3. Descriptions
Genus Philetaerobius Marshall, 1923
Philetaerobius Marshall, 1923: 546 [
1]; Schenkling & Marshall, 1929: 20 [
4]; Louw, 1998: 24 [
5]; Alonso-Zarazaga & Lyal, 1999: 144 [
6]; Oberprieler, 2010: 11 [
3].
Type species: Philetaerobius nidicola Marshall, 1923, by original designation.
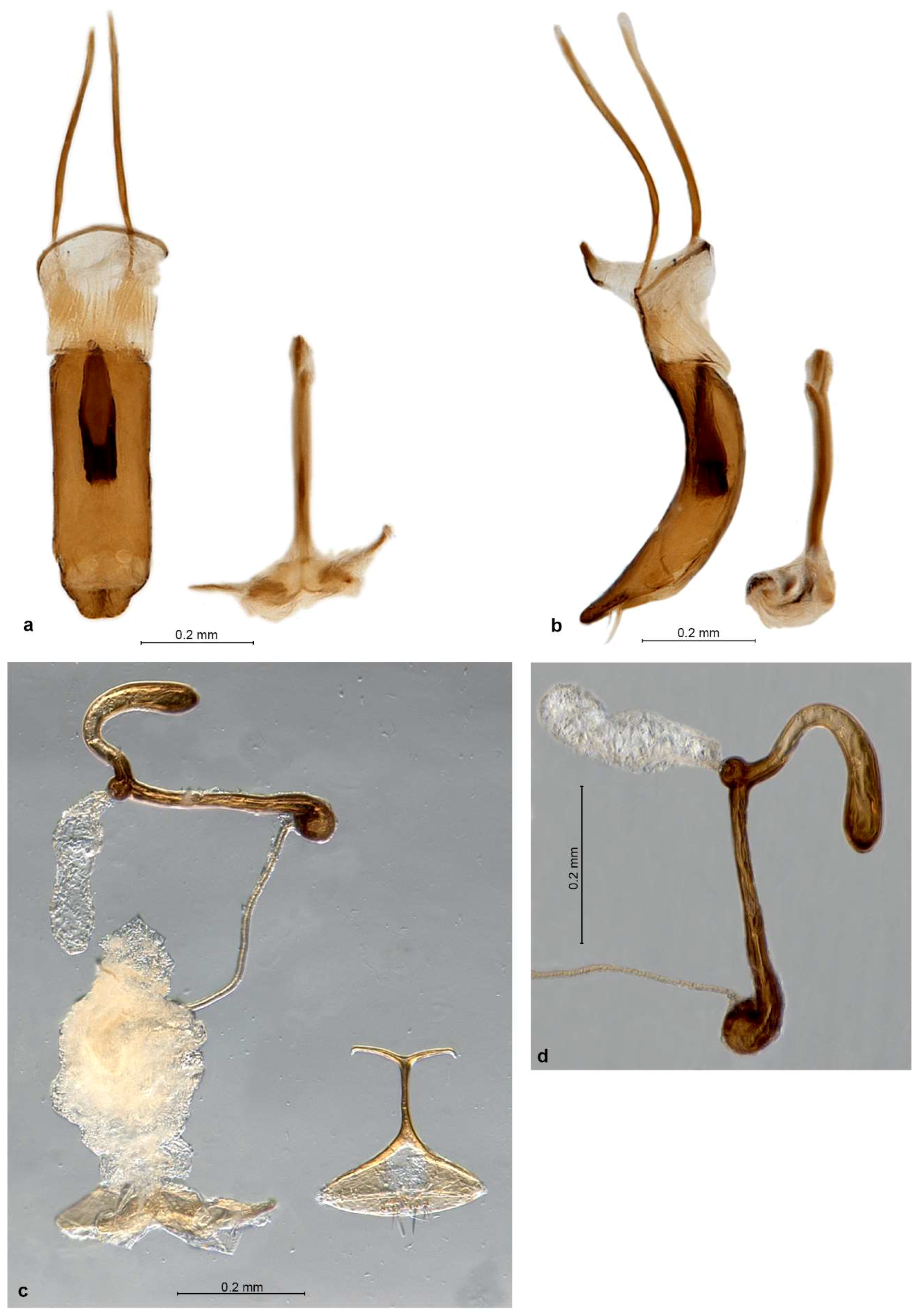
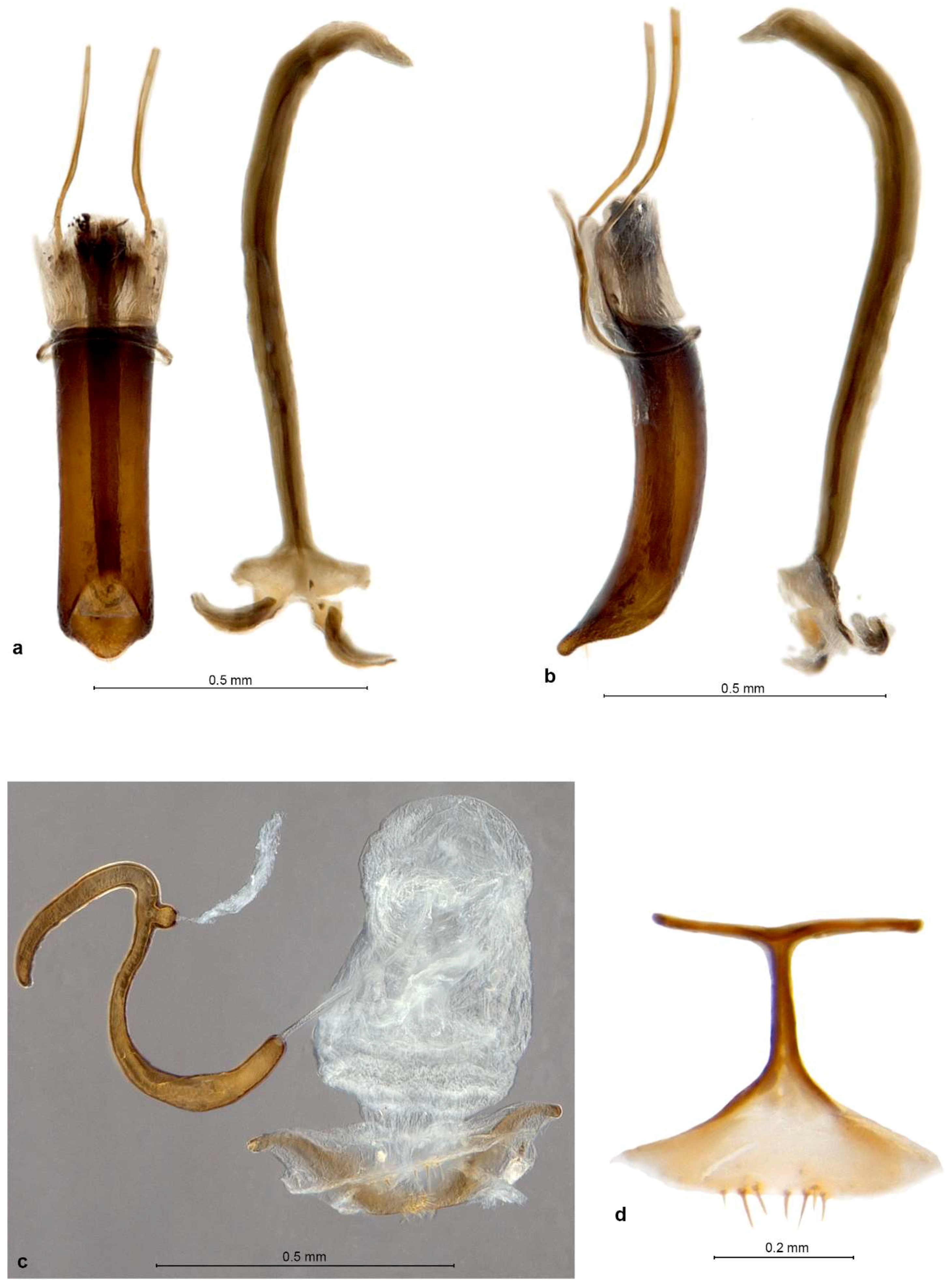
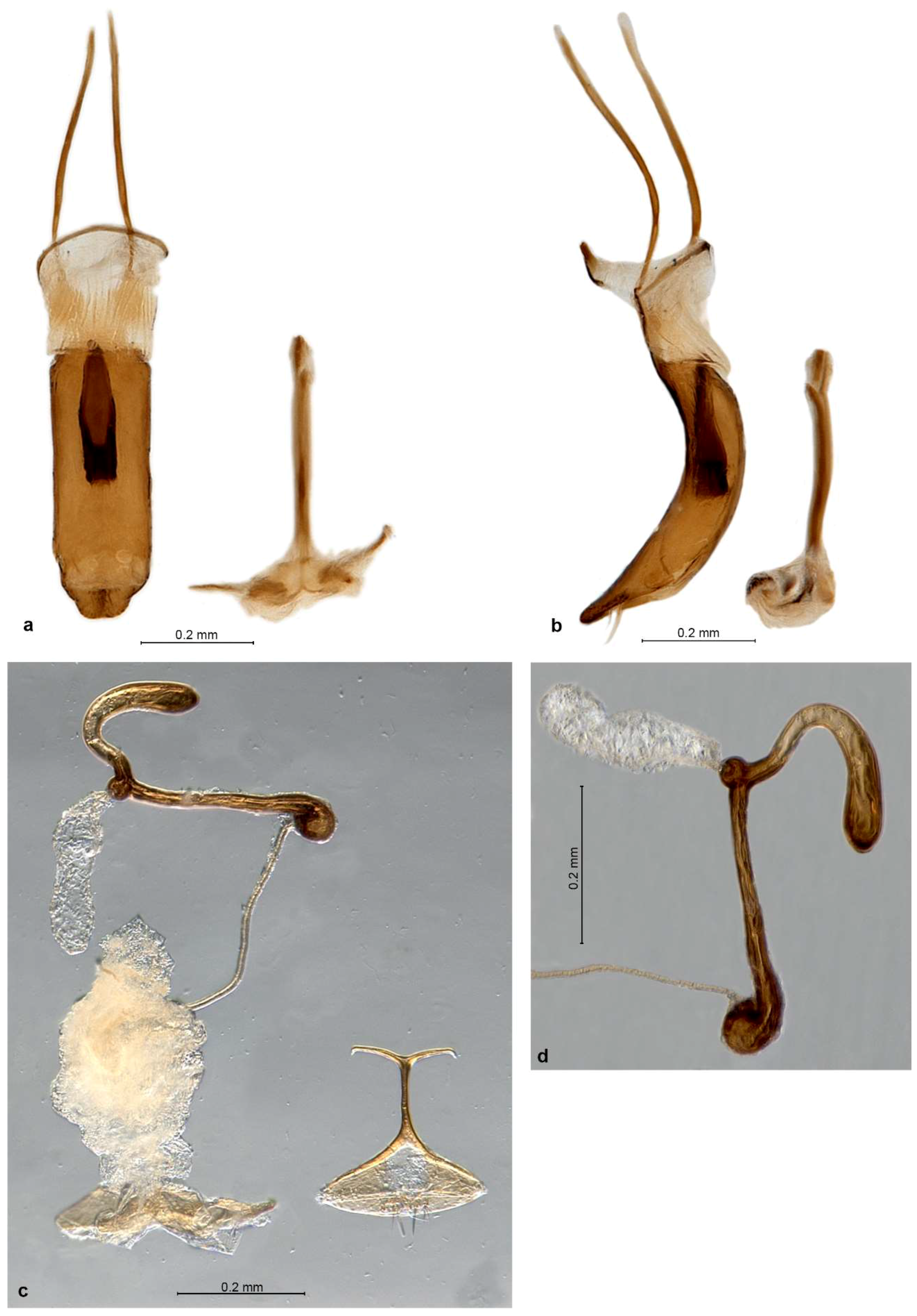
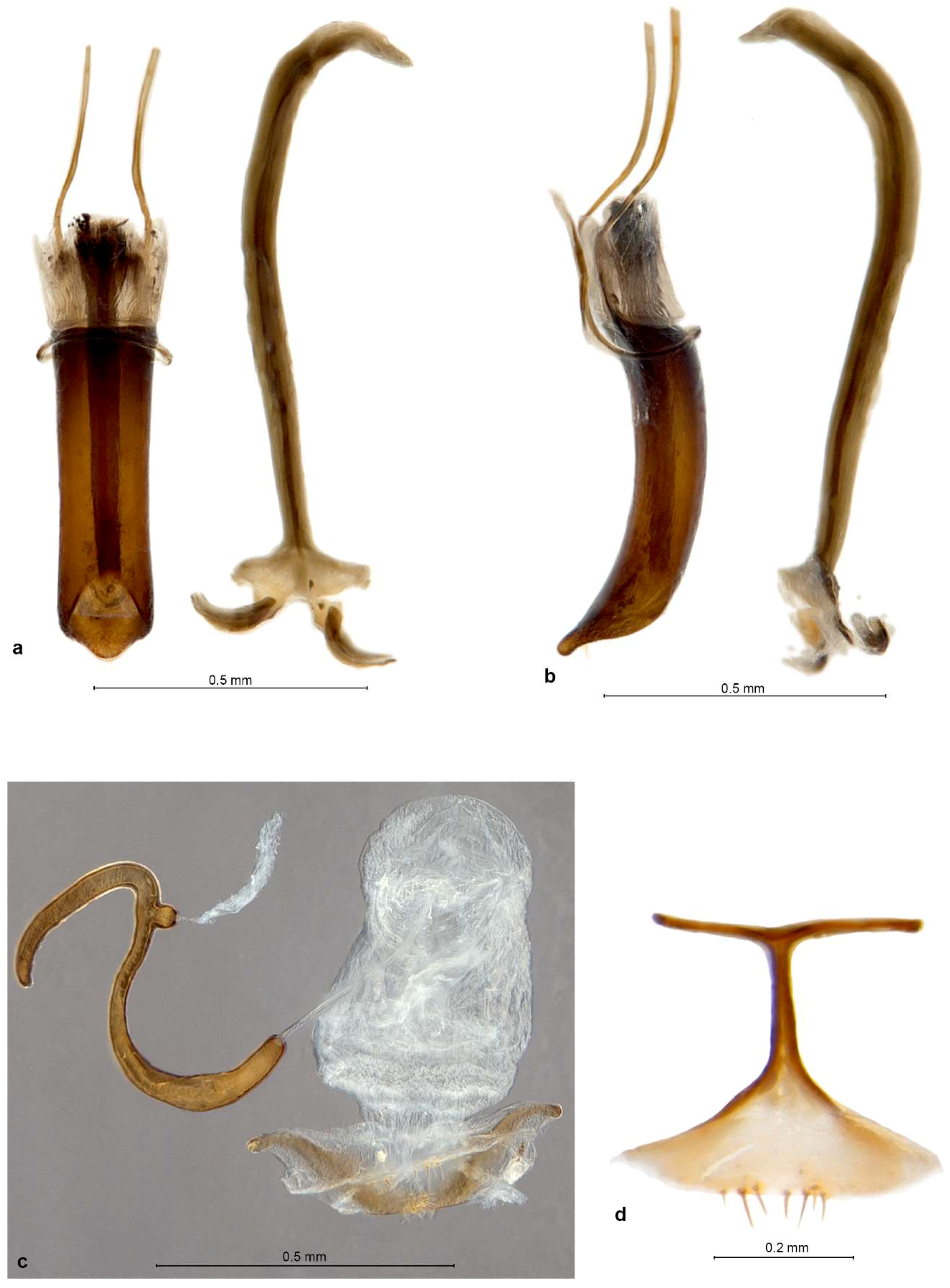
Diagnosis. Small Entiminae, with rostrum distinctly enlarged apicad, at apex broader than at base; frons large, indistinctly triangular, apically tectate, with 6–8 pairs of long setae; mandibles with spoon-shaped, vertical deciduous cusps; antennal scrobes narrow, straight, usually confluent on venter of rostrum; eyes in lateral view subtriangular to flatly reniform; antennal scapes significantly shorter than funicles; procoxal cavities nearer to posterior than anterior border of prothorax; metatibiae without corbels; tarsal claws single or inner one (outer one on protibiae) reduced to minute tooth at the base of other; tergite VII in both sexes with sclerotised margin, deeply V-shaped concave with straight sides and a row of coarse punctures; ventrite 1 in middle about as long as ventrite 2, ventrite 2 slightly shorter than 3–5 combined; gonocoxites small, flat, placed at angle to each other, without styli; sternite VIII of female with basal plate broadly triangular, its basal margin sclerotised, apodeme with apex transformed into a long transverse bar; spermatheca with collum longer than cornu, irregularly distorted, apically usually bulbous and curled over.
