Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
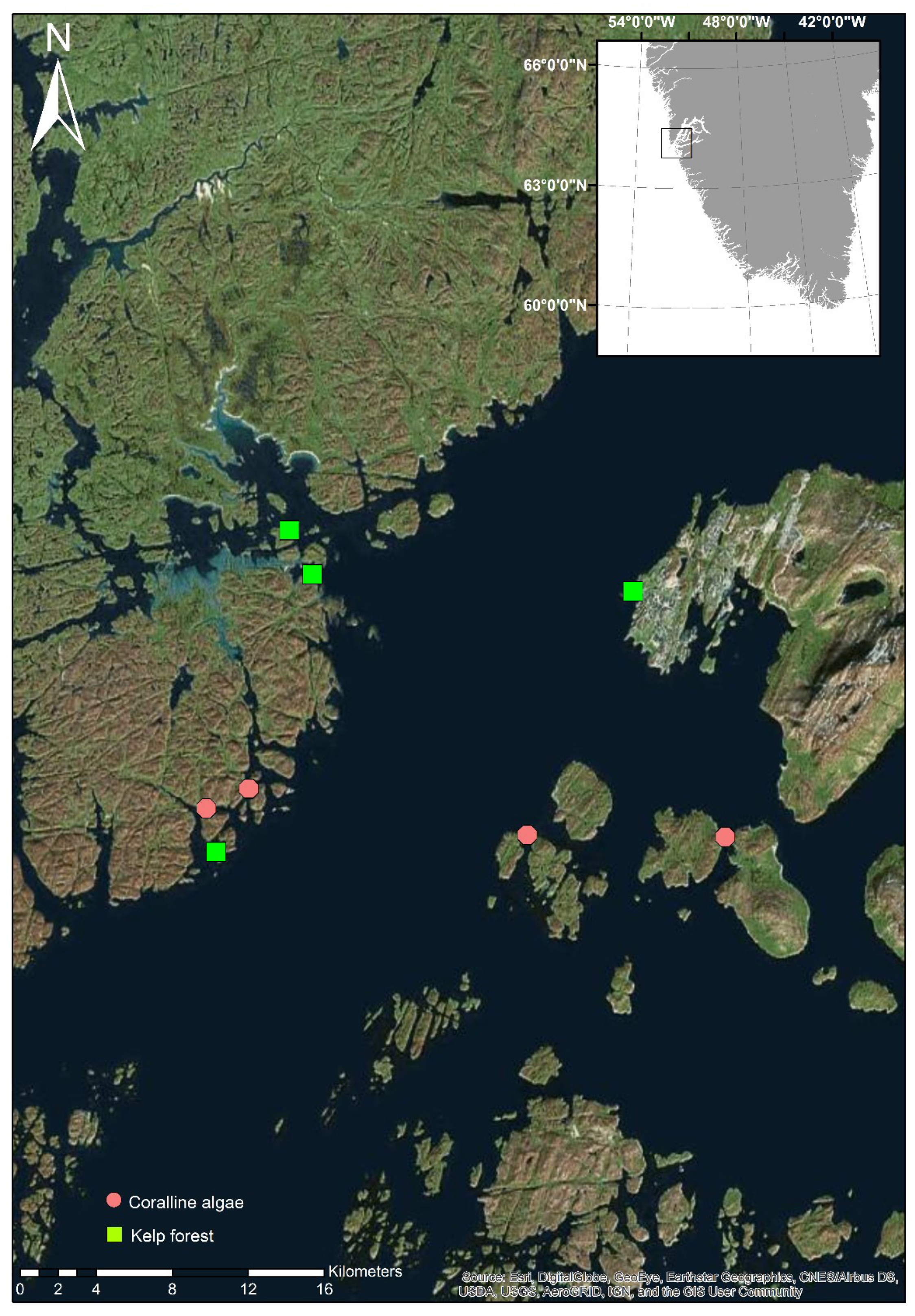
2.1. Field Work
2.2. Laboratory Analysis
2.3. Video and Image Analysis
2.4. Statistical Analysis
3. Results
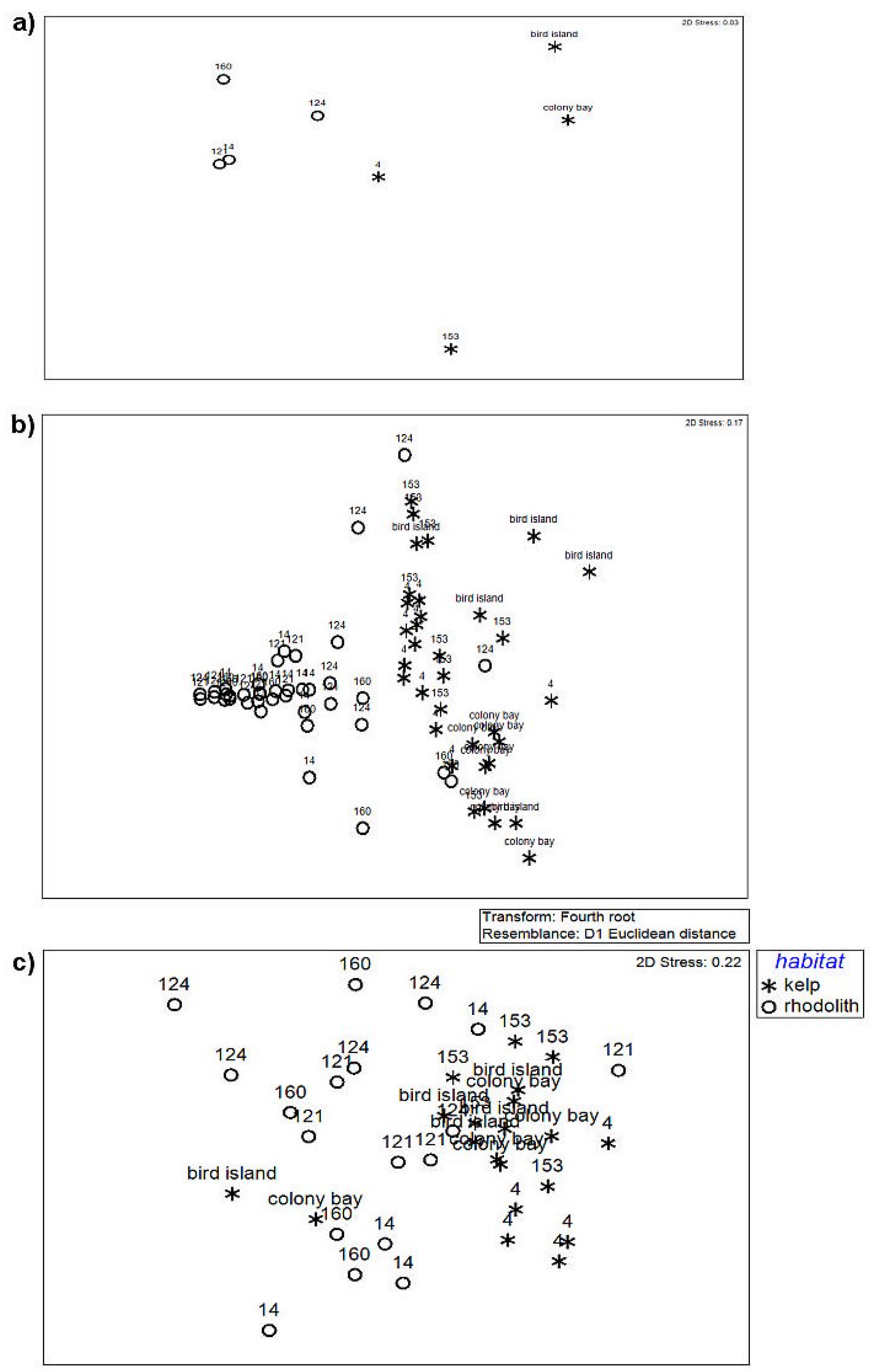
3.1. Habitat Diversity
3.2. Influence of Coralline Algae on Composition and Diversity of Habitats
4. Discussion
4.1. Kelp Versus Coralline Algal Habitats
4.2. Coralline Algae
4.3. Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blicher, M. Structure and Dynamics of Marine Macrozoobenthos in Greenland- and Link to Environmental Drivers; University of Copenhagen: København, Denmark, 2010. [Google Scholar]
- Wiencke, C.; Amsler, C.D. Seaweeds and their communities in polar regions. In Seaweed Biology; Wiencke, C., Bischof, K., Eds.; Springer: Berlin, Germany, 2012; pp. 265–294. [Google Scholar]
- Sakshaug, E. Primary and secondary production in the Arctic seas. In The Organic Carbon Cycle in the Arctic Ocean; Stein, R., MacDonald, R.W., Eds.; Springer: Berlin/Hiedelberg, Germany, 2004; pp. 57–81. [Google Scholar]
- Bonsell, C.; Dunton, K.H. Long-term patterns of benthic irradiance and kelp production in the central Beaufort Sea reveal implications of warming for Arctic inner shelves. Prog. Oceanogr. 2018, 162, 160–172. [Google Scholar] [CrossRef]
- Grebmeier, J.M. Shifting patterns of life in the Pacific Arctic and sub-Arctic seas. Annu. Rev. Mar. Sci. 2012, 4, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Krause-Jensen, D.; Marba, N.; Olesen, B.; Sejr, M.K.; Christensen, P.B.; Rodrigues, J.; Renaud, P.E.; Balsby, T.J.S.; Rysgaard, S. Seasonal sea ice cover as principal driver of spatial and temporal variation in depth extension and annual production of kelp in Greenland. Glob. Chang. Biol. 2012, 18, 2981–2994. [Google Scholar] [CrossRef] [PubMed]
- Dunton, K.H. Growth of dark-exposed Laminaria sacchrina (L.) Lamour. and Laminaria solidungula J. Ag. (Laminariales: Phaeophyta) in the Alaskan Beaufort Sea. J. Exp. Mar. Biol. Ecol. 1985, 94, 181–189. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Dayton, P.K. Ecology of kelp communities. Annu. Rev. Ecol. Syst. 1985, 16, 215–245. [Google Scholar] [CrossRef]
- Steneck, R.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 426–459. [Google Scholar] [CrossRef]
- Hop, H.; Wiencke, C.; Vogele, B.; Kovaltchouk, N.A. Species composition, zonation, and biomass of marine benthic macroalgae in Kongsfjorden, Svalbard. Bot. Mar. 2012, 55, 399–414. [Google Scholar] [CrossRef]
- Freiwald, A. Coralline algal maerl frameworks-islands within the phaeophytic kelp belt. Facies 1993, 29, 133–148. [Google Scholar] [CrossRef]
