaus, B.; Tramice, A. The hemicellulose extract from Cynara cardunculus: A source of value-added biomolecules produced by xylanolytic thermozymes. Green Chem. 2016, 18, 2460–2472. [Google Scholar] [CrossRef]
- Manca, M.C.; Lama, L.; Improta, R.; Esposito, E.; Gambacorta, A.; Nicolaus, B. Chemical Composition of Two Exopolysaccharides from Bacillus thermoantarcticus. Appl. Environ. Microbiol. 1996, 62, 3265–3269. [Google Scholar]
- Mastascusa, V.; Romano, I.; Di Donato, P.; Poli, A.; Della Corte, V.; Rotundi, A.; Bussoletti, E.; Quarto, M.; Pugliese, M.; Nicolaus, B. Extremophiles Survival to Simulated Space Conditions: An Astrobiology Model Study. Orig. Life Evol. Biosph. 2014, 44, 231–237. [Google Scholar] [CrossRef]
- Di Donato, P.; Romano, I.; Mastascusa, V.; Poli, A.; Orlando, P.; Pugliese, M.; Nicolaus, B. Survival and Adaptation of the Thermophilic Species Geobacillus thermantarcticus in Simulated Spatial Conditions. Orig. Life Evol. Biosph. 2018, 48, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; De Angelis, A.; Poli, A.; Ragni, P.; Lilla, L.; Zito, G.; Nicolaus, B.; De Luca, A.C.; Di Donato, P. Resistance and Raman spectroscopy analysis of Parageobacillus thermantarcticus spores after γ-ray exposure. Extremophiles 2018, 22, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Villada, J.C.; Lee, P.K.H. Modular Metabolic Engineering for Biobased Chemical Production. Trends Biotechnol. 2019, 37, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, B.; Lama, L.; Esposito, E.; Manca, M.C.; di Prisco, G.; Gambacorta, A. Validation of publication of new names and new combinations previously effectively published outside the IJSEM. Int. J. Syst. Evol. Microbiol. 2005, 55, 1743–1745. [Google Scholar]
- Chen, X.G.; Stabnikova, O.; Tay, J.H.; Wang, J.Y.; Tay, S.T.L. Thermoactive extracellular proteases of Geobacillus caldoproteolyticus, sp. nov., from sewage sludge. Extremophiles 2004, 8, 489–498. [Google Scholar] [CrossRef]
- Ash, C.; Farrow, J.A.E.; Wallbanks, S.; Collins, M.D. Phylogenetic heterogeneity of the genus Bacillus revealed by comparative analysis of small-subunit-ribosomal RNA sequences. Lett. Appl. Microbiol. 1991, 13, 202–206. [Google Scholar] [CrossRef]
- Rainey, F.A.; Fritze, D.; Stackebrandt, E. The phylogenetic diversity of thermophilic members of the genus Bacillus as revealed by 16S rDNA analysis. FEMS Microbiol. Lett. 1994, 115, 205–211. [Google Scholar] [CrossRef]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic study of aerobic thermophilic bacilli: descriptions of Geobacillus subterraneus gen. nov., sp. nov. and Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. th. Int. J. Syst. Evol. Microbiol. 2001, 51, 433–446. [Google Scholar] [CrossRef]
- Scholz, T.; Demharter, W.; Hensel, R.; Kandler, O. Bacillus pallidus sp. nov., a new thermophilic species from sewage. Syst. Appl. Microbiol. 1987, 9, 91–96. [Google Scholar] [CrossRef]
- Ahmad, S.; Scopes, R.K.; Rees, G.N.; Patel, B.K.C. Saccharococcus caldoxylosilyticus sp. nov., an obligately thermophilic, xylose-utilizing, endospore-forming bacterium. Int. J. Syst. Evol. Microbiol. 2000, 50, 517–523. [Google Scholar] [CrossRef]
- Caccamo, D.; Gugliandolo, C.; Stackebrandt, E.; Maugeri, T.L. Bacillus vulcani sp. nov., a novel thermophilic species isolated from a shallow marine hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2000, 50, 2009–2012. [Google Scholar] [CrossRef] [PubMed]
