Changes in Species Composition of Birds and Declining Number of Breeding Territories over 40 Years in a Nature Conservation Area in Southwest Germany



Abstract
1. Introduction
2. Materials and Methods
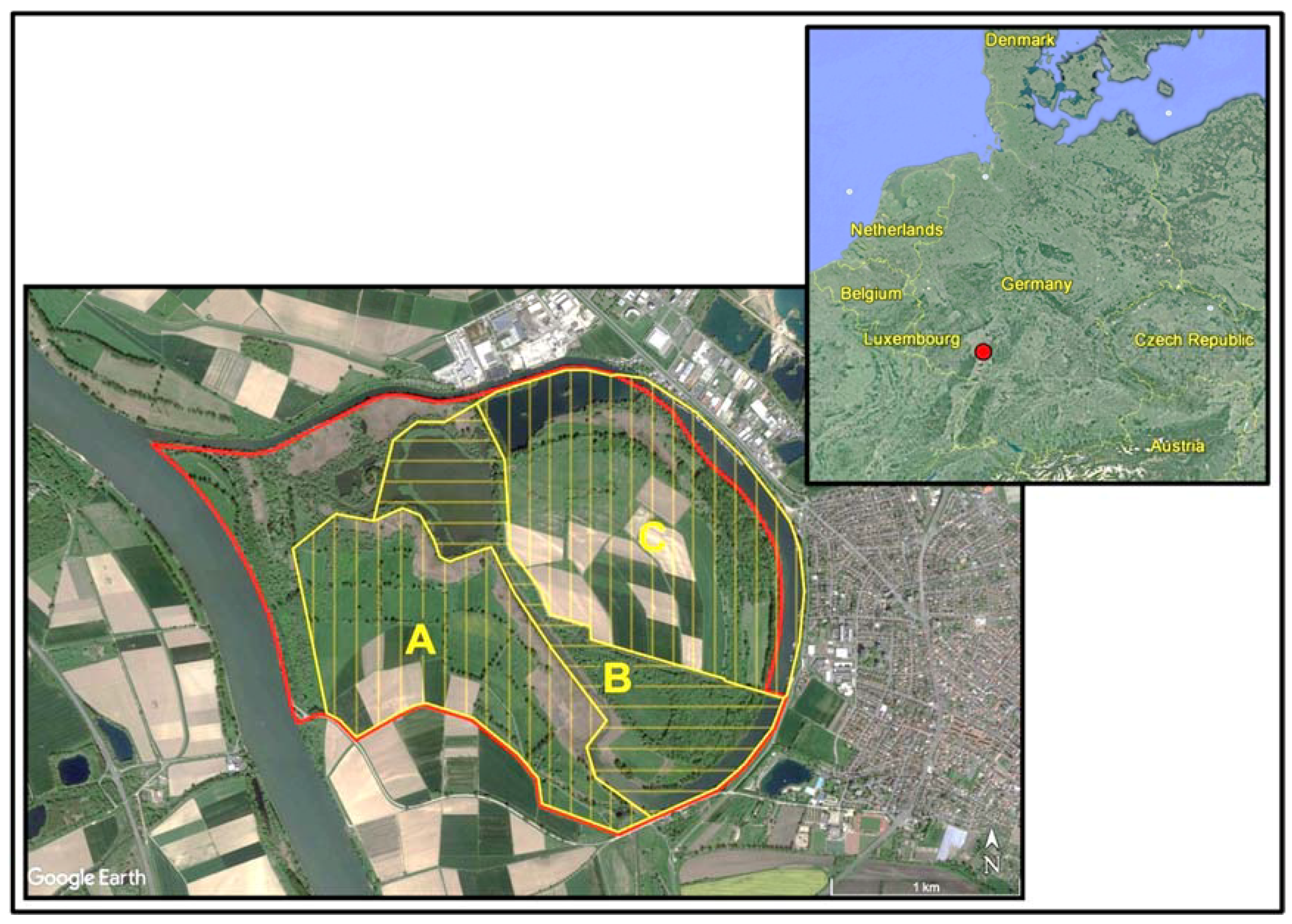
2.1. Study Area
2.2. Recording Breeding Birds
2.3. Data Analysis
2.4. Analysis of Changes in the Breeding Bird Community
3. Results
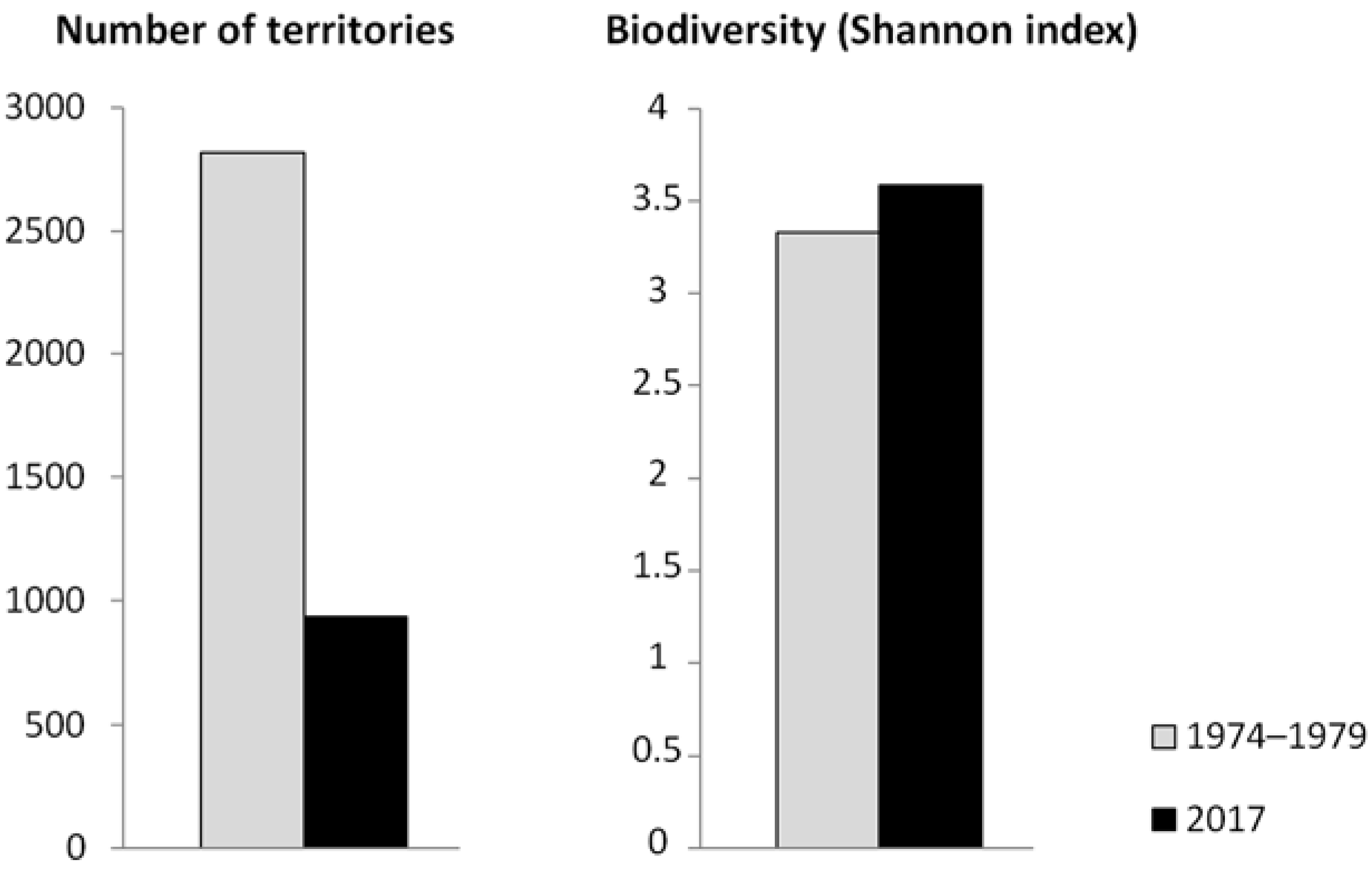
3.1. Abundance and Distribution of Breeding Birds
3.2. Development of the Breeding Bird Community
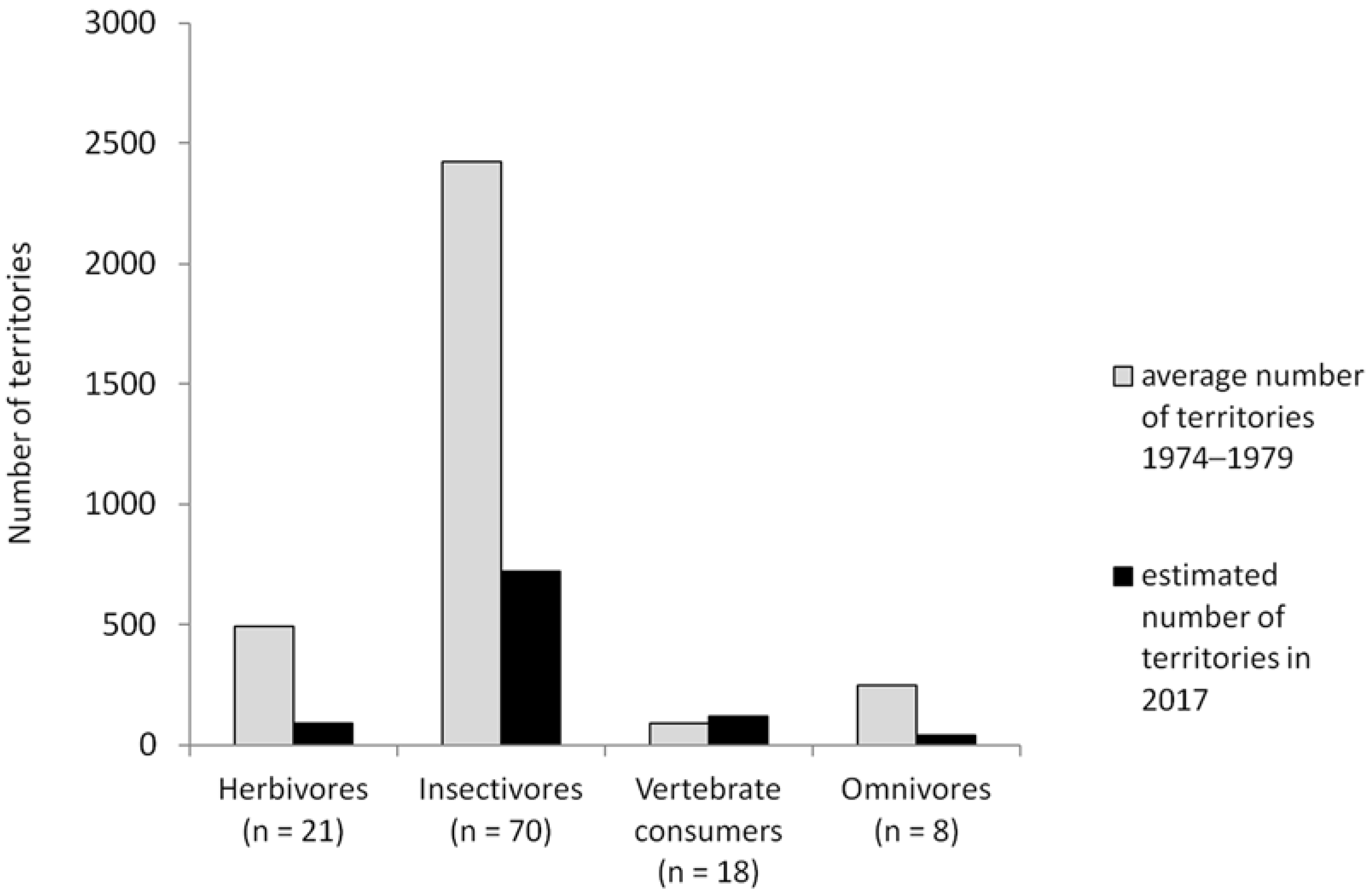
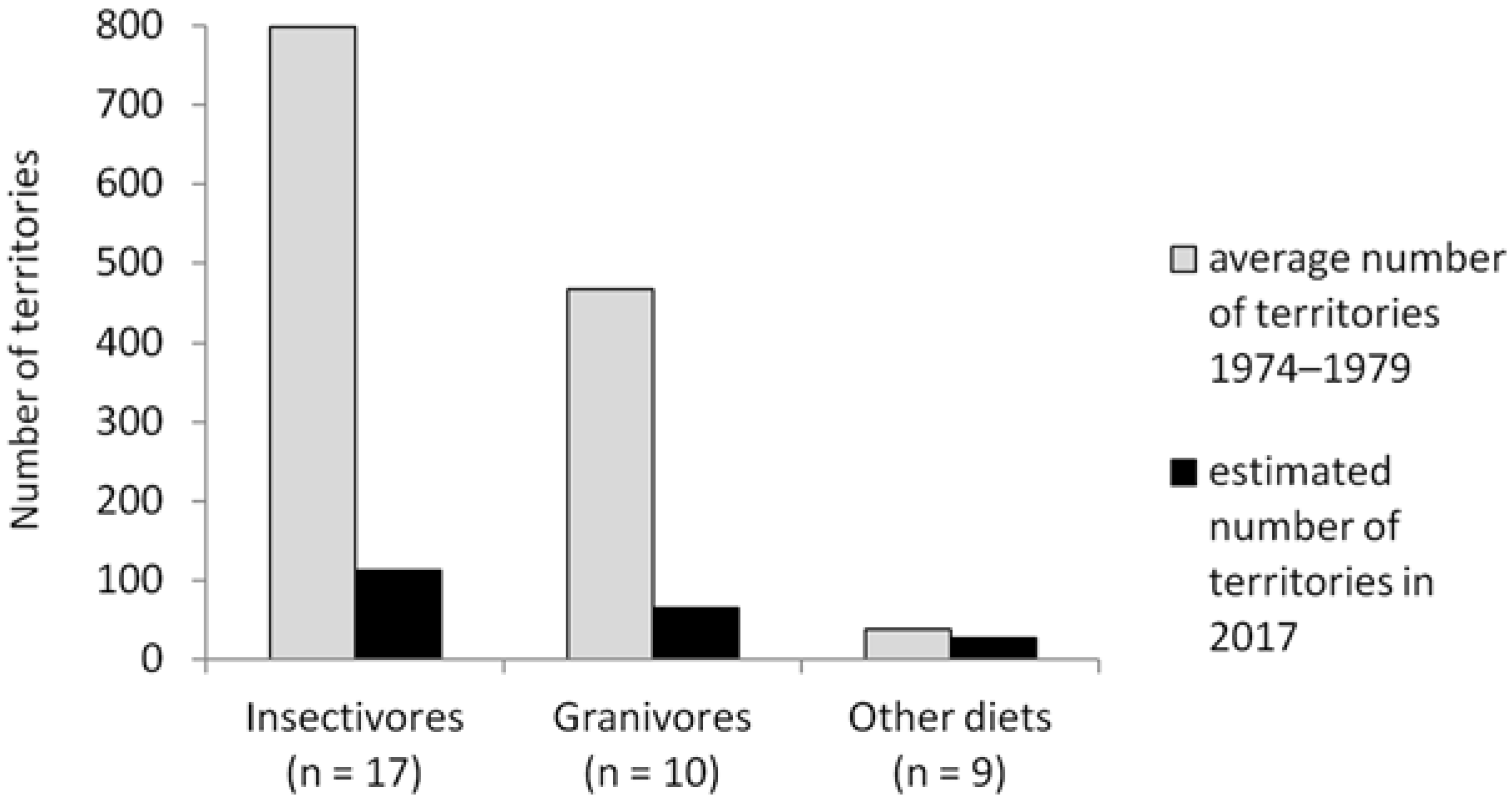
3.3. Population Trends in Relation to Food Availability
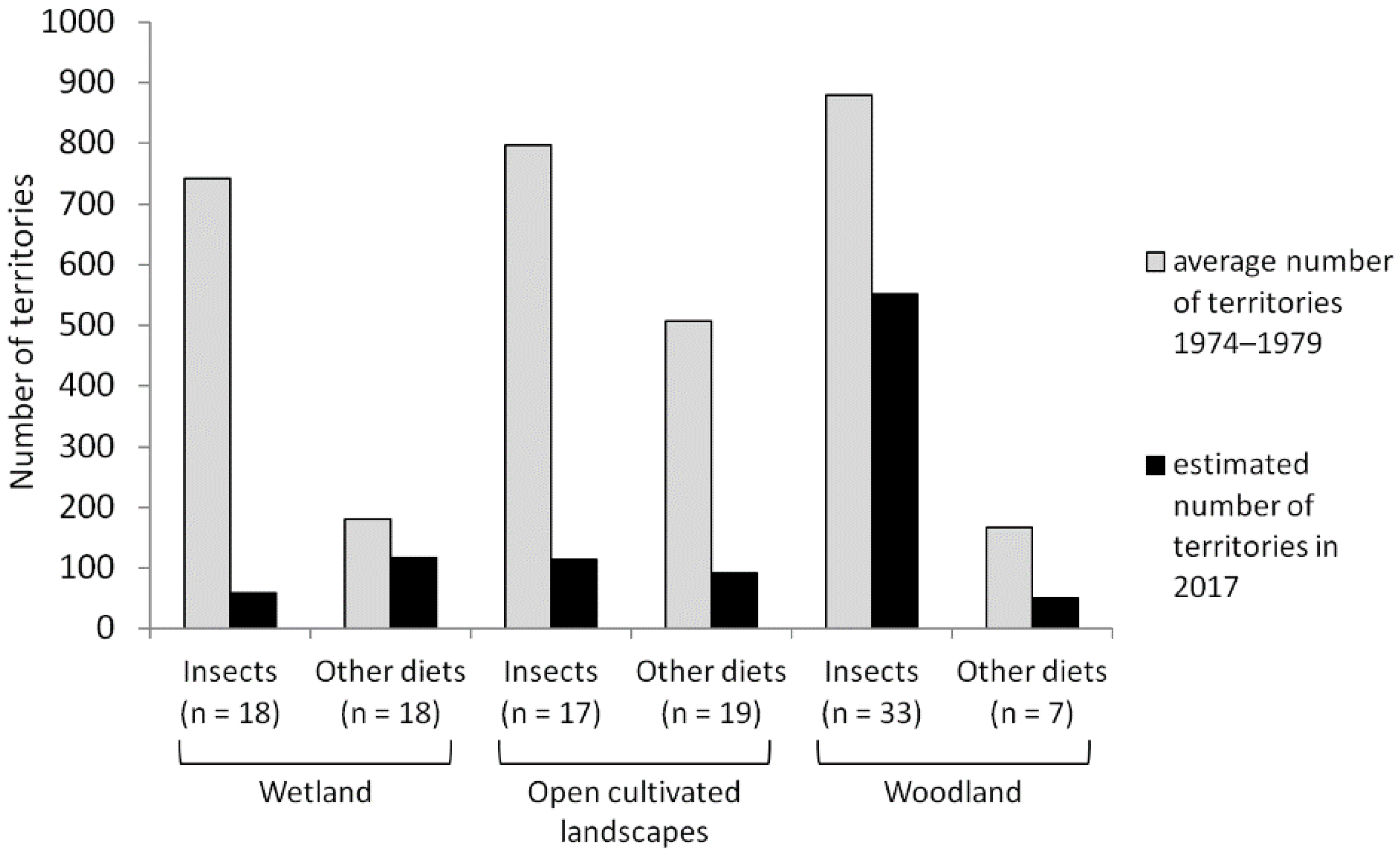
3.4. Population Trends in Relation to Habitat Utilisations
4. Discussion
4.1. Changes in the Breeding Bird Community
4.2. Population Trends in Relation to Food Availability
4.3. Population Trends in Relation to Habitat Utilisations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Butchart, S.H.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baille, J.E.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Rands, M.R.; Adams, W.M.; Bennun, L.; Butchart, S.H.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Gamero, A.; Brotons, L.; Brunner, A.; Foppen, R.; Fornasari, L.; Gregory, R.D.; Herrando, S.; Hořák, D.; Jiguet, F.; Kmecl, P.; et al. Tracking progress toward EU biodiversity strategy targets: EU policy effects in preserving its common farmland birds. Conserv. Lett. 2017, 10, 394–401. [Google Scholar] [CrossRef]
- Santana, J.; Reino, L.; Stoate, C.; Borralho, R.; Carvalho, C.R.; Schindler, S.; Moreira, F.; Bugalho, M.N.; Ribeiro, P.F.; Santos, J.L.; et al. Mixed effects of long-term conservation investment in Natura 2000 farmland. Conserv. Lett. 2014, 7, 467–477. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Brereton, T.M.; Botham, M.S.; Middlebrook, I.; Randle, Z.; Noble, D.; Roy, D.B. United Kingdom Butterfly Monitoring Scheme report for 2016; Centre for Ecology & Hydrology & Butterfly Conservation: Dorset, UK, 2017. [Google Scholar]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Wahl, J.; Dröschmeister, R.; Gerlach, B.; Grüneberg, C.; Langgemach, T.; Trautmann, S.; Sudfeldt, C. Vögel in Deutschland 2014; Dachverband Deutscher Avifaunisten e.V., Bundesamt für Naturschutz, Länderarbeitsgemeinschaft der Vogelschutzwarten: Münster, Germany, 2015; ISBN 978-3-9815543-5-9. [Google Scholar]