- Bartsch, I.; Wiencke, C.; Laepple, T. Global seaweed biogeography under a changing climate: The prospected effects of temperature. In Seaweed Biology; Wiencke, C., Bischof, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Assis, J.; Lucas, A.V.; Barbara, I.; Serrao, E.A. Future climate change is predicted to shift long-term persistence zones in the cold-temperate kelp Laminaria hyperborea. Mar. Environ. Res. 2016, 113, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Assis, J.; Araujo, M.B.; Serrao, E.A. Projected climate changes threaten ancient refugia of kelp forests in the north Atlantic. Glob. Chang. Biol. 2018, 24, e55–e66. [Google Scholar] [CrossRef] [PubMed]
- Brodie, J.; Williamson, C.J.; Smale, D.A.; Mieszkowska, N.; Santos, R.; Cunliffe, M.; Steinke, M.; Yesson, C.; Anderson, K.M.; Asnaghi, V.; et al. The future of the northeast Atlantic benthic flora in a high CO2 world. Ecol. Evol. 2014, 4, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, S.; Bartsch, I.; Wiencke, C. New additions to the benthic marine flora of Kongsfjorden, western Svlabard, and comparison between 1996/1998 and 2012/2013. Bot. Mar. 2014, 57, 203–216. [Google Scholar] [CrossRef]
- Schoenrock, K.M.; Bacquet, M.; Pearce, D.; Rea, B.; Schofield, J.E.; Lea, J.; Mair, D.; Kamenos, N. Influences of salinity on the physiology and distribution of the Arctic coalline algae, Lithothamnion glaciale Kjellman (Corallinales, Rhodophyta). J. Phycol. 2018, 54, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Schoenrock, K.M.; Stachnik, L.; Vad, J.; Kamenos, N.A.; Pearce, D.; Rea, B.; Schofield, J.E.; Lea, J.; Mair, D. Distribution of Benthic Communities in a Fjord-Marine System in Southwestern Greenland, with a Focus on Algal Dominated Habitats. In Proceedings of the International Youth Scientific and Practical Conference, “Arctic research: From extensive to integrated development”, Arkhangelsk, Russia, 23–25 April 2018. [Google Scholar]
- Gagnon, P.; Johnson, L.E.; Himmelman, J.H. Kelp patch dynamics in the face of intense herbivory: Stability of Agarum clathratum (Phaeophyta) stands and associated flora on urchin barrens. J. Phycol. 2005, 41, 498–505. [Google Scholar] [CrossRef]
- Olesen, B.; Krause-Jensen, D.; Marba, N.; Christensen, P.B. Eelgrass Zostera marina in subarctic Greenland: Dense meadows with slow biomass turnover in cold waters. Mar. Ecol. Prog. Ser. 2015, 518, 107–121. [Google Scholar] [CrossRef]
- Wulff, A.; Iken, K.; Quartino, M.L.; Al-Handal, A.; Weincke, C.; Clayton, M.N. Biodiversity, biogeography and zonation of marine benthic micro- and macroalgae in the Arctic and Antarctic. Bot. Mar. 2009, 52, 491–507. [Google Scholar] [CrossRef]
- Bolton, J.J. Global seaweed diversity: Patterns and anomalies. Bot. Mar. 1994, 37, 241–246. [Google Scholar] [CrossRef]