- Fortina, M.G.; Mora, D.; Schumann, P.; Parini, C.; Manachini, P.L.; Stackebrandt, E. Reclassification of Saccharococcus caldoxylosilyticus as Geobacillus caldoxylosilyticus (Ahmad et al. 2000) comb. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 2063–2071. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Banat, I.M.; Marchant, R.; Rahman, T.J. Geobacillus debilis sp. nov., a novel obligately thermophilic bacterium isolated from a cool soil environment, and reassignment of Bacillus pallidus to Geobacillus pallidus comb. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 2197–2201. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Lebedeva, E.V.; Poltaraus, A.B.; Tourova, T.P.; Grigoryan, A.A.; Sokolova, D.S.; Lysenko, A.M.; Osipov, G.A. Geobacillus gargensis sp. nov., a novel thermophile from a hot spring, and the reclassification of Bacillus vulcani as Geobacillus vulcani comb. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Miñana-Galbis, D.; Pinzón, D.L.; Lorén, J.G.; Manresa, À.; Oliart-Ros, R.M. Reclassification of Geobacillus pallidus (Scholz et al. 1988) Banat et al. 2004 as Aeribacillus pallidus gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 1600–1604. [Google Scholar] [CrossRef]
- Sung, M.H.; Kim, H.; Bae, J.W.; Rhee, S.K.; Jeon, C.O.; Kim, K.; Kim, J.J.; Hong, S.P.; Lee, S.G.; Yoon, J.H.; et al. Geobacillus toebii sp. nov., a novel thermophilic bacterium isolated from hay compost. Int. J. Syst. Evol. Microbiol. 2002, 52, 2251–2255. [Google Scholar] [CrossRef] [PubMed]
- Kuisiene, N.; Raugalas, J.; Chitavichius, D. Geobacillus lituanicus sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Schäffer, C.; Franck, W.L.; Scheberl, A.; Kosma, P.; McDermott, T.R.; Messner, P. Classification of isolates from locations in Austria and Yellowstone National Park as Geobacillus tepidamans sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 2361–2368. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Sokolova, D.S.; Grigoryan, A.A.; Shestakova, N.M.; Mikhailova, E.M.; Poltaraus, A.B.; Tourova, T.P.; Lysenko, A.M.; Osipov, G.A.; Belyaev, S.S. Geobacillus jurassicus sp. nov., a new thermophilic bacterium isolated from a high-temperature petroleum reservoir, and the validation of the Geobacillus species. Syst. Appl. Microbiol. 2005, 28, 43–53. [Google Scholar] [CrossRef]
- Coorevits, A.; Dinsdale, A.E.; Halket, G.; Lebbe, L.; de Vos, P.; Van Landschoot, A.; Logan, N.A. Taxonomic revision of the genus Geobacillus: Emendation of Geobacillus, G. stearothermophilus, G. jurassicus, G. toebii, G. thermodenitrificans and G. thermoglucosidans (nom. corrig., formerly ‘thermoglucosidasius’); transfer of Bacillus thermantarcticus to the genus as G. thermantarcticus comb. nov.; proposal of Caldibacillus debilis gen. nov., comb. nov.; transfer of G. tepidamans to Anoxybacillus as A. tepidamans comb. nov.; and proposal of Anoxybacillus caldiproteolyticus sp. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1470–1485. [Google Scholar] [CrossRef]
- Claus, D.; Berkeley, R.C.W. Genus Bacillus Cohn 1872. In Bergey’s Manual of Systematic Bacteriology; Sneath, P.H.A., Mair, N.S., Sharpe, M.E., Holt, J.G., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1986; Volume 2, pp. 1105–1140. [Google Scholar]
