
Complexity of Damage-Associated Molecular Pattern Molecule Expression Profile in Porcine Brain Affected by Ischemic Stroke

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Results
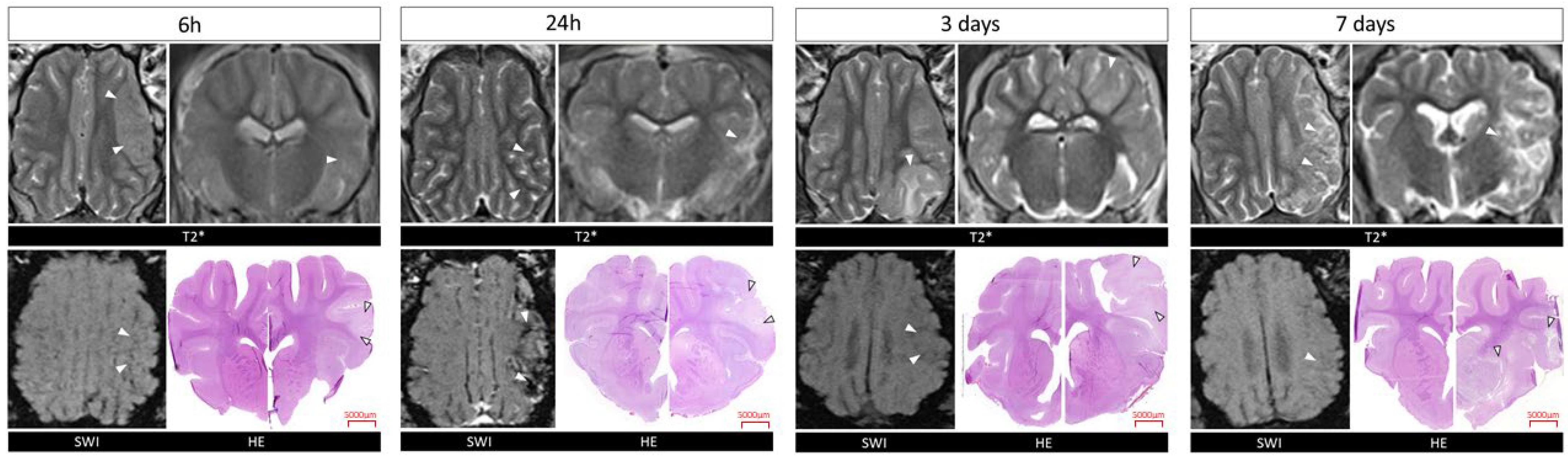
2.1. Confirmation of Ischemic Stroke Induction
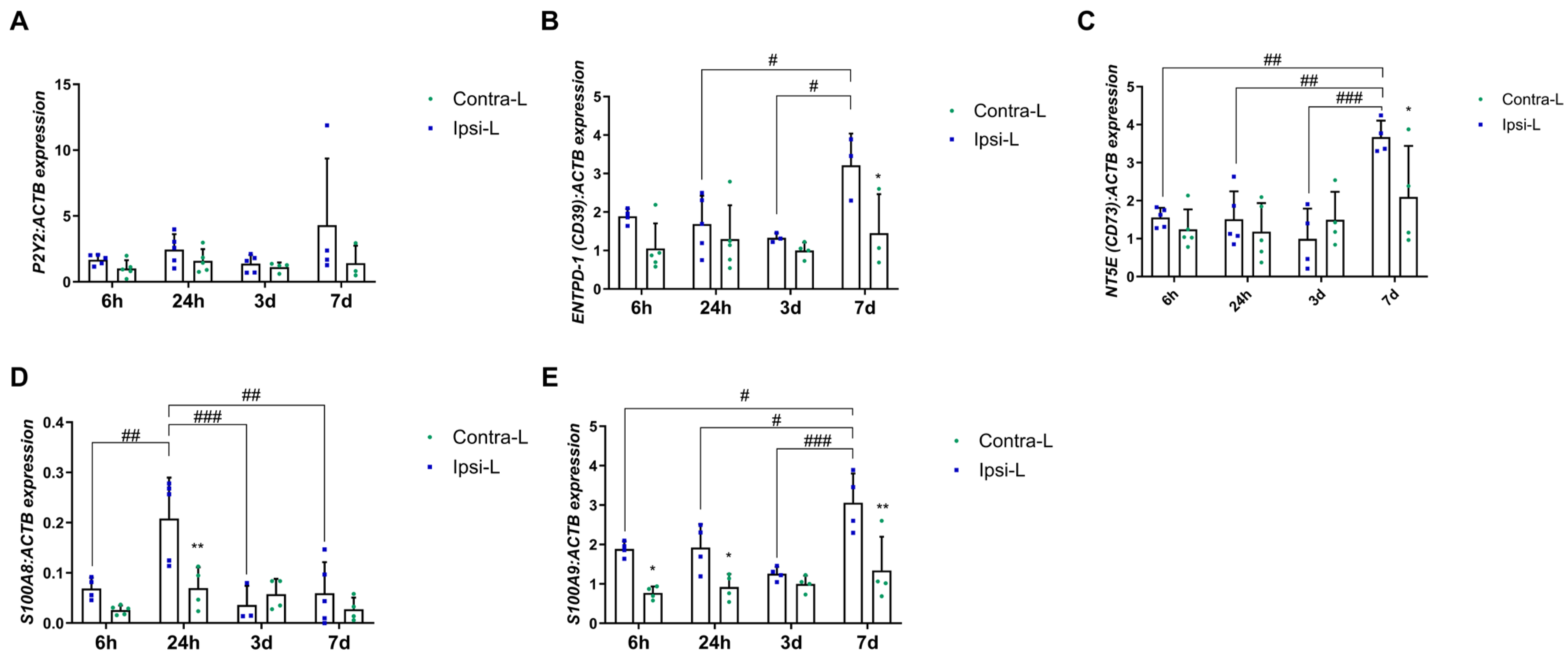
2.2. Purines and Calcium-Binding Proteins
2.3. Peroxiredoxins
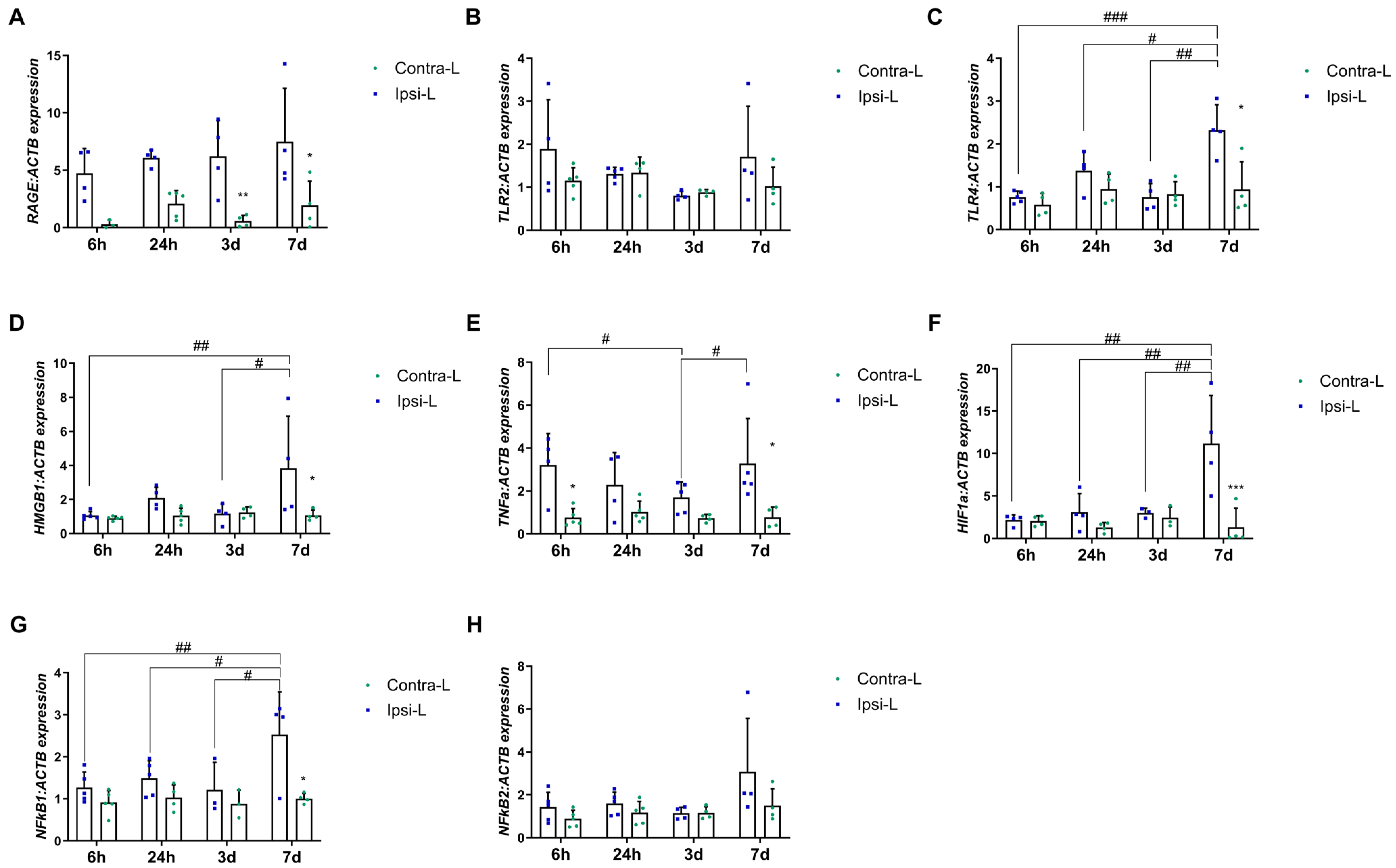
2.4. HMGB1, RAGE and TLR Receptors, TNFα and Transcription Factors
2.5. Interleukins
2.5.1. Gene Expression in the Brain Tissue
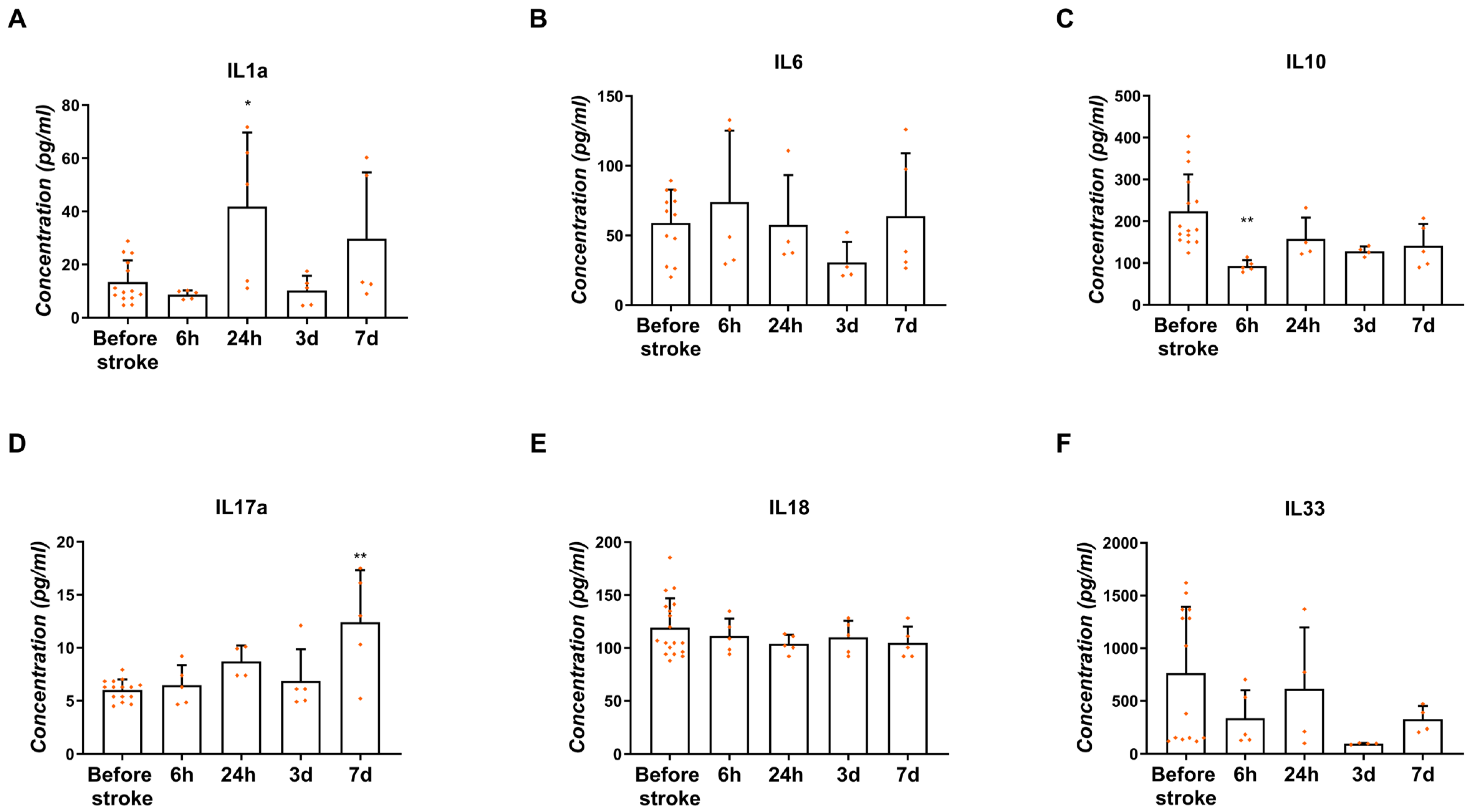
2.5.2. Concentration in the Blood Serum
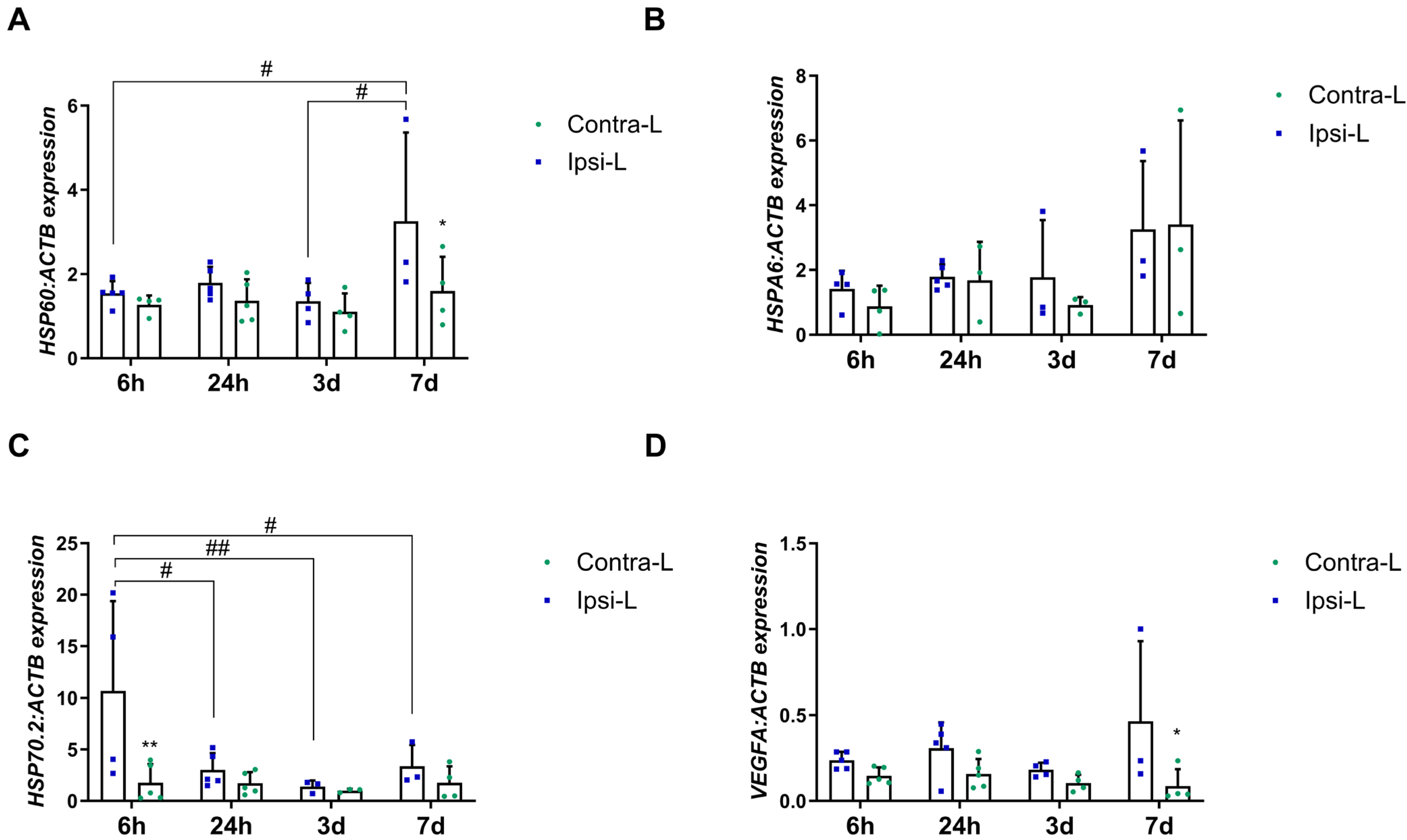
2.6. Heat Shock Protein Family
2.7. Vascular Endothelial Growth Factor A (VEGFA)
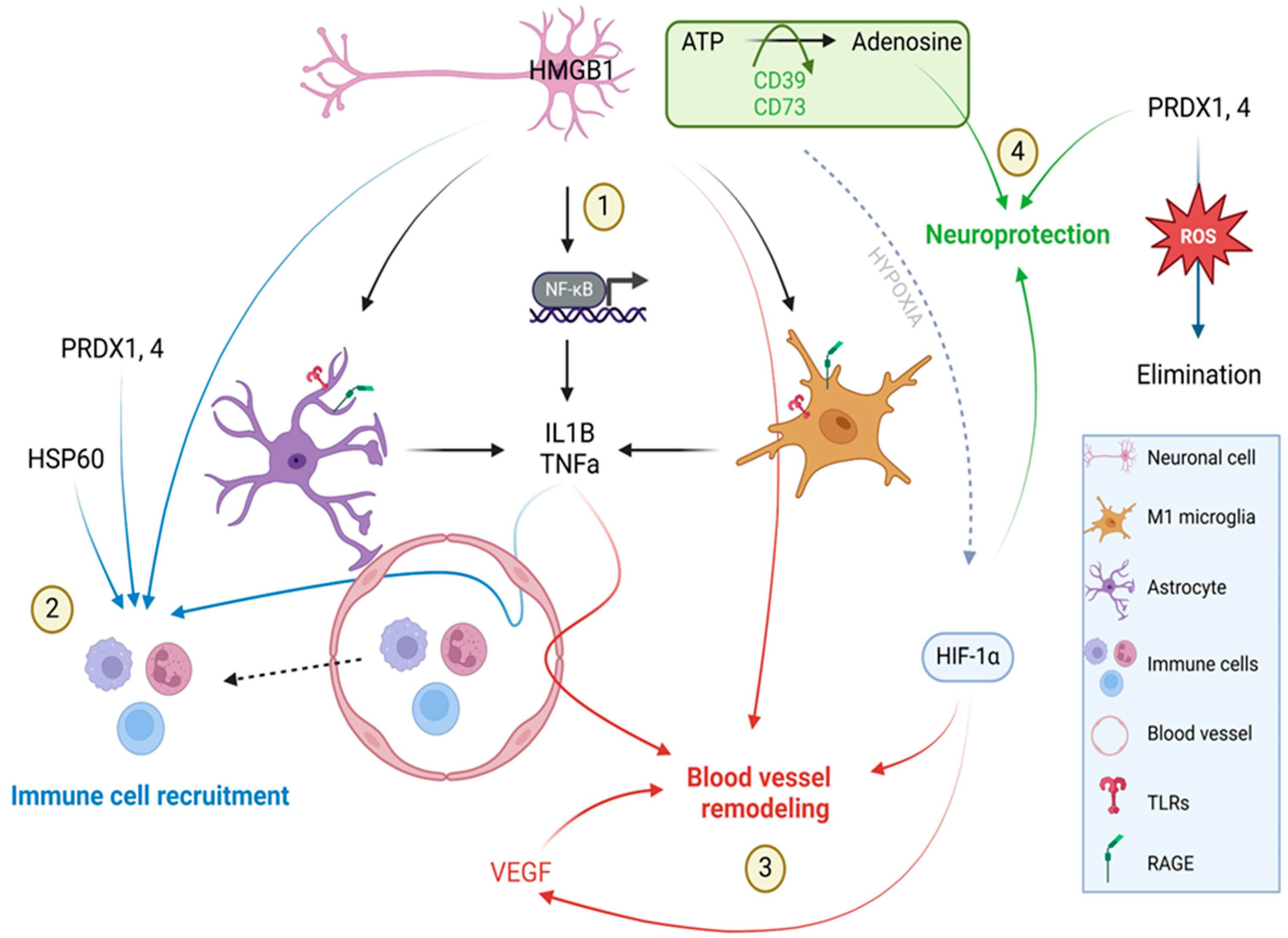
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Surgery and Sample Collection
4.3. Hematoxylin and Eosin Staining
4.4. Total RNA Extraction and Reverse Transcription
4.5. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.6. ELISA
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.-L.; Li, Y.; Zhao, B.-L.; Li, J.-S.; Zhang, N.; Ye, Z.-H.; Sun, X.-J.; Liu, W.-W. Necroptosis Signaling Pathways in Stroke: From Mechanisms to Therapies. Curr. Neuropharmacol. 2018, 16, 1327–1339. [Google Scholar]
- Schuhmann, M.K.; Kollikowski, A.M.; März, A.G.; Bieber, M.; Pham, M.; Stoll, G. Danger-associated molecular patterns are locally released during occlusion in hyper-acute stroke. Brain Behav. Immun. Health 2021, 15, 100270. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.; Zeng, D.; Zhou, W.; Hong, X. Prognostic value of plasma HMGB1 in ischemic stroke patients with cerebral ischemia-reperfusion injury after intravenous thrombolysis. J. Stroke Cerebrovasc. Dis. 2020, 29, 105055. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Mejorado, A.; Pérez-Samartín, A.; Gottlieb, M.; Matute, C. ATP signaling in brain: Release, excitotoxicity and potential therapeutic targets. Cell. Mol. Neurobiol. 2015, 35, 1–6. [Google Scholar] [CrossRef]