- Dunton, K.H.; Schell, D.M. Dependence of consumers on macroalgal (Laminaria solidundula) carbon in an Arctic kelp community: δ13C evidence. Mar. Biol. 1987, 93, 615–625. [Google Scholar] [CrossRef]
- Hernandez-Kantun, J.J.; Hall-Spencer, J.M.; Grall, J.; Adey, W.; Rindi, F.; Maggs, C.A.; Barbara, I.; Pena, V. North Atlantic rhodolith beds. In Rhodolith/Maerl Beds: A Global Perspective; Riosmena-Rodrigeua, R., Ed.; Springer: Cham, Switzerland, 2017; pp. 265–279. [Google Scholar]
- Filbee-Dexter, K.; Wernberg, T.; Fredriksen, S.; Norderhaug, K.M.; Pedersen, M.F. Arctic kelp forests: Diversity, resilience and future. Glob. Planet. Chang. 2019, 172, 1–14. [Google Scholar] [CrossRef]
- Blicher, M.E.; Sejr, M.K.; Rysgaard, S. High carbon demand of dominant macrozoobenthic species indicates their central role in ecosystem carbon flow in a sub-Arctic fjord. Mar. Ecol. Prog. Ser. 2009, 383, 127–140. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Marba, N.; Sanz-Martin, M.; Hendriks, I.E.; Thyrring, J.; Carstensen, J.; Sejr, M.K.; Duarte, C.M. Long photoperiods sustain high pH in Arctic kelp forests. Sci. Adv. 2016, 2, e1501938. [Google Scholar] [CrossRef] [PubMed]
- Krause-Jensen, D.; Duarte, C.M.; Hendriks, I.E.; Meire, L.; Blicher, M.E.; Marba, N.; Sejr, M.K. Macroalgae contribute to nested mosaics of pH variability in a subarctic fjord. Biogeosciences 2015, 12, 4895–4911. [Google Scholar] [CrossRef]
- Ramajo, L.; Perez-Leon, E.; Hendriks, I.E.; Marba, N.; Krause-Jensen, D.; Sejr, M.K.; Blicher, M.E.; Lagos, N.A.; Olsen, Y.S.; Duarte, C.M. Food supply confers calcifiers resistance to ocean acidification. Sci. Rep. 2016, 6, 19374. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.D.; Anderson, K.M.; Demes, K.W.; Kordas, J.P.; Coyle, T.A.; Graham, M.H. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef] [PubMed]
- McCoy, S.J.; Kamenos, N.A. Coralline algae (rhodophyta) in a changing world: Integrating ecological, physiological, and geochemical responses to global change. J. Phycol. 2015, 51, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, L.C.; Bischof, K. Ocean acidification effects of calcifying macroalgae. Aquat. Biol. 2014, 22, 261–279. [Google Scholar] [CrossRef]
- Foster, M.S. Rhodoliths: Between rocks and soft places. J. Phycol. 2001, 37, 659–667. [Google Scholar] [CrossRef]
- Steneck, R.S. The ecology of coralline algal crusts: Convergent patterns and adaptive strategies. Annu. Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar] [CrossRef]
- Chenelot, H.; Jewett, S.C.; Hoberg, M.K. Macrobenthos of the nearshore Aleutian Archipelago, with emphasis on invertebrates associated with Clathromorphum nereostratum (rhodophyta, corallinaceae). Mar. Biodivers. 2011, 41, 413–424. [Google Scholar] [CrossRef]