- Zeigler, D.R. Application of a recN sequence similarity analysis to the identification of species within the bacterial genus Geobacillus. Int. J. Syst. Evol. Microbiol. 2005, 5, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic re-assessment of the thermophilic genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of lignocellulosic biomass: Biochemical and molecular perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Jain, I.; Kumar, V.; Satyanarayana, T. Applicability of recombinant beta-xylosidase from the extremely thermophilic bacterium Geobacillus thermodenitrificans in synthesizing alkylxylosides. Bioresour. Technol. 2014, 170, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Subramaniyan, S.; Prema, P. Cellulase-free xylanases from Bacillus and other microorganisms. FEMS Microbiol. Lett. 2000, 183, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Xue, Y.; Wu, A.; Kataeva, I.; Pei, J.; Wu., H.; Wiegel, J. Characterization of a Novel beta-xylosidase, XylC, from Thermoanaerobacterium saccharolyticum JW/SL-YS485. Appl. Environ. Microbiol. 2011, 77, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Kumar, V.; Satyanarayana, T. Characteristics of thermostable endoxylanase and β-xylosidase of the extremely thermophilic bacterium Geobacillus thermodenitrificans TSAA1 and its applicability in generating xylooligosaccharides and xylose from agro-residues. Extremophiles 2013, 17, 357–366. [Google Scholar] [CrossRef]
- Shi, H.; Li, X.; Gu, H.; Zhang, Y.; Huang, Y.M.; Wang, L.; Wang, F. Biochemical properties of a novel thermostable and highly xylose-tolerant β-xylosidase/α-arabinosidase from Thermotoga thermarum. Biotechnol. Biofuels 2013, 6, 27. [Google Scholar] [CrossRef]
- Khasin, A.; Alchanati, I.; Shoham, Y. Purification and characterization of a thermostable xylanase from Bacillus stearothermophilus T-6. Appl. Environ. Microbiol. 1993, 59, 1725–1730. [Google Scholar]
- Beg, Q.; Kapoor, M.; Mahajan, L.; Hoondal, G.S. Microbial xylanases and their industrial applications: A review. Appl. Microbiol. Biotechnol. 2001, 56, 326–338. [Google Scholar] [CrossRef]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- FitzPatrick, M.; Champagne, P.; Cunningham, M.F.; Whitney, R.A. A biorefinery processing perspective: Treatment of lignocellulosic materials for the production of value-added products. Bioresour. Technol. 2010, 101, 8915–8922. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Xie, C.Y. Assessment of fibrolytic activities of 18 commercial enzyme products and their abilities to degrade the cell wall fraction of corn stalks in in vitro enzymatic and ruminal batch cultures. Anim. Feed Sci. Technol. 2010, 159, 110–121. [Google Scholar] [CrossRef]
- Chadha, B.S.; Kaur, B.; Basotra, N.; Tsang, A.; Pandey, A. Thermostable xylanases from thermophilic fungi and bacteria: Current perspective. Bioresour. Technol. 2019, 277, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Daas, M.J.A.; Murciano Martínez, P.; van de Weijer, A.H.P.; van der Oost, J.; de Vos, W.M.; Kabel, M.A.; van Kranenburg, R. Biochemical characterization of the xylan hydrolysis profile of the extracellular endoxylanase from Geobacillus thermodenitrificans T1. BMC Biotechnol. 2017, 17, 44. [Google Scholar] [CrossRef]
- Bhalla, A.; Bischoff, K.M.; Uppugundla, N.; Balan, V.; Sani, R.K. Novel thermostable endo-xylanase cloned and expressed from bacterium Geobacillus sp. WSUCF1. Bioresour. Technol. 2014, 165, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Bibra, M.; Kunreddy, V.; Sani, R. Thermostable Xylanase Production by Geobacillus sp. strain DUSELR13, and Its Application in Ethanol Production with Lignocellulosic Biomass. Microorganisms 2018, 6, 93. [Google Scholar] [CrossRef]
