Adaptations by Zostera marina Dominated Seagrass Meadows in Response to Water Quality and Climate Forcing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Seagrass Transect Surveys
2.2. Aerial Surveys
2.3. Water Quality
2.4. Data Analyses
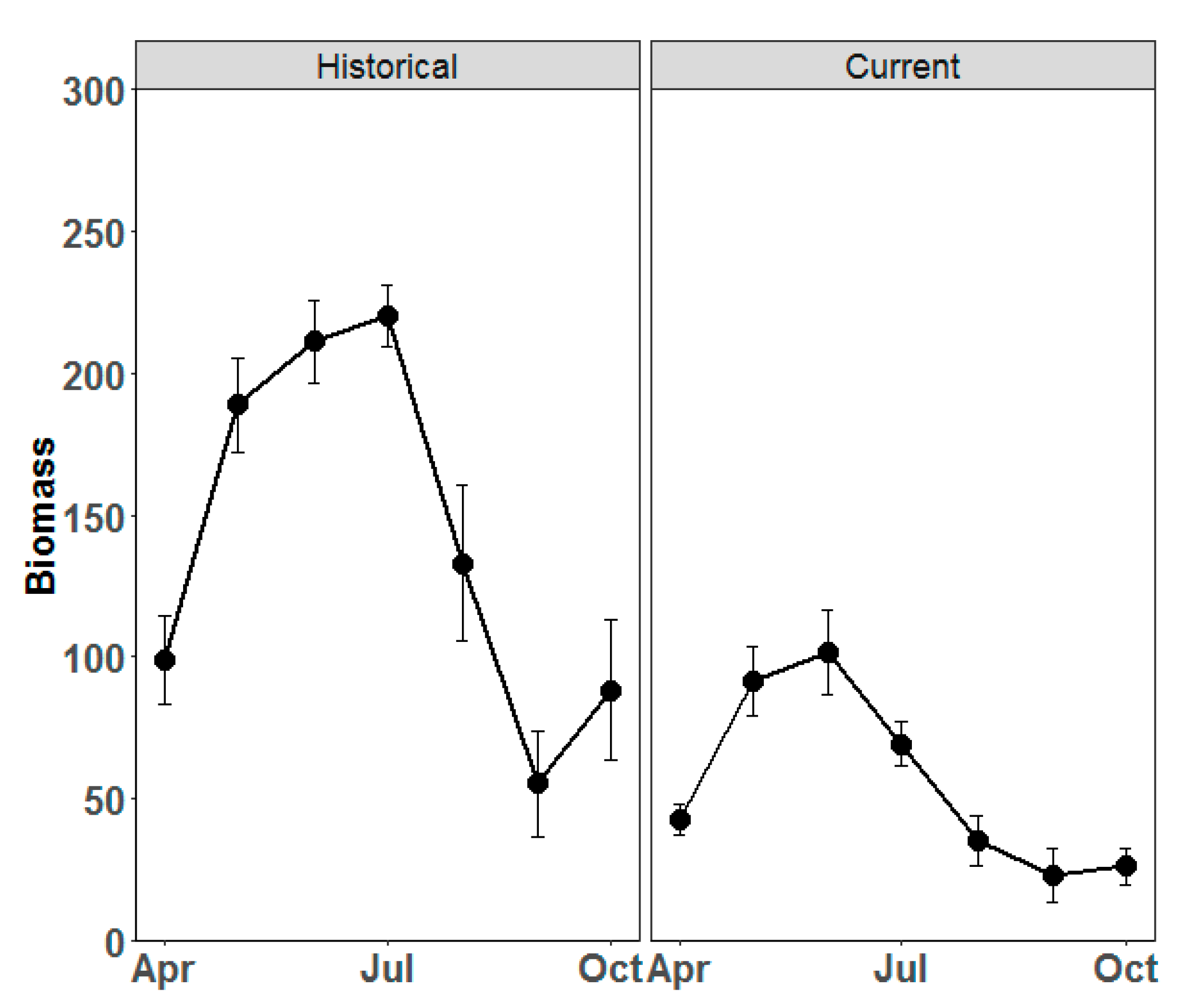
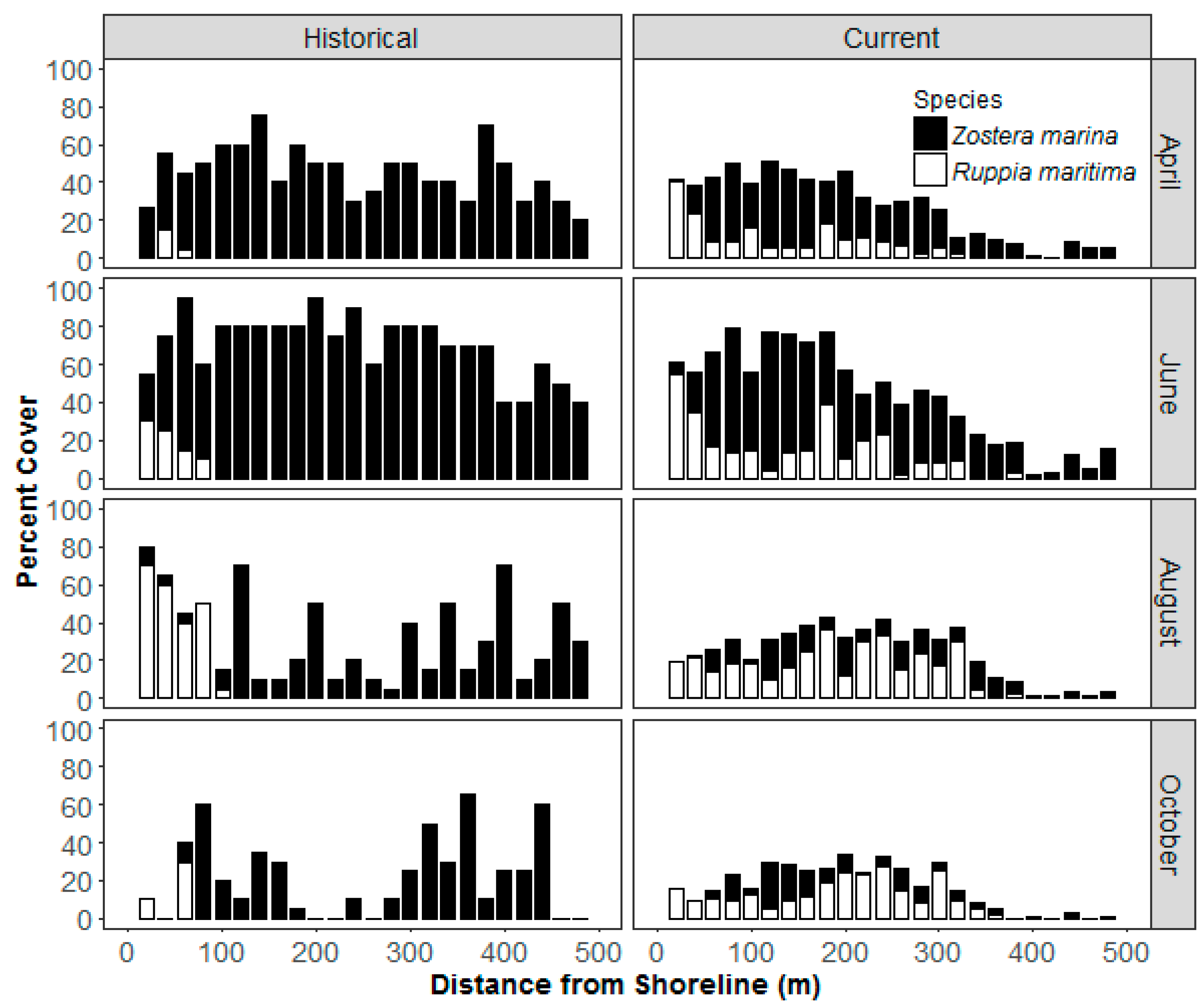
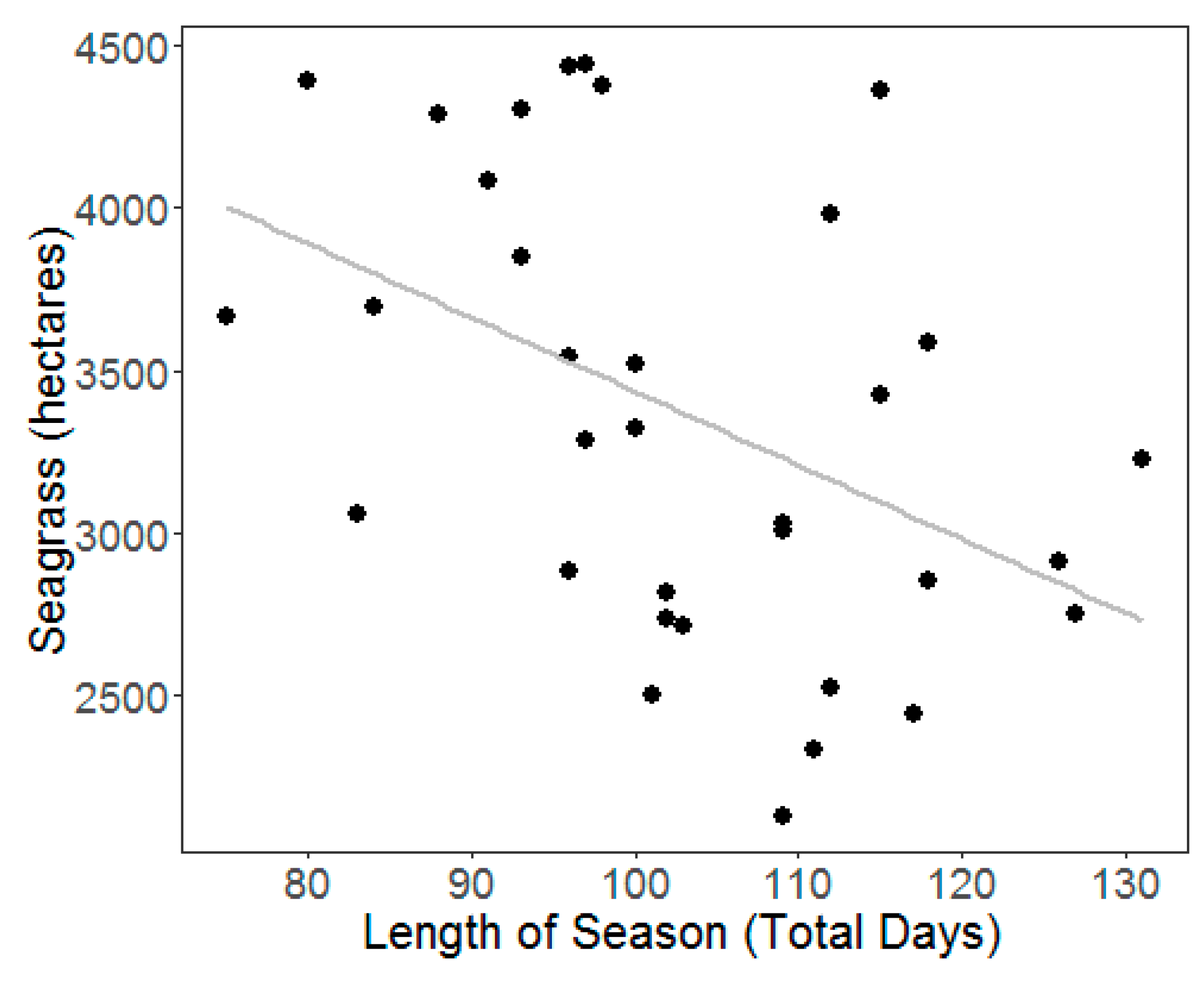
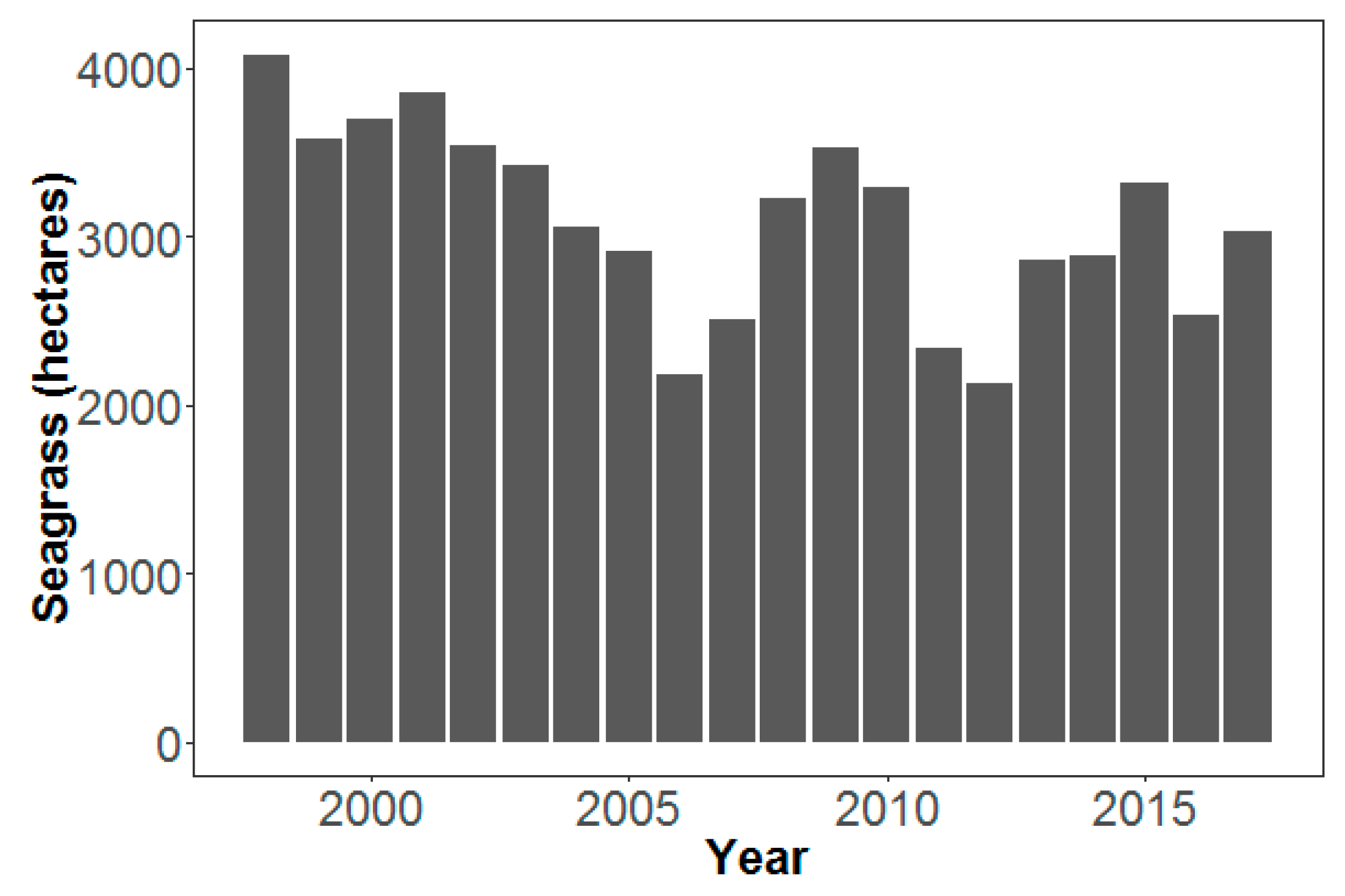
3. Results
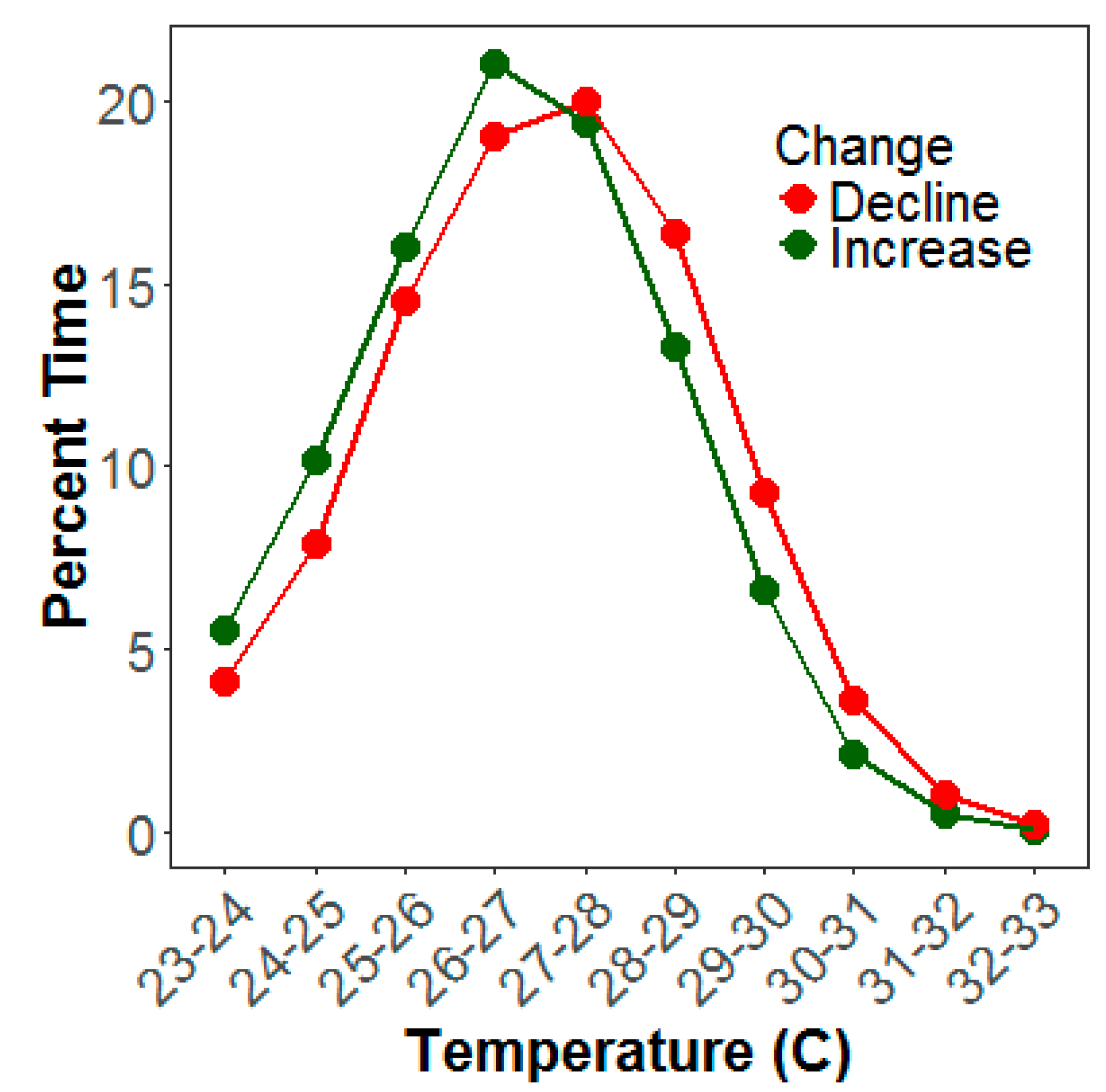
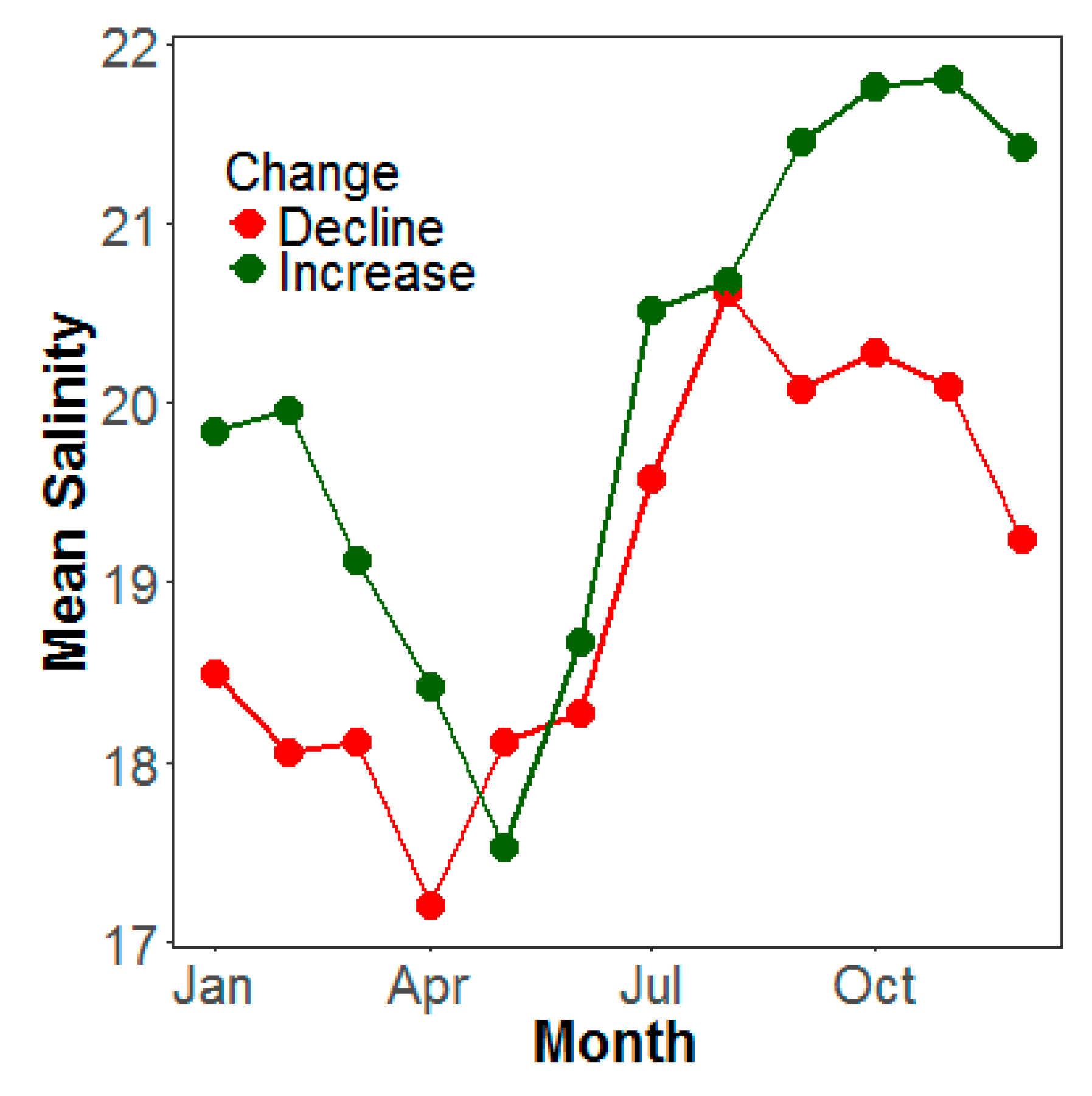
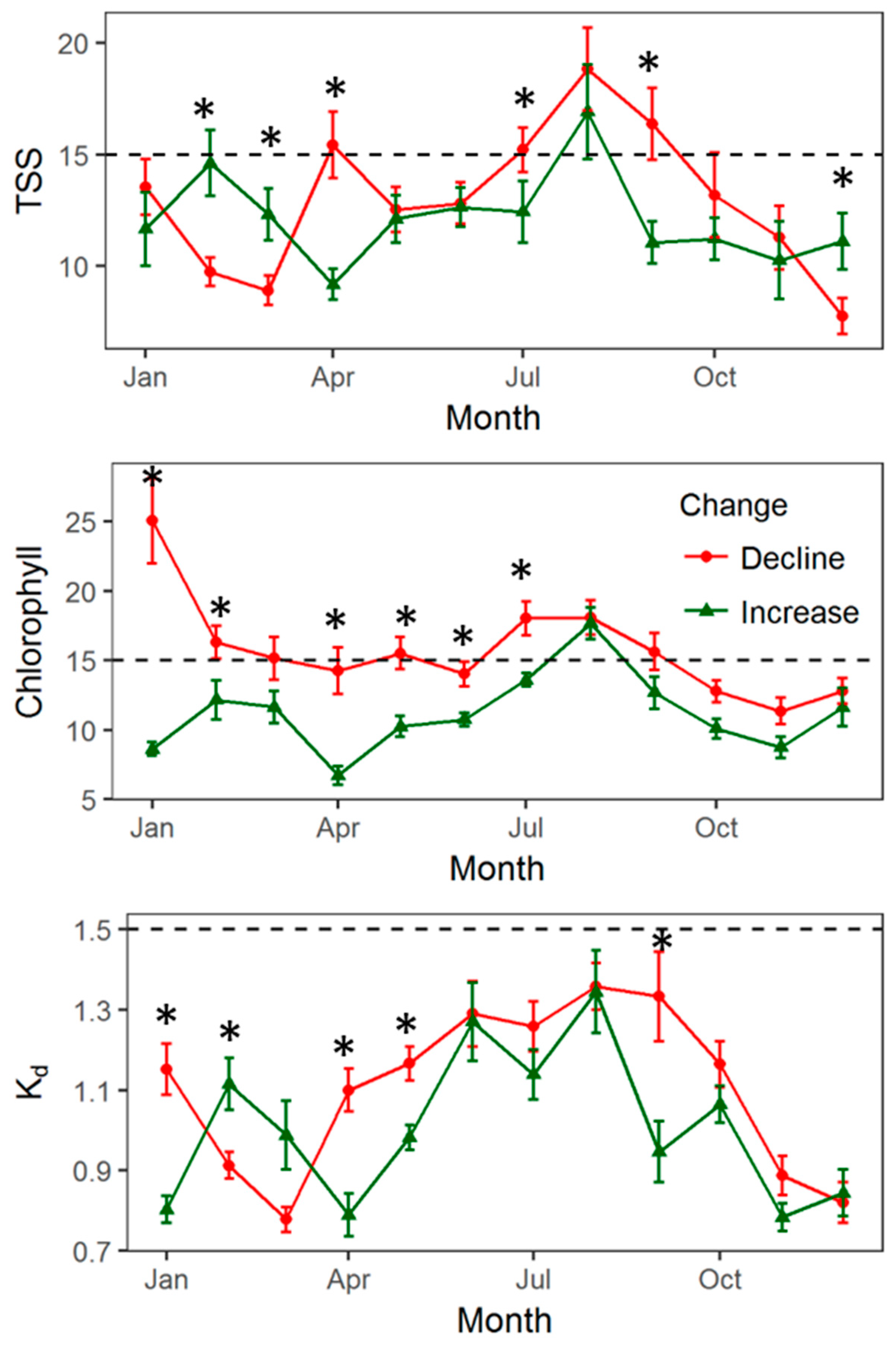
4. Discussion
4.1. Growth Pattern
4.2. Climate and Water Quality Conditions
4.3. Resiliency
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.R.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Menzel, A.; Estrella, N.; Fabian, P. Spatial and temporal variability of the phenological seasons in Germany from 1951 to 1996. Glob. Chang. Biol. 2001, 7, 657–666. [Google Scholar] [CrossRef]