- Freiwald, A.; Henrich, R. Reefal coralline algal build-ups within the Arctic circle: Morphology and sedimentary dynamics under extreme environmental seasonality. Sedimentology 1994, 41, 963–984. [Google Scholar] [CrossRef]
- Teichert, S. Hollow rhodoliths increase svalbard’s shelf biodiversity. Sci. Rep. 2014, 4, 6972. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.Q.; Feely, R.A.; Carter, B.R.; Greeley, D.J.; Gledhill, D.K.; Arzayus, K.M. Climatological distribution of aragonite saturation state in the global oceans. Glob. Biogeochem. Cycles 2015, 29, 1656–1673. [Google Scholar] [CrossRef]
- Büdenbender, J.; Riebesell, U.; Form, A. Calcification of the arctic coralline red algae Lithothamnion glaciale in response to elevated CO2. Mar. Ecol. Prog. Ser. 2011, 441, 79–87. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Duarte, C.M. Substatial role of macroalgae in marince carbon seqeustration. Nat. Geosci. 2016, 9, 737. [Google Scholar] [CrossRef]
- Van der Heijden, L.H.; Kamenos, N.A. Reviews and syntheses: Calculating the global contribution of coralline algae to total carbon burial. Biogeosciences 2015, 12, 6429–6441. [Google Scholar] [CrossRef]
- Rink, H. Grønland Geographisk og Statistisk Beskrevet; Andr. Fred. Høst: Copenhagen, Denmark, 1852; Volume I. [Google Scholar]
- Matsunga, K.; Kawaguchi, T.; Suzuki, Y.; Nigi, G. The role of terrestrial humic substances on the shift of kelp community to crustose coralline algae community of the southern Hokkaido Island in the Japan Sea. J. Exp. Mar. Biol. Ecol. 1999, 241, 193–205. [Google Scholar] [CrossRef]
- Gagnon, P.; Matheson, K.; Stapleton, M. Variation in rhodolith morphology and biogenic potential of newly discovered rhodolith beds in Newfoundland and Labrador (Canada). Bot. Mar. 2012, 55, 85–99. [Google Scholar] [CrossRef]
- Grall, J.; Loc’h, F.L.; Guyonnet, B.; Riera, P. Community structure and food web based on stable isotopes (δ15N and δ13C) analysis of a north eastern Atlantic maerl bed. J. Exp. Mar. Biol. Ecol. 2006, 338, 1–15. [Google Scholar] [CrossRef]
- Gabara, S. Community Structure and Energy Flow within Rhodolith Habitats at Santa Catalina Island, CA; San Jose State University: Moss Landing, CA, USA, 2014. [Google Scholar]
- Jørgensbye, H.I.; Halfar, J. Overview of coralline red algal crusts and rhodolith beds (Corallinales, Rhodophyta) and their possible ecological importance in Greenland. Polar Biol. 2017, 40, 517–531. [Google Scholar] [CrossRef]
- Gabrielson, P. (University of North Carolina Chapel Hill, Chapel Hill, NC, USA). Personal communication, 2018.