- Marcolongo, L.; La Cara, F.; Morana, A.; Di Salle, A.; del Monaco, G.; Paixão, S.M.; Alves, L.; Ionata, E. Properties of an alkali-thermo stable xylanase from Geobacillus thermodenitrificans A333 and applicability in xylooligosaccharides generation. World J. Microbiol. Biotechnol. 2015, 31, 633–648. [Google Scholar] [CrossRef]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Ratnadewi, A.A.; Fanani, M.; Kurniasih, S.D.; Sakka, M.; Wasito, E.B.; Sakka, K.; Nurachman, Z.; Puspaningsih, N.N. β-D-Xylosidase from Geobacillus thermoleovorans IT-08: Biochemical Characterization and Bioinformatics of the Enzyme. J. Ind. Microbiol. Biotechnol. 2014, 41, 65–74. [Google Scholar] [CrossRef]
- Sari, B.; Faiz, O.; Genc, B.; Sisecioglu, M.; Adiguzel, A.; Adiguzel, G. New xylanolytic enzyme from Geobacillus galactosidasius BS61 from a geothermal resource in Turkey. Int. J. Biol. Macromol. 2018, 119, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Lama, L.; Calandrelli, V.; Gambacorta, A.; Nicolaus, B. Purification and characterization of thermostable xylanase and β-xylosidase by the thermophilic bacterium Bacillus thermantarcticus. Res. Microbiol. 2004, 155, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.W.F.; dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef]
- Watanabe, T.; Suzuki, K.; Sato, I.; Morita, T.; Koike, H.; Shinozaki, Y.; Ueda, H.; Koitabashi, M.; Kitamoto, H.K. Simultaneous bioethanol distillery wastewater treatment and xylanase production by the phyllosphere yeast Pseudozyma antarctica GB-4(0). AMB Express 2015, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Finore, I.; Orlando, P.; Di Donato, P.; Leone, L.; Nicolaus, B.; Poli, A. Nesterenkonia aurantiaca sp. nov., an alkaliphilic actinobacterium isolated from Antarctica. Int. J. Syst. Evol. Microbiol. 2016, 66, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.; Salas, M.; Nwosu, V.C. High fructose corn syrup: Production, uses and public health concerns. Biotechnol. Molecul. Biol. Rev. 2010, 5, 71–78. [Google Scholar]
- Lee, S.M.; Jellison, T.; Alper, H.S. Directed Evolution of Xylose Isomerase for Improved Xylose Catabolism and Fermentation in the Yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 5708–5716. [Google Scholar] [CrossRef] [PubMed]
- Lama, L.; Nicolaus, B.; Calandrelli, V.; Romano, I.; Basile, R.; Gambacorta, A. Purification and characterization of thermostable xylose(glucose) isomerase from Bacillus thermoantarcticus. J. Ind. Microbiol. Biotechnol. 2001, 27, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P. Enzymes in Food Processing: A Condensed Overview on Strategies for Better Biocatalysts. Enzym. Res. 2010, 2010, 862537. [Google Scholar] [CrossRef]
- Bhosale, S.H.; Rao, M.B.; Deshpande, V.V. Molecular and Industrial Aspects of Glucose Isomerase. Microbiol. Rev. 1996, 60, 280–300. [Google Scholar]
- Lama, L.; Nicolaus, B.; Calandrelli, V.; Esposito, E.; Gambacorta, A. Xylanase produced by Bacillus thermoantarcticus, a new thermophilic bacillus. Ann. N. Y. Acad. Sci. Pap. 1996, 799, 284–289. [Google Scholar] [CrossRef]
- Brown, S.H.; Sjoholm, C.; Kelly, R.M. Purification and characterization of a highly thermostable glucose isomerase produced by the extremely thermophilic eubacterium, Thermotoga maritima. Biotechnol. Bioeng. 1993, 41, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Dekker, K.; Sugiura, A.; Yamagata, H.; Sakaguchi, K.; Udaka, S. Efficient production of thermostable Thermus thermophilus xylose isomerase in Escherichia coli and Bacillus brevis. Appl. Microbiol. Biotechnol. 1992, 36, 727–732. [Google Scholar] [CrossRef]
- Hess, J.M.; Tchernajenko, V.; Vieille, C.; Zeikus, J.G.; Kelly, R.M. Thermotoga neapolitana Homotetrameric Xylose Isomerase Is Expressed as a Catalytically Active and Thermostable Dimer in Escherichia coli. Appl. Environ. Microbiol. 1998, 64, 2357–2360. [Google Scholar] [PubMed]
