
Annexin–Membrane Interactions Across Eukaryotic Domains of Life—A Comparative Approach


Abstract
1. Introduction
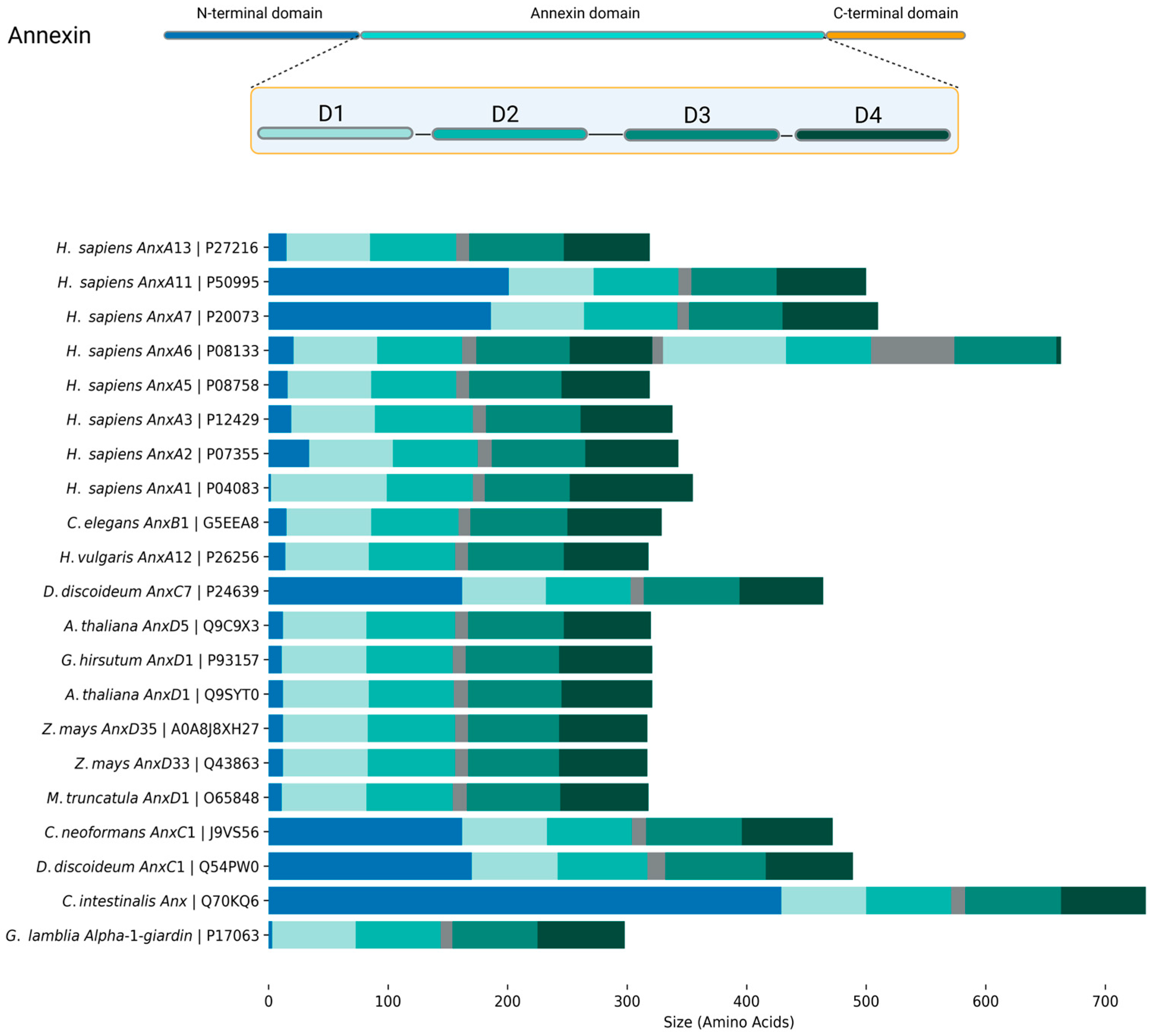
2. Annexins: The Basics
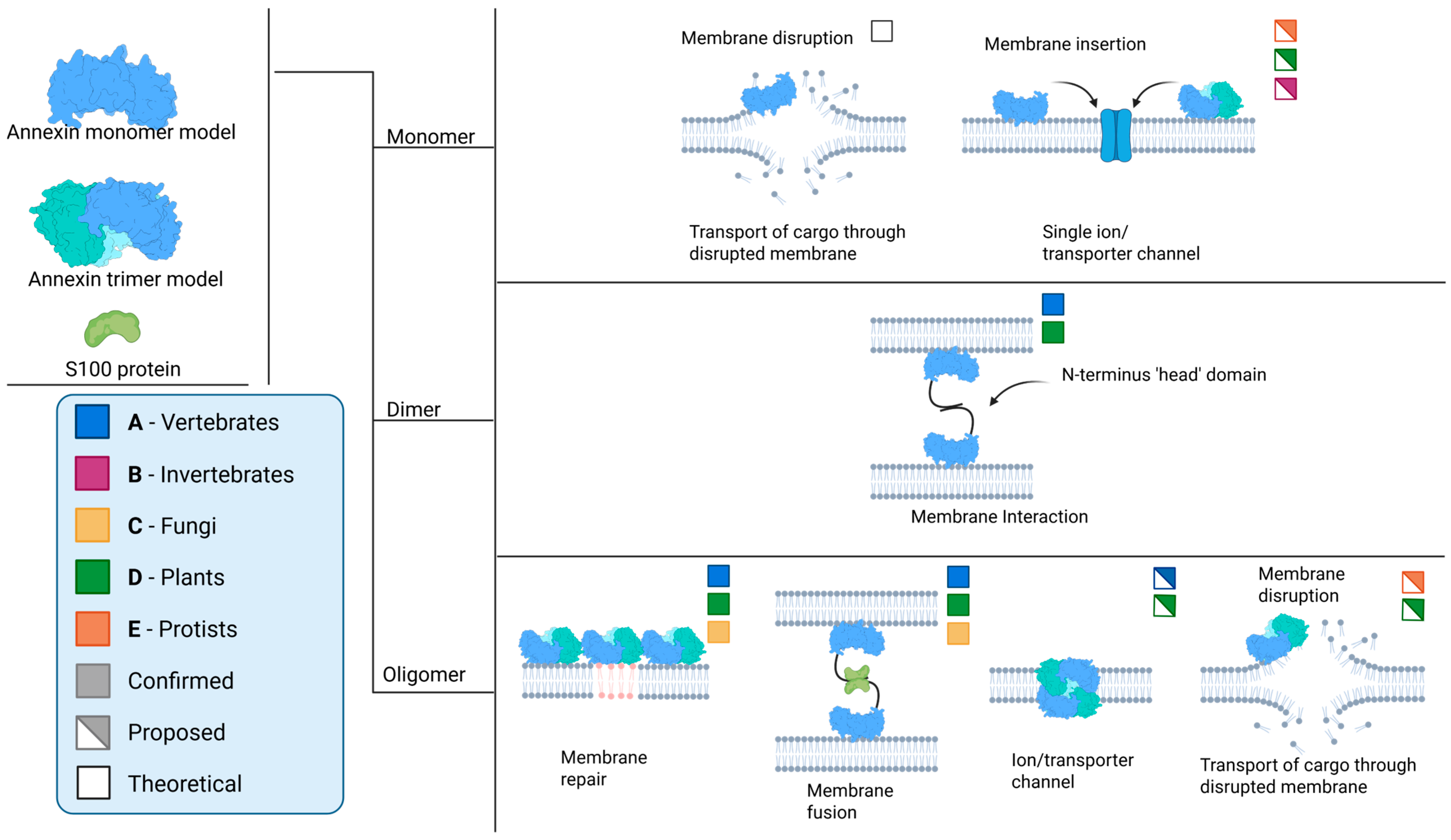
3. Annexin–Membrane Interactions: Modes and Functions
3.1. Annexins and Membrane Repair
3.1.1. Mammalian Cell Systems
3.1.2. Plant Cell Systems
3.1.3. Other Cell Systems
3.2. Annexins as Calcium Ion Channels
3.2.1. Mammalian Cell Systems
3.2.2. Plant Cell Systems
3.2.3. The Debate Around Annexins as True Ca2+ Ion Channels
3.3. Annexins and Protein Trafficking
3.3.1. Mammalian Cell Systems
3.3.2. Plant Cell Systems
3.3.3. Other Cell Systems
3.4. Membrane Traversal
4. Regulation of Annexin Function and Membrane Interaction
4.1. The Role of the N-Terminal Head Region
4.2. Calcium-Dependent Regulation and Notable Exceptions
4.3. Oligomerization
5. Open Questions and Future Research Directions
Author Contributions
Funding
Conflicts of Interest
References
- Gatenby, R.A. The Role of Cell Membrane Information Reception, Processing, and Communication in the Structure and Function of Multicellular Tissue. Int. J. Mol. Sci. 2019, 20, 3609. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Moss, S.E. Annexins: From Structure to Function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef]
- Moss, S.E.; Morgan, R.O. The annexins. Genome Biol. 2004, 5, 219. [Google Scholar] [CrossRef]
- Kodavali, P.K.; Dudkiewicz, M.; Pikuła, S.; Pawłowski, K. Bioinformatics Analysis of Bacterial Annexins—Putative Ancestral Relatives of Eukaryotic Annexins. PLoS ONE 2014, 9, e85428. [Google Scholar] [CrossRef]
- Liemann, S.; Lewit-Bentley, A. Annexins: A novel family of calcium-and membrane-binding proteins in search of a function. Structure 1995, 3, 233–237. [Google Scholar] [CrossRef]
- Mo, Y.; Campos, B.; Mealy, T.R.; Commodore, L.; Head, J.F.; Dedman, J.R.; Seaton, B.A. Interfacial basic cluster in annexin V couples phospholipid binding and trimer formation on membrane surfaces. J. Biol. Chem. 2003, 278, 2437–2443. [Google Scholar] [CrossRef]
