
Development of Sheep Duodenum Intestinal Organoids and Implementation of High-Throughput Screening Platform for Veterinary Applications

 ,
,  ,
,  , , , ,
, , , ,  ,
, 
Abstract

1. Introduction
2. Results
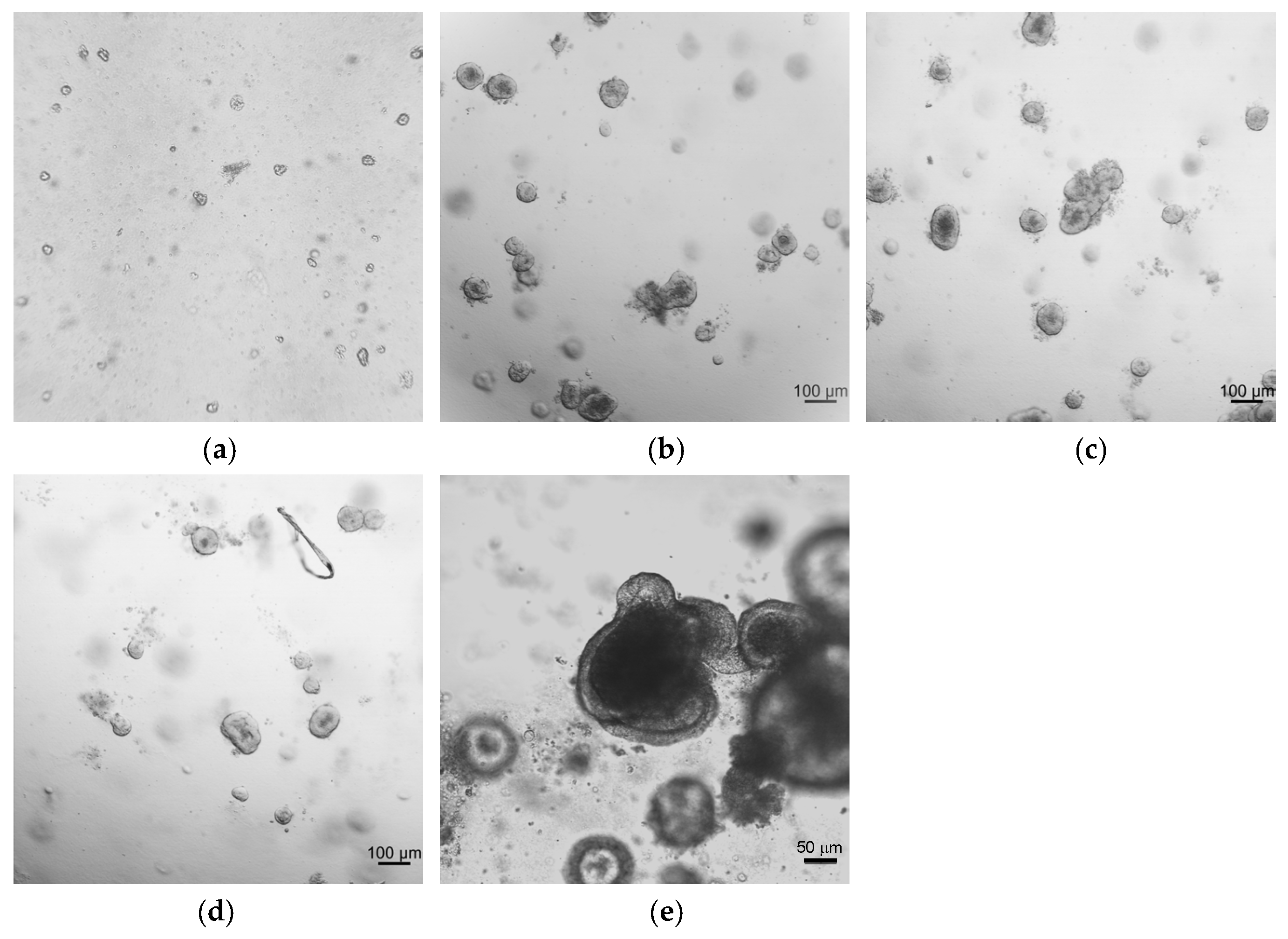
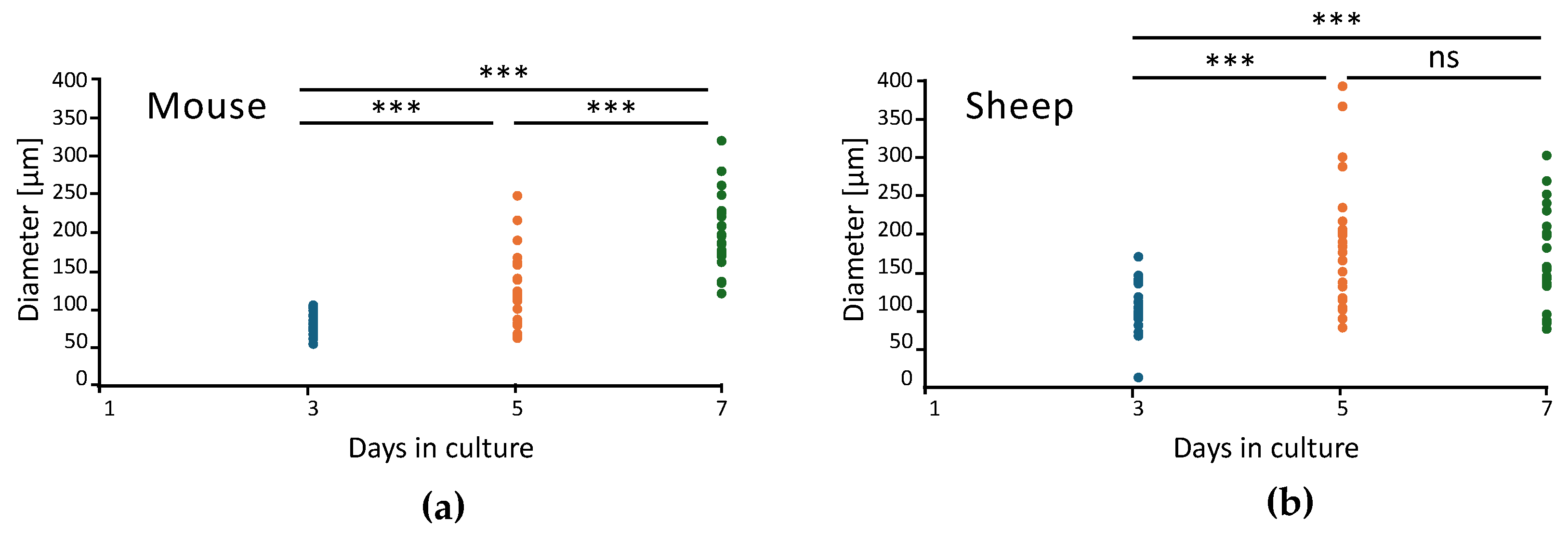
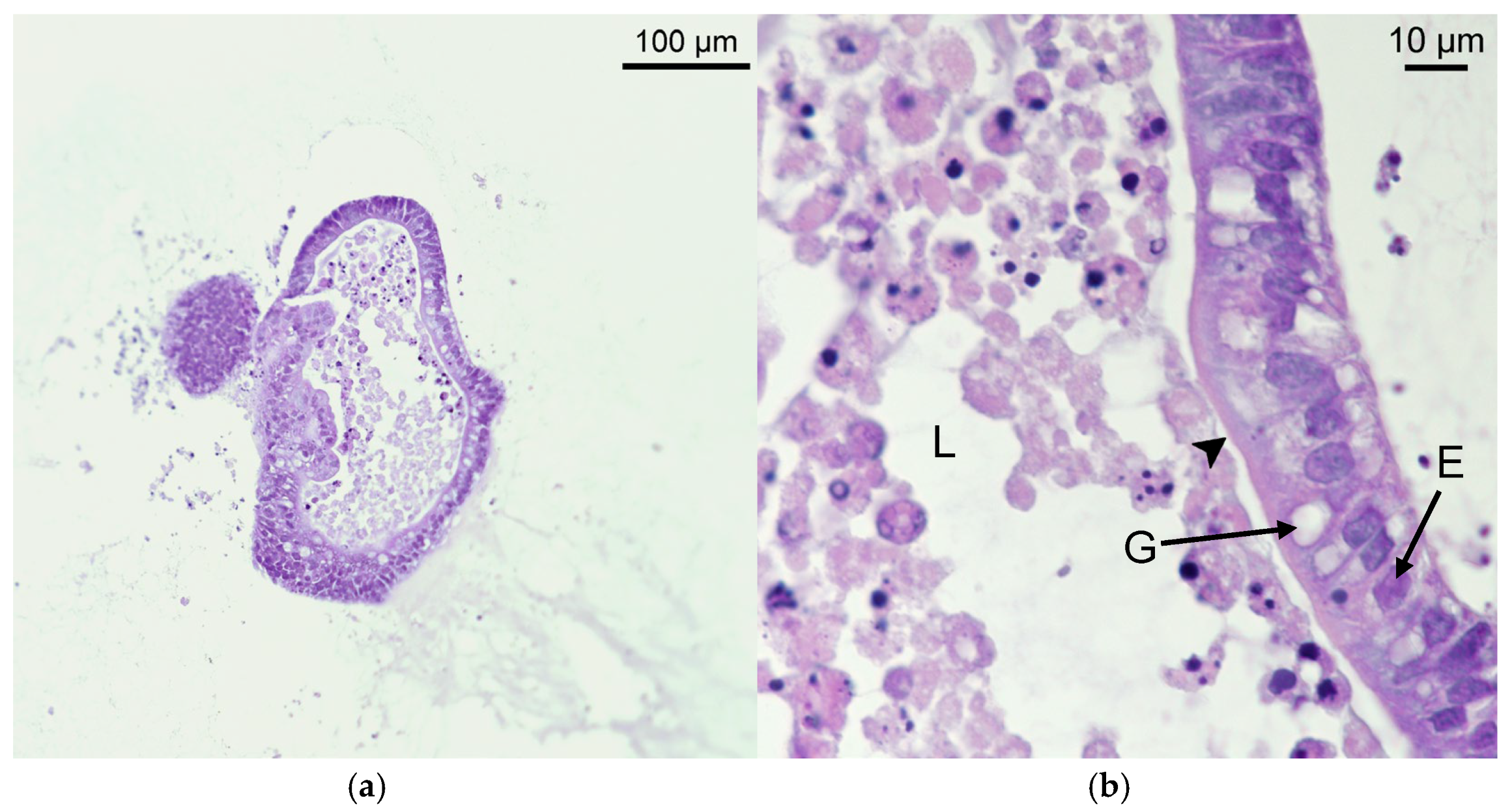
2.1. Development and Morphological Characterization of Sheep Duodenum Intestinal Organoids
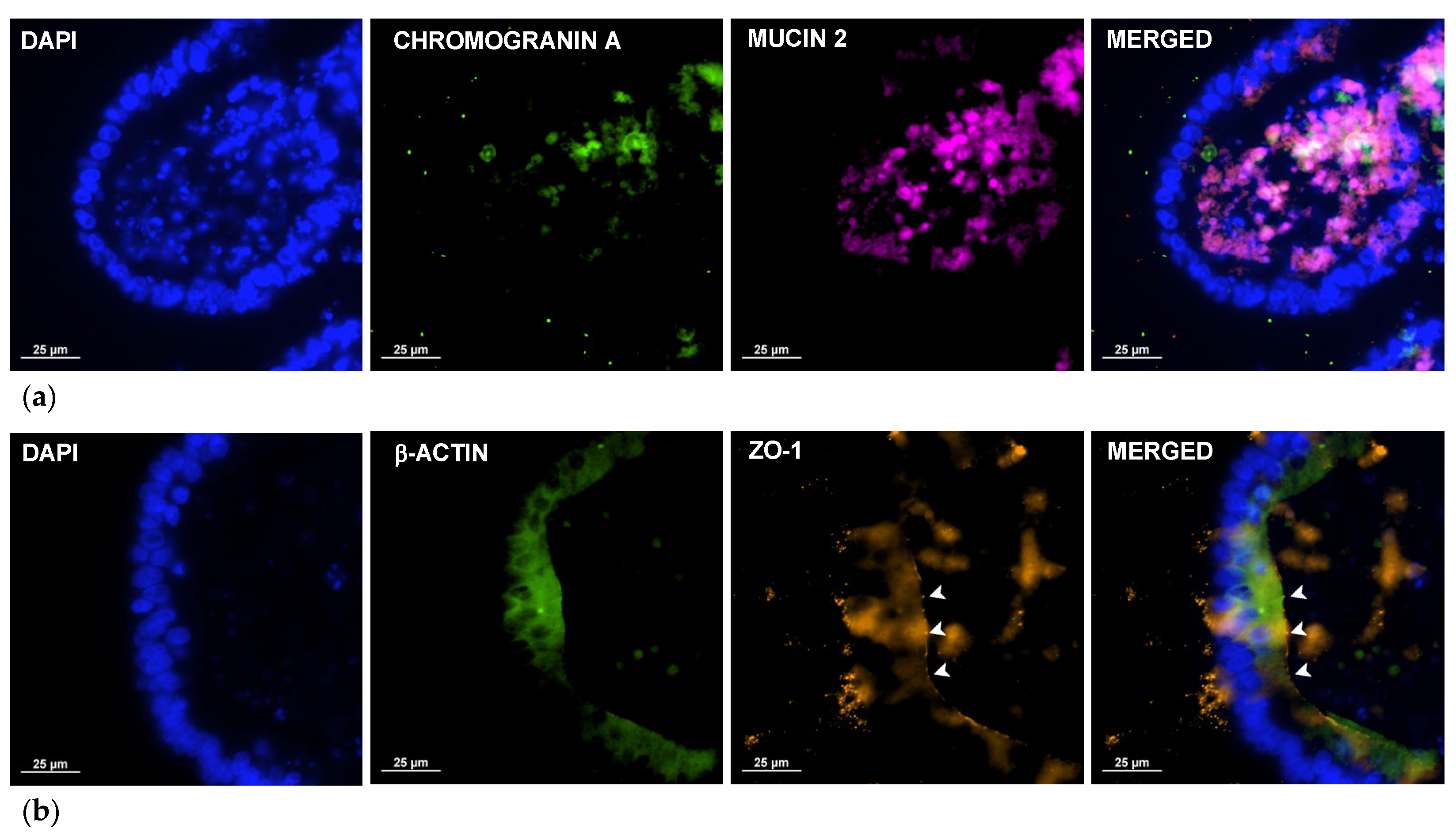
2.2. Immunocytochemistry Reveals Key Structures of Sheep Duodenum Intestinal Organoids
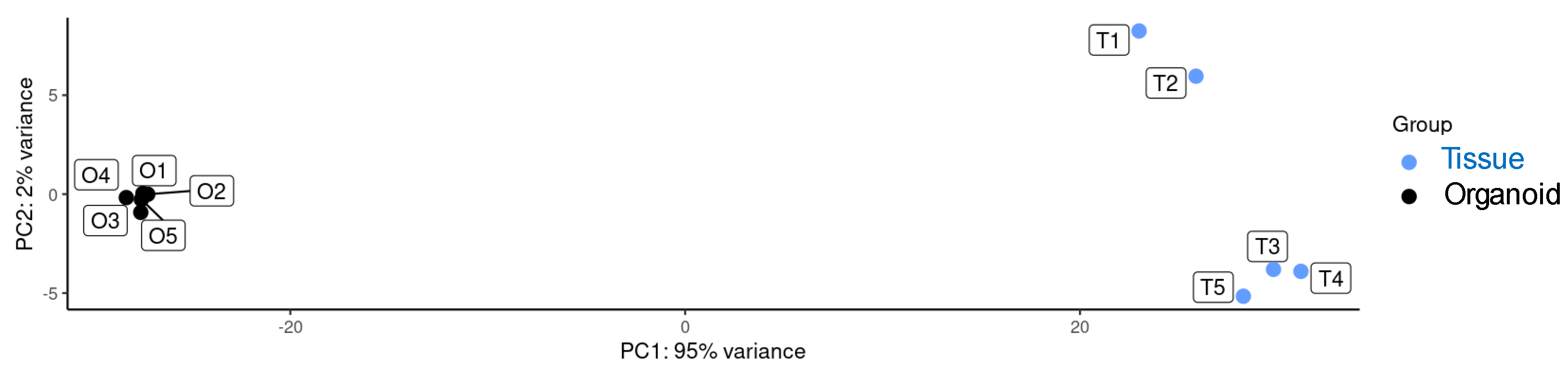
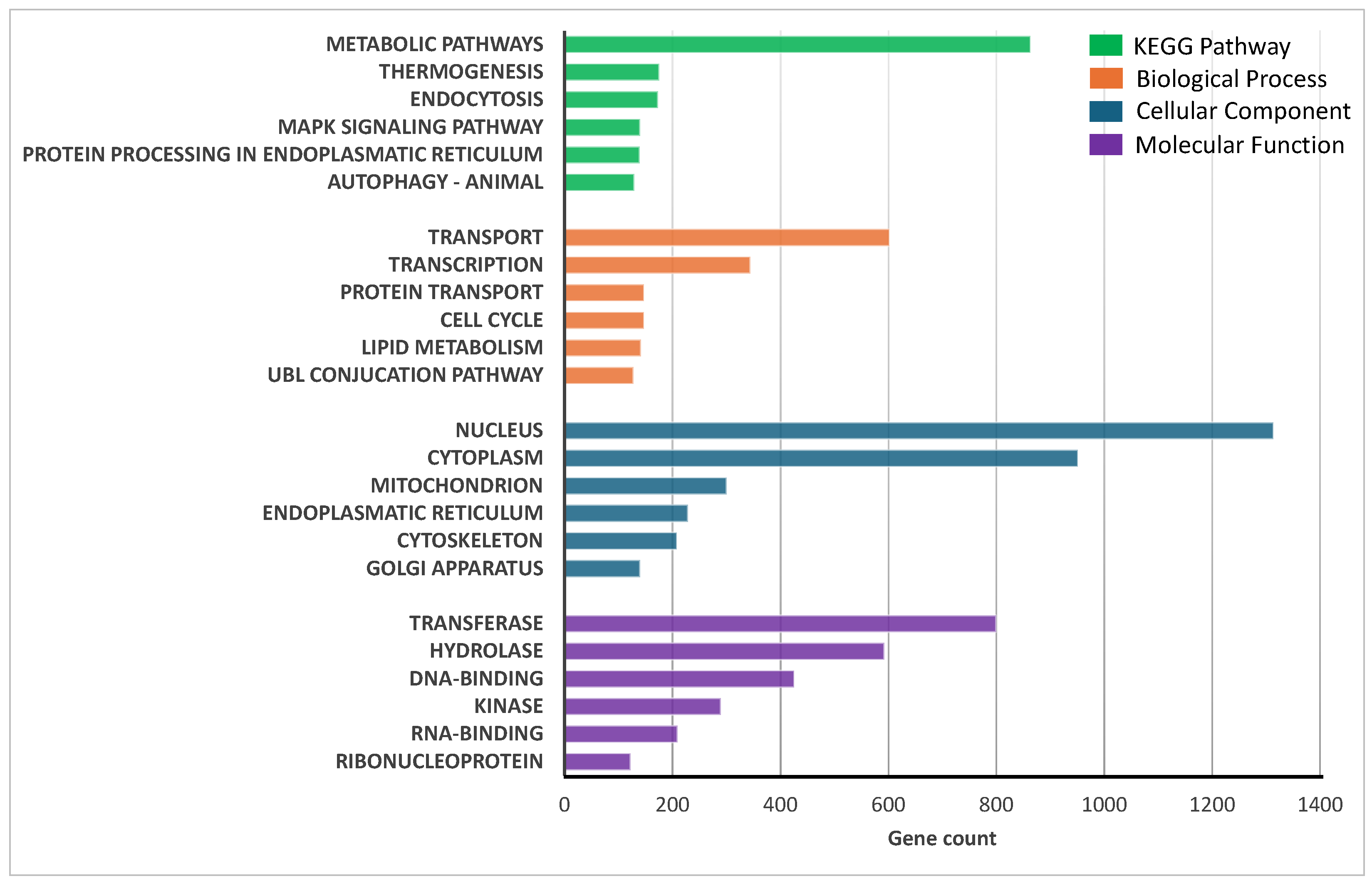
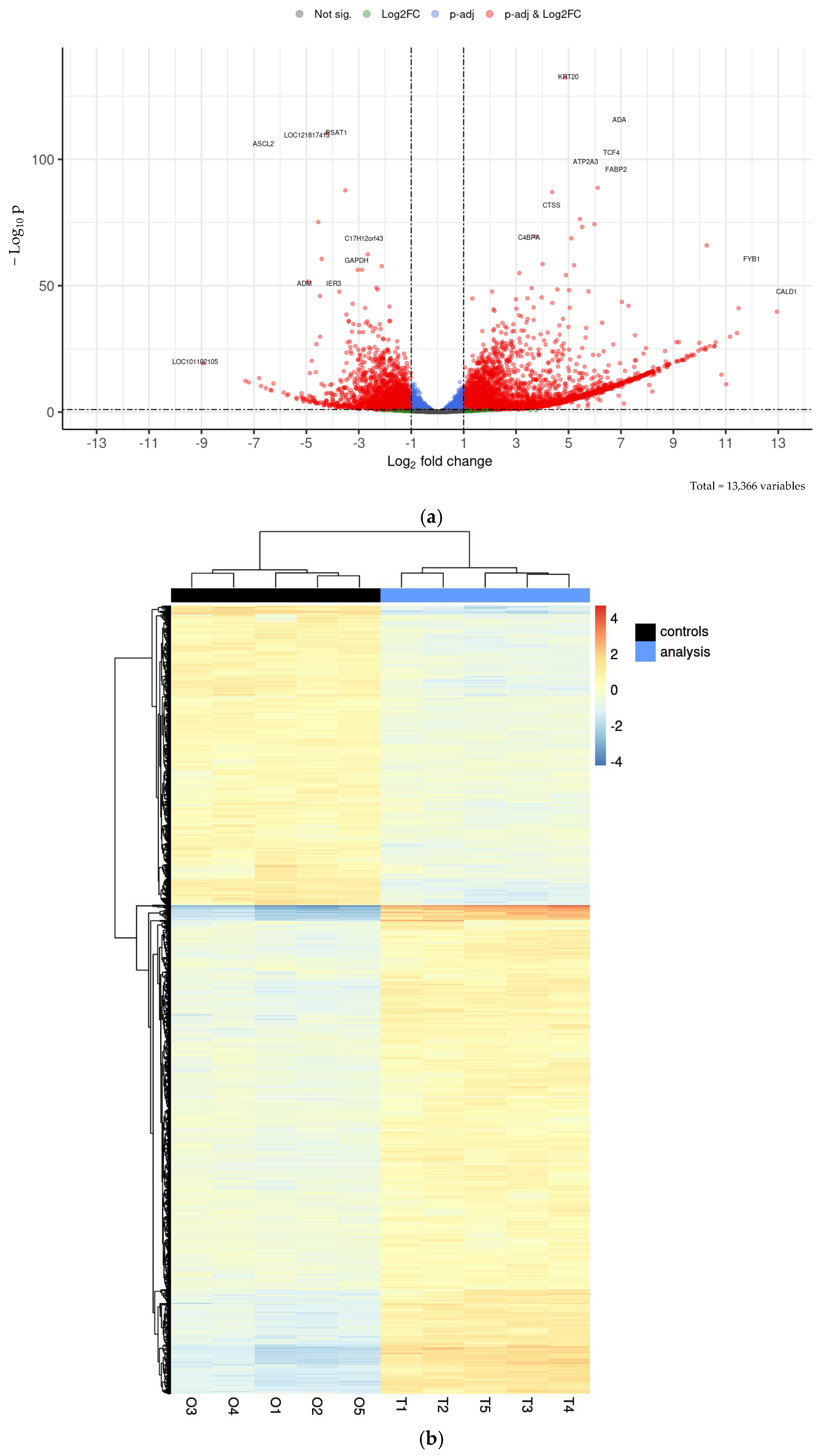
2.3. Transcriptional Analysis of Sheep Duodenum Intestinal Organoids and Expression of Cell- and Tissue-Specific Genes in Intestinal Organoids and Tissue
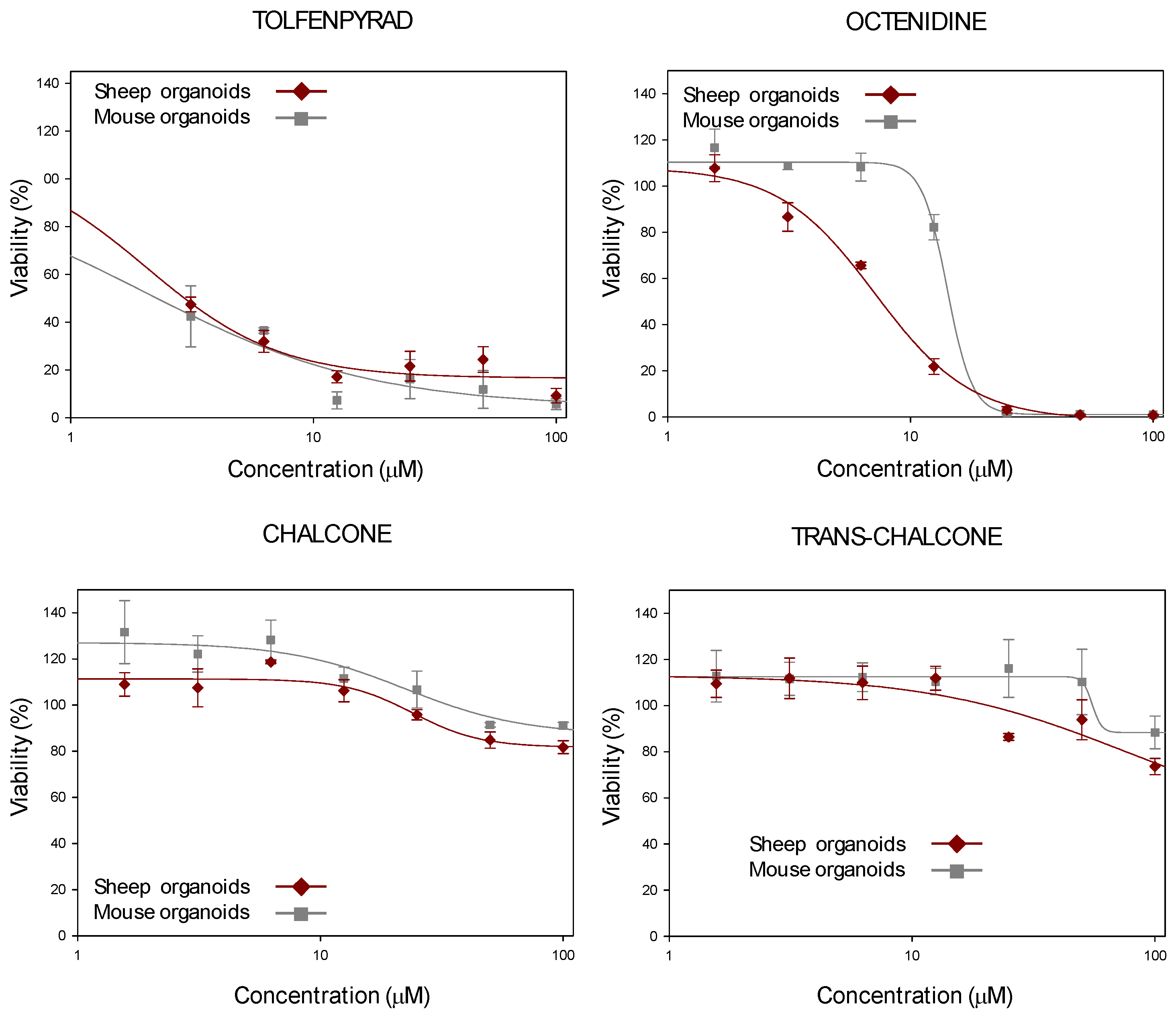
2.4. Development of HTS for Drug Discovery Purposes Based on Sheep Duodenum Organoids
3. Discussion
4. Materials and Methods
4.1. Preparation and Culture of Sheep Duodenum Intestinal Organoids and Mouse Intestinal Organoids
4.2. Histological Microscopy and Immunofluorescence Microscopy
4.3. RNAseq Analysis
4.4. Cytotoxicity Test in 384-Well Plates
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | Bidimensional |
| 3D | Three-dimensional |
| ADMET | Absorption, Distribution, Metabolism, Excretion, Toxicity |
| CC50 | Cytotoxic Concentration 50% |
| CES | Carboxylesterases |
| CYP | Cytochrome P450 |
| DMEs | Drug-metabolizing enzymes |
| H&E | Haematoxylin and eosin staining |
| HTS | High-throughput screening |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| R&D | Research and Development |
| UGT | Uridine diphosphate-glucuronosyltransferase |
References
- Kaplan, R.M.; Vidyashankar, A.N. An inconvenient truth: Global worming and anthelmintic resistance. Vet. Parasitol. 2012, 186, 70–78. [Google Scholar] [PubMed]
- Papadopoulos, E.; Gallidis, E.; Ptochos, S. Anthelmintic resistance in sheep in Europe: A selected review. Vet. Parasitol. 2012, 189, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Vázquez, F.A.; Hosking, B.C. A telephone survey of internal parasite control practices on sheep farms in Spain. Vet. Parasitol. 2013, 192, 166–172. [Google Scholar] [PubMed]