- Suekane, M.M.; Tamura, M.; Tomimura, C. Physico-chemical and Enzymatic Properties of Purified Glucose Isomerases from Streptomyces olivochromogenes and Bacillus stearothermophilus. Agric. Biol. Chem. 1978, 42, 909–917. [Google Scholar] [CrossRef]
- Inyang, C.U.; Gebhart, U.; Obi, S.K.C.; Bisswanger, H. Isolation and characterization of a d-glucose/xylose isomerase from a new thermophilic strain Streptomyces sp. (PLC). Appl. Microbiol. Biotechnol. 1995, 43, 632–638. [Google Scholar] [CrossRef]
- Lee, C.; Zeikus, G. Purification and characterization of thermostable glucose isomerase from Clostridium thermosulfurogenes and Thermoanaerobacter strain B6A. Biochem. J. 1991, 273, 565–571. [Google Scholar] [CrossRef]
- Lee, Y.E.; Ramesh, M.V.; Zeikus, J.G. Cloning, sequencing and biochemical characterization of xylose isomerase from Thermoanaerobacterium saccharolyticum strain B6A-RI. J. Gen. Microbiol. 1993, 139, 1227–1234. [Google Scholar] [CrossRef]
- Srih-Belghith, K.; Bejar, S. A thermostable glucose isomerase having a relatively low optimum pH: Study of activity and molecular cloning of the corresponding gene. Biotechnol. Lett. 1998, 20, 553–556. [Google Scholar] [CrossRef]
- Vieille, C.; Hess, J.M.; Kelly, R.M.; Zeikus, J.G. XylA cloning and sequencing and biochemical characterization of xylose isomerase from Thermotoga neapolitana. Appl. Environ. Microbiol. 1995, 61, 1867–1875. [Google Scholar]
- Konak, L.; Kolcuoğlu, Y.; Ozbek, E.; Colak, A.; Ergenoglu, B. Purification and characterization of an extremely stable glucose isomerase from Geobacillus thermodenitrificans TH2. Appl. Biochem. Microbiol. 2014, 50, 25–29. [Google Scholar] [CrossRef]
- Chauthaiwale, J.; Rao, M. Production and Purification of Extracellular D-Xylose Isomerase from an Alkaliphilic, Thermophilic Bacillus sp. Appl. Environ. Microb. 1994, 60, 4495–4499. [Google Scholar]
- Kumar, C.G.; Takagi, H. Microbial alkaline proteases: from a bioindustrial viewpoint. Biotechnol. Adv. 1999, 17, 561–594. [Google Scholar] [CrossRef]
- Rahman, R.N.Z.A.; Razak, C.N.; Ampon, K.; Basri, M.; Zin, W.M.; Yunus, W.; Salleh, A.B. Purification and characterization of a heat-stable alkaline protease from Bacillus stearothermophilus F1. Appl. Microbiol. Biotechnol. 1994, 40, 822–827. [Google Scholar] [CrossRef]
- Hawumba, J.F.; Theron, J.; Brözel, V.S. Thermophilic protease-producing Geobacillus from Buranga hot springs in Western Uganda. Curr. Microbiol. 2002, 45, 144–150. [Google Scholar] [CrossRef]
- Contesini, F.J.; de Melo, R.R.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef]
- Dipasquale, L.; Calandrelli, V.; Romano, I.; Nicolaus, B.; Gambacota, A.; Lama, L. Purification and characterisation of a highly thermostable extracellular protease from Bacillus thermantarcticus, strain M1. Ann. Microbiol. 2008, 58, 253–259. [Google Scholar] [CrossRef]
- Sako, Y.; Croocker, P.C.; Ishida, Y. An extremely heat-stable extracellular proteinase (aeropyrolysin) from the hyperthermophilic archaeon Aeropyrum pernix K. FEBS Lett. 1997, 415, 329–334. [Google Scholar] [CrossRef]
- Johnvesly, B.; Naik, G.R. Studies on production of thermostable alkaline protease from thermophilic and alkaliphilic Bacillus sp. JB-99 in a chemically defined medium. Proc. Biochem. 2001, 37, 139–144. [Google Scholar] [CrossRef]
- Zhu, W.; Cha, D.; Cheng, G.; Peng, Q.; Shen, P. Purification and characteriztion of a thermostable protease from a newly isolated Geobacillus sp. YMTC 1049. Enzym. Microbiol. Technol. 2007, 40, 1592–1597. [Google Scholar] [CrossRef]