- Südbeck, P.; Andretzke, H.; Fischer, S.; Gedeon, K.; Schikore, T.; Schröder, K.; Sudfeldt, C. Methodenstandards zur Erfassung der Brutvögel Deutschlands; Länderarbeitsgemeinschaft der Vogelschutzwarten und Dachverband Deutscher Avifaunisten e.V.: Radolfzell, Germany, 2005; ISBN 3-00-015261-X. [Google Scholar]
- Schwarz, J.; Flade, M. Ergebnisse des DDA-Monitoringprogramms. Teil 1: Bestandsänderungen von Vogelarten der Siedlungen seit 1989. Die Vogelwelt 2000, 121, 87–106. [Google Scholar]
- Eppler, G.; Petermann, P. Grunddatenerhebung für das EU-Vogelschutzgebiet “Lampertheimer Altrhein” (6316-401); Regierungspräsidium Darmstadt: Darmstadt, Germany, 2012. [Google Scholar]
- Petermann, P. Die Vogelwelt. In 75 Jahre Naturschutzgebiet “Lampertheimer Altrhein”. Geschichte, Lebensräume und Arten einer Landschaft am Rhein, 2nd ed.; Regierungspräsidium Darmstadt: Darmstadt, Germany, 2012; pp. 11–14. [Google Scholar]
- Hagemeijer, E.J.; Blair, M.J. The EBCC Atlas of European Breeding Birds: Their Distribution and Abundance; T & A.D. Poyser: London, UK, 1997. [Google Scholar]
- Handke, K.; Handke, U. Ergebnisse sechsjähriger Brutvogelbestandsaufnahmen im NSG “Lampertheimer Altrhein”, Kr. Bergstraße (1974–1979). Vogel Umw. 1982, 2, 75–124. [Google Scholar]
- Eppler, G. Arten der Vogelschutzrichtlinie. In Grunddatenerfassung zum FFH-Gebiet “Lampertheimer Altrhein” (6316-401); Regierungspräsidium Darmstadt: Darmstadt, Germany, 2003; pp. 35–54. [Google Scholar]
- Bauer, H.-G.; Bezzel, E.; Fiedler, W. Das Kompendium der Vögel Mitteleuropas, 2nd ed.; AULA-Verlag: Wiebelsheim, Germany, 2012; ISBN 978-3-89104-758-3. [Google Scholar]
- Siegel, H. Bemerkenswerte Brutzeitbeobachtungen in Hessen 1985. Kormoran-Phalocrocorax carbo. Vogel Umw. 1985, 3, 368. [Google Scholar]
- Boatman, N.D.; Brickle, N.W.; Hart, J.D.; Milsom, T.P.; Morris, A.J.; Murray, A.W.; Murray, K.A.; Robertson, P.A. Evidence for the indirect effects of pesticides on farmland birds. IBIS 2004, 146, 131–143. [Google Scholar] [CrossRef]
- Jahn, T.; Hötker, H.; Oppermann, R.; Bleil, R.; Vele, L. Protection of Biodiversity of Free Living Birds and Mammals in Respect of the Effects of Pesticides; Umweltbundesamt: Dessau-Roßlau, Germany, 2014. [Google Scholar]
- Rutschke, E. Zur Dynamik und Funktion von Vogelrevieren. Ann. Naturhist. Mus. Wien 1986, 88/89, 171–180. [Google Scholar]
- Stenger, J. Food habits and available food of ovenbirds in relation to territory size. Auk 1958, 75, 335–346. [Google Scholar]
- Adams, E.S. Approaches to the Study of Territory Size and Shape. Annu. Rev. Ecol. Syst. 2001, 32, 277–303. [Google Scholar] [CrossRef]