- Mazinani, M.; Rude, B. Population, world production and quality of sheep and goat products. Am. J. Anim. Vet. Sci. 2020, 15, 291–299. [Google Scholar] [CrossRef]
- McKellar, Q.A. The health of the sheep industry and the medicines to maintain it. Small Rumin. Res. 2006, 62, 7–12. [Google Scholar]
- Charlier, J.; Rinaldi, L.; Musella, V.; Ploeger, H.W.; Chartier, C.; Vineer, H.R.; Hinney, B.; von Samson-Himmelstjerna, G.; Băcescu, B.; Mickiewicz, M.; et al. Initial assessment of the economic burden of major parasitic helminth infections to the ruminant livestock industry in Europe. Prev. Vet. Med. 2020, 182, 105103. [Google Scholar]
- Charlier, J.; van der Voort, M.; Kenyon, F.; Skuce, P.; Vercruysse, J. Chasing helminths and their economic impact on farmed ruminants. Trends Parasitol. 2014, 30, 361–367. [Google Scholar]
- Martínez-Valladares, M.; Geurden, T.; Bartram, D.J.; Martínez-Pérez, J.M.; Robles-Pérez, D.; Bohórquez, A.; Flórez, E.; Meana, A.; Rojo-Vázquez, F.A. Resistance of gastrointestinal nematodes to the most commonly used anthelmintics in sheep, cattle and horses in Spain. Vet. Parasitol. 2015, 211, 228–233. [Google Scholar]
- Kaminsky, L.S.; Zhang, Q.-Y. The small intestine as a xenobiotic-metabolizing organ. Drug Metab. Dispos. 2003, 31, 1520–1525. [Google Scholar]
- Hedrich, W.D.; Panzica-Kelly, J.M.; Chen, S.-J.; Strassle, B.; Hasson, C.; Lecureux, L.; Wang, L.; Chen, W.; Sherry, T.; Gan, J.; et al. Development and characterization of rat duodenal organoids for ADME and toxicology applications. Toxicology 2020, 446, 152614. [Google Scholar]
- Lu, W.; Rettenmeier, E.; Paszek, M.; Yueh, M.-F.; Tukey, R.H.; Trottier, J.; Barbier, O.; Chen, S. Crypt organoid culture as an in vitro model in drug metabolism and cytotoxicity studies. Drug Metab. Dispos. 2017, 45, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, J.; Bertero, A.; Coccini, T.; Baderna, D.; Buzanska, L.; Caloni, F. Organoids are promising tools for species-specific in vitro toxicological studies. J. Appl. Toxicol. 2019, 39, 1610–1622. [Google Scholar] [CrossRef] [PubMed]