Redescription. Body length 2.7–4.2 mm. Integument of body black, legs and antennae blackish or reddish brown with apical parts of femora, basal parts of tibiae and clubs blackish, claws black, apical tibial spines yellowish brown. Vestiture of body consisting of very dense, tessellate, appressed scales, completely hiding integument; scales on elytra large and flat, 3 across one elytral interstria, irregularly angular, leaving only very slender spaces between them, sometimes concave and with irregular puncture in centre; scales on pronotum, head and rostrum, legs, antennae except clubs and underside slightly smaller than elytral ones, imbricate, with a distinct puncture in centre. Raised setae on whole body inconspicuous, very sparse and very short, pale to dark brownish, hardly visible in lateral view, shorter than diameter of one adherent scale; pronotum dorsally with anterior fringe of short semi-erect setae; antennal funicle, tibiae and tarsi with short, suberect to recumbent, subspatulate greyish setae. Color pattern of body various, in fresh specimens basic colour of pronotum and elytra brown, often with small irregular blackish spots on dorsal part of elytra and greyish on outer interstriae and two slender blackish longitudinal stripes on pronotum; in some specimens predominant color of pronotum and elytra dark brown to blackish, with irregular small spots of pale brownish and whitish scales, in others creamy with few scattered white scales; head and rostrum blackish, frons greyish, sometimes with pearly sheen; antennae and legs greyish; only vestiture of P. garibebi paler, grey with admixed blackish and greenish or bronze scales; underside white greyish, legs and underside in some specimens opalescent.
Rostrum 1.13× broader than long to 1.14× longer than broad, ventral part in dorsal view not visible below epifrons, in apical half to two-thirds distinctly widened apicad, at apex broader than at base and slightly narrower than head including eyes; in lateral view flat, at same level as head, anteriorly more or less abruptly declivous beyond antennal insertions. Epifrons broad, flat or longitudinally depressed, tapering anteriad with straight or weakly concave sides, at base as wide as interocular space, with slender median longitudinal stria along the whole length. Frons (
Figure 1d,
Figure 2d,
Figure 4d,
Figure 6d,
Figure 8d) large, vaguely triangular to trapezoidal but not raised, reaching antennal insertions, squamose, border with epifrons indicated by lines between 2–4 pairs of long, stout setae; epistome distincty tectate, squamose, border with frons indicated by 2–3 pairs of setae, one anteriorly and more widely spaced and the others further back. Mandibles very small, not projecting, base squamose; deciduous mandibular processes (
Figure 4e,f) vertical, flatly, bluntly asymmetrically spoon-shaped, with concavity on the outside and base, there thickened and strongly protruding, situated just inside of squamose base of mandible and leaving a narrow, vertical, subtriangular scar when broken off. Prementum with two setae. Antennal scrobes (
Figure 1e,
Figure 2e,
Figure 4e,
Figure 6e,
Figure 8e) in dorsal view invisible; in lateral view slender, sharp, curved obliquely downwards in front of eyes, mostly (except in
P. garibebi) shallowly to deeply confluent at back of venter of rostrum, creating a small flat median projection of venter (
Figure 6e). Head broad, flat to slightly concave, with slender longitudinal stria continuing into median stria on epifrons; laterally with prominent, rounded projection partly covering eyes in dorsal view. Eyes moderately large, kidney-shaped to subtriangular in outline, flat to slightly convex, placed in about middle of head height in lateral view, in dorsal view hardly to distinctly prominent from outline of head. Antennal scapes short, hardly reaching anterior border of eyes when folded back, at apex distinctly narrower than clubs, weakly curved, faintly regularly enlarged apicad. Funicles 7-segmented, 1.2–1.3× longer than scape, with segment 1 longer than broad, 2 isodiametric to transverse and 3–7 cupular to transverse, progressively shorter and broader towards club. Clubs oval with segment 1 conspicuously largest.
Pronotum flat, narrow, 1.08× longer than broad to 1.10× broader than long, sides strongly rounded, broadest near the midlength, strongly tapered anteriad and more gradually posteriad, anteriorly slightly broader than posteriorly, anteriorly constricted. Disc finely and densely punctate, flat or slightly vaulted with faint, shallow, curved depressions next to faint median longitudinal elevation, sometimes concealed below appressed scales. Base faintly to strongly arched. Anterior border in lateral view straight, without ocular lobes or setae. Procoxal cavities contiguous, round, nearer to posterior than to anterior border but not reaching it; procoxae subglobular. Scutellar shield indistinct.
Elytra very flat or slightly convex, subrectangular, together 1.48–1.71× longer than wide, subparallel-sided or weakly and regularly curved, bases jointly strongly arched and embracing posterior part of pronotum, sides straight or moderately curved, at apex shortly elongated and rounded, with regularly rounded shoulders; 9-striate, striae distinctly narrowly punctate; interstriae flat to slightly convex, with single row of sparse, short, blunt, semi-erect setae but these often largely or totally absent on even interstriae, in all species except P. garibebi interstriae 1 and 3 raised at base and transversely connected, 5–8 also raised at base into shallow humeral callus, in P. garibebi all interstriae equally wide and even ones also with complete row of sparse but small, translucent setae. Mesocoxae semiglobular, mesoventral process narrow, about as wide as third of mesocoxal width, not reaching posterior margin of mesocoxae, prominently raised in P. garibebi but not in other species. Metacoxae semiglobular, not reaching elytra (cavities laterally closed by metaventrite), metaventral process arched, about twice as wide as metacoxa. Tergite VII short and broad, translucent, with well sclerotised narrow margins, deeply V-shaped concave with straight sides and row of coarse punctures; tergite VIII rounded, well sclerotised, coarsely punctate. Tergite VII and VIII identical in both sexes. Femora faintly swollen, unarmed. Tibiae moderately long and slender, straight, apical surface (surrounding tarsal socket) densely squamose; protibiae apically rounded, mucronate, with 6–7 short, fine, yellowish, sparse, well separate spines, with lateral margin straight and mesal margin faintly bisinuate, at apex enlarged; mesotibiae apically with 6, metatibiae with 8 subequal spines, mesotibiae also mucronate but metatibiae amucronate; metatibiae without corbels. Tarsi slender and long, underside with several sparse long setae, with segment 2 slightly broader than long, 3 faintly broader than 2, bilobed, segment 5 slightly shorter than 3; claws single or with minute remnant of second claw (on outside in protibiae, on inside in meso- and metatibiae), about 0.3× as long as major claw.
Abdomen ventrally stretched subtriangular, about 1.3–1.4× longer than broad; ventrite 1 in middle about as long as 2, 2 slightly longer than 3 + 4 combined, 5 short, apically rounded; suture between 1 and 2 faintly sinuate, very fine and inconspicuous, other sutures somewhat arched towards apex, fine and narrow; all ventrites regularly, finely and densely punctate but obscured by dense cover of scales; ventrites 1, 2 and 5 with irregularly scattered, semierect to recumbent, moderately long, subspatulate setae, ventrites 3 and 4 each with single transverse row of the same setae; ventrites 1 and 2 in male longitudinally shallowly concave, in female convex.
Genitalia. Penis moderately short, well sclerotised, tubular, curved, apex ventrally attenuate, dorsally below ostium with a tuft of two or more long subapical setae; inside in basal half with thick, single but complex sclerite about 0.5× as long as body (
Figure 3a,
Figure 5a,
Figure 7a), in
P. garibebi with tubular sclerite as long as body and protruding from base of penis (
Figure 9a); temones 1.4–1.7× longer than body of penis and 4× longer than tegminal manubrium. Tegmen with slender ring without parameres, manubrium slightly shorter than diameter of ring. Sternite IX of male with spiculum gastrale moderately long, anteriorly curved and tapered, posteriorly with short, transverse basal plate; sternite VIII consisting of two sickle-shaped, sclerotised hemisternites. Gonocoxites lightly sclerotised, flat, short and broad, roundly subtriangular (in
P. garibebi longer and laterally curved outwards), at about right angle to each other, apex with a row of 3–7 stout setae, stylus absent. Sternite VIII of female (
Figure 3c,
Figure 5c,
Figure 7c,
Figure 9d) short and broad, basal plate broadly triangular, wider than long, in centre weakly sclerotised, often forming a round fenestra, basal margin thickened and sclerotised, apical margin thin, arcuate, medially usually with 1 pair of long stiff setae (sometimes 2 pairs, and 4 in
P. garibebi) and some smaller setae, sometimes medially notched; apodeme about as long as basal plate, conspicuously T-shaped, caput forming a slender transverse rod as long as or longer than stem of apodeme. Spermatheca (
Figure 3d,
Figure 5d,
Figure 7d,
Figure 9c) extraordinarily large and conspicuous, well sclerotised; cornu long, variously curved or bent, apically pointed or bluntly inflated; ramus short, subglobular, faintly longer than wide, sessile or on short stem, gland stalked, globular or elongate (in
P. garibebi); nodulus not differentiated; collum distinctly longer than cornu, variously curved and twisted, apex usually bulbous and curled so that duct arises next to collum (in
P. garibebi not curled and duct arising apically); spermathecal duct stiff, more or less straight, about as long as or shorter than spermatheca, inserted in middle of underside of bursa.