- Gulke, E.; Gelderblom, M.; Magnus, T. Danger signals in stroke and their role on microglia activation after ischemia. Ther. Adv. Neurol. Disord. 2018, 11, 1756286418774254. [Google Scholar] [CrossRef]
- Kim, J.Y.; Yenari, M.A. The immune modulating properties of the heat shock proteins after brain injury. Anat. Cell Biol. 2013, 46, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shevtsov, M.A.; Dobrodumov, A.V.; Mikhrina, A.L.; Yakovenko, I.V.; Guzhova, I.V.; Yakovleva, L.Y.; Vilisov, A.D.; Ischenko, A.M.; Margulis, B.; Galibin, O.V.; et al. Neurotherapeutic activity of the recombinant heat shock protein Hsp70 in a model of focal cerebral ischemia in rats. Drug Des. Dev. Ther. 2014, 8, 639–650. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, S.; Li, Y.; Sun, Y.; Xiong, X.; Hu, X.; Chen, J.; Qiu, S. Interleukins and Ischemic Stroke. Front. Immunol. 2022, 13, 828447. [Google Scholar] [CrossRef]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in Inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Hu, X.; Li, P.; Guo, Y.; Wang, H.; Leak, R.K.; Chen, S.; Gao, Y.; Chen, J. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke 2012, 43, 3063–3070. [Google Scholar] [CrossRef]
- Abdullahi, W.; Tripathi, D.; Ronaldson, P.T. Blood-brain barrier dysfunction in ischemic stroke: Targeting tight junctions and transporters for vascular protection. Am. J. Physiol. Cell Physiol. 2018, 315, C343–C356. [Google Scholar] [CrossRef]
- Justicia, C.; Panés, J.; Solé, S.; Cervera, Á.; Deulofeu, R.; Chamorro, Á.; Planas, A.M. Neutrophil infiltration increases matrix metalloproteinase-9 in the ischemic brain after occlusion/reperfusion of the middle cerebral artery in rats. J. Cereb. Blood Flow Metab. 2003, 23, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Pan, M.-X.; Tang, J.-C.; Zhang, Y.; Liao, H.-B.; Zhuang, Y.; Zhao, D.; Wan, Q. Role of neuroinflammation in ischemic stroke. Neuroimmunol. Neuroinflamm. 2017, 4, 158. [Google Scholar] [CrossRef]