- Taelman, J.; Diaz, M.; Guiu, J. Human intestinal organoids: Promise and challenge. Front. Cell Dev. Biol. 2022, 10, 854740. [Google Scholar] [CrossRef]
- Tenreiro, M.F.; Branco, M.A.; Cotovio, J.P.; Cabral, J.M.S.; Fernandes, T.G.; Diogo, M.M. Advancing organoid design through co-emergence, assembly, and bioengineering. Trends Biotechnol. 2023, 41, 923–938. [Google Scholar] [CrossRef]
- Wang, Y.; Jeon, H. 3D cell cultures toward quantitative high-throughput drug screening. Trends Pharmacol. Sci. 2022, 43, 569–581. [Google Scholar] [CrossRef]
- Yang, S.; Hu, H.; Kung, H.; Zou, R.; Dai, Y.; Hu, Y.; Wang, T.; Lv, T.; Yu, J.; Li, F. Organoids: The current status and biomedical applications. MedComm 2023, 4, e274. [Google Scholar] [CrossRef]
- Borlak, J.; Zwadlo, C. Expression of drug-metabolizing enzymes, nuclear transcription factors and ABC transporters in Caco-2 cells. Xenobiotica 2003, 33, 927–943. [Google Scholar] [CrossRef]
- Pang, K.S. Modeling of intestinal drug absorption: Roles of transporters and metabolic enzymes (for the gillette review series). Drug Metab. Dispos. 2003, 31, 1507–1519. [Google Scholar] [CrossRef]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut 2015, 64, 911–920. [Google Scholar] [CrossRef]
- Iakobachvili, N.; Peters, P.J. Humans in a dish: The potential of organoids in modeling immunity and infectious diseases. Front. Microbiol. 2017, 8, 2402. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [PubMed]
- Moreira, L.; Bakir, B.; Chatterji, P.; Dantes, Z.; Reichert, M.; Rustgi, A.K. Pancreas 3D organoids: Current and future aspects as a research platform for personalized medicine in pancreatic cancer. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 289–298. [Google Scholar] [PubMed]
- Doherty, M.M.; Charman, W.N. The mucosa of the small intestine. Clin. Pharmacokinet. 2022, 41, 235–253. [Google Scholar]
- Fritz, A.; Busch, D.; Lapczuk, J.; Ostrowski, M.; Drozdzik, M.; Oswald, S. Expression of clinically relevant drug-metabolizing enzymes along the human intestine and their correlation to drug transporters and nuclear receptors: An intra-subject analysis. Basic Clin. Pharmacol. Toxicol. 2019, 124, 245–255. [Google Scholar]