- Van Dijk, T.C.; van Staalduinen, M.A.; van der Sluijs, J.P. Macro-invertebrate decline in surface water polluted with imidacloprid. PLoS ONE 2013, 8, e62374. [Google Scholar] [CrossRef] [PubMed]
- Petermann, P. Das NSG Lampertheimer Altrhein-ein Jahr nach den Eingriffen in den Wasserhaushalt-eine erste Bilanz. Collurio 2015, 31, 1–21. [Google Scholar]
- Silber, A. Die Geschichte der Naturschutzgebietsverordnung. In 75 Jahre Naturschutzgebiet “Lampertheimer Altrhein”. Geschichte, Lebensräume und Arten einer Landschaft am Rhein, 2nd ed.; Regierungspräsidium Darmstadt: Darmstadt, Germany, 2012; pp. 8–9. [Google Scholar]
- Wink, M.; Dietzen, C.; Gießing, B. Allgemeine Ergebnisse zur Vogelverbreitung im Rheinland (Nordrhein). In Die Vögel des Rheinlandes (Nordrhein). Ein Atlas der Brut- und Wintervogelverbreitung 1990 bis 2000. Beiträge zur Avifauna Nordrhein-Westfalens; Wink, M., Dietzen, C., Gießing, B., Eds.; Romneya Verlag: Dossenheim, Germany, 2005; Volume 36, pp. 17–31. ISBN 3-931921-07-7. [Google Scholar]
- Wretenberg, J.; Pärt, T.; Berg, Å. Changes in local species richness of farmland birds in relation to land-use changes and landscape structure. Biol. Conserv. 2010, 143, 375–381. [Google Scholar] [CrossRef]
- Berg, A.; Wretenberg, J.; Zmihorski, M.; Hiron, M.; Pärt, T. Linking occurrence and changes in local abundance of farmland bird species to landscape composition and land-use changes. Agric. Ecosyst. Environ. 2015, 204, 1–7. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Foppen, R.P.; van Turnhout, C.A.; de Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 2014, 511, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.H.; Avery, M.L.; Donald, P.; Evans, A.D.; Greem, R.E.; Wilson, J.D. A Review of the Indirect Effects of Pesticides on Birds; JNCC Report. No. 227; Joint Nature Conservation Committee: Peterborough, UK, 1997. [Google Scholar]
- Kluge, W. Die Waldbewirtschaftung. In 75 Jahre Naturschutzgebiet “Lampertheimer Altrhein”. Geschichte, Lebensräume und Arten einer Landschaft am Rhein, 2nd ed.; Regierungspräsidium Darmstadt: Darmstadt, Germany, 2012; pp. 18–20. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name (IOC World Bird List) 1 | Scientific Name (IOC World Bird List) 1 | Number of Confirmed Territories | Abundance Total Study Area (Territories/10 ha) | Abundance Suitable Habitat (Territories/10 ha) |
|---|---|---|---|---|
| Grey Partridge | Perdix perdix | 1 | 0.02 | 0.15 |
| Common Pheasant | Phasianus colchicus | 10 | 0.22 | 0.61 |
| Canada Goose | Branta canadensis | 1 | 0.02 | 0.15 |
