Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions


 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
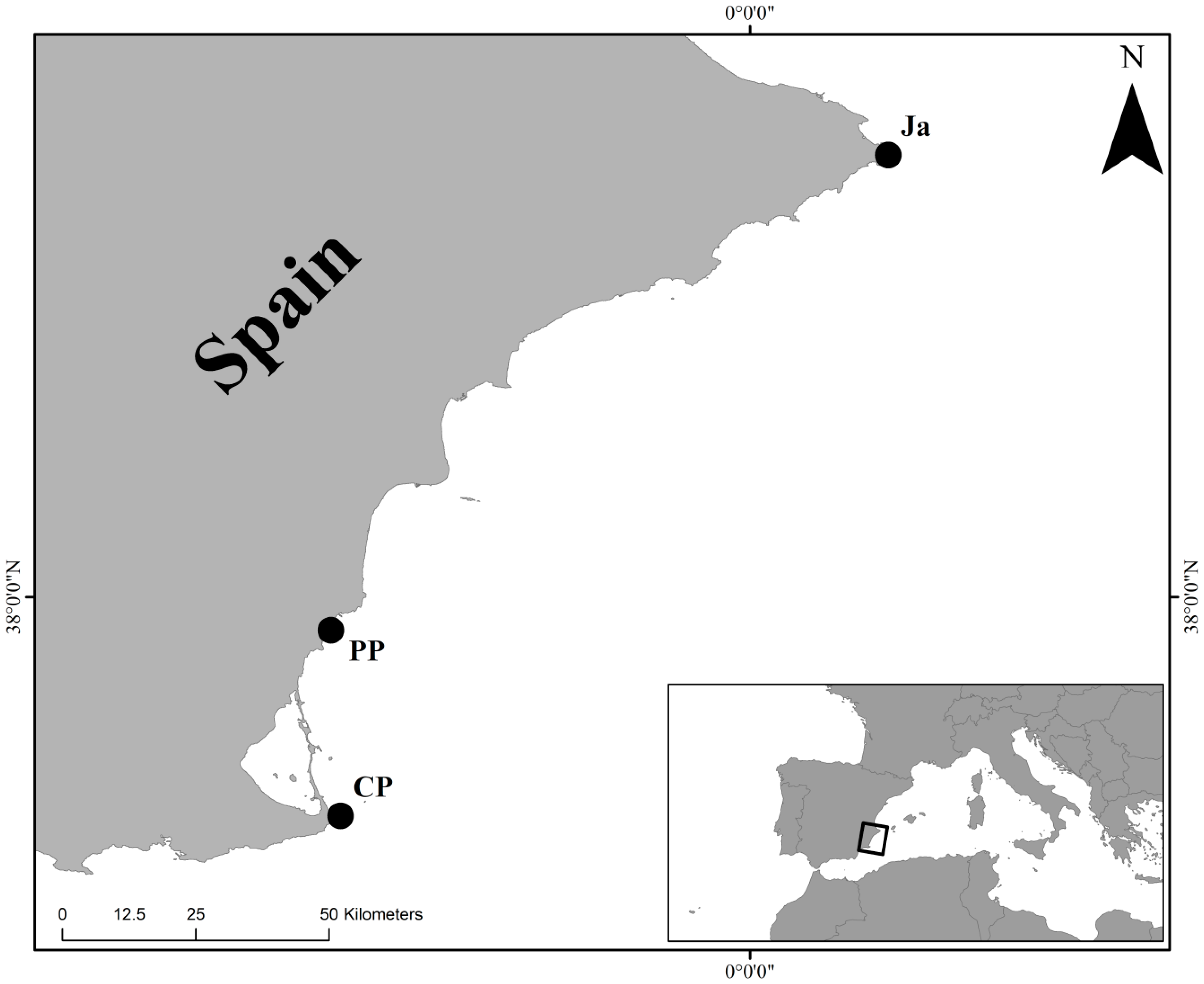
2.1. Sampling Design
2.2. Data Analysis
3. Results
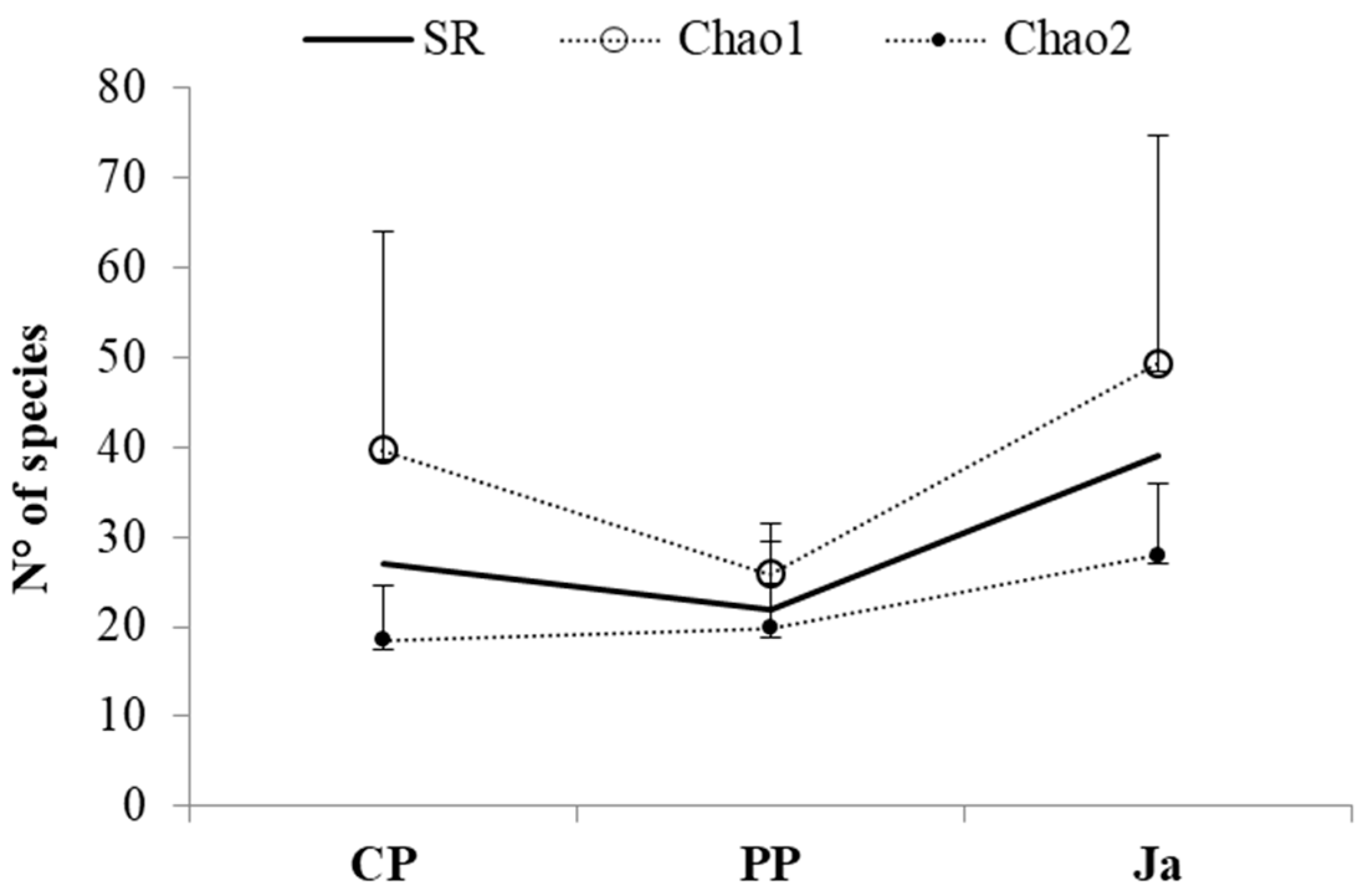
3.1. Composition of Mollusc Assemblages
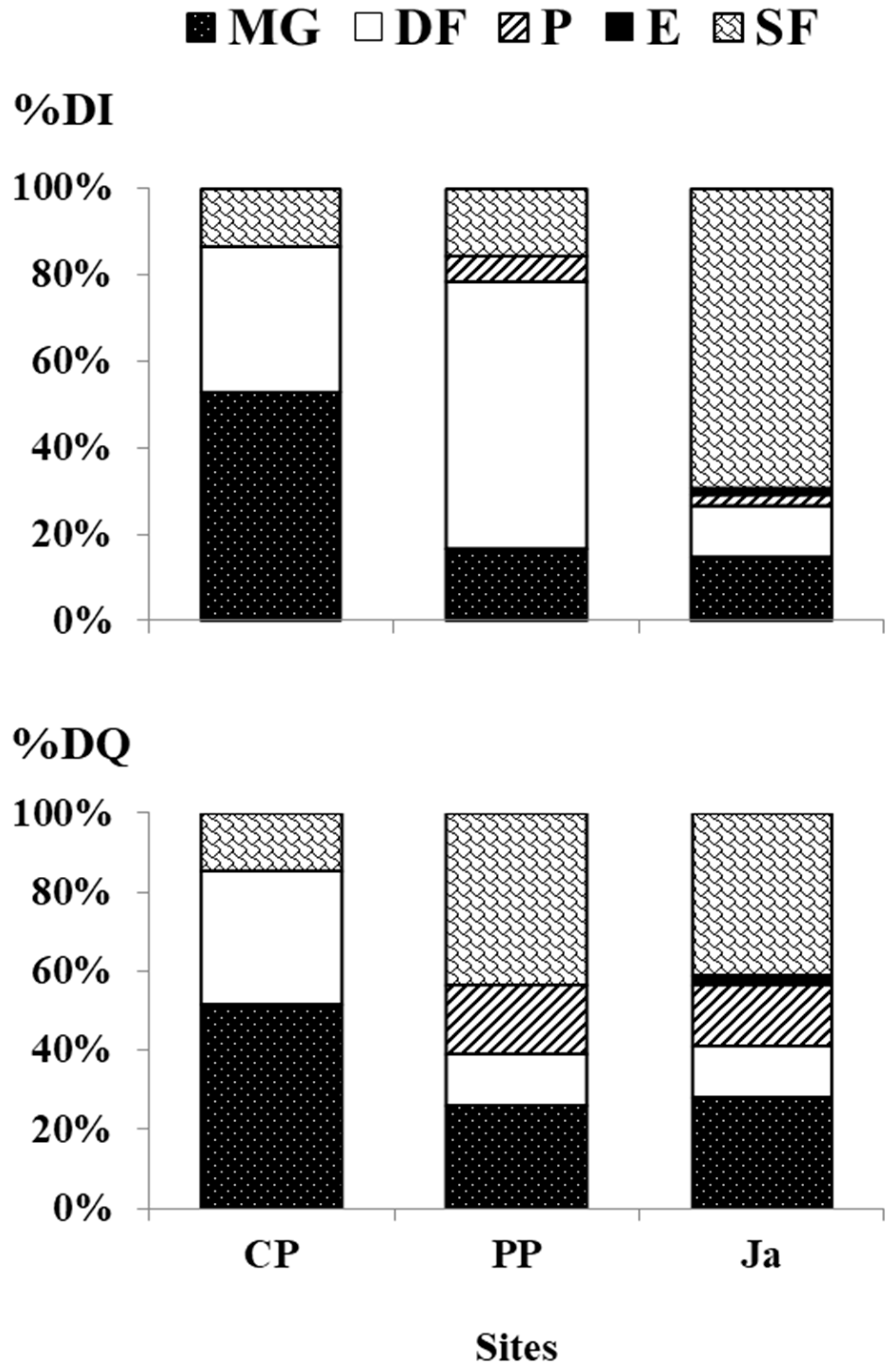
3.2. Trophic Diversity
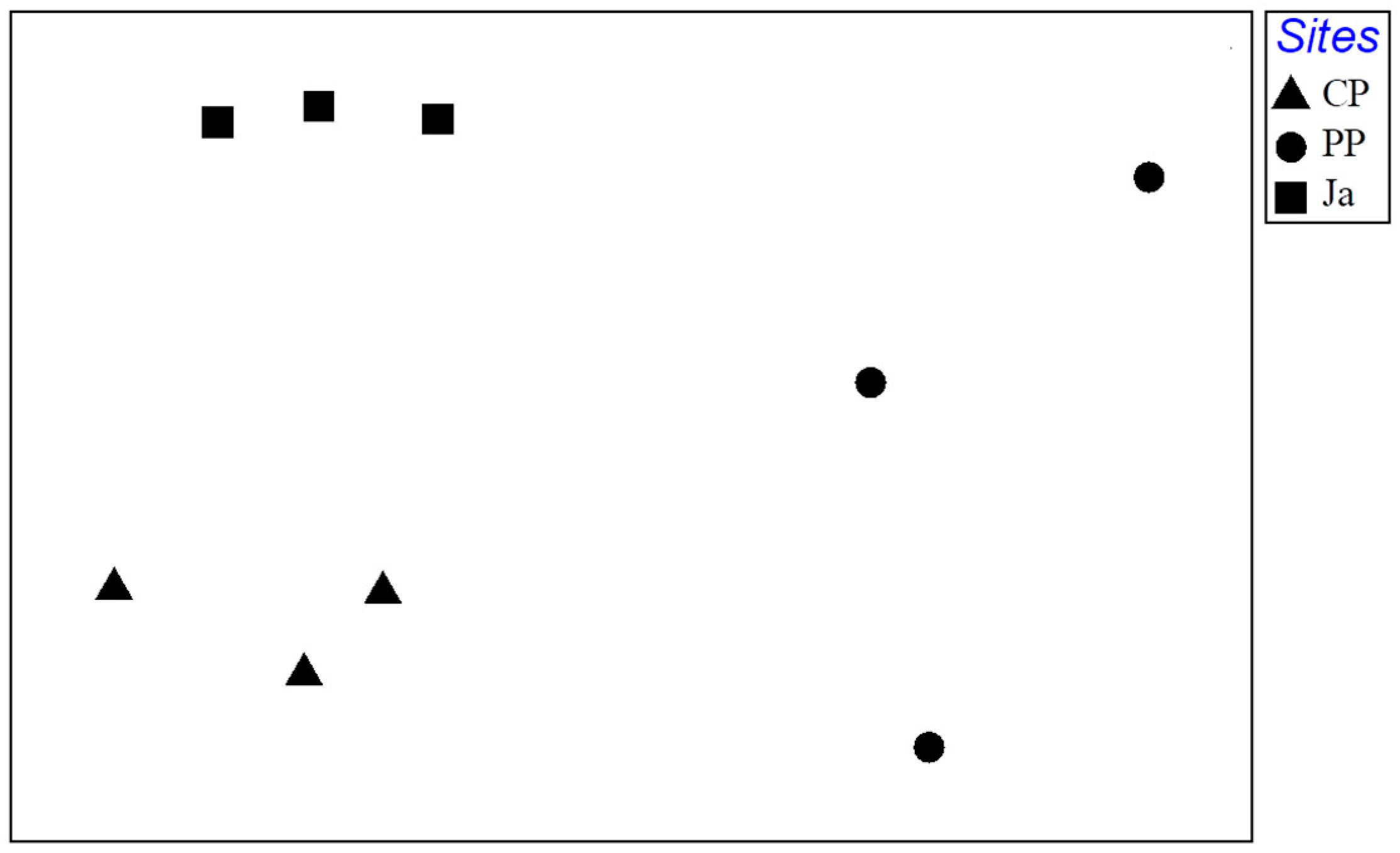
3.3. Structural Analysis of Mollusc Assemblage
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Danovaro, R. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Kružić, P. Bioconstructions in the Mediterranean: Present and Future. In The Mediterranean Sea; Goffredo, S., Dubinsky, Z., Eds.; Springer: Berlin, Germany, 2014; pp. 435–447. ISBN 978-94-007-6703-4. [Google Scholar]
- Antonioli, F.; Chemello, R.; Improta, S.; Riggio, S. Dendropoma lower intertidal reef formations and their palaeoclimatological significance NW Sicily. Mar. Geol. 1999, 161, 155–170. [Google Scholar] [CrossRef]
- Silenzi, S.; Antonioli, F.; Chemello, R. A new marker for sea surface temperature trend during the last centuries in temperate areas: Vermetid reef. Glob. Planet Chang. 2004, 40, 105–114. [Google Scholar] [CrossRef]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Di Stefano, F.; Russo, G.F. Morpho-structural and ecological features of a shallow vermetid bioconstruction in the Tyrrhenian Sea (Mediterranean Sea, Italy). J. Sea Res. 2018, 131, 61–68. [Google Scholar] [CrossRef]
- Ben Eliahu, M.N.; Safriel, U.N. A comparison between species diversities of polychaetes from tropical and temperate structurally similar rocky intertidal habitats. J. Biogeogr. 1982, 9, 371–390. [Google Scholar] [CrossRef]
- Pandolfo, A.; Chemello, R.; Riggio, S. Notes sur la signification écologique de la malacofaune d’un “Trottoir à Vermets” le long de la côte de Palerme (Sicile). Rapp. Comm. Int. Mer. Medit. 1992, 33, 47. [Google Scholar]
- Chemello, R.; Ciuna, I.; Pandolfo, A.; Riggio, S. Molluscan assemblages associated with intertidal vermetid formations: A morpho-functional approach. Boll. Malacol. 1998, 33, 105–114. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Menge, B.A.; Sutherland, J.P. Species diversity gradients: Synthesis of the roles of predation, competition, and temporal heterogeneity. Am. Nat. 1976, 110, 351–369. [Google Scholar] [CrossRef]