- Beaumont, M.; Blanc, F.; Cherbuy, C.; Egidy, G.; Giuffra, E.; Lacroix-Lamandé, S.; Wiedemann, A. Intestinal organoids in farm animals. Vet. Res. 2021, 52, 33. [Google Scholar]
- Powell, R.H.; Behnke, M.S. WRN conditioned media is sufficient for in vitro propagation of intestinal organoids from large farm and small companion animals. Biol. Open 2017, 6, 698–705. [Google Scholar]
- Smith, D.; Price, D.R.G.; Burrells, A.; Faber, M.N.; Hildersley, K.A.; Chintoan-Uta, C.; Chapuis, A.F.; Stevens, M.; Stevenson, K.; Burgess, S.T.G.; et al. The development of ovine gastric and intestinal organoids for studying ruminant host-pathogen interactions. Front. Cell. Infect. Microbiol. 2021, 11, 733811. [Google Scholar]
- Melcón-Fernandez, E.; Martín-Encinas, E.; Palacios, F.; Galli, G.; Reguera, R.M.; Martínez-Valladares, M.; Balaña-Fouce, R.; Alonso, C.; Pérez-Pertejo, Y. Antileishmanial effect of 1,5- and 1,8-Substituted fused naphthyridines. Molecules 2023, 29, 74. [Google Scholar] [CrossRef]
- Melcón-Fernández, E.; Galli, G.; Balaña-Fouce, R.; García-Fernández, N.; Martínez-Valladares, M.; Reguera, R.M.; García-Estrada, C.; Pérez-Pertejo, Y. In vitro and ex vivo synergistic effect of pyrvinium pamoate combined with miltefosine and paromomycin against Leishmania. Trop. Med. Infect. Dis. 2024, 9, 30. [Google Scholar] [CrossRef]
- Galli, G.; Ruiz-Somacarrera, M.; González del Palacio, L.; Melcón-Fernández, E.; González-Pérez, R.; García-Estrada, C.; Martinez-Valladares, M.; Balaña-Fouce, R. High-Throughput Screening of five compound libraries for anthelmintic activity and toxicity leads to the discovery of two flavonoid compounds. Int. J. Mol. Sci. 2025, 26, 1595. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [PubMed]
- Wang, H.; He, X.; Zhang, M.; Fan, N.; Yang, Z.; Shen, T.; Guo, J.; Song, Y.; Cao, G.; Liu, Y.; et al. Development of sheep intestinal organoids for studying deoxynivalenol-induced toxicity. Int. J. Mol. Sci. 2025, 26, 955. [Google Scholar] [CrossRef] [PubMed]
- Co, J.Y.; Margalef-Català, M.; Li, X.; Mah, A.T.; Kuo, C.J.; Monack, D.M.; Amieva, M.R. Controlling epithelial polarity: A human enteroid model for host-pathogen interactions. Cell Rep. 2019, 26, 2509–2520.e4. [Google Scholar] [PubMed]