- Wang, S.; Sun, H.; Tanowitz, M.; Liang, X.; Crooke, S.T. Annexin A2 facilitates endocytic trafficking of antisense oligonucleotides. Nucleic. Acids. Res. 2016, 44, 7314–7330. [Google Scholar] [CrossRef]
- Tcatchoff, L.; Andersson, S.; Utskarpen, A.; Klokk, T.I.; Skånland, S.S.; Pust, S.; Gerke, V.; Sandvig, K. Annexin A1 and A2: Roles in Retrograde Trafficking of Shiga Toxin. PLoS ONE 2012, 7, e40429. [Google Scholar] [CrossRef]
- Isas, J.M.; Cartailler, J.-P.; Sokolov, Y.; Patel, D.R.; Langen, R.; Luecke, H.; Hall, J.E.; Haigler, H.T. Annexins V and XII Insert into Bilayers at Mildly Acidic pH and Form Ion Channels. Biochemistry 2000, 39, 3015–3022. [Google Scholar] [CrossRef]
- Huber, R.; Schneider, M.; Mayr, I.; Römisch, J.; Paques, E.-P. The calcium binding sites in human annexin V by crystal structure analysis at 2.0 A resolution Implications for membrane binding and calcium channel activity. FEBS Lett. 1990, 275, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Berendes, R.; Voges, D.; Demange, P.; Huber, R.; Burger, A. Structure-Function Analysis of the Ion Channel Selectivity Filter in Human Annexin V. Science 1993, 262, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Golczak, M.; Kicinska, A.; Bandorowicz-Pikula, J.; Buchet, R.; Szewczyk, A.; Pikula, S. Acidic pH-induced folding of annexin VI is a prerequisite for its insertion into lipid bilayers and formation of ion channels by the protein molecules. FASEB J. 2001, 15, 1083–1085. [Google Scholar] [CrossRef]
- Balmer, E.A.; Wirdnam, C.D.; Faso, C. A core UPS molecular complement implicates unique endocytic compartments at the parasite–host interface in Giardia lamblia. Virulence 2023, 14, 2174288. [Google Scholar] [CrossRef]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef]
- Al-Ali, H.N.; Crichton, S.J.; Fabian, C.; Pepper, C.; Butcher, D.R.; Dempsey, F.C.; Parris, C.N. A therapeutic antibody targeting annexin-A1 inhibits cancer cell growth in vitro and in vivo. Oncogene 2024, 43, 608–614. [Google Scholar] [CrossRef]