| Greylag Goose | Anser anser | 2 | 0.04 | 0.31 |
| Mute Swan | Cygnus olor | 5 | 0.11 | 0.76 |
| Egyptian Goose | Alopochen aegyptiaca | 5 | 0.11 | 0.76 |
| Gadwall | Mareca strepera | 1 | 0.02 | 0.15 |
| Mallard | Anas platyrhynchos | 6 | 0.13 | 0.92 |
| Common Cuckoo | Cuculus canorus | 7 | 0.16 | 0.31 |
| Stock Dove | Columba oenas | 4 | 0.09 | 0.28 |
| Common Wood Pigeon | Columba palumbus | 17 | 0.38 | 1.07 |
| European Turtle Dove | Streptopelia turtur | 1 | 0.02 | 0.11 |
| Eurasian Coot | Fulica atra | 2 | 0.04 | 0.31 |
| Great Crested Grebe | Podiceps cristatus | 4 | 0.09 | 0.61 |
| Great Cormorant | Phalacrocorax carbo | 71 | 1.58 | - |
| Western Marsh Harrier | Circus aeruginosus | 1 | 0.02 | - |
| Red Kite | Milvus milvus | 1 | 0.02 | - |
| Black Kite | Milvus migrans | 3 | 0.07 | - |
| Common Buzzard | Buteo buteo | 3 | 0.07 | - |
| Tawny Owl | Strix aluco | 4 | 0.09 | - |
| Common Kingfisher | Alcedo atthis | 1 | 0.02 | 0.15 |
| Middle Spotted Woodpecker | Dendrocoptes medius | 5 | 0.11 | 0.54 |
| Lesser Spotted Woodpecker | Dryobates minor | 3 | 0.07 | 0.33 |
| Great Spotted Woodpecker | Dendrocopos major | 20 | 0.44 | 1.00 |
| Black Woodpecker | Dryocopus martius | 3 | 0.07 | 0.33 |
| Grey-headed Woodpecker | Picus canus | 4 | 0.09 | 0.27 |
| European Green Woodpecker | Picus viridis | 11 | 0.24 | 0.73 |
| Common Kestrel | Falco tinnunculus | 2 | 0.04 | - |
| Eurasian Hobby | Falco subbuteo | 1 | 0.02 | - |
| Eurasian Golden Oriole | Oriolus oriolus | 8 | 0.18 | 0.40 |
| Red-backed Shrike | Lanius collurio | 11 | 0.24 | 0.81 |
| Eurasian Jay | Garrulus glandarius | 9 | 0.20 | 0.40 |
| Carrion Crow | Corvus corone | 2 | 0.04 | 0.22 |
| Marsh Tit | Poecile palustris | 8 | 0.18 | 0.68 |
| Eurasian Blue Tit | Cyanistes caeruleus | 41 | 0.91 | 1.80 |
| Great Tit | Parus major | 61 | 1.36 | 3.07 |
| Eurasian Skylark | Alauda arvensis | 8 | 0.18 | 0.81 |
| Long-tailed Tit | Aegithalos caudatus | 10 | 0.22 | 0.57 |
| Common Chiffchaff | Phylloscopus collybita | 42 | 0.93 | 1.85 |
| Eurasian Reed Warbler | Acrocephalus scirpaceus | 36 | 0.80 | 5.94 |
| Icterine Warbler | Hippolais icterina | 3 | 0.07 | 0.28 |
| Eurasian Blackcap | Sylvia atricapilla | 61 | 1.36 | 2.38 |
| Garden Warbler | Sylvia borin | 10 | 0.22 | 0.44 |
| Common Whitethroat | Sylvia communis | 10 | 0.22 | 0.39 |
| Eurasian Wren | Troglodytes troglodytes | 14 | 0.31 | 1.52 |
| Eurasian Nuthatch | Sitta europaea | 26 | 0.58 | 1.73 |
| Eurasian Treecreeper | Certhia familiaris | 2 | 0.04 | 0.22 |