- Bradley, N.L.; Leopold, A.C.; Ross, J.; Huffaker, W. Phenological changes reflect climate change in Wisconsin. Proc. Natl. Acad. Sci. USA 1999, 96, 9701–9704. [Google Scholar] [CrossRef] [PubMed]
- Overpeck, J.T.; Rind, D.; Goldberg, R. Climate-induced changes in forest disturbance and vegetation. Nature 1990, 343, 51–53. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A Global Crisis for Seagrass Ecosystems. BioScience 2006, 56, 987–996. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377. [Google Scholar] [CrossRef] [PubMed]
- Vaudrey, J.M.P.; Kremer, J.N.; Branco, B.F.; Short, F.T. Eelgrass recovery after nutrient enrichment reversal. Aquat. Bot. 2010, 93, 237–243. [Google Scholar] [CrossRef]
- Short, F.T.; Coles, R.; Fortes, M.D.; Victor, S.; Salik, M.; Isnain, I.; Andrew, J.; Seno, A. Monitoring in the Western Pacific region shows evidence of seagrass decline in line with global trends. Mar. Pollut. Bull. 2014, 83, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Short, F.T.; Neckles, H.A. The effects of global climate change on seagrasses. Aquat. Bot. 1999, 63, 169–196. [Google Scholar] [CrossRef]