- Huang, J.M.; Hu, J.; Chen, N.; Hu, M.-L. Relationship between plasma high-mobility group box-1 levels and clinical outcomes of ischemic stroke. J. Crit. Care 2013, 28, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Kunze, A.; Zierath, D.; Tanzi, P.; Cain, K.; Becker, K. Peroxiredoxin 5 (PRX5) is correlated inversely to systemic markers of inflammation in acute stroke. Stroke 2014, 45, 608–610. [Google Scholar] [CrossRef] [PubMed]
- Stanzione, R.; Forte, M.; Cotugno, M.; Bianchi, F.; Marchitti, S.; Rubattu, S. Role of DAMPs and of Leukocytes Infiltration in Ischemic Stroke: Insights from Animal Models and Translation to the Human Disease. Cell. Mol. Neurobiol. 2022, 42, 545–556. [Google Scholar] [CrossRef]
- Golubczyk, D.; Kalkowski, L.; Kwiatkowska, J.; Zawadzki, M.; Holak, P.; Glodek, J.; Milewska, K.; Pomianowski, A.; Janowski, M.; Adamiak, Z.; et al. Endovascular model of ischemic stroke in swine guided by real-time MRI. Sci. Rep. 2020, 10, 17318. [Google Scholar] [CrossRef]
- Chovsepian, A.; Berchtold, D.; Winek, K.; Mamrak, U.; Álvarez, I.R.; Dening, Y.; Golubczyk, D.; Weitbrecht, L.; Dames, C.; Aillery, M.; et al. A Primeval Mechanism of Tolerance to Desiccation Based on Glycolic Acid Saves Neurons in Mammals from Ischemia by Reducing Intracellular Calcium-Mediated Excitotoxicity. Adv. Sci. 2022, 9, e2103265. [Google Scholar] [CrossRef]
- Schadlich, I.S.; Winzer, R.; Stabernack, J.; Tolosa, E.; Magnus, T.; Rissiek, B. The role of the ATP-adenosine axis in ischemic stroke. Semin. Immunopathol. 2023, 45, 347–365. [Google Scholar] [CrossRef]
- Wilmes, M.; Espinoza, C.P.; Ludewig, P.; Stabernack, J.; Liesz, A.; Nicke, A.; Gelderblom, M.; Gerloff, C.; Falzoni, S.; Tolosa, E.; et al. Blocking P2X7 by intracerebroventricular injection of P2X7-specific nanobodies reduces stroke lesions. J. Neuroinflamm. 2022, 19, 256. [Google Scholar] [CrossRef]