- Chan, C.W.; Wong, N.A.; Liu, Y.; Bicknell, D.; Turley, H.; Hollins, L.; Miller, C.J.; Wilding, J.L.; Bodmer, W.F. Gastrointestinal differentiation marker Cytokeratin 20 is regulated by homeobox gene CDX1. Proc. Natl. Acad. Sci. USA 2009, 106, 1936–1941. [Google Scholar]
- van Es, J.H.; Haegebarth, A.; Kujala, P.; Itzkovitz, S.; Koo, B.-K.; Boj, S.F.; Korving, J.; van den Born, M.; van Oudenaarden, A.; Robine, S.; et al. A critical role for the Wnt effector Tcf4 in adult intestinal homeostatic self-renewal. Mol. Cell. Biol. 2012, 32, 1918–1927. [Google Scholar]
- Xing, Z.; Li, X.; He, J.; Chen, Y.; Zhu, L.; Zhang, X.; Huang, Z.; Tang, J.; Guo, Y.; He, Y. OLFM4 modulates intestinal inflammation by promoting IL-22+ILC3 in the gut. Commun. Biol. 2024, 7, 914. [Google Scholar]
- Bradford, K.L.; Moretti, F.A.; Carbonaro-Sarracino, D.A.; Gaspar, H.B.; Kohn, D.B. Adenosine deaminase (ADA)-deficient severe combined immune deficiency (SCID): Molecular pathogenesis and clinical manifestations. J. Clin. Immunol. 2017, 37, 626–637. [Google Scholar]
- Huang, X.; Zhou, Y.; Sun, Y.; Wang, Q. Intestinal fatty acid binding protein: A rising therapeutic target in lipid metabolism. Prog. Lipid Res. 2022, 87, 101178. [Google Scholar]
- Vié, N.; Copois, V.; Bascoul-Mollevi, C.; Denis, V.; Bec, N.; Robert, B.; Fraslon, C.; Conseiller, E.; Molina, F.; Larroque, C.; et al. Overexpression of phosphoserine aminotransferase PSAT1 stimulates cell growth and increases chemoresistance of colon cancer cells. Mol. Cancer 2008, 7, 14. [Google Scholar]
- Schuijers, J.; Junker, J.P.; Mokry, M.; Hatzis, P.; Koo, B.-K.; Sasselli, V.; van der Flier, L.G.; Cuppen, E.; van Oudenaarden, A.; Clevers, H. Ascl2 acts as an R-spondin/Wnt-responsive switch to control stemness in intestinal crypts. Cell Stem Cell 2015, 16, 158–170. [Google Scholar]
- Miller, S.C.; MacDonald, C.C.; Kellogg, M.K.; Karamysheva, Z.N.; Karamyshev, A.L. Specialized ribosomes in health and disease. Int. J. Mol. Sci. 2023, 24, 6334. [Google Scholar] [CrossRef] [PubMed]