- Jordà, G.; Marbà, N.; Duarte, C.M. Mediterranean seagrass vulnerable to regional climate warming. Nat. Clim. Chang. 2012, 2, 821–824. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Wilcox, D.J.; Murphy, R.R.; Marion, S.R.; Orth, R.J. Multiple stressors threaten the imperiled coastal foundation species eelgrass (Zostera marina) in Chesapeake Bay, USA. Glob. Chang. Biol. 2017, 23, 3474–3483. [Google Scholar] [CrossRef] [PubMed]
- Orth, R.J.; Moore, K.A. Chesapeake Bay: An Unprecedented Decline in Submerged Aquatic Vegetation. Science 1983, 222, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Batiuk, R.A.; Orth, R.J.; Moore, K.A.; Dennison, W.C.; Stevenson, J.C.; Staver, L.W.; Carter, V.; Rybicki, N.B.; Hickman, R.E.; Kollar, S.; et al. Chesapeake Bay Submerged Aquatic Vegetation Habitat Requirements and Restoration Targets: A Technical Synthesis; Chesapeake Bay Program; EPA: Annapolis, MD, USA, 1992; 170p. [Google Scholar]
- Batiuk, R.; Bergstrom, P.; Kemp, M.; Koch, E.; Murray, L.; Stevenson, C.; Bartleson, R.; Carter, V.; Rybicki, N.; Landwehr, J.; et al. Chesapeake Bay Submerged Aquatic Vegetation Water Quality and Habitat-Based Requirements and Restoration Targets: A Second Technical Synthesis; Chesapeake Bay Program; EPA: Annapolis, MD, USA, 2000; 220p. [Google Scholar]
- Orth, R.J.; Dennison, W.C.; Lefcheck, J.S.; Gurbisz, C.; Hannam, M.; Keisman, J.; Landry, J.B.; Moore, K.A.; Murphy, R.R.; Patrick, C.J.; et al. Submersed Aquatic Vegetation in Chesapeake Bay: Sentinel Species in a Changing World. BioScience 2017, 67, 698–712. [Google Scholar] [CrossRef]
- Moore, K.A.; Wilcox, D.J.; Orth, R.J. Analysis of the Abundance of Submersed Aquatic Vegetation Communities in the Chesapeake Bay. Estuaries 2000, 23, 115. [Google Scholar] [CrossRef]
- Koch, E.W.; Orth, R.J. Seagrasses of the Mid-Atlantic Coast of the United States. In World Atlas of Seagrasses; Green, E.P., Short, F.T., Eds.; University of California Press: Berkeley, CA, USA, 2003; pp. 216–223. ISBN 0520240472. [Google Scholar]