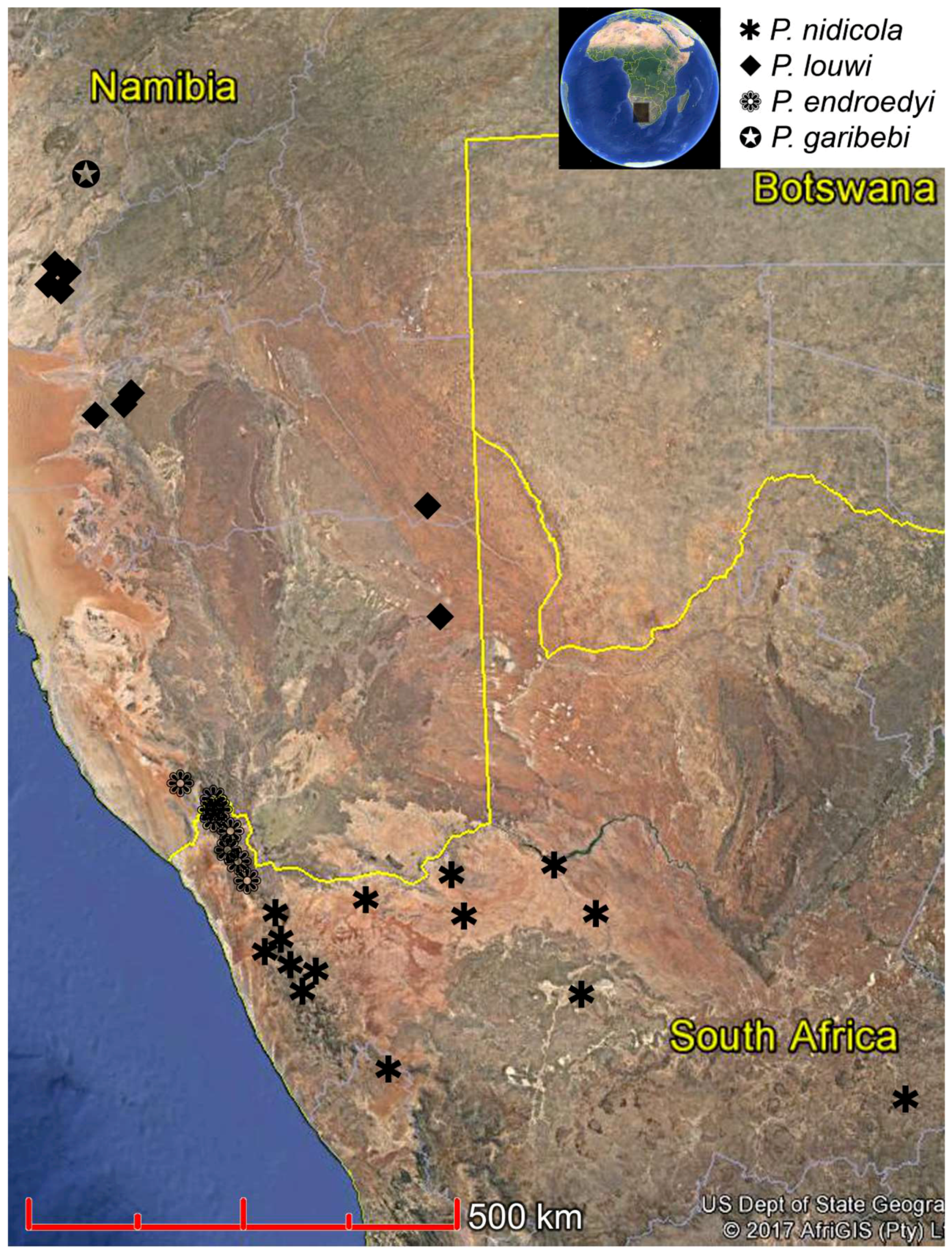
Distribution.Philetaerobius as known is endemic to the southern half of Namibia (the Erongo, Hardap and Karas regions) and the north-western parts of South Africa (the Northern and Western Cape provinces) (
Figure 10).
Habitat and life-history. Marshall described
P. nidicola from specimens found in a nest of the Sociable Weaver, a bird species endemic to southern Africa and occurring from central and south-eastern Namibia southwards through south-western Botswana into the Northern, North-West and Free State provinces of South Africa [
10]. Two additional short series of specimens have been collected from such nests, at least one evidently representing the same collecting event (see under
P. nidicola below). However, all newer specimens have been collected in pitfall traps [
2,
11], sifted from plant debris beneath shrubs or collected from grasses [
3]. Given also that the adults are wingless, it seems that the specimens collected from Sociable Weaver nests were carried into the nests with plant material collected by the birds to construct their nests. The flat body and cryptic coloration of all species except
P. garibebi suggest a specialized lifestyle on the ground, under leaf litter or stones or in crevices in the soil, but
P. garibebi appears to lead a more exposed way of life on plants, as other entimines generally do. The weevils have invariably been collected in xeric habitats, such as dry grassy plains adjacent to dunes [
11], but little further is known about their habitats and nothing about the larvae and their hostplants and feeding sites. The deciduous mandibular cusps of teneral adults indicate that the weevils pupate in the ground and have soil-dwelling, root-feeding larvae like other entimines do. The peculiar, shovel-like shape of the deciduous cusps suggests a specialized mode of pupation, perhaps in loose sand rather than in firm soil as it occurs in other entimines, and possibly the larvae of
Philetaerobius also lead a specialized way of life. Closer observation of the weevils in their habitats is required to properly assess their habits and life-history.
Remarks.Philetaerobius is a unique genus among the entimine fauna of southern Africa, differing most significantly from all other genera in the unusual shape of its spermatheca and sternite VIII of the female and, as far as known, of its deciduous mandibular cusps. Other characteristic features are the flat body shape, squamose epistome and frons, ventrally confluent or almost confluent scrobes, single or almost single tarsal claws, tubular internal sclerite of the penis, apical tuft of setae on the penis and tessellate scales arranged on the elytra in rosettes around the interstrial setae. The only entimine genus in southern Africa with a similar spermatheca is Spartecerus Schoenherr, 1834, to which Philetaerobius appears to be related on this account, although Spartecerus is very different in shape and other external features. A tubular internal sclerite of the penis also occurs in Spartecerus and, to a lesser degree, in Mimaulus, which furthermore has a similar rosette-like arrangement of scales on its pronotum and elytra. The likely relationships of Philetaerobius to these genera are explored in more detail below (see Discussion).
Philetaerobius can be regarded as a monophyletic taxon mainly on the shape of the apodeme of sternite VIII of the female, featuring a conspicuous transverse bar at its apex, and, as far as known, on the shovel-like mandibular processes, the squamose epistome and the apical tuft of setae on the penis. The structure of its spermatheca is also rather unique, differing from the similar one of Spartecerus in not having a nodulus differentiated at all. In Philetaerobius, P. garibebi occupies an isolated position, differing in several characters from all the other species (see description and key). Some of these characters appear to be more derived, e.g., the single claws and absence of humeri, whereas others seem more plesiomorphic, e.g., the shape of the elytra (especially at the base) and eyes, the long tubular penis sclerite and the development and setation of the gonocoxites and sternite VIII of the female. While these numerous differences may warrant placing P. garibebi in a separate genus, we prefer to assign it to Philetaerobius as it shares all the critical genital characters of the other species and evidently belongs to the same monophyletic taxon. When or if additional species of Philetaerobius are discovered and perhaps accentuate these differences, the taxonomic position of P. garibebi may be revised.
- 1.
Body in lateral view dorsally gently convex, elytral declivity in apical third; eyes vertically aligned on head in lateral view; scrobes not confluent on underside of rostrum; body dorsally covered with greyish-white scales, partly with a pearl to coppery sheen; tarsal claws single; penis with long, thick, straight, simple, cylindrical internal sclerite.....................P. garibebi sp. n.
- -
Body in lateral view dorsally flat, elytral declivity in apical fifth; eyes obliquely to horizontally aligned on head in lateral view; scrobes confluent on underside of rostrum; body dorsally covered with mainly brown scales, admixed with some black and white scales arranged in rosettes; tarsal claws unequal, tiny remnant of second one present; penis with short, thick, dorsally open, apically double internal sclerite.................................................................................2
- 2.
Eyes in lateral view subtriangular, regularly tapered posteriad, with ventroposterior margin straight, in dorsal view almost flat, hardly protruding from outline of head; epifrons flat; pronotum slightly longer than broad; internal sclerite of penis with dorsal apical arm as long and about as thick as ventral arm; spermatheca with cornu thickened apicad, apex globular......................................................................................................................P. endroedyi sp. n.