- Welin, M.; Egeblad, L.; Johansson, A.; Stenmark, P.; Wang, L.; Flodin, S.; Nyman, T.; Trésaugues, L.; Kotenyova, T.; Johansson, I.; et al. Structural and functional studies of the human phosphoribosyltransferase domain containing protein 1. FEBS J. 2010, 277, 4920–4930. [Google Scholar] [PubMed]
- Cruz-Rojo, M.A.; Martínez-Valladares, M.; Álvarez-Sánchez, M.A.; Rojo-Vázquez, F.A. Effect of infection with Teladorsagia circumcincta on milk production and composition in Assaf dairy sheep. Vet. Parasitol. 2012, 185, 194–200. [Google Scholar]
- Martínez-Valladares, M.; Martín-Ramos, E.; Esteban-Ballesteros, M.; Balaña-Fouce, R.; Rojo-Vázquez, F.A. Effect of level of infection by gastrointestinal nematodes and anthelmintic treatment on milk yield in dairy sheep. Parasite 2021, 28, 71. [Google Scholar]
- Gopallawa, I.; Gupta, C.; Jawa, R.; Cyril, A.; Jawa, V.; Chirmule, N.; Gujar, V. Applications of organoids in advancing drug discovery and development. J. Pharm. Sci. 2024, 113, 2659–2667. [Google Scholar]
- Banstola, A.; Reynolds, J.N.J. The sheep as a large animal model for the investigation and treatment of human disorders. Biology 2022, 11, 1251. [Google Scholar] [CrossRef]
- Stankiewicz, M.; Pernthaner, A.; Cabaj, W.; Jonas, W.E.; Douch, P.G.; Bisset, S.A.; Rabel, B.; Pfeffer, A.; Green, R.S. Immunization of sheep against parasitic nematodes leads to elevated levels of globule leukocytes in the small intestine lumen. Int. J. Parasitol. 1995, 25, 389–394. [Google Scholar]
- Keane, O.M.; Zadissa, A.; Wilson, T.; Hyndman, D.L.; Greer, G.J.; Baird, D.B.; McCulloch, A.F.; Crawford, A.M.; McEwan, J.C. Gene expression profiling of naïve sheep genetically resistant and susceptible to gastrointestinal nematodes. BMC Genom. 2006, 7, 42. [Google Scholar]
- Makovcová, K.; Langrová, I.; Vadlejch, J.; Jankovská, I.; Lytvynets, A.; Borkovcová, M. Linear distribution of nematodes in the gastrointestinal tract of tracer lambs. Parasitol. Res. 2008, 104, 123–126. [Google Scholar]
- Szymański, P.; Markowicz, M.; Mikiciuk-Olasik, E. Adaptation of high-throughput screening in drug discovery—toxicological screening tests. Int. J. Mol. Sci. 2012, 13, 427–452. [Google Scholar]