- Safriel, U.N.; Ben Eliahu, M.N. The influence of habitat structure and environmental stability on the species diversity of polychaetes in vermetid reefs. In Habitat Structure. The Physical Arrangement of Objects in Space; Bell, S.S., McCoy, E.D., Mushinsky, H.R., Eds.; Chapman and Hall, Ltd.: London, UK, 1991; pp. 349–369. ISBN 978-94-011-3076-9. [Google Scholar]
- Calvo, M.; Templado, J.; Penchaszadeh, P.E. Reproductive biology of the gregarious Mediterranean vermetid gastropod Dendropoma petraeum. J. Mar. Biol. Assoc. UK 1998, 78, 525–549. [Google Scholar] [CrossRef]
- D’Ancona Lunetta, G.; Damiani, F. Spermiogenesis in the vermetid gastropod Dendropoma petraeum (Gastropoda, Prosobranchia). Eur. J. Histochem. 2002, 46, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Klerman, A.; Fine, M.; Galil, B.S. Reproductive biology of a threatened reef building vermetid (Mollusca: Gastropoda) off the coast of Israel. Rapp. Comm. Int. Mer. Médit. 2004, 37, 7–11. [Google Scholar]
- Usvyatsov, S. Dendropoma petraeum (Monterosato, 1884): A Mediterranean species complex based on reproductive characteristics? In Proceedings of the 38th CIESM (the Mediterranean Science Commission) Congress, Istanbul, Turkey, 9–13 April 2007; p. 629. [Google Scholar]
- Templado, J.; Richter, A.; Calvo, M. Reef building Mediterranean vermetid gastropods: Disentangling the Dendropoma petraeum species complex. Mediterr. Mar. Sci. 2016, 17, 13–31. [Google Scholar] [CrossRef]
- Goren, M.; Galil, B.S. Fish biodiversity in the vermetid reef of Shiqmona (Israel). Mar. Ecol. 2001, 22, 369–378. [Google Scholar] [CrossRef]
- Mannino, A.M. Studio fitosociologico della vegetazione mesolitorale a Lithophyllum lichenoides PHILIPPI (Rhodophyceae, Corallinales). Nat. Sicil. 1992, 16, 3–25. [Google Scholar]
- Consoli, P.; Romeo, T.; Giongrandi, U.; Andaloro, F. Differences among fish assemblages associated with a nearshore vermetid reef and two other rocky habitats along the shores of Cape Milazzo (northern Sicily, central Mediterranean Sea). J. Mar. Biol. Assoc. UK 2008, 88, 401–410. [Google Scholar] [CrossRef]
- Pandolfo, A.; Chemello, R.; Riggio, S. Prime note sui popolamenti associati ai “trottoir” a vermetidi delle coste siciliane: I Molluschi. Oebalia 1992, 17, 379–382. [Google Scholar]