| Short-toed Treecreeper | Certhia brachydactyla | 22 | 0.49 | 1.54 |
| Common Starling | Sturnus vulgaris | 23 | 0.51 | 1.14 |
| Common Blackbird | Turdus merula | 19 | 0.42 | 1.12 |
| Song Thrush | Turdus philomelos | 16 | 0.36 | 1.63 |
| Spotted Flycatcher | Muscicapa striata | 9 | 0.20 | 0.45 |
| European Robin | Erithacus rubecula | 9 | 0.20 | 0.98 |
| Bluethroat | Luscinia svecica | 6 | 0.13 | 1.08 |
| Common Nightingale | Luscinia megarhynchos | 15 | 0.33 | 0.66 |
| European Pied Flycatcher | Ficedula hypoleuca | 1 | 0.02 | 0.11 |
| Dunnock | Prunella modularis | 5 | 0.11 | 0.22 |
| Eurasian Tree Sparrow | Passer montanus | 9 | 0.20 | 0.83 |
| Western Yellow Wagtail | Motacilla flava flava | 11 | 0.24 | 1.12 |
| Common Chaffinch | Fringilla coelebs | 51 | 1.13 | 2.54 |
| Hawfinch | Coccothraustes coccothraustes | 6 | 0.13 | 0.65 |
| European Greenfinch | Chloris chloris | 1 | 0.02 | 0.11 |
| European Goldfinch | Carduelis carduelis | 6 | 0.13 | 0.30 |
| European Serin | Serinus serinus | 1 | 0.02 | 0.11 |
| Yellowhammer | Emberiza citrinella | 20 | 0.44 | 0.78 |
| Common Reed Bunting | Emberiza schoeniclus | 3 | 0.07 | 0.54 |
| Common Name (IOC World Bird List) 1 | Scientific Name (IOC World Bird List) 1 | Investigation on Breeding Birds 1974–1979 (Number of Territories) | Investigation on Selected Species 2003 (Number of Territories) | Current Results 2017 (Number of Territories) | Long-Term Trend (%) | Short-Term Trend (%) | |
|---|---|---|---|---|---|---|---|
| Range of the Number of Territories | Average Number of Territories | ||||||
| Marsh Warbler | Acrocephalus palustris | 110 | 110 | n.r. | 0 | X | |
| Willow Warbler | Phylloscopus trochilus | 24–39 | 32 | n.r. | 0 | X | |
| Common Moorhen | Gallinula chloropus | 2–60 | 31.2 | n.r. | 0 | X | |
| Fieldfare | Turdus pilaris | 17–40 | 25.7 | n.r. | 0 | X | |
| Common Redstart | Phoenicurus phoenicurus | 6–24 | 14.1 | 6 | 0 | X | X |
| Tree Pipit | Anthus trivialis | 11–12 | 11.8 | n.r. | 0 | X | |
| Grey Heron | Ardea cinerea | 0–27 | 9.2 | 60 | 0 | X | X |
| Willow Tit | Poecile montanus | 7–14 | 8.8 | n.r. | 0 | X | |
| Little Owl | Athene noctua | 8–9 | 8.4 | 1 | 0 | X | X |
| Great Reed Warbler | Acrocephalus arundinaceus | 5–12 | 8.3 | 0 | 0 | X | |
| White Wagtail | Motacilla alba | 6 | 6 | n.r. | 0 | X | |
| Common Grasshopper Warbler | Locustella naevia | 1–15 | 5 | 2 | 0 | X | X |
| Water Rail | Rallus aquaticus | 0–9 | 4.9 | 0 | 0 | X | |
| Long-eared Owl | Asio otus | >3–5 | >4.3 | 2 | 0 | X | X |
| Eurasian Magpie | Pica pica | 1–5 | 3 | n.r. | 0 | X | |