- Hayward, P.J.; Ryland, J.S. Marine Fauna of North-West Europe, 2nd ed.; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Kohler, K.E.; Gill, S.M. Coral point count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, 2.4-2. 2017. Available online: https://cran.r-project.org, https://github.com/vegandevs/vegan (accessed on 22 August 2018).
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Foster, M.S. Algal succession in a Macrocystis pyrifera forest. Mar. Biol. 1975, 32, 313–329. [Google Scholar] [CrossRef]
- Noble-James, T.; Jesus, A.; McBreen, F. Monitoring Guidance for Marine Benthic Habitats; JNCC Report No. 598; Joint Nature Conservation Committee: Peterborough, UK, 2017. [Google Scholar]
- Fabry, V.J.; McClintock, J.B.; Mathis, J.T.; Grebmeier, J.M. Ocean acidification at high latitudes: The bellweather. Oceanography 2009, 22, 160–171. [Google Scholar] [CrossRef]
- Reed, D.; Washburn, L.; Rassweiler, A.; Miller, R.; Bell, T.; Harrer, S. Extreme warming challenges sentinel status of kelp forests as indicators of climate change. Nat. Commun. 2016, 7, 12757. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; Bettignies, T.D.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U. Research note: Salinity tolerance of arctic kelps from Spitsbergen. Phycol. Res. 2007, 55, 257–262. [Google Scholar] [CrossRef]
- Polis, G.A.; Hurd, S.D. Linking marine and terrestrial food webs: Allochthonous input from the ocean supports high secondary productivity on small islands and coastal land communities. Am. Nat. 1996, 147, 396–423. [Google Scholar] [CrossRef]
- Brand, M.; Fischer, P. Species composition and abundance of the shallow water fish community of Kongsfjorden, Svalbard. Polar Biol. 2016, 39, 2155–2167. [Google Scholar] [CrossRef]
- Kamenos, N.A.; Moore, P.G.; Hall-Spencer, J.M. Nursery-area function of maerl grounds for juvenile Queen scallops Aequipecten opercularis and other invertebrates. Mar. Ecol. Prog. Ser. 2004, 274, 183–189. [Google Scholar] [CrossRef]
- Steller, D.L.; Caceres-Martinez, C. Coralline algal rhodoliths enhance larval settlement and early growth of the Pacific Calico Scallop Argopecten ventricosus. Mar. Ecol. Prog. Ser. 2009, 396, 49–60. [Google Scholar] [CrossRef]
- Foster, M.S.; McConnico, L.C.; Lundsten, L.; Wadsworth, T.; Kimball, T.; Brooks, L.B.; Medina-Lopez, M.; Riosmena-Rodriguez, R.; Hernandez-Carmona, G.; Vasquez-Elizondo, R.M.; et al. Diversity and natural history of a Lithothamnion muelleri-Sargassum horridum community in the Gulf of California. Cienc. Mar. 2007, 33, 367–384. [Google Scholar] [CrossRef]
- Stuart, V.; Klump, D.W. Evidence for food-resource partitioning by kelp-bed filter feeders. Mar. Ecol. Prog. Ser. 1984, 16, 27–37. [Google Scholar] [CrossRef]
- Yesson, C.; Simon, P.; Chamshirova, I.; Gorham, T.; Turner, C.J.; Arboe, N.H.; Blicher, M.E.; Kemp, K.M. Community composition of epibenthic megafauna on the west Greenland shelf. Polar Biol. 2015, 38, 2085–2096. [Google Scholar] [CrossRef]
- Leinaas, H.P.; Christie, H. Effecs of removing sea urchins (Strongylocentrotus droebachiensis): Stability of the barren state and succession of kelp forest recovery in the east Atlantic. Oecologia 1996, 105, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Steller, D.L.; Riosmena-Rodriguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquat. Conserv. 2003, 13, S5–S20. [Google Scholar] [CrossRef]