- Yan, Z.; Cheng, X.; Wang, T.; Hong, X.; Shao, G.; Fu, C. Therapeutic potential for targeting Annexin A1 in fibrotic diseases. Genes Dis. 2022, 9, 1493–1505. [Google Scholar] [CrossRef]
- Wang, X.; Shao, G.; Hong, X.; Shi, Y.; Zheng, Y.; Yu, Y.; Fu, C. Targeting Annexin A1 as a Druggable Player to Enhance the Anti-Tumor Role of Honokiol in Colon Cancer through Autophagic Pathway. Pharmaceuticals 2023, 16, 70. [Google Scholar] [CrossRef]
- Li, Z.; Yu, L.; Hu, B.; Chen, L.; Jv, M.; Wang, L.; Zhou, C.; Wei, M.; Zhao, L. Advances in cancer treatment: A new therapeutic target, Annexin A2. J. Cancer 2021, 12, 3587–3596. [Google Scholar] [CrossRef]
- Geisow, M.J.; Walker, J.H.; Boustead, C.; Taylor, W. Annexins—New family of Ca2+-regulated-phospholipid binding protein. Biosci. Rep. 1987, 7, 289–298. [Google Scholar] [CrossRef]
- Creutz, C.E.; Pazoles, C.J.; Pollard, H.B. Identification and purification of an adrenal medullary protein (synexin) that causes calcium-dependent aggregation of isolated chromaffin granules. J. Biol. Chem. 1978, 253, 2858–2866. [Google Scholar] [CrossRef] [PubMed]
- Flower, R.J. Background and discovery of lipocortins. Agents Actions 1986, 17, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-S.; Wallner, B.P.; Mattaliano, R.J.; Tizard, R.; Burne, C.; Frey, A.; Hession, C.; McGray, P.; Sinclair, L.K.; Chow, E.P.; et al. Two human 35 kd inhibitors of phospholipase A2 are related to substrates of pp60v-src and of the epidermal growth factor receptor/kinase. Cell 1986, 46, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Saris, C.J.M.; Tack, B.F.; Kristensen, T.; Glenney, J.R.; Hunter, T. The cDNA sequence for the protein-tyrosine kinase substrate p36 (calpactin I heavy chain) reveals a multidomain protein with internal repeats. Cell 1986, 46, 201–212. [Google Scholar] [CrossRef]
- Glenney, J.R.; Tack, B.; Powell, M.A. Calpactins: Two distinct Ca++-regulated phospholipid- and actin-binding proteins isolated from lung and placenta. J. Cell Biol. 1987, 104, 503–511. [Google Scholar] [CrossRef]