| Little Grebe | Tachybaptus ruficollis | 0–5 | 2.8 | 0 | 0 | X | |
| Common Linnet | Linaria cannabina | 1–3 | 2.5 | n.r. | 0 | X | |
| Little Bittern | Ixobrychus minutus | 0–3 | 2.3 | 0 | 0 | X | |
| Sedge Warbler | Acrocephalus schoenobaenus | 1–5 | 1.9 | n.r. | 0 | X | |
| Corn Bunting | Emberiza calandra | 0–2 | 1.4 | n.r. | 0 | X | |
| Eurasian Collared Dove | Streptopelia decaocto | 1–2 | 1.2 | n.r. | 0 | X | |
| Western Jackdaw | Coloeus monedula | 1 | 1 | n.r. | 0 | X | |
| Northern Lapwing | Vanellus vanellus | 0–3 | 1 | n.r. | 0 | X | |
| Black-crowned Night Heron | Nycticorax nycticorax | 0–3 | 0.9 | 0 | 0 | X | |
| Purple Heron | Ardea purpurea | 0–3 | 0.9 | 0 | 0 | X | |
| Savi’s Warbler | Locustella luscinioides | 0–5 | 0.8 | n.r. | 0 | X | |
| Lesser Whitethroat | Sylvia curruca | 0–2 | 0.7 | n.r. | 0 | X | |
| Wood Warbler | Phylloscopus sibilatrix | 0–2 | 0.7 | n.r. | 0 | X | |
| House Sparrow | Passer domesticus | 0–2 | 0.5 | n.r. | 0 | X | |
| Little Crake | Porzana parva | 0–2 | 0.5 | n.r. | 0 | X | |
| Corn Crake | Crex crex | 0–2 | 0.4 | n.r. | 0 | X | |
| Little Ringed Plover | Charadrius dubius | 0–1 | 0.3 | n.r. | 0 | X | |
| River Warbler | Locustella fluviatilis | 0–2 | 0.3 | n.r. | 0 | X | |
| Garganey | Spatula querquedula | 0–1 | 0.2 | 0 | 0 | X | |
| Swan Goose | Anser cygnoides | 0 | 0 | 1 | 0 | X | |
| Yellow-legged Gull | Larus michahellis | 0 | 0 | 1 | 0 | X | |
| Common Reed Bunting | Emberiza schoeniclus | ca. 250 | ca. 250 | n.r. | 3 | −99 | |
| Eurasian Coot | Fulica atra | 2–106 | 59.6 | n.r. | 2 | −97 | |
| Eurasian Tree Sparrow | Passer montanus | ca. 350 | ca. 350 | n.r. | 9 | −97 | |
| European Turtle Dove | Streptopelia turtur | 29–45 | 37.2 | 11 | 1 | −97 | −91 |
| European Greenfinch | Chloris chloris | 19–27 | 23.3 | n.r. | 1 | −96 | |
| Great Crested Grebe | Podiceps cristatus | 4–70 | 47.3 | 4–5 | 4 | −92 | −11 |
| Eurasian Reed Warbler | Acrocephalus scirpaceus | ca. 400 | ca. 400 | 180–200 | 36 | −91 | −81 |
| Common Starling | Sturnus vulgaris | ca. 220 | ca. 220 | n.r. | 23 | −90 | |
| Dunnock | Prunella modularis | 46–51 | 49 | n.r. | 5 | −90 | |
| Carrion Crow | Corvus corone | 14–19 | 16.5 | n.r. | 2 | −88 | |
| Common Blackbird | Turdus merula | 130 | 130 | n.r. | 19 | −85 | |
| Eurasian Wren | Troglodytes troglodytes | 64–75 | 69.5 | n.r. | 14 | −80 | |
| Eurasian Skylark | Alauda arvensis | 38 | 38 | 3–4 | 8 | −79 | +129 |
| Common Nightingale | Luscinia megarhynchos | 55–66 | 59.3 | n.r. | 15 | −75 | |