- Hofmann, L.C.; Schoenrock, K.M.; Beer, D.D. Arctic coralline algae elevate surface pH and carbonate in the dark. Front. Plant Sci. 2018, 9, 1416. [Google Scholar] [CrossRef] [PubMed]
- Comeau, S.; Edmunds, P.J.; Lantz, C.A.; Carpenter, R.C. Water flow modulates the response of coral reef communities to ocean acidification. Sci. Rep. 2014, 4, 6681. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Blake, C.; Berges, J.A.; Maggs, C.A. Environmental tolerances of free-living coralline algae (maerl): Importance for European marine conservation. Biol. Conserv. 2004, 120, 279–289. [Google Scholar] [CrossRef]
- Roberts, R.D.; Kühl, M.; Glud, R.N.; Rysgaard, S. Primary production of crustose coralline red algae in a high Arctic fjord. J. Phycol. 2002, 38, 273–283. [Google Scholar] [CrossRef]
- Adey, W.H.; Hayek, L.C. Elucidating marine biogeography with macrophytes: Quantitative analysis of the North Atlantic supports the thermogeographic model and demonstrates a distinct subarctic region in the northwest Atlantic. Northeast Nat. 2011, 18, 1–128. [Google Scholar] [CrossRef]
- Teichert, S.; Woelkerling, W.; Rüggeberg, A.; Wisshak, M.; Piepenburg, D.; Meyerhöfer, M.; Form, A.; Büdenbender, J.; Freiwald, A. Rhodolith beds (Corallinales, Rhodophyta) and their physical and biological environment at 80°31′ N in Nordkappbukta (Nordaustlandet, Svalbard Archipelago, Norway). Phycologia 2012, 51, 371–390. [Google Scholar] [CrossRef]
- Johnson, C.R.; Mann, K.H. The crustose coralline alga, Phymatolithon foslie, inhibits the overgrowth of seaweeds without relying on herbivores. J. Exp. Mar. Biol. Ecol. 1986, 96, 127–146. [Google Scholar] [CrossRef]
- Bulleri, F.; Bertocci, I.; Micheli, F. Interplay of encrusting coralline algae and sea urchins in maintaining alternative habitats. Mar. Ecol. Prog. Ser. 2002, 243, 101–109. [Google Scholar] [CrossRef]
- Muth, A. (University of Texas at Austin, Port Aransas, TX, USA). Personal observation, 2018.
- Burek, K.E.; Brien, J.M.; Scheibling, R.E. Wasted effort: Recruitment and persistence of kelp on algal turf. Mar. Ecol. Prog. Ser. 2018, 600, 3–19. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C. Ocean acidification causes ecosystem shifts via altered competetive interactions. Nat. Clim. Chang. 2013, 3, 156–159. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Wernberg, T. Rise of turfs: A new battlefront for globally declining kelp forests. Bioscience 2018, 68, 64–76. [Google Scholar] [CrossRef]
- Voronkov, A.; Hop, H.; Gulliksen, B. Diversity of hard-bottom fauna relative to environmental gradients in Kongsfjorden, Svalbard. Polar Res. 2013, 32, 11208. [Google Scholar] [CrossRef][Green Version]
- Paar, M.; Voronkov, A.; Hop, H.; Brey, T.; Bartsch, I.; Schwanitz, M.; Wiencke, C. Temporal shift in biomass and production of macrozoobenthos in the macroalgal belt at Hansneset, Kongsfjorden, after 15 years. Polar Biol. 2016, 39, 2065–2076. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Site Name | Habitat | Sediment Level | Latitude | Longitude | Depth (m) |
|---|---|---|---|---|---|
| Colony Bay | kelp | Medium | 64°10′34.76″ N | 51°44′58.57″ W | 9.1 |