- Xue, L.X.; Chen, S.-F.; Xue, S.-X.; Zhang, X.-Z.; Lian, Y.-J. P2RY2 Alleviates Cerebral Ischemia-Reperfusion Injury by Inhibiting YAP Phosphorylation and Reducing Mitochondrial Fission. Neuroscience 2022, 480, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, D.J.; Broekman, M.J.; Peschon, J.J.; Stocking, K.L.; Fujita, T.; Ramasamy, R.; Connolly, E.S., Jr.; Huang, J.; Kiss, S.; Zhang, Y.; et al. Elucidation of the thromboregulatory role of CD39/ectoapyrase in the ischemic brain. J. Clin. Investig. 2002, 109, 1031–1040. [Google Scholar] [CrossRef]
- Petrovic-Djergovic, D.; Hyman, M.C.; Ray, J.J.; Bouis, D.; Visovatti, S.H.; Hayasaki, T.; Pinsky, D.J. Tissue-resident ecto-5’ nucleotidase (CD73) regulates leukocyte trafficking in the ischemic brain. J. Immunol. 2012, 188, 2387–2398. [Google Scholar] [CrossRef]
- Pruenster, M.; Vogl, T.; Roth, J.; Sperandio, M. S100A8/A9: From basic science to clinical application. Pharmacol. Ther. 2016, 167, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Li, Q.; Zhang, Q.; Xu, L.; Han, J.-Y. Upregulated expression of S100A8 in mice brain after focal cerebral ischemia reperfusion. World J. Emerg. Med. 2013, 4, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Zhu, Z.; Xu, T.; Zhong, C.; Wang, A.; Xie, X.; Peng, Y.; Peng, H.; Li, Q.; Ju, Z.; et al. Plasma S100A8/A9 Concentrations and Clinical Outcomes of Ischemic Stroke in 2 Independent Multicenter Cohorts. Clin. Chem. 2020, 66, 706–717. [Google Scholar] [CrossRef]
- Olmez, I.; Ozyurt, H. Reactive oxygen species and ischemic cerebrovascular disease. Neurochem. Int. 2012, 60, 208–212. [Google Scholar] [CrossRef]
- Rodrigo, R.; Fernandez-Gajardo, R.; Gutierrez, R.; Matamala, J.M.; Carrasco, R.; Miranda-Merchak, A.; Feuerhake, W. Oxidative stress and pathophysiology of ischemic stroke: Novel therapeutic opportunities. CNS Neurol. Disord. Drug Targets 2013, 12, 698–714. [Google Scholar] [CrossRef]