- Boronat, J.; Acuna, D.; Fresneda, M. Ensayo de caracterizacion malacologica de tres unidades bionomicas en las costas de Jávea (Alicante). Bol. Inst. Esp. Oceanogr. 1985, 2, 73–84. [Google Scholar]
- Fernández, M.T. Caracterización de las fitocenosis de las plataformas de abrasión con vermétidos del sureste ibérico. Ph.D. Thesis, University of Alicante, Alicante, Spain, 2014. [Google Scholar]
- Fernández, M.T.; Gómez, C.B.; Urrea, M.V.; Zubcoff, J.J.; Espla, A.R. The dynamics of phytobenthos and its main drivers on abrasion platforms with vermetids (Alicante, Southeastern Iberian Peninsula). Mediterr. Mar. Sci. 2018, 19, 58–68. [Google Scholar] [CrossRef]
- WoRMS Editorial Board—World Register of Marine Species. Available online: http://www.marinespecies.org at VLIZ (accessed on 19 June 2018).
- Purchon, R.D. The Biology of the Mollusca, 2nd ed.; Pergamon Press Ltd.: Oxford, UK, 1977; p. 587. ISBN 1483285421. [Google Scholar]
- Hughes, R.N. A Functional Biology of Marine Gastropods; Croom Helm: London, UK; Sydney, Australia, 1986; p. 245. ISBN 0 7099 3746 6. [Google Scholar]
- Fretter, V.; Graham, A. British Prosobranch Molluscs. Their Functional Anatomy and Ecology; The Ray Society: London, UK, 1994; ISBN 0-903874-23-7. [Google Scholar]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; p. 283. ISBN 1439839123. [Google Scholar]
- Gotelli, N.J.; Colwell, R.K. Estimating species richness. In Frontiers in Measuring Biodiversity; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 39–54. ISBN 9780199580675. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Terlizzi, A.; Anderson, M.J.; Fraschetti, S.; Benedetti-Cecchi, L. Scales of spatial variation in Mediterranean subtidal sessile assemblages at different depths. Mar. Ecol. Prog. Ser. 2007, 332, 25–39. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2010; ISBN 1855311402. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v.6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008; p. 214. [Google Scholar]
- Gabriele, M.; Bellot, A.; Gallotti, D.; Brunetti, R. Sublittoral hard substrate communities of the northern Adriatic Sea. Cah. Biol. Mar. 1999, 40, 65–76. [Google Scholar]
- Chemello, R.; Russo, G.F. The molluscan taxocoene of photophilic algae from the Island of Lampedusa (Strait of Sicily, southern Mediterranean). Boll. Malacol. 1997, 33, 95–104. [Google Scholar]