- Gerke, V.; Gavins, F.N.E.; Geisow, M.; Grewal, T.; Jaiswal, J.K.; Nylandsted, J.; Rescher, U. Annexins—A family of proteins with distinctive tastes for cell signaling and membrane dynamics. Nat. Commun. 2024, 15, 1574. [Google Scholar] [CrossRef]
- Bitto, E.; Li, M.; Tikhonov, A.M.; Schlossman, M.L.; Cho, W. Mechanism of Annexin I-Mediated Membrane Aggregation. Biochemistry 2000, 39, 13469–13477. [Google Scholar] [CrossRef]
- Langen, R.; Isas, J.M.; Hubbell, W.L.; Haigler, H.T. A transmembrane form of annexin XII detected by site-directed spin labeling. Proc. Natl. Acad. Sci. USA 1998, 95, 14060–14065. [Google Scholar] [CrossRef]
- Ladokhin, A.S.; Haigler, H.T. Reversible Transition between the Surface Trimer and Membrane-Inserted Monomer of Annexin 12. Biochemistry 2005, 44, 3402–3409. [Google Scholar] [CrossRef]
- Oling, F.; Bergsma-Schutter, W.; Brisson, A. Trimers, dimers of trimers, and trimers of trimers are common building blocks of annexin A5 two-dimensional crystals. J. Struct. Biol. 2001, 133, 55–63. [Google Scholar] [CrossRef]
- Ayala-Sanmartin, J.; Zibouche, M.; Illien, F.; Vincent, M.; Gallay, J. Insight into the location and dynamics of the annexin A2 N-terminal domain during Ca2+-induced membrane bridging. Biochim. Biophys. Acta—Biomembr. 2008, 1778, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, J.C.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.H.; Davies, J.M. Annexins: Multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Posokhov, Y.O.; Rodnin, M.V.; Lu, L.; Ladokhin, A.S. Membrane Insertion Pathway of Annexin B12: Thermodynamic and Kinetic Characterization by Fluorescence Correlation Spectroscopy and Fluorescence Quenching. Biochemistry 2008, 47, 5078–5087. [Google Scholar] [CrossRef] [PubMed]
- Bouter, A.; Gounou, C.; Bérat, R.; Tan, S.; Gallois, B.; Granier, T.; d’Estaintot, B.L.; Pöschl, E.; Brachvogel, B.; Brisson, A.R. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nat. Commun. 2011, 2, 270. [Google Scholar] [CrossRef]
- McNeil, A.K.; Rescher, U.; Gerke, V.; McNeil, P.L. Requirement for Annexin A1 in Plasma Membrane Repair. J. Biol. Chem. 2006, 281, 35202–35207. [Google Scholar] [CrossRef]