- Liu, J.; Su, G.; Gao, J.; Tian, Y.; Liu, X.; Zhang, Z. Effects of Peroxiredoxin 2 in Neurological Disorders: A Review of its Molecular Mechanisms. Neurochem. Res. 2020, 45, 720–730. [Google Scholar] [CrossRef]
- Tang, B.; Ni, W.; Zhou, J.; Ling, Y.; Niu, D.; Lu, X.; Chen, T.; Ramalingam, M.; Hu, J. Peroxiredoxin 6 secreted by Schwann-like cells protects neuron against ischemic stroke in rats via PTEN/PI3K/AKT pathway. Tissue Cell 2021, 73, 101635. [Google Scholar] [CrossRef]
- Turovsky, E.A.; EVarlamova, G.; Plotnikov, E.Y. Mechanisms Underlying the Protective Effect of the Peroxiredoxin-6 Are Mediated via the Protection of Astrocytes during Ischemia/Reoxygenation. Int. J. Mol. Sci. 2021, 22, 8805. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bonilla, L.; Iadecola, C. Peroxiredoxin sets the brain on fire after stroke. Nat. Med. 2012, 18, 858–859. [Google Scholar] [CrossRef]
- Shichita, T.; Hasegawa, E.; Kimura, A.; Morita, R.; Sakaguchi, R.; Takada, I.; Sekiya, T.; Ooboshi, H.; Kitazono, T.; Yanagawa, T.; et al. Peroxiredoxin family proteins are key initiators of post-ischemic inflammation in the brain. Nat. Med. 2012, 18, 911–917. [Google Scholar] [CrossRef]
- Nitika; Porter, C.M.; Truman, A.W.; Truttmann, M.C. Post-translational modifications of Hsp70 family proteins: Expanding the chaperone code. J. Biol. Chem. 2020, 295, 10689–10708. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Han, Y.; Lee, J.E.; Yenari, M.A. The 70-kDa heat shock protein (Hsp70) as a therapeutic target for stroke. Expert Opin. Ther. Targets 2018, 22, 191–199. [Google Scholar] [CrossRef]
- Izaki, K.; Kinouchi, H.; Watanabe, K.; Owada, Y.; Okubo, A.; Itoh, H.; Kondo, H.; Tashima, Y.; Tamura, S.; Yoshimoto, T.; et al. Induction of mitochondrial heat shock protein 60 and 10 mRNAs following transient focal cerebral ischemia in the rat. Brain Res. Mol. Brain Res. 2001, 88, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Zeb, A.; Ali, T.; Muhammad, T.; Faheem, M.; Alam, S.I.; Saeed, K.; Koh, P.