- Sánchez-Moyano, J.E.; Estacio, F.J.; García-Adiego, E.M.; García-Gómez, J.C. The molluscan epifauna of the alga Halopteris scoparia in Southern Spain as a bioindicator of coastal environmental conditions. J. Mollus. Stud. 2000, 66, 431–448. [Google Scholar] [CrossRef]
- Chemello, R.; Milazzo, M. Effect of algal architecture on associated fauna: Some evidence from phytal molluscs. Mar. Biol. 2002, 140, 981–990. [Google Scholar]
- Pitacco, V.; Orlando-Bonaca, M.; Mavrič, B.; Popović, A.; Lipej, L. Mollusc fauna associated with the Cystoseira algal associations in the Gulf of Trieste (Northern Adriatic Sea). Mediterr. Mar. Sci. 2014, 15, 225–238. [Google Scholar] [CrossRef]
- Österling, M.; Pihl, L. Effects of filamentous green algal mats on benthic macrofaunal functional feeding groups. J. Exp. Mar. Biol. Ecol. 2001, 263, 159–183. [Google Scholar] [CrossRef]
- Underwood, A.J.; Chapman, M.G. Scales of spatial patterns of distribution of intertidal invertebrates. Oecologia 1996, 107, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Benedetti-Cecchi, L. Variability in abundance of algae and invertebrates at different spatial scales on rocky sea shores. Mar. Ecol. Prog. Ser. 2001, 215, 79–92. [Google Scholar] [CrossRef]
- Fraschetti, S.; Terlizzi, A.; Benedetti-Cecchi, L. Patterns of distribution of marine assemblages from rocky shores: Evidence of relevant scales of variation. Mar. Ecol. Prog. Ser. 2005, 296, 13–29. [Google Scholar] [CrossRef]
- Mykrä, H.; Heino, J.; Muotka, T. Scale-related patterns in the spatial and environmental components of stream macroinvertebrate assemblage variation. Glob. Ecol. Biogeogr. 2007, 16, 149–159. [Google Scholar] [CrossRef]
- Gofas, S. The genus Tricolia in the Eastern Atlantic and the Mediterranean. J. Mollus. Stud. 1982, 48, 182–213. [Google Scholar]
- Gofas, S. Notes on some Ibero-Moroccan and Mediterranean Tricolia (Gastropoda, Tricoliidae), with descriptions of new species. J. Mollus. Stud. 1993, 59, 351–361. [Google Scholar] [CrossRef]
- Scuderi, D.; Russo, G.F. Due nuovi Gasteropodi per le acque italiane: Melibe fimbriata Alder e Hancock, 1864 e Tricolia tingitana Gofas 1982 (Mollusco, Gastropoda). Biol. Mar. Mediterr. 2003, 10, 618–621. [Google Scholar]