- Li, L.; Zhou, Q.; Voss, T.C.; Quick, K.L.; LaBarbera, D.V. High-throughput imaging: Focusing in on drug discovery in 3D. Methods 2016, 96, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Taki, A.C.; Byrne, J.J.; Wang, T.; Sleebs, B.E.; Nguyen, N.; Hall, R.S.; Korhonen, P.K.; Chang, B.C.H.; Jackson, P.; Jabbar, A.; et al. High-Throughput phenotypic assay to screen for anthelmintic activity on Haemonchus contortus. Pharmaceuticals 2021, 14, 616. [Google Scholar] [CrossRef]
- Bhat, A.H.; Tak, H.; Malik, I.M.; Ganai, B.A.; Zehbi, N. Trichostrongylosis: A zoonotic disease of small ruminants. J. Helminthol. 2023, 97, e26. [Google Scholar] [CrossRef] [PubMed]
- Xuan, L.; Luo, J.; Qu, C.; Guo, P.; Yi, W.; Yang, J.; Yan, Y.; Guan, H.; Zhou, P.; Huang, R. Predictive metabolomic signatures for safety assessment of three plastic nanoparticles using intestinal organoids. Sci. Total. Environ. 2024, 913, 169606. [Google Scholar] [CrossRef] [PubMed]
- Almeqdadi, M.; Mana, M.D.; Roper, J.; Yilmaz, Ö.H. Gut organoids: Mini-tissues in culture to study intestinal physiology and disease. Am. J. Physiol. Cell Physiol. 2019, 317, C405–C419. [Google Scholar] [CrossRef]
- Shuhendler, A.J.; Prasad, P.; Cai, P.; Hui, K.K.W.; Henderson, J.T.; Rauth, A.M.; Wu, X.Y. Matrigel alters the pathophysiology of orthotopic human breast adenocarcinoma xenografts with implications for nanomedicine evaluation. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 795–805. [Google Scholar] [CrossRef]
- Dunn, D.A.; Feygin, I. Challenges and solutions to ultra-high-throughput screening assay miniaturization: Submicroliter fluid handling. Drug Discov. Today 2000, 5, 84–91. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kumar, R.A.; Sukumaran, S.M.; Hogg, M.G.; Clark, D.S.; Dordick, J.S. Three-dimensional cellular microarray for high-throughput toxicology assays. Proc. Natl. Acad. Sci. USA 2008, 105, 59–63. [Google Scholar] [CrossRef]
- Document #28223, Version 2.0.1, December 2016. Stemcell Enteroids Tech Bulletin. Available online: https://cdn.stemcell.com/media/files/techbulletin/TB28223-Intestinal_Epithelial_Organoid_Culture_with_IntestiCult_Organoid_Growth_Medium_(Mouse).pdf?_ga=1.255806536.1701332796.1451577632 (accessed on 6 September 2022).