- Jarvis, J.C.; Moore, K.A. The role of seedlings and seed bank viability in the recovery of Chesapeake Bay, USA, Zostera marina populations following a large-scale decline. Hydrobiologia 2010, 649, 55–68. [Google Scholar] [CrossRef]
- Moore, K.A.; Shields, E.C.; Parrish, D.B. Impacts of Varying Estuarine Temperature and Light Conditions on Zostera marina (Eelgrass) and its Interactions with Ruppia maritima (Widgeongrass). Estuar. Coasts 2014, 37, 20–30. [Google Scholar] [CrossRef]
- Carr, J.A.; D’Odorico, P.; McGlathery, K.J.; Wiberg, P.L. Modeling the effects of climate change on eelgrass stability and resilience: Future scenarios and leading indicators of collapse. Mar. Ecol. Prog. Ser. 2012, 448, 289–301. [Google Scholar] [CrossRef]
- Abe, M.; Kurashima, A.; Maegawa, M. Temperature requirements for seed germination and seedling growth of Zostera marina from central Japan. Fish. Sci. 2008, 74, 589–593. [Google Scholar] [CrossRef]
- Moore, K.A.; Jarvis, J.C. Environmental Factors Affecting Recent Summertime Eelgrass Diebacks in the Lower Chesapeake Bay: Implications for Long-term Persistence. J. Coast. Res. 2008, 135–147. [Google Scholar] [CrossRef]