- Soininen, J.; McDonald, R.; Hillebrand, H. The distance decay of similarity in ecological communities. Ecography 2007, 30, 3–12. [Google Scholar] [CrossRef]
- Ape, F.; Gristina, M.; Chemello, R.; Sarà, G.; Mirto, S. Meiofauna associated with vermetid reefs: The role of macroalgae in increasing habitat size and complexity. Coral Reefs 2018, 37, 875–889. [Google Scholar] [CrossRef]
- Dubois, S.; Commito, J.A.; Olivier, F.; Retière, C. Effects of epibionts on Sabellaria alveolata (L.) biogenic reefs and their associated fauna in the Bay of Mont Saint-Michel. Estuar. Coast. Shelf Sci. 2006, 68, 635–646. [Google Scholar] [CrossRef]
- La Porta, B.; Nicoletti, L. Sabellaria alveolata (Linnaeus) reefs in the central Tyrrhenian Sea (Italy) and associated polychaete fauna. Zoosymposia 2009, 2, 527–536. [Google Scholar]
- Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. Impacts of zebra mussels on aquatic communities and their role as ecosystem engineers. In Invasive Aquatic Species of Europe. DISTRIBUTION, Impacts and Management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Kluwer Accademic Publishers: London, UK, 2002; pp. 433–446. ISBN 978-94-015-9956-6. [Google Scholar]
- Arribas, L.P.; Donnarumma, L.; Palomo, M.G.; Scrosati, R.A. Intertidal mussels as ecosystem engineers: Their associated invertebrate biodiversity under contrasting wave exposures. Mar. Biodivers. 2014, 44, 203–211. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trophic Groups | Feeding Guilds | |
|---|---|---|
| Description | Code | |
| Herbivores | Deposit feeders, feeding on both diatoms and/or microalgae of sandy sediment | DF |
| Micro- and macro-grazers, feeding on both diatoms and/or algae growing on rocks, shells or on large plants | MG | |
| Carnivores | Predators, feeding on sedentary or motile animals | P |
| Ectoparasites, feeding on much larger animals on which they live during their life cycle | E | |
| Omnivores | Suspension feeders, feeding on the organic particles suspended in the water, which mainly include algae or, in coastal habitats, bacteria, detritus and nano-zooplankton | SF |
| Taxa | Species | TC | FG | N | %DI | %F | ||
|---|---|---|---|---|---|---|---|---|
| CP | PP | Ja | ||||||
| Polyplacophora | ||||||||