-O.; Lee, K.W.; Kim, M.O. Identification of Proteins Differentially Expressed in the Striatum by Melatonin in a Middle Cerebral Artery Occlusion Rat Model-a Proteomic and in silico Approach. Front. Neurosci. 2018, 12, 888. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Ding, Q.; He, Y.; Li, H. Role of IL-17A in different stages of ischemic stroke. Int. Immunopharmacol. 2023, 117, 109926. [Google Scholar] [CrossRef]
- DeLong, J.H.; Ohashi, S.N.; O’connor, K.C.; Sansing, L.H. Inflammatory Responses After Ischemic Stroke. Semin. Immunopathol. 2022, 44, 625–648. [Google Scholar] [CrossRef]
- Zhang, Q.; Liao, Y.; Liu, Z.; Dai, Y.; Li, Y.; Li, Y.; Tang, Y. Interleukin-17 and ischaemic stroke. Immunology 2021, 162, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zheng, Y.; Wu, Y.; Ni, B.; Shi, S. Imbalance between IL-17A-producing cells and regulatory T cells during ischemic stroke. Mediat. Inflamm. 2014, 2014, 813045. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Zhang, Y.; Shu, H.; Xie, B.; Tao, Y.; Yuan, Y.; Shang, Y.; Yuan, S.; Zhang, J. Hyperforin Promotes Post-stroke Neuroangiogenesis via Astrocytic IL-6-Mediated Negative Immune Regulation in the Ischemic Brain. Front. Cell Neurosci. 2019, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Lambertsen, K.L.; Finsen, B.; Clausen, B.H. Post-stroke inflammation-target or tool for therapy? Acta Neuropathol. 2019, 137, 693–714. [Google Scholar] [CrossRef] [PubMed]
- Shaafi, S.; Sharifipour, E.; Rahmanifar, R.; Hejazi, S.; Andalib, S.; Nikanfar, M.; Baradarn, B.; Mehdizadeh, R. Interleukin-6, a reliable prognostic factor for ischemic stroke. Iran. J. Neurol. 2014, 13, 70–76. [Google Scholar]
- Wu, F.; Liu, Z.; Zhou, L.; Ye, D.; Zhu, Y.; Huang, K.; Weng, Y.; Xiong, X.; Zhan, R.; Shen, J. Systemic immune responses after ischemic stroke: From the center to the periphery. Front. Immunol. 2022, 13, 911661. [Google Scholar] [CrossRef]