- Marsh, J.A.; Dennison, W.C.; Alberte, R.S. Effects of temperature on photosynthesis and respiration in eelgrass (Zostera marina L.). J. Exp. Mar. Biol. Ecol. 1986, 101, 257–267. [Google Scholar] [CrossRef]
- Moore, K.A.; Wetzel, R.L.; Orth, R.J. Seasonal pulses of turbidity and their relations to eelgrass (Zostera marina L.) survival in an estuary. J. Exp. Mar. Biol. Ecol. 1997, 215, 115–134. [Google Scholar] [CrossRef]
- Holmer, M.; Bondgaard, E.J. Photosynthetic and growth response of eelgrass to low oxygen and high sulfide concentrations during hypoxic events. Aquat. Bot. 2001, 70, 29–38. [Google Scholar] [CrossRef]
- Holmer, M.; Hasler-Sheetal, H. Sulfide intrusion in seagrasses assessed by stable sulfur isotopes—A synthesis of current results. Front. Mar. Sci. 2014, 1, 64. [Google Scholar] [CrossRef]
- Orth, R.J.; Marion, S.R.; Moore, K.A.; Wilcox, D.J. Eelgrass (Zostera marina L.) in the Chesapeake Bay Region of Mid-Atlantic Coast of the USA: Challenges in Conservation and Restoration. Estuar. Coasts 2010, 33, 139–150. [Google Scholar] [CrossRef]
- Moore, K.A. Influence of seagrasses on water quality in shallow regions of the lower Chesapeake Bay. J. Coast. Res. 2004, 45, 162–178. [Google Scholar] [CrossRef]
- SAV in Chesapeake Bay and Coastal Bays. Available online: http://web.vims.edu/bio/sav/index.html (accessed on 16 August 2018).
- Virginia Estuarine and Coastal Observing System (VECOS). Available online: http://web2.vims.edu/vecos/ (accessed on 16 August 2018).
- Shoaf, W.T.; Lium, B.W. Improved extraction of chlorophyll a and b from algae using dimethyl sulfoxide. Limnol. Oceanogr. 1976, 21, 926–928. [Google Scholar] [CrossRef]
- RStudio Team. Integrated Development for RStudio; RStudio Inc.: Boston, MA, USA, 2016; Available online: http://www.rstudio.com/ (accessed on 6 January 2018).
- Changing Chesapeake. Available online: http://www.chesapeakedata.com/changingchesapeake/ (accessed on 16 August 2018).