- Thebti, W.; Riahi, Y.; Belhadj, O. Purification and Characterization of a New Thermostable, Haloalkaline, Solvent Stable, and Detergent Compatible Serine Protease from Geobacillus toebii strain LBT 77. BioMed Res. Int. 2016, 2016, 9178962. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.Y.; Wang, G.L.; Li, D.; Zhao, D.L.; Qin, Q.L.; Chen, X.L.; Chen, B.; Zhou, B.C.; Zhang, X.Y.; Zhang, Y.Z. Diversity of Both the Cultivable Protease-Producing Bacteria and Bacterial Extracellular Proteases in the Coastal Sediments of King George Island, Antarctica. PLoS ONE 2013, 8, e79668. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, S.C.; Coria, S.H.; Mac Cormack, W.P. Extracellular proteases from eight psychrotolerant antarctic strains. Microbiol. Res. 2004, 159, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, S.S.; Sharma, J.; Battan, B. Industrial applications and future prospects of microbial xylanases: A review. BioResources 2008, 3, 1377–1402. [Google Scholar]
- Muzard, M.; Aubry, N.; Plantier-Royon, R.; O’Donohue, M.; Remond, C. Evaluation of the transglycosylation activities of a GH 39 β-D-xylosidase for the synthesis of xylose-based glycosides. J. Mol. Catal. B Enzym. 2009, 58, 1–5. [Google Scholar] [CrossRef]
- Nacos, M.K.; Katapodis, P.; Pappas, C.; Daferera, D.; Tarantilis, P.A.; Christakopoulos, P.; Polissiou, M. Kenaf xylan—A source of biologically active acidic oligosaccharides. Carbohydr. Polym. 2006, 66, 126–134. [Google Scholar] [CrossRef]
- Vazquez, M.J.; Alonso, J.L.; Dominguez, H.; Parajo, J.C. Xylooligosaccharides: Manufacture and applications. Trends Food Sci. Technol. 2000, 11, 387–393. [Google Scholar] [CrossRef]
- Samanta, A.C.; Jayapal, N.; Jayaram, C.; Roy, S.; Kolte, A.P.; Senani, S.; Sridhar, M. Xylooligosaccharides as prebiotics from agricultural by-products: Production and applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 62–71. [Google Scholar] [CrossRef]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (XOS) as an Emerging Prebiotic: Microbial Synthesis, Utilization, Structural Characterization, Bioactive Properties and Applications. Compr. Rev. Food Sci. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Kabel, M.A.; Kortenoeven, L.; Schols, H.A.; Voragen, A.G. In vitro fermentability of differently substituted xylo-oligosaccharides. J. Agric. Food Chem. 2002, 50, 6205–6210. [Google Scholar] [CrossRef]
- Immerzeel, P.; Falck, P.; Galbe, M.; Adlercreutz, P.; Nordberg Karlsson, E.; Stålbrand, H. Extraction of water-soluble xylan from wheat bran and utilization of enzymatically produced xylooligosaccharides by Lactobacillus bifidobacterium and Weissella spp., LWT. Food Sci. Technol. 2014, 56, 321–327. [Google Scholar] [CrossRef]
- Driss, D.; Zouari-Ellouzi, S.; Chaari, F.; Kallel, F.; Ghazala, I.; Bouaziz, F.; Chaabouni, S.E. Production and in vitro evaluation of xylooligosaccharides generated from corncobs using immobilized Penicillium occitanis xylanase. J. Mol. Catal. B Enzym. 2014, 102, 146–153. [Google Scholar] [CrossRef]
- Manju, S.; Chadha, B.S. Biofuels: Alternative Feedstocks and Conversion Processes; Pandey, A., Larroche, C., Ricke, S.C., Dussap, C.G., Gnansounou, E., Eds.; Academic Press: New York, NY, USA, 2011; Volume 9, pp. 203–228. [Google Scholar]
- Verbruggen, M.A.; Beldman, G.; Voragen, A.G.J. The Selective Extraction of Glucuronoarabinoxylans from Sorghum Endosperm Cell Walls using Barium and Potassium Hydroxide Solutions. J. Cereal Sci. 1995, 21, 271–282. [Google Scholar] [CrossRef]