- Muhammad, S.; Barakat, W.; Stoyanov, S.; Murikinati, S.; Yang, H.; Tracey, K.J.; Bendszus, M.; Rossetti, G.; Nawroth, P.P.; Bierhaus, A.; et al. The HMGB1 receptor RAGE mediates ischemic brain damage. J. Neurosci. 2008, 28, 12023–12031. [Google Scholar] [CrossRef]
- Murad, S. Toll-like receptor 4 in inflammation and angiogenesis: A double-edged sword. Front. Immunol. 2014, 5, 313. [Google Scholar] [CrossRef]
- Voronov, E.; Carmi, Y.; Apte, R.N. The role IL-1 in tumor-mediated angiogenesis. Front. Physiol. 2014, 5, 114. [Google Scholar] [CrossRef]
- Yao, J.S.; Zhai, W.; Young, W.L.; Yang, G.-Y. Interleukin-6 triggers human cerebral endothelial cells proliferation and migration: The role for KDR and MMP-9. Biochem. Biophys. Res. Commun. 2006, 342, 1396–1404. [Google Scholar] [CrossRef]
- Huang, H.; Huang, Q.; Wang, F.; Milner, R.; Li, L. Cerebral ischemia-induced angiogenesis is dependent on tumor necrosis factor receptor 1-mediated upregulation of alpha5beta1 and alphaVbeta3 integrins. J. Neuroinflamm. 2016, 13, 227. [Google Scholar] [CrossRef]
- Le, K.; Mo, S.; Lu, X.; Ali, A.I.; Yu, D.; Guo, Y. Association of circulating blood HMGB1 levels with ischemic stroke: A systematic review and meta-analysis. Neurol. Res. 2018, 40, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Li, Q.; Han, H. HIF-1alpha in cerebral ischemia (Review). Mol. Med. Rep. 2022, 25, 41. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Mol. biology 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Taq Man Probes | Gene ID |
|---|---|---|
| IL1β | Ss03393804 | 397122 |
| IL1α | Ss03391335 | 397094 |
| IL6 | Ss03384604 | 399500 |
| IL8 | Ss03392437 | 396880 |
| IL10 | Ss03382372 | 397106 |
| IL17α | Ss03391803 | 449530 |
| RAGE | Ss03390846 | 396591 |
| HMGB1 | Ss03378573 | 445521 |
| TLR2 | Ss03381278 | 396623 |
| TLR4 | Ss03389780 | 399541 |
| TNF | Ss03391318 | 397086 |
| HiF1α | Ss03390447 | 396696 |
| PRDX1 | Ss06880613 | 100512476 |