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Abu-Asab, M.S.; Peterson, P.M.; Shetler, S.G.; Orli, S.S. Earlier plant flowering in spring as a response to global warming in the Washington, DC, area. Biodivers. Conserv. 2001, 10, 597–612. [Google Scholar] [CrossRef]
- Ho, C.-H.; Lee, E.-J.; Lee, I.; Jeong, S.-J. Earlier spring in Seoul, Korea. Int. J. Climatol. 2006, 26, 2117–2127. [Google Scholar] [CrossRef]
- Linderholm, H.W. Growing season changes in the last century. Agric. For. Meteorol. 2006, 137, 1–14. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Burke, M.K.; Dennison, W.C.; Moore, K.A. Non-structural carbohydrate reserves of eelgrass Zostera marina. Mar. Ecol. Prog. Ser. 1996, 137, 195–201. [Google Scholar] [CrossRef]
- Shields, E.C.; Moore, K.A.; Parrish, D.B. Short-term temperature stress results in seagrass community shift in a temperate estuary. Estuar. Coasts 2018. under review. [Google Scholar]
- Boström, C.; Baden, S.; Bockelmann, A.-C.; Dromph, K.; Fredriksen, S.; Gustafsson, C.; Krause-Jensen, D.; Möller, T.; Nielsen, S.L.; Olesen, B.; et al. Distribution, structure and function of Nordic eelgrass (Zostera marina) ecosystems: Implications for coastal management and conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 410–434. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.G.; Pyke, C.R.; Adams, M.B.; Breitburg, D.; Hershner, C.; Kemp, M.; Howarth, R.; Mulholland, M.R.; Paolisso, M.; Secor, D.; et al. Potential climate-change impacts on the Chesapeake Bay. Estuar. Coast. Shelf Sci. 2010, 86, 1–20. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Collier, C.J.; Waycott, M.; Mckenzie, L.J.; Cullen-Unsworth, L.C. A framework for the resilience of seagrass ecosystems. Mar. Pollut. Bull. 2015, 100, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Plus, M.; Deslous-Paoli, J.-M.; Dagault, F. Seagrass (Zostera marina L.) bed recolonisation after anoxia-induced full mortality. Aquat. Bot. 2003, 77, 121–134. [Google Scholar] [CrossRef]
- Greve, T.M.; Krause-Jensen, D.; Rasmussen, M.B.; Christensen, P.B. Means of rapid eelgrass (Zostera marina L.) recolonisation in former dieback areas. Aquat. Bot. 2005, 82, 143–156. [Google Scholar] [CrossRef]
- Jarvis, J.C.; Moore, K.A.; Kenworthy, W.J. Characterization and ecological implication of eelgrass life history strategies near the species’ southern limit in the western North Atlantic. Mar. Ecol. Prog. Ser. 2012, 444, 43–56. [Google Scholar] [CrossRef]
- Silberhorn, G.M.; Orth, R.J.; Moore, K.A. Anthesis and seed production in Zostera marina L. (eelgrass) from the Chesapeake Bay. Aquat. Bot. 1983, 15, 133–144. [Google Scholar] [CrossRef]
- Orth, R.J.; Moore, K.A. Seasonal and year-to-year variations in the growth of Zostera marina L. (eelgrass) in the lower Chesapeake Bay. Aquat. Bot. 1986, 24, 335–341. [Google Scholar] [CrossRef]
- Johnson, A.J.; Moore, K.A.; Orth, R.J. The influence of resource availability on flowering intensity in Zostera marina (L.). J. Exp. Mar. Biol. Ecol. 2017, 490, 13–22. [Google Scholar] [CrossRef]
- Jarvis, J.C.; Moore, K.A.; Kenworthy, W.J. Persistence of Zostera marina L. (eelgrass) seeds in the sediment seed bank. J. Exp. Mar. Biol. Ecol. 2014, 459, 126–136. [Google Scholar] [CrossRef]
- Elmqvist, T.; Folke, C.; Nyström, M.; Peterson, G.; Bengtsson, J.; Walker, B.; Norberg, J. Response diversity, ecosystem change, and resilience. Front. Ecol. Environ. 2003, 1, 488–494. [Google Scholar] [CrossRef]
- Johnson, M.R.; Williams, S.L.; Lieberman, C.H.; Solbak, A. Changes in the abundance of the seagrasses Zostera marina L. (eelgrass) and Ruppia maritima L. (widgeongrass) in San Diego, California, following and El Niño Event. Estuaries 2003, 26, 106. [Google Scholar] [CrossRef]
- Richardson, J.P.; Lefcheck, J.S.; Orth, R.J. Warming temperatures alter the relative abundance and distribution of two co-occurring foundational seagrasses in Chesapeake Bay, USA. Mar. Ecol. Prog. Ser. 2018, 599, 65–74. [Google Scholar] [CrossRef]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shields, E.C.; Moore, K.A.; Parrish, D.B. Adaptations by Zostera marina Dominated Seagrass Meadows in Response to Water Quality and Climate Forcing. Diversity 2018, 10, 125. https://doi.org/10.3390/d10040125
Shields EC, Moore KA, Parrish DB. Adaptations by Zostera marina Dominated Seagrass Meadows in Response to Water Quality and Climate Forcing. Diversity. 2018; 10(4):125. https://doi.org/10.3390/d10040125
Chicago/Turabian StyleShields, Erin C., Kenneth A. Moore, and David B. Parrish. 2018. "Adaptations by Zostera marina Dominated Seagrass Meadows in Response to Water Quality and Climate Forcing" Diversity 10, no. 4: 125. https://doi.org/10.3390/d10040125
APA StyleShields, E. C., Moore, K. A., & Parrish, D. B. (2018). Adaptations by Zostera marina Dominated Seagrass Meadows in Response to Water Quality and Climate Forcing. Diversity, 10(4), 125. https://doi.org/10.3390/d10040125