- Fujii, E.; Yamazaki, M.; Kawai, S.; Ohtani, Y.; Watanabe, T.; Kato, A.; Suzuki, M. A simple method for histopathological evaluation of organoids. J. Toxicol. Pathol. 2018, 31, 81–85. [Google Scholar] [CrossRef]
- Du, Y.; Li, X.; Niu, Q.; Mo, X.; Qui, M.; Ma, T.; Kuo, C.J.; Fu, H. Development of a miniaturized 3D organoid culture platform for ultra-high-throughput screening. J. Mol. Cell Biol. 2020, 12, 630–643. [Google Scholar] [CrossRef]
- Iversen, P.W.; Eastwood, B.J.; Sittampalam, G.S.; Cox, K.L. A comparison of assay performance measures in screening assays: Signal window, Z′ factor, and assay variability ratio. J. Biomol. Screen. 2006, 11, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-H.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of High Throughput Screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [PubMed]
- Malo, N.; Hanley, J.A.; Cerquozzi, S.; Pelletier, J.; Nadon, R. Statistical practice in high-throughput screening data analysis. Nat. Biotechnol. 2006, 24, 167–175. [Google Scholar] [PubMed]
- Sui, Y.; Wu, Z. Alternative statistical parameter for High-Throughput Screening assay quality assessment. J. Biomol. Screen. 2007, 12, 229–234. [Google Scholar] [CrossRef]
- Thorne, N.; Auld, D.S.; Inglese, J. Apparent activity in high-throughput screening: Origins of compound-dependent assay interference. Curr. Opin. Chem. Biol. 2010, 14, 315–324. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metrics 1 | P1 | P2 | P3 | P4 | Average |
|---|---|---|---|---|---|
| S/B | 36 | 89 | 85 | 37 | 61 |
| S/N | 54 | 259 | 169 | 26 | 127 |
| Z’ | 0.7 | 0.4 | 0.5 | 0.6 | 0.5 |
| SW | 7.0 | 2.0 | 3.5 | 5.2 | 4.4 |
| AVR | 0.3 | 0.6 | 0.5 | 0.4 | 0.5 |
| CC50 Values (μM) | ||
|---|---|---|
| Sheep Intestinal Organoids | Mouse Intestinal Organoids 1 | |
| Tolfenpyrad | <3 | <1 |
| Octenidine | 7.2 ± 0.2 | 14.2 ± 0.41 |
| Chalcone | >50 | >50 |
| Trans-chalcone | >50 | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galli, G.; Melcón-Fernández, E.; de Garnica García, M.G.; Martínez-Fernández, B.; Dehnavi, M.; Andrés, S.; Pérez-Pertejo, Y.; Reguera, R.M.; García-Estrada, C.; Martínez-Valladares, M.; et al. Development of Sheep Duodenum Intestinal Organoids and Implementation of High-Throughput Screening Platform for Veterinary Applications. Int. J. Mol. Sci. 2025, 26, 3452. https://doi.org/10.3390/ijms26073452
Galli G, Melcón-Fernández E, de Garnica García MG, Martínez-Fernández B, Dehnavi M, Andrés S, Pérez-Pertejo Y, Reguera RM, García-Estrada C, Martínez-Valladares M, et al. Development of Sheep Duodenum Intestinal Organoids and Implementation of High-Throughput Screening Platform for Veterinary Applications. International Journal of Molecular Sciences. 2025; 26(7):3452. https://doi.org/10.3390/ijms26073452
Chicago/Turabian StyleGalli, Giulio, Estela Melcón-Fernández, María Gracia de Garnica García, Beatriz Martínez-Fernández, Mahsa Dehnavi, Sonia Andrés, Yolanda Pérez-Pertejo, Rosa M. Reguera, Carlos García-Estrada, María Martínez-Valladares, and et al. 2025. "Development of Sheep Duodenum Intestinal Organoids and Implementation of High-Throughput Screening Platform for Veterinary Applications" International Journal of Molecular Sciences 26, no. 7: 3452. https://doi.org/10.3390/ijms26073452
APA StyleGalli, G., Melcón-Fernández, E., de Garnica García, M. G., Martínez-Fernández, B., Dehnavi, M., Andrés, S., Pérez-Pertejo, Y., Reguera, R. M., García-Estrada, C., Martínez-Valladares, M., & Balaña-Fouce, R. (2025). Development of Sheep Duodenum Intestinal Organoids and Implementation of High-Throughput Screening Platform for Veterinary Applications. International Journal of Molecular Sciences, 26(7), 3452. https://doi.org/10.3390/ijms26073452