| Grey Partridge | Perdix perdix | 3–5 | 4 | 2 | 1 | −75 | −50 |
| Garden Warbler | Sylvia borin | 30–51 | 38.7 | n.r. | 10 | −74 | |
| European Robin | Erithacus rubecula | 28–37 | 32.8 | n.r. | 9 | −73 | |
| Mallard | Anas platyrhynchos | 16–31 | 22.2 | n.r. | 6 | −73 | |
| Bluethroat | Luscinia svecica | 12–31 | 20.6 | 21–30 | 6 | −71 | −76 |
| European Goldfinch | Carduelis carduelis | 16–20 | 18 | n.r. | 6 | −67 | |
| Icterine Warbler | Hippolais icterina | 8 | 8 | 11 | 3 | −63 | −73 |
| Common Wood Pigeon | Columba palumbus | 39 | 39 | n.r. | 17 | −56 | |
| Common Cuckoo | Cuculus canorus | 15 | 15 | n.r. | 7 | −53 | |
| Common Kestrel | Falco tinnunculus | 3–5 | 4.1 | n.r. | 2 | −51 | |
| Eurasian Golden Oriole | Oriolus oriolus | 14–20 | 16.3 | 17 | 8 | −51 | −53 |
| Song Thrush | Turdus philomelos | 23–39 | 29 | n.r. | 16 | −45 | |
| Eurasian Blackcap | Sylvia atricapilla | 95–120 | 107.5 | n.r. | 61 | −43 | |
| Eurasian Blue Tit | Cyanistes caeruleus | 70 | 70 | n.r. | 41 | −41 | |
| Common Chiffchaff | Phylloscopus collybita | 65–75 | 70 | n.r. | 42 | −40 | |
| Lesser Spotted Woodpecker | Dryobates minor | 3–7 | 4.6 | 2–3 | 3 | −35 | +20 |
| Short-toed Tree creeper | Certhia brachydactyla | 31–35 | 33 | n.r. | 22 | −33 | |
| Great Tit | Parus major | 90 | 90 | n.r. | 61 | −32 | |
| Common Whitethroat | Sylvia communis | 1–22 | 12.5 | 39 | 10 | −20 | −74 |
| Black Kite | Milvus migrans | 2–5 | 3.7 | 11–15 | 3 | −18 | −77 |
| Common Chaffinch | Fringilla coelebs | 48–55 | 51.5 | n.r. | 51 | −1 | |
| Common Buzzard | Buteo buteo | 2–4 | 3 | n.r. | 3 | ±0 | |
| Long-tailed Tit | Aegithalos caudatus | 6–14 | 9.5 | n.r. | 10 | +5 | |
| Grey-headed Woodpecker | Picus canus | 2–5 | 3.5 | 0–1 | 4 | +14 | * |
| Tawny Owl | Strix aluco | 2–4 | 3.2 | 3 | 4 | +25 | +33 |
| Yellowhammer | Emberiza citrinella | 14–16 | 15 | n.r. | 20 | +33 | |
| Spotted Flycatcher | Muscicapa striata | 1–13 | 5.3 | n.r. | 9 | +70 | |
| Great Spotted Woodpecker | Dendrocopos major | 10 | 10 | n.r. | 20 | +100 | |
| Eurasian Jay | Garrulus glandarius | 4–5 | 4.3 | n.r. | 9 | +108 | |
| European Green Woodpecker | Picus viridis | 3–5 | 4 | 2–4 | 11 | +175 | +267 |
| Eurasian Nuthatch | Sitta europaea | 6–12 | 8.3 | n.r. | 26 | +212 | |
| Western Yellow Wagtail | Motacilla flava flava | 0–7 | 2.3 | 3–4 | 11 | +371 | +214 |
| Common Kingfisher | Alcedo atthis | 0–1 | 0.2 | 1 | 1 | * | ±0 |
| European Pied Flycatcher | Ficedula hypoleuca | 0–1 | 0.2 | n.r. | 1 | * | |
| Gadwall | Mareca strepera | 0–1 | 0.2 | 0 | 1 | * | new |