- -
Eyes in lateral view kidney-shaped, in posterior half abruptly tapered posteriad, with ventroposterior margin distinctly concave, in dorsal view faintly vaulted, protruding from outline of head; epifrons longitudinally depressed; pronotum slightly broader than long; internal sclerite of penis with dorsal apical arm shorter and thinner than ventral arm; spermatheca with cornu tapered apicad, apex acute...........................................................................3
- 3.
Rostrum shorter, 1.1× broader than long; elytra broader, 1.45–1.55× longer than broad, laterally at apex beak-shaped elongate; odd elytral interstriae more elevated than even ones on disc; funicle segments moniliform, well separated; internal sclerite of penis broad, dorsal arm divided into two laterally projecting teeth.........................................................P. nidicola Marshall
- -
Rostrum longer, 1.05× longer than broad; elytra slender, 1.6–1.7× longer than broad, laterally at apex regularly convex; elytral interstriae flat on disc; funicle segments cylindrical, closely approximated; internal sclerite of penis narrow, dorsal arm not divided into laterally projecting prongs...............................................................................................................................P. louwi sp. n.
Philetaerobius nidicola Marshall, 1923: 547 [
1]; Schenkling & Marshall, 1929: 20 [
4]; Alonso-Zarazaga & Lyal, 1999: 144 [
6]; Oberprieler, 2010: 11 [
3].
Redescription. Body length 3.16–4.25 mm, lectotype 4.06 mm. Color pale brownish, elytra with only several dark brownish or blackish and whitish spots, whitish spots sometimes forming irregular lateral stripes on interstriae 6 and 7, blackish spots dominant sometimes in posterior declivity. Pronotum with two irregular dark brownish longitudinal stripes. Rostrum 1.05–1.13× broader than long, in basal half faintly tapering anteriad, in apical half distinctly enlarged anteriad, with straight sides; in lateral view short and robust. Epifrons evenly tapered anteriad along whole length, with straight to slightly convex sides, longitudinally shallowly regularly deepened along the median longitudinal stria. Scrobes shallowly confluent at back of venter of rostrum. Eyes in dorsal view vaulted, weakly prominent from outline of head; in lateral view slender, horizontal, kidney-shaped, tapered posteriad, with ventral margin concave. Antennae with funicle with segments well separated, bead-shaped; segment 1 conical, 1.1–1.2× longer than broad and 1.1–1.2× longer than segment 2; segment 2 1.1–1.3× broader than long to isodiametric; segments 3–5 1.1–1.5× broader than long; segment 6 1.2–1.6× broader than long; segment 7 1.4–1.7× broader than long. Clubs 1.6–2.1× longer than broad. Pronotum 1.07–1.18× broader than long, broadest at midlength, in lateral view flat to slightly convex, with swollen anterior margin. Elytra 1.45–1.57× longer than broad, parallel-sided, apically broadly rounded, evenly tapered posteriad; interstriae flat, only 1, 3 and 6 behind base more elevated than others, 1 also on declivity slightly elevated. Tarsi with segment 2 isodiametric to 1.3× broader than long, segment 3 1.1–1.3× broader than long and 1.1–1.2× broader than segment 2, onychium 1.1–1.2× longer than segment 3. Genitalia. Penis parallel-sided, in apical fifth regularly tapered with slightly convex sides, with apex truncate, in lateral view regularly curved, equal in width, in apical third regularly tapered with lengthened apex; internal sclerite with dorsal arm apically divided into 2 stout short teeth directed lateroposteriad but not reaching apex of ventral part (giving the appearance of a clove), ventral part dorsoapically raised into narrow spout. Gonocoxites flat to shallowly convex, spatulate, broader in apical half; apex broadly rounded, with 1–2 stiff setae; orientated at about 90–135° to each other. Spermatheca with cornu abruptly bent at basal third of its length, apically slightly narrowed and bluntly pointed, often slightly bent; ramus globular, shortly stalked; gland ovoid, ca. 2× broader than ramus, on narrow stalk of same length; collum long, more or less straight but with slight irregular bends along the length, apex bulbous and curled over; duct stiff, straight to faintly twisted, shorter than spermatheca.
Material examined (60 exx.). Types. Lectotype (here designated), ♂: Type [printed, rounded with red margin] / Philetaerobius nidicola, Mshl. TYPE [handwritten] / S. Africa [printed] / In nest of Social Weaver Bird (Philetaerus socius) [handwritten] / Pres. by Imp. Bur. Ent. Brit. Mus. 1923-253. [printed] / LECTOTYPUS Philetaerobius nidicola Marshall // Borovec, Oberprieler & Meregalli desig. 2018 [printed, red] (BMNH). Paralectotypes: 1 ♀: SYNTYPE [printed, rounded with blue margin] / Philetaerobius nidicola, Mshl. COTYPE [handwritten] / S. Africa, fr. nest of Sociable Weaver bird [handwritten] / G.A.K. Marshall Coll. B.M. 1950-255 [printed] / PARALECTOTYPUS Philetaerobius nidicola Marshall // Borovec, Oberprieler & Meregalli desig. 2018 [printed, red] (BMNH); 1 ♂, 1 ♀?: from nest of / Social Weaver / bird [handwritten] // Philetaerobius / nidicola, Mshl. / COTYPES. [in Marshall’s hand] // Type / SAM/Ent / 3706 [on green card] / PARALECTOTYPUS Philetaerobius nidicola Marshall // Borovec, Oberprieler & Meregalli desig. 2018 [printed, red] (SAMC). Other specimens: 1 ♂, 8 exx.: TransKei / A. L. du Toit / 1910 [handwritten] // from nest of / Social Weaver / bird [handwritten] // Philetaerobius / nidicola Mshl [handwritten] // SAM-COL- / A048230 (SAMC); 6 exx.: [no locality], from thorn tree / bearing nest of / social weaver / bird [handwritten] // Philetaerobius / nidicola Mshl [handwritten] // SAM-COL- / A048229 (SAMC); 1 ♂: Hanover [31°4.100′ S 24°26.383′ E] / C. C. / Cronr. Schrein. / 1901 [handwritten] // Philetaerobius nidicola Mshl // SAM-COL- / A048227 (SAMC); 1 ♀, 5 exx.: Kenhardt [29°22.909′ S 21°11.211′ E] / 1911 // Philetaerobius / nidicola Mshl [handwritten] // SAM-COL- / A063585 (SAMC); 1 ♂: Kakamas [28°47.339′ S 20°38.214′ E] / Kenhardt Div. [typed] // R. F. Lawrence / May 1934 [underside of label] // SAM-COL- / A048228 (SAMC); 1 ♀: S. Afr. [Northern Cape], Richtersveld / Buffelsriv. Valley/29°33′ S 17°27′ E // 31.8.1976, E-Y: 1193 / groundtraps, 35 days / leg. Endrödy-Younga // groundtraps with / banana bait (TMSA); 1 ♂: [South Africa, Northern Cape], S.Afr., Namaqualand / Onseepkans–Kakamas / 28°52′ S 19°37′ E // 9.9.1976; E-Y:1244 / groundtraps, 24days / leg. Endrödy-Younga // groundtrap / with faeces bait (TMSA); 2 ♂, 5 exx.: [South Africa, Northern Cape], S.Afr., Namaqualand / Onseepkans–Kakamas / 28°52′ S 19°37′ E // 2.10.1976; E-Y:1280 / groundtraps, 12days / leg. Endrödy&Breyten. (TMSA, ANIC); 1 ♀: [South Africa, Northern Cape], S. Afr., Richtersveld / farm Haramoep / 29°06′ S 18°40′ E // 13.10.1976, E-Y: 1276 / cattle dung / Endrödy & Breytenb. (TMSA); 1 ♀: S. Afr. Cape [Northern Cape], Karroo / Struisputs farm / 30°02′ S 20°55′ E // 1.5.1985, E-Y: 2202 / under stones, plants / leg. M.-L. Penrith (TMSA); 1 ♀: SOUTH AFRICA, C. P. / 24 km W Springbok / 29°42′ S 17°44′ E / 10.ix.1986 / R. Oberprieler // collected off / Zygophyllum / morgsana (SANC); 1 ♂, 1 ♀: Gannapo[o]rt farm / 29°16′ S, 19°39′ E / 17.iii.[1988] // S. Afr. Cape // W. Wittmer (SANC); 1 ♀: RSA, Western Cape, 811 m / R 358 12 km S Kliprand / 30°40.169′ S, 18°42.597′ E / 30.x.2011, R. Borovec lgt. // sifted detritus and dead / leaves below / Euphorbia dregeana shrubs (RBSC); 1 ♂: RSA, Northern Cape, 541 m / 40 km S Springbok, Die Drif / 1 km from Koringhuis, sifting / 29°59.139′ S 17°51.875′ E / 31.x.2011, R. Borovec legit (MMTI); 1 ♂, 2 ♀, 4 exx.: RSA Northern Cape 757 m / Ca 2 km NE Nigramoep / 29°27.147′ S 17°37.968′ E / R. Borovec lgt. 15.xi.2016 / Sifting below Euphorbia dregeana (TMSA, RBSC, ANIC); 1 ♀, 1 ex.: RSA Northern Cape 545 m / W Anenous Pass / 29°14.475′ S 17°35.922′ E / R. Borovec lgt. 16.xi.2016 // Sifting of detritus died / leaves and branches / below shrubby Euphorbia dregeana (RBSC, ANIC); 2 ♀, 7 exx.: RSA Northern Cape 908 m / R355 SE Springbok / 29°43.641′ S 18°01.413′ E / Sifting below Euphorbia dregeana / R. Borovec lgt. 20.xi.2016 (RBSC, ANIC).