| Acanthochitonidae | Acanthochitona fascicularis | Herbivore | MG | 7 | 4 | 2 | 2.17 | 66.67 |
| Acanthochitonidae | Acanthochitona crinite | Herbivore | MG | 4 | 3 | 2 | 1.50 | 66.67 |
| Lepidochitonidae | Lepidochitona sp. | Herbivore | MG | 14 | 0 | 0 | 2.33 | 33.33 |
| Chitonidae | Chiton (Rhyssoplax) olivaceus | Herbivore | MG | 0 | 0 | 1 | 0.17 | 11.11 |
| Gastropoda | ||||||||
| Fissurellidae | Fissurella nubecula | Herbivore | MG | 5 | 2 | 4 | 1.83 | 55.56 |
| Fissurellidae | Puncturella noachina | Herbivore | MG | 5 | 0 | 8 | 2.17 | 55.56 |
| Fissurellidae | Diodora gibberula | Herbivore | MG | 0 | 1 | 0 | 0.17 | 11.11 |
| Fissurellidae | Diodora graeca | Herbivore | MG | 0 | 0 | 1 | 0.17 | 11.11 |
| Planaxidae | Fossarus ambiguous | Herbivore | DF | 11 | 0 | 0 | 1.83 | 33.33 |
| Scissurellidae | Sinezona cingulata | Herbivore | DF | 24 | 0 | 10 | 5.67 | 66.67 |
| Cingulopsidae | Eatonina pumila | Herbivore | DF | 2 | 0 | 15 | 2.83 | 55.56 |
| Cingulopsidae | Tubbreva micrometrica | Herbivore | DF | 1 | 0 | 0 | 0.17 | 11.11 |
| Anabathridae | Pisinna glabrata | Herbivore | DF | 3 | 0 | 0 | 0.50 | 22.22 |
| Trochidae | Jujubinus cfr gravinae | Herbivore | MG | 0 | 0 | 1 | 0.17 | 11.11 |
| Trochidae | Gibbula drepanensis | Herbivore | MG | 11 | 0 | 26 | 6.17 | 66.67 |
| Phasianellidae | Tricolia miniata var. albino | Herbivore | MG | 1 | 0 | 0 | 0.17 | 11.11 |
| Phasianellidae | Tricolia puncture | Herbivore | MG | 3 | 0 | 0 | 0.50 | 11.11 |
| Phasianellidae | Tricolia tingitana | Herbivore | MG | 36 | 0 | 0 | 6.00 | 33.33 |
| Rissoidae | Crisilla semistriata | Herbivore | DF | 1 | 42 | 9 | 8.67 | 66.67 |
| Rissoidae | Setia amabilis | Herbivore | DF | 10 | 5 | 1 | 2.67 | 66.67 |
| Rissoidae | Crisilla simulans | Herbivore | DF | 3 | 0 | 0 | 0.50 | 11.11 |
| Rissoidae | Rissoa similis | Herbivore | DF | 3 | 5 | 6 | 2.33 | 100 |
| Rissoidae | Alvania tenera | Herbivore | MG | 0 | 4 | 0 | 0.67 | 11.11 |
| Rissoidae | Alvania sp. | Herbivore | MG | 1 | 0 | 0 | 0.17 | 11.1 |
| Eulimidae | Vitreolina philippi | Carnivore | E | 0 | 0 | 5 | 0.83 | 22.22 |
| Cerithiidae | Bittium latreillii | Herbivore | MG | 1 | 0 | 0 | 0.17 | 11.11 |
| Cerithiopsidae | Cerithiopsis minima | Carnivore | P | 0 | 0 | 1 | 0.17 | 11.11 |
| Cerithiopsidae | Cerithiopsis scalaris | Carnivore | P | 0 | 0 | 2 | 0.33 | 22.22 |