| Western Marsh Harrier | Circus aeruginosus | 0–1 | 0.3 | 0 | 1 | * | new |
| Black Woodpecker | Dryocopus martius | 1–2 | 1.3 | 1–2 | 3 | * | * |
| Middle Spotted Woodpecker | Dendrocoptes medius | 1–2 | 1.5 | 4–5 | 5 | * | +11 |
| Mute Swan | Cygnus olor | 0–3 | 1.2 | n.r. | 5 | * | |
| Hawfinch | Coccothraustes coccothraustes | 1–2 | 1.4 | n.r. | 6 | * | |
| Canada Goose | Branta canadensis | 0 | 0 | 2 | 1 | new | −50 |
| Eurasian Hobby | Falco subbuteo | 0 | 0 | 1 | 1 | new | ±0 |
| European Serin | Serinus serinus | 0 | 0 | n.r. | 1 | new | |
| Red Kite | Milvus milvus | 0 | 0 | n.r. | 1 | new | |
| Eurasian Treecreeper | Certhia familiaris | 0 | 0 | n.r. | 2 | new | |
| Greylag Goose | Anser anser | 0 | 0 | 1 | 2 | new | +100 |
| Stock Dove | Columba oenas | 0 | 0 | 2 | 4 | new | +100 |
| Egyptian Goose | Alopochen aegyptiaca | 0 | 0 | n.r. | 5 | new | |
| Marsh Tit | Poecile palustris | 0 | 0 | n.r. | 8 | new | |
| Red-backed Shrike | Lanius collurio | 0 | 0 | 7 | 11 | new | +57 |
| Great Cormorant | Phalacrocorax carbo | 0 | 0 | 240 | 71 | new | −70 |
| Food 1 | Increase | Constant | Decrease |
|---|---|---|---|
| Herbivores (n = 21) | 43% | 0% | 57% |
| Insectivores (n = 70) | 20% | 6% | 74% |
| Vertebrate consumers (n = 18) | 33% | 17% | 50% |
| Omnivores (n = 8) | 13% | 0% | 88% |
| Habitat | Increase | Constant | Decrease |
|---|---|---|---|
| Wetland (n = 29) | 28% | 0% | 72% |
| Open cultivated landscapes (n = 27) | 22% | 11% | 67% |
| Woodland (n = 39) | 31% | 10% | 59% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrauth, F.E.; Wink, M. Changes in Species Composition of Birds and Declining Number of Breeding Territories over 40 Years in a Nature Conservation Area in Southwest Germany. Diversity 2018, 10, 97. https://doi.org/10.3390/d10030097
Schrauth FE, Wink M. Changes in Species Composition of Birds and Declining Number of Breeding Territories over 40 Years in a Nature Conservation Area in Southwest Germany. Diversity. 2018; 10(3):97. https://doi.org/10.3390/d10030097
Chicago/Turabian StyleSchrauth, Fabian Etienne, and Michael Wink. 2018. "Changes in Species Composition of Birds and Declining Number of Breeding Territories over 40 Years in a Nature Conservation Area in Southwest Germany" Diversity 10, no. 3: 97. https://doi.org/10.3390/d10030097
APA StyleSchrauth, F. E., & Wink, M. (2018). Changes in Species Composition of Birds and Declining Number of Breeding Territories over 40 Years in a Nature Conservation Area in Southwest Germany. Diversity, 10(3), 97. https://doi.org/10.3390/d10030097