- Mathew, S.; Aronsson, A.; Karlsson, E.N.; Adlercreutz, P. Xylo-and arabinoxylooligosaccharides from wheat bran by endoxylanases, utilisation by probiotic bacteria, and structural studies of the enzymes. Appl. Microbiol. Biotechnol. 2018, 102, 3105–3120. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Peng, P.; Xu, F.; Sun, R.C. Fractional purification and bioconversion of hemicelluloses. Biotechnol. Adv. 2012, 30, 879–903. [Google Scholar] [CrossRef] [PubMed]
- Bernfeld, P. Amylases α and β. In Methods in Enzymology; Kaplan, N., Colowick, S., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 149–158. [Google Scholar]
- Ratanakhanokchai, K.; Kyu, K.L.; Tanticharoen, M. Purification and Properties of a Xylan-Binding Endoxylanase from Alkaliphilic Bacillus sp. strain K-1. Appl. Environ. Microbiol. 1999, 65, 694–697. [Google Scholar] [PubMed]
- Huang, D.; Liu, J.; Qi, Y.; Yang, K.; Xu, Y.; Feng, L. Synergistic hydrolysis of xylan using novel xylanases, β-xylosidases, and an α-L-arabinofuranosidase from Geobacillus thermodenitrificans NG80-2. Appl. Microbiol. Biotechnol. 2017, 101, 6023–6037. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; Van de Wiele, T.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef]
- Yan, Q.J.; Wang, L.; Jiang, Z.Q.; Yang, S.Q.; Zhu, H.F.; Li, L.T. A xylose-tolerant beta-xylosidase from Paecilomyces thermophila: characterization and its coaction with the endogenous xylanase. Bioresour. Technol. 2008, 99, 5402–5410. [Google Scholar] [CrossRef]
- Horneck, G.; Walter, N.; Westall, F.; Grenfell, J.L.; Martin, W.F.; Gomez, F.; Leuko, S.; Lee, N.; Onofri, S.; Tsiganis, K.; et al. AstRoMap European Astrobiology Roadmap. Astrobiology 2016, 16, 201–243. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D.P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from Marine and Marine Extremophilic Bacteria: Structures, Properties, Ecological Roles and Applications. Mar. Drugs 2018, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring the marine environments for the identification of extremophiles and their enzymes for sustainable and green bioprocesses. Review. Sustainabylity 2019, 11, 149. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial Exo-xylanase: A mini Review. Appl. Biochem. Biotechnol. 2014, 174, 81–92. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Molecular Weight (kDa) | Optimum Temperature (°C) | Optimum pH | Thermostability |
|---|---|---|---|---|
| Parageobacillus thermantarcticus | 200 | 90 | 7.0 | 90 °C |
| Geobacillus stearothermophilus | 130 | 80 | 7.5–8.0 | 75 °C |
| Geobacillus thermodenitrificans TH2 | 49 * | 80 | 7.5 | 50 °C × 4 days |
| Bacillus sp. (NCIM 59) | 160 | 85 | 8.0 | 80 °C |
| Thermotoga maritima | 200 | 105–110 | 6.5–7.5 | 100 °C |
| Thermotoga neapolitana | 200 | 95 | 7.1 | Half life 95 °C × 24’ |
| Thermus thermophilus | 200 | 95 | 7.0 | 85 °C × 8 h |
| Thermus aquaticus HB8 | 196 | 85 | 7.5 | 70 °C × 1 month |
| Thermoanaerobacterium saccharolyticum B6AR1 | 200 | 80 | 7.0–7.5 | 85 °C × 1 h |
| Thermoanaerobacterium JW/SL-YS 489 | 200 | 80 | 6.8 | 80 °C × 1 h |
| Clostridium thermosulfurogenes | 200 | 80 | 7.0–7.5 | 85 °C × 1 h |
| Streptomyces sp. (PLC) | 183 | 80 | 7.0 | 53 °C × 10 days |
| Streptomyces sp. SK | 185 | 95 | 6.0 | 80 °C × 5 h |
| Media | Cell Growth (O.D.540 nm) | Protease Total Units | Protease Production (U/O.D.540 nm) * |
|---|---|---|---|
| YN | 1.705 | 17.6 | 10.3 |
| YN + milk 0.1% | 1.915 | 18.4 | 9.6 |
| Medium A | 0.918 | 30.0 | 32.7 |