Distribution (
Figure 10). The species occurs in the Northern Cape province of South Africa, from Namaqualand eastwards into the Great Karoo, apparently as far east as Hanover (near De Aar). It does not seem to occur north of the Orange River and also not in the Richtersveld; the two localities so-labelled (Buffelsrivier Valley and Haramoep) lie further south, the former west of Springbok and the latter near Goodhouse in the Riemvasmaak Community Conservancy. The distribution range of
P. nidicola thus apparently does not overlap with those of
P. endroedyi and
P. louwi. The implied type locality of the species (“Transkei”, see below) is evidently erroneous as the Sociable Weaver does not occur in this region.
Habitat and life-history. The type and two other series of specimens in the SAMC were found in the communal nest of the Sociable Weaver (Philetairus socius, Ploceidae), but the species also occurs outside the range of this bird and more recently collected specimens have been taken on the ground in pitfall traps, among detritus under plants and under stones. A single specimen was beaten off a Zygophyllum bush, but this plant is unlikely to represent a host for the species. It remains to be seen whether P. nidicola may occur in Sociable Weaver nests (or other bird nests) more regularly and how the flightless specimens end up in such nests.
Derivation of name. Marshall [
1] did not state how he derived the name of the species, but it is obviously formed from the Latin noun
nidus, a nest, and the verb
colere, to live or inhabit. A
nidícola is a nest-dweller, and the species name is a masculine noun in apposition.
Remarks. Marshall [
1] based his description of
P. nidicola on five specimens. Two are housed in the BMNH, a male labelled “TYPE” and a female labelled “COTYPE”, and another two specimens are in the SAMC, glued in the top corners of a large card and labelled “COTYPES”. The space between these two specimens, with the remains of a ring of glue there, indicates that the card originally held a third specimen between the outer two, thus accounting for Marshall’s remaining fifth syntype. Although Marshall labelled the male in the BMNH as type, he did not designate a holotype in his description of
P. nidicola, and we therefore here designate the male in the BMNH as lectotype and the other three existing syntypes as paralectotypes.
The lectotype is glued on a triangular card and lacks the protarsi and several segments of the other tarsi. It was dissected by someone (not by Marshall or us) as its ventrites are glued on a card, and a second triangular card holds a blob of glue but without any genitalia. A search by one of us (M.M.) in the drawer in which the specimen is housed failed to find the aedeagus, which is apparently lost. The paralectotype female in the BMNH, glued on a rectangular card, is missing the entire right front leg. It has also been dissected and the ventrites, tergites VII and VIII, spermatheca and slightly damaged sternite VIII are glued on another rectangular card placed beneath it. The two paralectotypes in the SAMC were not dissected; the one on the right of the card, glued on its back, appears to be a male and the other is possibly a female. Although study of the genitalia of at least one type specimen is desirable for unequivocal assignment of other specimens to P. nidicola, we chose not to attempt dissection of the only two remaining intact types due to the insufficient dissecting facilities in the SAMC during our (R.G.O.) visit there, instead dissecting a male from the longer series of specimens (SAM-COL-A048230) bearing the same host label as the paralectotypes and evidently representing the same collecting event and a female from the series of six specimens from Kenhardt (SAM-COL-A048229). The internal sclerite of the dissected male has the characteristic clove-type shape and the spermatheca of the dissected female is consistent with that of other specimens assigned to P. nidicola here.
Marshall (1923) provided no locality for P. nidicola in his description, and none of the types have such attached to their pins. However, a series of nine specimens in the SAMC (SAM-COL-A048230) with an identical host label (handwritten “from nest of Social Weaver bird”), thus apparently representing the same collecting event, is labelled as being from the Transkei, the former region in the Eastern Cape province of South Africa lying northeast of the Great Kei River. This region is far removed from the Northern Cape province as well as from the range of the Sociable Weaver and therefore cannot be regarded as representing a conceivable type locality for P. nidicola. The date on the label, 1910, is plausible, but it appears that a wrong locality label has been attached to this series.
Philetaerobius nidicola differs from the other species most conspicuously in the characteristic clove-type internal sclerite of its penis. Its spermatheca differs from that of P. endroedyi in having an acute cornu and from those of P. louwi and P. garibebi in having the collum more or less straight, not widely curved. Externally P. nidicola differs from the former two species (the other flat ones of the genus) mainly by its shorter and more robust rostrum, which is broader than long in dorsal view. In other characters it is somewhat variable. The elytra and pronotum vary from being slender to broader and in lateral view from flat to slightly convex, and the range of the length-width ratio of the antennal clubs is larger than in the other species, the shape varying from slender and spindle-shaped to oval.
The COX1 sequence of a male from Nigramoep has been deposited on GenBank.
Philetaerobius nidicola: Louw, 1986: 310 [
2].
Description. Body length 3.16–4.50 mm, holotype 4.05 mm. Integument black but densely covered with tessellate scales, these mostly pale orange-brown or greyish in color but on pronotum and elytra also irregularly scattered black and whitish scales, arranged in partial or complete rosettes around strial punctures on lateral and posterior parts of elytra, and white scales sometimes forming a large drop-shaped macula on pronotal disc. Rostrum 1.04–1.06× longer than broad, in basal half faintly narrowing anteriad, in apical half distinctly widening anteriad; in lateral view moderately short and robust, abruptly declivous in front of antennal insertions. Epifrons inflated, sides straight but slightly converging anteriad, disc longitudinally deeply trough-shaped impressed towards a narrow median stria. Scrobes shallowly confluent at back of venter of rostrum. Eyes flat but slightly curved, in dorsal view barely visible below broad and raised forehead; in lateral view narrowly elongate, horizontal, anteroventrally extended into acute angle, ventral margin concave and raised on blunt canthus from posterior and ventral part of head. Antennae with funicle segments cupular, closely approximated; segment 1 more cylindrical, 1.1–1.2× longer than broad and 1.1–1.2× longer than segment 2, segment 2 1.1× broader than long, segments 3–5 1.2–1.3× broader than long, segment 6 1.4–1.5× broader than long, segment 7 1.6–1.7× broader than long. Clubs 1.6–1.7× longer than broad, very compact. Pronotum 1.06–1.16× broader than long, broadest at midlength, sides rounded; in lateral view flat with swollen anterior margin. Elytra long and slender, together 1.61–1.71× longer than broad, broadest at basal part and slightly, regularly tapered posteriad, base deeply emarginate and embracing base of pronotum, broadly rounded at apex; interstriae mostly flat, 3, 5 and 7 slightly roundly raised, 1 also raised on declivity, only just behind basal margin odd interstriae more elevated than even ones. Tarsi with segment 2 1.2× broader than long, segment 3 1.2–1.3× broader than long and 1.1–1.2× broader than segment 2, onychium 1.1× longer than segment 3. Genitalia. Penis parallel-sided, posteriorly abruptly truncate with narrow, attenuated, truncate apex, in lateral view regularly curved, basally slightly thicker than apically, posteriorly sharply tapered to narrow, flat ventral apex; internal sclerite narrowly navicular in dorsal view, in lateral view cleft in apical third, with dorsal arm shorter than ventral one and only about half as thick. Gonocoxites flat to shallowly convex, sublenticular, about equally broad throughout or slightly broader in apical half; apex broadly rounded, with 2–3 stiff setae; orientated at about 90–120° to each other. Spermatheca with cornu sharply curved into right angle at basal third of its length, nearly straight, apically usually shortly bent, blunt but not inflated; ramus small, globular, sessile or shortly stalked; gland elongate, without distinct stalk; collum very long (longer than spermathecal duct), strongly variously bent and coiled, often faintly hook-shaped, apex bulbous but not curled (insertion of duct rotated at most 90°); duct stiff, straight to slightly twisted, only about half as long as spermatheca.