- Lichocka, M.; Krzymowska, M.; Górecka, M.; Hennig, J. Arabidopsis annexin 5 is involved in maintenance of pollen membrane integrity and permeability. J. Exp. Bot. 2022, 73, 94–109. [Google Scholar] [CrossRef]
- Maryam, M.; Fu, M.S.; Alanio, A.; Camacho, E.; Goncalves, D.S.; Faneuff, E.E.; Grossman, N.T.; Casadevall, A.; Coelho, C. The enigmatic role of fungal annexins: The case of Cryptococcus neoformans. Microbiology 2019, 165, 852–862. [Google Scholar] [CrossRef]
- Pervin, M.S.; Itoh, G.; Talukder, M.U.; Fujimoto, K.; Morimoto, Y.V.; Tanaka, M.; Ueda, M.; Yumura, S. A study of wound repair in Dictyostelium cells by using novel laserporation. Sci. Rep. 2018, 8, 7969. [Google Scholar] [CrossRef]
- Laohavisit, A.; Mortimer, J.C.; Demidchik, V.; Coxon, K.M.; Stancombe, M.A.; Macpherson, N.; Brownlee, C.; Hofmann, A.; Webb, A.A.R.; Miedema, H.; et al. Zea mays Annexins Modulate Cytosolic Free Ca2+ and Generate a Ca2+-Permeable Conductance. Plant Cell 2009, 21, 479–493. [Google Scholar] [CrossRef]
- Hofmann, A.; Proust, J.; Dorowski, A.; Schantz, R.; Huber, R. Annexin 24 from Capsicum annuum. J. Biol. Chem. 2000, 275, 8072–8082. [Google Scholar] [CrossRef]
- Kodavali, P.K.; Skowronek, K.; Koszela-Piotrowska, I.; Strzelecka-Kiliszek, A.; Pawlowski, K.; Pikula, S. Structural and functional characterization of annexin 1 from Medicago truncatula. Plant Physiol. Biochem. 2013, 73, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Gorecka, K.M.; Thouverey, C.; Buchet, R.; Pikula, S. Potential Role of Annexin AnnAt1 from Arabidopsis thaliana in pH-Mediated Cellular Response to Environmental Stimuli. Plant Cell Physiol. 2007, 48, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Reviakine, I. When a transmembrane channel isn’t, or how biophysics and biochemistry (mis)communicate. Biochim. Biophys. Acta—Biomembr. 2018, 1860, 1099–1104. [Google Scholar] [CrossRef]
- Lizarbe, M.; Barrasa, J.; Olmo, N.; Gavilanes, F.; Turnay, J. Annexin-Phospholipid Interactions. Functional Implications. Int. J. Mol. Sci. 2013, 14, 2652–2683. [Google Scholar] [CrossRef]