| Site 153 | kelp | High | 64°10′45.24″ N | 51°53′27.61″ W | 5.2 |
| Site 4 | kelp | Low | 64°7′19.96″ N | 51°56′49.34″ W | 6.4 |
| Bird Island | kelp | Medium | 64°11′16.91″ N | 51°54′24.52″ W | 8.7 |
| Site 124 | rhodolith | Low | 64°8′17.37″ N | 51°55′55.62″ W | 4.6 |
| Site 121 | rhodolith | Low | 64°8′3.11″ N | 51°57′5.48″ W | 5.2 |
| Site 14 | rhodolith | Low | 64°7′38.51″ N | 51°42′21.30″ W | 5.4 |
| Site 160 | rhodolith | Low | 64°7′39.31″ N | 51°47′56.82″ W | 5.3 |
| Grabs | Surveys | Quadrats | ||||
|---|---|---|---|---|---|---|
| Rhodolith Bed | Kelp Forest | Rhodolith Bed | Kelp Forest | Rhodolith Bed | Kelp Forest | |
| Algae | ||||||
| filamentous algae (A) | ||||||
| Rhodophyta | ||||||
| Rhodolith (A) | ||||||
| CCA * (A) | ||||||
| Porphyra spp. (A) | ||||||
| Palmaria palmata (A) | ||||||
| Phaeophyta | ||||||
| kelp drift (A) | ||||||
| Saccharina longicruis (A) | ||||||
| Agarum clathratum (A) | ||||||
| Alaria esculenta (A) | ||||||
| Desmarestia aculeata (A) | ||||||
| Chlorophyta | ||||||
| Ulva spp. (A) | ||||||
| Animalia | ||||||
| Porifera | ||||||
| Haliclona sp. (F, S) | ||||||
| Mycale sp. (F, S) | ||||||
| Myxilla sp. (F, S) | ||||||
| Reniera rufescens (F, S) | ||||||
| Mollusca | ||||||
| Class Bivalvia | ||||||
| Hiatella arctica (F) | ||||||
| Mytilus spp. (F) | ||||||
| Macoma calcarea (Dep) | ||||||
| Ennucula tenuis (Det) | ||||||
| Chlamys islandica (F) | ||||||
| Class Gastropoda | ||||||
| Siphonodentalium lobatum (Dep) | ||||||
| Gibbula magus (G, Dep) | ||||||
| Gibbula umbilicalis (G, Dep) | ||||||
| Littorina saxatilis (G) | ||||||
| Cryptonatica affinis (P) | ||||||
| Natica clausa (P) | ||||||
| Margarites helicinus (G, Dep) | ||||||
| Margarites sp. (G, Dep) | ||||||
| Tonicella rubra (G) | ||||||
| Tectura testudinalis (G) | ||||||
| Dendronotus frondosus (P) | ||||||
| Echinodermata | ||||||
| Asterias rubens (S) | ||||||
| Psolus fabricii (S) | ||||||
| Strongylocentrotus droebachiensis (P, Dep) | ||||||
| Ophiura sarsi (S) | ||||||
| Leptasterias polaris (S) | ||||||
| Sargatiogeton laceratus (S) | ||||||
| Cucumaria frondosa (S) | ||||||
| Annelida | ||||||
| Gattyana cirrosa (U) | ||||||
| Euchurian sp. (U) | ||||||
| Order Phoronida (F) | ||||||
| Phyllodoce groenlandica (P) | ||||||
| Nipponnemertes pulchra (U) | ||||||
| Arthropoda | ||||||
| Semibalanus balanoides (F) | ||||||
| Order Amphipoda (F, Det, S, P) | ||||||
| Hyas coarctatus (O) | ||||||
| Weyprechtia pinguis (U) | ||||||
| Acanthonotozoma inflatum (U) | ||||||
| Calanus sp. (U) | ||||||
| Order Mysida (U) | ||||||
| unknown micrograzers (G) | ||||||
| Bryozoa | ||||||
| Cellaria sp. (F) | ||||||
| Cnidaria | ||||||
| Bolinopsis infundibulum (P) | ||||||
| Chordata | ||||||
| Gadus morhua (P) | ||||||
| Leptocottus armatus (P) | ||||||
| Myoxocephalus scorpius (P) |
| Swath Survey | Photo-Quadrat | Benthic Grabs | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Habitat | n | S | H’ | J | n | S | H’ | J | n | S | H’ | J |
| Kelp forest | ||||||||||||
| Colony Bay | 1 | 4 | 0.82 | 0.59 | 8 | 2.5 ± 0.19 | 0.66 ± 0.08 | 0.74 ± 0.08 | 5 | 6.6 ± 0.75 | 1.23 ± 0.11 | 0.66 ± 0.03 |
| Site 153 | 1 | 12 | 1.66 | 0.67 | 9 | 4.33 ± 0.67 | 1.01 ± 0.13 | 0.76 ± 0.04 | 5 | 5 ± 0.63 | 1.16 ± 0.06 | 0.75 ± 0.04 |