Material examined (54 exx.). Types. Holotype, ♂: [Namibia, Erongo], S.W.Afr., Namib / Us Pass, Park Gate / 23°04′ S 15°35′ E // 15.11.1974, E-Y: 468 / groundtraps 70 days / leg. Endrödy-Younga // ground traps with / ferm.banana bait (TMSA). Paratypes. 3 ♀, 13 exx.: same data as holotype (TMSA, ANIC); 1 ♂, 1 ♀: South West Africa / ??? [obtained in 1973 from old school collection in Windhoek, probably collected on the Khomas Hochland] (ANIC); 2 exx.: [Namibia, Hardap], Bullspoort S.W.A. [24°8.943′ S 16°21.783′ E] / R. G. Strey (TMSA); 2 exx.: [Namibia, Hardap], S.W.Afr., Naukluft / Felseneck farm / 24°21′ S 16°00′ E // 25.10.1974, E-Y: 417 / groundtraps, 136 day / leg. Endrödy-Younga (TMSA); 1 ♀: [Namibia, Hardap], S.W.Afr., Nauwkluft / Nauwkluft Park / 24°16′ S 16°15′ E // 26.10.1974, E-Y: 425 / groundtraps, 88 day / leg. Endrödy-Younga // groundtrap with / banana bait (TMSA); 1 ex.: [Namibia, Hardap], S.W.Afr., Nauwkluft / Naukluft Park / 24°16′ S 16°15′ E // 26.10.1974, E-Y: 428 / groundtraps, 88 day / leg. Endrödy-Younga // ground traps / unbaited (TMSA); 3 exx.: [Namibia, Erongo], S.W.Afr., Namib / Ganab water / 23°06′ S 15°32′ E // 1.11.1974, E-Y: 437 / groundtraps, 17 day / leg. Endrödy-Younga // ground traps with / ferm. banana bait (TMSA); 1 ex.: [Namibia, Erongo], S.W.Afr., Namib / Ganab NE range / 23°08′ S 15°36′ E // 18.11.1974, E-Y: 483 / groundtraps, 65 day / leg. Endrödy-Younga // ground traps with / ferm. banana bait (TMSA); 1 ♂: [Namibia, Hardap], S.Afr., Kalah. Park / Farm Mara / 25°25′ S 19°30′ E // 19.12.1974, E-Y: 513 / ground traps, 73 d / leg. Endrödy-Younga // ground traps / with meat bait (TMSA); 3 exx.: [Namibia, Erongo], S.W.Afr., Namib / Ganab NE range / 23°08′ S 15°36′ E // 1.3.1975, E-Y: 711 / groundtrap: 90 day / leg. Endrödy-Younga (TMSA); 2 exx.: [Namibia, Erongo], S.W.Afr., Namib / Us Pass, 10 km Park / 23°03′ S 15°40′ E // 5.6.1975, E-Y: 852 / groundtraps 88 days / leg. Endrödy-Younga // ground traps / with faeces bait (TMSA); 1 ♂, 2 ♀, 1 exx.: [Namibia, Erongo], S.W.Afr., Namib / Us Pass, 10 km Park / 23°03′ S 15°40′ E // 1.9.1975, E-Y: 900 / groundtraps, 75 day / leg. Endrödy-Younga (TMSA); 1 ♂, 2 ♀, 8 exx.: [Namibia, Erongo], S.W.Afr. KhomasHl. / Us Pass, 10 km Park / 23°03′ S 15°40′ E // 7.7.1978; E-Y:1472 / groundtraps, 3years / leg. Endrödy-Younga (TMSA); 3 exx.: [Namibia, Erongo], S.W.Afr., c. Namib / Ganab, N. E. Hillgap / 23°08′ S 15°35′ E // 7.7.1978, E-Y: 1470 / groundtraps, 3 years / leg. Endrödy-Younga (TMSA); 1 ♀: [Namibia], Wildheim Ost 384 / SE 2619 Bc [26°28′ S 19°34′ E] / KEETMANSHOOP / 26–29 October 1976 / S. Louw, M.-L. Penrith // H33608 (SANC).
Distribution (
Figure 10). The species as known occurs in south-central Namibia, in the west along the edge of the Namib Desert from the Khomas-Hochland west of Windhoek southwards to the Naukluft, but also in the east along the western edge of the Kalahari Desert. It is probably distributed throughout south-central Namibia and may also occur in south-western Botswana and the Northern Cape province along the eastern border of Namibia, but its distribution range does not seem to overlap with that of
P. endroedyi in the south-west and with that of
P. nidicola in the south-east.
Habitat and life-history. Most specimens examined were collected in pitfall traps, some of which had been baited with fermented bananas or faeces, but it is unlikely that the weevils were attracted by the bait. Louw [
2] recorded the species (as
P. nidicola) to be common in the Kalahari in spring, occurring in large numbers on plains dominated by grasses and
Rhigozum trichotomum (in 1976 at Wildheim Ost) [
11]. It appears that the species is associated with grasses, but its larva and life-history remain unknown.
Derivation of name.Philetaerobius louwi is cordially named after the late Schalk van der Merwe Louw, Professor of Zoology and Entomology at the University of the Free State in Bloemfontein, South Africa, who collected 150 specimens of this species during an ecological study of ground-living Coleoptera in the Namib and Kalahari Deserts and who also made significant contributions to the taxonomy of other taxa of terricolous weevils in southern Africa. Schalk sadly and unexpectedly passed away while this paper was in the proof stage.
Remarks. This species is distinguishable from P. nidicola by its slender rostrum and elytra and from P. endroedyi by its kidney-shaped and vaulted eyes and longitudinally depressed epifrons. Its internal penis sclerite is also distinctive, being deeply cleft and with the dorsal arm shorter and narrower than the ventral one, and its spermatheca is characteristic in having a long, curved to twisted collum. It is one of three Philetaerobius species known from Namibia and apparently the most widespread and common one, P. garibebi and P. endroedyi being known from only a few specimens and occurring to the north and south of it, respectively.
Description. Body length 2.66–4.38 mm, holotype 4.38 mm. Integument black on body, dark testaceous on antennae and tarsi, densely covered with tessellate variegated golden brown and black scales, laterally and posteriorly on elytra admixed with a few white scales arranged in complete or partial rosettes around strial punctures. Rostrum 1.09–1.14× longer than broad, in basal third slightly tapering anteriad, in apical two-thirds widening anteriad; in lateral view moderately long and slender, abruptly declivous in front of antennal insertions. Epifrons dorsally tapering anteriad, sides almost straight, at declivity widening again, with faintly concave sides; disc flat, not impressed but with thin median stria along whole length. Scrobes deeply confluent at back of venter of rostrum. Eyes flat, in dorsal view narrowly visible below broad and raised forehead; in lateral view obliquely subtriangular, with ventroposterior margin straight, slightly raised on weak canthus from posterior and ventral part of head. Antennae with funicle segments subcylindrical to frustocone-shaped, closely approximated; segment 1 almost parallel-sided, 1.2× longer than broad and 1.5–1.6× longer than segment 2; segment 2 isodiametric to 1.1× longer than broad; segments 3–5 1.1–1.2× broader than long; segment 6 1.3–1.4× broader than long; segment 7 1.5× broader than long. Clubs oval, 1.4–1.6× longer than broad. Pronotum 1.03–1.08× longer than broad, broadest before midlength to anterior third, distinctly constricted behind anterior margin, in dorso-lateral view flat or faintly depressed on disc. Elytra together 1.56–1.67× longer than wide, parallel-sided, base deeply emarginate and embracing base of pronotum, broadly rounded at apex; interstriae flat to faintly convex, odd ones only in very short basal part somewhat more elevated than even ones and sutural ones elevated on declivity. Tarsi with segment 2 1.1× broader than long, segment 3 1.3–1.4× broader than long and 1.3–1.4× broader than previous segment, onychium 0.9× as long as segment 3. Genitalia. Penis subparallel-sided, posteriorly weakly attenuate, apex truncate, in lateral view regularly curved, equal in width, in apical third regularly tapered to ventrally placed apex; internal sclerite narrowly to broadly navicular in dorsal view, in lateral view cleft in apical quarter, with dorsal arm as long as ventral one and almost as thick. Gonocoxites flat to shallowly convex, subspatulate, broader in apical half; apex subrectangular, with 1–2 stiff setae; orientated at about 90–135° to each other. Spermatheca with cornu abruptly curved at basal third of its length, then evenly weakly C-shaped, apically inflated (twice thicker than at base) and broadly globular; ramus globular, broadly sessile to shortly stalked; gland elongate, medially constricted, not stalked; collum long but shorter than spermathecal duct, more or less straight but with slight irregular bends along the length, apex bulbous and tightly curled over (insertion of duct rotated 180°); duct stiff, straight to slightly twisted, shorter than spermatheca.