| PRDX2 | Ss04327514 | 100512521 |
| PRDX4 | Ss06879099 | 100152260 |
| PRDX5 | Ss03394180 | 397273 |
| PRDX6 | Ss03384662 | 399538 |
| P2Y2 | Ss03378709 | 450248 |
| S100a8 | Ss04246257 | 100127488 |
| S100a9 | Ss04246618 | 100127489 |
| HSP60 | Ss01036749 | 492279 |
| HSPA6 | Ss03387784 | 396906 |
| HSP 70.2 | Ss03392270 | 396648 |
| NFKB1 | Ss03388575 | 751869 |
| NFKB2 | Ss06883748 | 100153829 |
| ENTPD1 (CD39) | Ss03394207 | 397298 |
| NT5E (CD73) | Ss06882629 | 100157995 |
| VEGFA | Ss03393993 | 397157 |
| ACTB | Ss03376563 | 414396 |
| GAPDH | Ss03375629 | 396823 |
| SDHA | Ss03376909 | 780433 |
| HPRT1 | Ss03388274 | 397351 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golubczyk, D.; Mowinska, A.; Holak, P.; Walczak, P.; Janowski, M.; Malysz-Cymborska, I. Complexity of Damage-Associated Molecular Pattern Molecule Expression Profile in Porcine Brain Affected by Ischemic Stroke. Int. J. Mol. Sci. 2025, 26, 3702. https://doi.org/10.3390/ijms26083702
Golubczyk D, Mowinska A, Holak P, Walczak P, Janowski M, Malysz-Cymborska I. Complexity of Damage-Associated Molecular Pattern Molecule Expression Profile in Porcine Brain Affected by Ischemic Stroke. International Journal of Molecular Sciences. 2025; 26(8):3702. https://doi.org/10.3390/ijms26083702
Chicago/Turabian StyleGolubczyk, Dominika, Aleksandra Mowinska, Piotr Holak, Piotr Walczak, Miroslaw Janowski, and Izabela Malysz-Cymborska. 2025. "Complexity of Damage-Associated Molecular Pattern Molecule Expression Profile in Porcine Brain Affected by Ischemic Stroke" International Journal of Molecular Sciences 26, no. 8: 3702. https://doi.org/10.3390/ijms26083702
APA StyleGolubczyk, D., Mowinska, A., Holak, P., Walczak, P., Janowski, M., & Malysz-Cymborska, I. (2025). Complexity of Damage-Associated Molecular Pattern Molecule Expression Profile in Porcine Brain Affected by Ischemic Stroke. International Journal of Molecular Sciences, 26(8), 3702. https://doi.org/10.3390/ijms26083702