| Medium A + milk 0.1% | 1.334 | 253 | 190 |
| Medium B | 1.290 | 28.8 | 22.3 |
| Medium B + milk 0.1% | 1.660 | 35.2 | 21.2 |
| Microorganisms | Molecular Mass (kDa) | Optimal pH and Temperature | pH and Temperature of the Stability |
|---|---|---|---|
| B. megaterium RRM2 | 27 | pH 10 and 60 °C | pH 7–11 and 60 °C for 2 h of incubation |
| B. koreensis BK-P21A | 48 | pH 9.0 and 60 °C | pH 7–10 and 70 °C for 1 h of incubation |
| B. alveayuensis CAS5 | 33 | pH 9.0 and 50 °C | pH 8–11 and 80 °C for 1 h of incubation |
| B. amyloliquefaciens SYB-001 | 36.8 | pH 7.0 and 50 °C | pH 6–10 and 60 °C for 1 h of incubation |
| B. subtilis GA CAS | 41 | pH 9.0 and 50 °C | pH 9–11 and 60 °C for 24 h of incubation |
| P. thermantarcticus | 42 | pH 7.0 and 70 °C | pH 7–10 and 70 °C for 24 h of incubation |
| Geobacillus YMTC 1049 | 59.2 | pH 7 and 85 °C | pH 6.0–9.0 and 65 °C for 10 h of incubation |
| Geobacillus toebii LBT 77 | 30 | pH 13 and 95 °C | pH 9.0–13.0 and 95 °C for 70 min |
| Hemicellulose Source | Amount of Hemicellulose (mg/mL) | Biocatalyst | S/B | RS% | Xylose % |
|---|---|---|---|---|---|
| A. donax | 4 | ES-GTM | 13.5 | 73.5 | 26.5 |
| A. donax | 4.4 | ES-GTM | 36.3 | 63.1 | 9.85 |
| C. cardunculus | 4 | ES-GTM | 3.58 | 62 | 44 |
| C. cardunculus | 4 | ES-GTM endo-xyl | 17.5 | 43.7 | 6.8 |
| C. cardunculus | 4 | ES-GTM endo-xyl + ES-GTM beta-xyl | 17.5 | 73.6 | 62.6 |
| Temperature Variations (from −196 °C to 80 °C) | Desiccation (up to 1 Year) | UV-Ray Irradiation (at λ 254 nm) | X-Rays Exposure (up to 1 kGy) | |
|---|---|---|---|---|
| Growth’s kinetics | n.m. | Slight delay | n.m. | |
| Membranes lipids’ fatty acids | Increase of branched fatty acids fraction | |||
| Membranes’ quinones | Partial depletion of demethylated menaquinone | n.m. | Partial depletion of demethylated menaquinone | |
| Protein pattern | n.m. | n.m. | n.m. | n.m. |
| DNA | n.m. | n.m. | Minor modifications | n.m. |
| Species | Bacillus pumilus | Bacillus subtilis | Bacillus thuringiensis | Salibacillus marismortui | Bacillus megaterium | Bacillus subtilis luxAB | Parageobacillus thermantarcticus | Bacillus cereus |
|---|---|---|---|---|---|---|---|---|
| D10 | 1.26 kGy | 1.5 kGy | 2.0 kGy | 2.0 kGy | 2.50 kGy | 2.50 kGy | >2.50 kGy | 3.0 kGy |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finore, I.; Lama, L.; Di Donato, P.; Romano, I.; Tramice, A.; Leone, L.; Nicolaus, B.; Poli, A. Parageobacillus thermantarcticus, an Antarctic Cell Factory: From Crop Residue Valorization by Green Chemistry to Astrobiology Studies. Diversity 2019, 11, 128. https://doi.org/10.3390/d11080128
Finore I, Lama L, Di Donato P, Romano I, Tramice A, Leone L, Nicolaus B, Poli A. Parageobacillus thermantarcticus, an Antarctic Cell Factory: From Crop Residue Valorization by Green Chemistry to Astrobiology Studies. Diversity. 2019; 11(8):128. https://doi.org/10.3390/d11080128
Chicago/Turabian StyleFinore, Ilaria, Licia Lama, Paola Di Donato, Ida Romano, Annabella Tramice, Luigi Leone, Barbara Nicolaus, and Annarita Poli. 2019. "Parageobacillus thermantarcticus, an Antarctic Cell Factory: From Crop Residue Valorization by Green Chemistry to Astrobiology Studies" Diversity 11, no. 8: 128. https://doi.org/10.3390/d11080128
APA StyleFinore, I., Lama, L., Di Donato, P., Romano, I., Tramice, A., Leone, L., Nicolaus, B., & Poli, A. (2019). Parageobacillus thermantarcticus, an Antarctic Cell Factory: From Crop Residue Valorization by Green Chemistry to Astrobiology Studies. Diversity, 11(8), 128. https://doi.org/10.3390/d11080128