Material examined (59 exx.). Types. Holotype, ♂: RSA Northern Cape / Richtersveld 19.ix.2013 / rd to Akkedis pass 450 m / 28°09.880′ S 17°01.497′ E // Sifting of detritus, died / leaves and branches / below shrubby Euphorbia sp. / R. Borovec, M. Meregalli lgt. (TMSA). Paratypes: 1 ♂, 1 ♀, 7 exx.: same data as holotype (RBSC, MMTI, ANIC); 1 ♂, 1 ♀, 6 exx.: S. Afr. Richtersveld / Rooiberg Valley / 28°12′ S 17°07′ E // 4. 9. 1976; E-Y: 1217 / groundtraps, 30day / leg. Endrödy-Younga // ground traps with banana bait (TMSA); 3 ♀, 7 exx.: S. Afr., Richtersveld / GanakomRiv. Valley / 28°15′ S 17°07′ E // 5.9.1976; E-Y: 1223 / groundtraps, 35 days / leg. Endrödy-Younga // groundtrap / with banana bait (TMSA); 1 ♂, 8 exx.: RSA Northern Cape / Richtersveld 465 m / Koeroegab 19.ix.2013/28°17.298′ S 17°02.606′ E // Sifting of detritus, died / leaves and branches / below shrubby Euphorbia sp. / R. Borovec, M. Meregalli lgt. (RBSC, MMTI, ANIC); 1 ♀, RSA, Northern Cape / NE Eksteenfontein, 640 m // Pass E Jenkinskopf, 17.ix.2013 // 28°41.612′ S 17°16.559′ E (RBSC); 1 ♀, 3 exx., RSA, Northern Cape / S Eksteenfontein, 612 m / dir. Vioolsdrift, 23.ix.2013 / 28°51.957′ S 17°21.503′ E // Sifting of detritus, died / leaves and branches / below shrubby Euphorbia / R. Borovec, M. Meregalli lgt. (RBSC, MMTI); 1 ♂: RSA Northern Cape 463 m / Richtersveld NP 18.xi.2016 / Pass 2km S X RT 14 / 28°17.309′ S, 17°02.657′ E [R. Borovec lgt.] (ANIC); 1 ♂, 2 exx.: RSA Northern Cape 359 m / Richtersveld NP 19.xi.2016 / Gannakouriep / 28°23.558′ S 17°09.285′ E / R. Borovec lgt. sifting Euphorbia sp. (RBSC, ANIC); 1 ♀, 11 exx.: South Namibia Karas / 7 km N Rosh Pinah / 22.ix.2013 470 m / 27°53.711′ S 16°42.627′ E // Sifting of detritus, died / leaves and branches / below shrubby Euphorbia dregeana / R. Borovec, M. Meregalli lgt. (RBSC, MMTI, ANIC).
Distribution (
Figure 10). The species is known only from the Richtersveld area, from Eksteenfontein in the north-western part of the Northern Cape province of South Africa northwards across the Orange River to Rosh Pinah in the south-western part of Namibia. It appears not to overlap in distribution with
P. nidicola, which occurs in Namaqualand slightly further south, nor with
P. louwi, which occurs further north in Namibia.
Habitat and life-history. Most of the recently collected specimens were obtained by sifting detritus and dead branches under shrubby Euphorbia plants, E. dregeana and other species of similar appearance. This suggests that the weevils live among plant debris and walk around on the ground (perhaps at night). This probably accounts for them having been collected in pitfall traps, rather than being attracted to any bait placed in these.
Derivation of name.Philetaerobius endroedyi is named in memory of the late Sebestyén (Sebastian) Endrődy-Younga (1934–1999), erstwhile coleopterist at the former Transvaal Museum in Pretoria (now Ditsong Museum), who extensively collected beetles throughout South Africa and Namibia, mainly terricolous taxa such as Tenebrionidae but also many weevils, including several Philetaerobius specimens. In the formation of the species name endroedyi, we adopt the spelling of his surname as he used it in South Africa both in his publications and on his specimen labels, Endrödi-Younga, and as used in the names of many other species named after him.
Remarks. This species is readily distinguishable from the other three flat species by its subtriangular, dorsally flat eyes, flat epifrons, longer pronotum and spermatheca with an apically enlarged, blunt cornu. The sclerite in the penis is also characteristic, similar to that of P. louwi but posteriorly only cleft in the apical quarter, and the dorsal arm as long and about as thick as the ventral one.
Philetaerobius undescribed species: Oberprieler, 2010: 11 [
3].
Description. Body length 3.52–4.36 mm. Integument black on body, testaceous on antennae and tarsi, uniformly covered with tessellate scales, scales on dorsum, venter and legs pale grey with slight greenish (in males) or stronger bronze to coppery (in females) tinge, admixed with white scales and sparser black ones, these not forming distinct pattern but white scales often clustered around interstrial setae. Rostrum slightly (1.1×) longer than wide at base and apex, narrower in middle of length (at antennal insertions); in lateral view dorsal outline mostly flat, continuous with that of head, but more or less abruptly but only slightly declivous in anterior third. Epifrons broadly shallowly impressed in middle, with deep broad median sulcus from base to frons but largely obscured (closed) by scales. Scrobes not confluent at back of venter of rostrum. Eyes in dorsal view flat, not raised from outline of head, barely visible; in lateral view inversely ovate, with sharp ventral point. Funicle with all 7 segments distinct, segments 1 and 5–7 obconical, 2–4 subcylindrical; segment 1 1.4× longer than broad and 1.4× longer than segment 2, segment 2 1.4× longer than broad, segments 3–7 about as long as broad. Clubs 1.8× longer than broad. Pronotum 1.1× broader than long, broadest at midlength, in lateral view flat (males) to slightly convex (females), with swollen anterior margin. Elytra together 1.57× (males)–1.42× (females) longer than broad, laterally gently rounded, widest in apical third (males) to middle (females), base straight (male) to slightly emarginate (female) but not embracing base of pronotum. All interstriae very slightly convex, none raised above others, all with single row of sparse, short, translucent, recumbent setae. Tarsi with segment 2 1.18× broader than long, segment 3 about as long but broader (1.4×), onychium 1.16× longer than segment 3; claws single, without remnant of second claw. Genitalia. Penis stout, cylindrical, in dorsal view broadest at base, then narrowing fairly abruptly, middle portion subparallel-sided, apical part slightly flaring out, apex ventrally shortly, roundedly attenuate; in lateral view slightly arcuate, with base curved up and apex curved down, subparallel-sided except tapering down at apex and ending in narrow tip; internal sclerite thickly tubular, straight, longer than body of penis; temones very slender, about as long as body of penis. Tegmen very slender, without parameres, apodeme about half as long as temones. Gonocoxites longer, sinuately sclerotised with proximal end curved outwards, placed at >90 angle to each other, apex blunt, with a row of 7 stout setae. Spermatheca very long and slender, S-shaped; cornu sharply bent into acute angle just after junction with ramus, then straight before gently bent in apical third and tapering to blunt point; ramus small, globular, sessile, gland as long as cornu, with narrow stalk and elongate body; collum evenly curved, thicker near duct insertion, apex narrow, not bulbous or curled; duct stiff, straight, shorter than collum.
Material examined (2 exx.). Types. Holotype, ♂: “S.W.Africa/Namibia / 10 km E Karibib / 21°57′ S 15°57′ E / 10.iii.1987 / R. Oberprieler // collected / on grass” (SANC). Paratype, 1 ♀: same data as holotype (SANC).
Distribution (
Figure 10)
. The species is thus far known from a single locality in the western part of central Namibia.
Habitat and life-history. The only known specimens were collected clinging to the stems of green grasses.
Derivation of name. This species is named after its type locality, Karibib, in central Namibia, but using the original Nama name ǂgaribeb, which apparently means a place of preparing an edible plant (seemingly the fruits of the nara, Acanthosicyos horridus). The epithet garibebi is a latinized genitive singular noun derived from the Nama name.
Remarks. This species is quite different from all other Philetaerobius species, most obviously in its convex body, vertically aligned eyes, and predominantly greyish scales. It also differs in many additional features from the other three species (see description), but it shares the distinctive female genitalia and several other characters with these and we therefore place it in the same genus. In many of its features, such as the shape of its elytra, eyes, scrobes, spermatheca and internal sclerite of the penis, it appears to be less derived than the other species, but its single claws and total absence of humeri are evidently more apomorphic characters.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}