| Site 4 | 1 | 8 | 1.42 | 0.68 | 10 | 5.25 ± 0.49 | 1.37 ± 0.11 | 0.85 ± 0.03 | 5 | 8.4 ± 0.68 | 1.51 ± 0.18 | 0.71 ± 0.07 |
| Bird Island | 1 | 1 | 0 | 0 | 5 | 3 ± 0.45 | 0.62 ± 0.13 | 0.59 ± 0.06 | 5 | 4.8 ± 0.73 | 1.15 ± 0.19 | 0.74 ± 0.05 |
| Average | 6.25 ± 4.79 | 0.96 ± 0.36 | 0.49 ± 0.33 | 4.03 ± 1.85 | 1 ± 0.45 | 0.76 ± 0.16 | 6.2 ± 2.07 | 1.26 ± 0.33 | 0.71 ± 0.11 | |||
| Rhodolith Bed | ||||||||||||
| Site 124 | 1 | 4 | 0.17 | 0.12 | 10 | 3.2 ± 0.39 | 0.69 ± 0.14 | 0.58 ± 0.08 | 5 | 9.6 ± 1.69 | 2.06 ± 0.14 | 0.93 ± 0.01 |
| Site 121 | 1 | 6 | 0.71 | 0.4 | 9 | 2.67 ± 0.24 | 0.65 ± 0.1 | 0.68 ± 0.08 | 5 | 11.8 ± 0.8 | 2.15 ± 0.1 | 0.87 ± 0.02 |
| Site 14 | 1 | 5 | 0.11 | 0.07 | 10 | 3.7 ± 0.3 | 0.88 ± 0.06 | 0.69 ± 0.04 | 4 | 14 ± 0.71 | 2.07 ± 0.09 | 0.78 ± 0.04 |
| Site 160 | 1 | 4 | 0.17 | 0.12 | 10 | 3.36 ± 0.2 | 0.78 ± 0.7 | 0.65 ± 0.04 | 4 | 10.25 ± 1.31 | 2 ± 0.17 | 0.87 ± 0.05 |
| Average | 4.75 ± 0.96 | 0.26 ± 0.18 | 0.18 ± 0.15 | 3.25 ± 0.95 | 0.75 ± 0.31 | 0.65 ± 0.19 | 11.33 ± 2.93 | 2.07 ± 0.26 | 0.87 ± 0.08 |
| Sample Method | Grabs | Swath | Photo-Quadrat | |||
|---|---|---|---|---|---|---|
| Rhodolith | Kelp | Rhodolith | Kelp | Rhodolith | Kelp | |
| Autotroph | 12% | 4% | 30% | 16% | 75% | 63% |
| Filter feeder | 12% | 12% | 10% | 16% | 8% | 13% |
| Deposit feeder | 6% | 8% | 0% | 0% | 0% | 0% |
| Detritivore | 3% | 4% | 0% | 0% | 0% | 0% |
| Suspension feeder | 12% | 8% | 10% | 11% | 0% | 6% |
| Grazer | 9% | 4% | 0% | 0% | 0% | 0% |
| Omnivore | 3% | 0% | 0% | 0% | 0% | 0% |
| Predator | 9% | 16% | 30% | 26% | 0% | 6% |
| Mixed | 18% | 20% | 20% | 32% | 17% | 13% |
| Unknown | 15% | 24% | 0% | 0% | 0% | 0% |
| Site | DW100 | % DW550 | % DWCO3 | CaCO3 (g) m−3 | |
|---|---|---|---|---|---|
| site 124 | dead | 9.55 ± 4.24 | 5.03 ± 0.75 | 40.6 ± 19.7 | 28.06 ± 5.02 |
| 95% | live | 214.56 ± 43.91 | 4.98 ± 0.63 | 34 ± 9 | |
| site 121 | dead | 32.63 ± 8.73 | 2.34 ± 0.42 | 61.9 ± 15.9 | 72.09 ± 12.99 |
| 90% | live | 288.49 ± 24.24 | 4.56 ± 0.25 | 52.3 ± 7.3 | |
| site 14 | dead | 18.44 ± 8.9 | 4.93 ± 1.46 | 43.2 ± 18.9 | 39.12 ± 23.16 |
| 92% | live | 211.18 ± 37.98 | 6.25 ± 0.44 | 79.6 ± 15.5 | |
| site 160 | dead | 30.53 ± 20.48 | 1.64 ± 0.18 | 48.8 ± 34 | 103.73 ± 34.37 |
| 93% | live | 420.89 ± 59.74 | 5.24 ± 0.84 | 54 ± 17.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoenrock, K.M.; Vad, J.; Muth, A.; Pearce, D.M.; Rea, B.R.; Schofield, J.E.; Kamenos, N.A. Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland. Diversity 2018, 10, 117. https://doi.org/10.3390/d10040117
Schoenrock KM, Vad J, Muth A, Pearce DM, Rea BR, Schofield JE, Kamenos NA. Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland. Diversity. 2018; 10(4):117. https://doi.org/10.3390/d10040117
Chicago/Turabian StyleSchoenrock, Kathryn M., Johanne Vad, Arley Muth, Danni M. Pearce, Brice R. Rea, J. Edward Schofield, and Nicholas A. Kamenos. 2018. "Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland" Diversity 10, no. 4: 117. https://doi.org/10.3390/d10040117
APA StyleSchoenrock, K. M., Vad, J., Muth, A., Pearce, D. M., Rea, B. R., Schofield, J. E., & Kamenos, N. A. (2018). Biodiversity of Kelp Forests and Coralline Algae Habitats in Southwestern Greenland. Diversity, 10(4), 117. https://doi.org/10.3390/d10040117