| Triphoridae | Obesula sp. | Carnivore | P | 0 | 2 | 0 | 0.33 | 11.11 |
| Triphoridae | Marshallora adversa | Carnivore | P | 0 | 1 | 2 | 0.50 | 33.33 |
| Triphoridae | Similiphora similior | Carnivore | P | 0 | 0 | 1 | 0.17 | 11.11 |
| Columbellidae | Columbella rustica | Herbivore | MG | 1 | 0 | 3 | 0.67 | 33.33 |
| Muricidae | Ocinebrina edwardsii | Carnivore | P | 0 | 0 | 2 | 0.33 | 22.22 |
| Muricidae | Stramonita haemastoma | Carnivore | P | 0 | 1 | 0 | 0.17 | 11.11 |
| Dorididae | Doris bicolor | Carnivore | P | 0 | 0 | 1 | 0.17 | 11.11 |
| Dorididae | indet. | Carnivore | P | 0 | 1 | 0 | 0.17 | 11.11 |
| Boselliidae | Bosellia mimetica | Herbivore | MG | 1 | 0 | 1 | 0.33 | 22.22 |
| Plakobranchidae | indet. | Herbivore | MG | 1 | 0 | 0 | 0.17 | 11.11 |
| Aplysiidae | Petalifera gravieri | Herbivore | MG | 0 | 0 | 1 | 0.17 | 11.11 |
| Bivalvia | ||||||||
| Mytilidae | Mytilaster solidus | Omnivore | SF | 16 | 3 | 150 | 28.17 | 100 |
| Mytilidae | Mytilus galloprovincialis | Omnivore | SF | 0 | 0 | 7 | 1.17 | 11.11 |
| Mytilidae | Modiolus barbatus | Omnivore | SF | 0 | 2 | 1 | 0.50 | 11.11 |
| Mytilidae | Lithophaga lithophaga | Omnivore | SF | 1 | 1 | 7 | 1.50 | 44.44 |
| Mytilidae | Musculus costulatus | Omnivore | SF | 0 | 0 | 6 | 1.00 | 33.33 |
| Mytilidae | Gregariella semigranata | Omnivore | SF | 0 | 1 | 14 | 2.50 | 44.44 |
| Gastrochaenidae | Rocellaria dubia | Omnivore | SF | 0 | 0 | 17 | 2.83 | 33.33 |
| Arcidae | Arca noae | Omnivore | SF | 0 | 0 | 3 | 0.50 | 33.33 |
| Arcidae | Arca tetragona | Omnivore | SF | 0 | 0 | 1 | 0.17 | 11.11 |
| Hiatellidae | Hiatella arctica | Omnivore | SF | 0 | 1 | 12 | 2.17 | 44.44 |
| Veneridae | Irus irus | Omnivore | SF | 1 | 0 | 0 | 0.17 | 11.11 |
| Veneridae | Petricola lithophaga | Omnivore | SF | 0 | 0 | 2 | 0.33 | 22.22 |
| Carditidae | Cardita calyculata | Omnivore | SF | 5 | 1 | 0 | 1.00 | 33.33 |
| Site | N | SR | H’ | J’ |
|---|---|---|---|---|
| CP | 57.33 ± 22.01 | 16.67 ± 4.62 | 3.51 ± 0.29 | 0.87 ± 0.03 |
| PP | 28 ± 30.35 | 10.33 ± 2.52 | 2.77 ± 0.59 | 0.84 ± 0.24 |
| Ja | 114.67 ± 41.06 | 24.67 ± 6.11 | 3.35 ± 0.70 | 0.73 ± 0.15 |
| F | 5.6565 | 7.1436 | 1.4629 | 0.65047 |
| p(perm) | 0.0318 | 0.0374 | 0.2972 | 0.5208 |
| Unique perms | 256 | 88 | 280 | 280 |
| PERMANOVA | df | SS | MS | F | Unique Perms | p(Perm) |
| Sites | 2 | 11450 | 5724.8 | 4.2157 | 280 | 0.0032 |
| Residual | 6 | 8147.7 | 1358 | |||
| Total | 8 | 19597 | ||||
| PAIR-WISE TEST | t | Unique perms | p(MC) | |||
| CP, PP | 1.8227 | 10 | 0.0598 | |||
| CP, Ja | 2.6485 | 10 | 0.0072 | |||
| PP, Ja | 1.9317 | 10 | 0.037 |
| Average Similarity | Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% |
|---|---|---|---|---|---|---|
| Ja—61.88% | Mytilaster solidus | 3.76 | 10.83 | 10.77 | 17.50 | 17.50 |
| Rocellaria dubia | 1.84 | 5.08 | 4.31 | 8.22 | 25.72 | |
| Gibbula drepanensis | 2.08 | 4.96 | 2.34 | 8.01 | 33.73 | |
| Gregariella semigranata | 1.71 | 4.92 | 5.35 | 7.95 | 41.67 | |
| Hiatella rugosa | 1.60 | 4.69 | 22.85 | 7.58 | 49.26 | |
| CP—58.06% | Sinezona cingulata | 2.18 | 9.84 | 4.26 | 16.94 | 16.94 |
| Tricolia tingitana | 2.41 | 9.19 | 4.86 | 15.82 | 32.76 | |
| Lepidochitona sp. | 1.71 | 7.44 | 3.12 | 12.81 | 45.57 | |
| PP—29.94% | Setia amabilis | 0.92 | 7.50 | 3.29 | 25.06 | 25.06 |
| Rissoa similis | 0.92 | 7.50 | 3.29 | 25.06 | 50.11 |
| Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% |
|---|---|---|---|---|---|
| Mytilaster solidus | 2.07 | 5.81 | 2.37 | 18.50 | 18.50 |
| Rissoa similis | 0.89 | 3.99 | 2.41 | 12.71 | 31.20 |
| Sinezona cingulata | 1.16 | 2.30 | 0.67 | 7.32 | 38.53 |
| Setia amabilis | 0.77 | 2.16 | 0.72 | 6.89 | 45.42 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donnarumma, L.; Sandulli, R.; Appolloni, L.; Sánchez-Lizaso, J.L.; Russo, G.F. Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions. Diversity 2018, 10, 96. https://doi.org/10.3390/d10030096
Donnarumma L, Sandulli R, Appolloni L, Sánchez-Lizaso JL, Russo GF. Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions. Diversity. 2018; 10(3):96. https://doi.org/10.3390/d10030096
Chicago/Turabian StyleDonnarumma, Luigia, Roberto Sandulli, Luca Appolloni, José Luis Sánchez-Lizaso, and Giovanni Fulvio Russo. 2018. "Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions" Diversity 10, no. 3: 96. https://doi.org/10.3390/d10030096
APA StyleDonnarumma, L., Sandulli, R., Appolloni, L., Sánchez-Lizaso, J. L., & Russo, G. F. (2018). Assessment of Structural and Functional Diversity of Mollusc Assemblages within Vermetid Bioconstructions. Diversity, 10(3), 96. https://doi.org/10.3390/d10030096