- Demange, P.; Voges, D.; Benz, J.; Liemann, S.; Göttig, P.; Berendes, R.; Burger, A.; Huber, R. Annexin V: The key to understanding ion selectivity and voltage regulation? Trends Biochem. Sci. 1994, 19, 272–276. [Google Scholar] [CrossRef]
- Eden, E.R.; Sanchez-Heras, E.; Tsapara, A.; Sobota, A.; Levine, T.P.; Futter, C.E. Annexin A1 Tethers Membrane Contact Sites that Mediate ER to Endosome Cholesterol Transport. Dev. Cell 2016, 37, 473–483. [Google Scholar] [CrossRef]
- Naidu, D.G.; Raha, A.; Chen, X.-L.; Spitzer, A.R.; Chander, A. Partial truncation of the NH2-terminus affects physical characteristics and membrane binding, aggregation, and fusion properties of annexin A7. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2005, 1734, 152–168. [Google Scholar] [CrossRef]
- Harder, T.; Gerke, V. The subcellular distribution of early endosomes is affected by the annexin II2p11(2) complex. J. Cell Biol. 1993, 123, 1119–1132. [Google Scholar] [CrossRef]
- Garbuglia, M.; Verzini, M.; Hofmann, A.; Huber, R.; Donato, R. S100A1 and S100B interactions with annexins. Biochim. Biophys. Acta—Mol. Cell Res. 2000, 1498, 192–206. [Google Scholar] [CrossRef]
- Dempsey, A.C.; Walsh, M.P.; Shaw, G.S. Unmasking the Annexin I Interaction from the Structure of Apo-S100A11. Structure 2003, 11, 887–897. [Google Scholar] [CrossRef]
- Chang, N.; Sutherland, C.; Hesse, E.; Winkfein, R.; Wiehler, W.B.; Pho, M.; Veillette, C.; Li, S.; Wilson, D.P.; Kiss, E.; et al. Identification of a novel interaction between the Ca2+ -binding protein S100A11 and the Ca2+- and phospholipid-binding protein annexin A6. Am. J. Physiol.-Cell Physiol. 2007, 292, C1417–C1430. [Google Scholar] [CrossRef] [PubMed]
- Illien, F.; Finet, S.; Lambert, O.; Ayala-Sanmartin, J. Different molecular arrangements of the tetrameric annexin 2 modulate the size and dynamics of membrane aggregation. Biochim. Biophys. Acta—Biomembr. 2010, 1798, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Ecsédi, P.; Kiss, B.; Gógl, G.; Radnai, L.; Buday, L.; Koprivanacz, K.; Liliom, K.; Leveles, I.; Vértessy, B.; Jeszenői, N.; et al. Regulation of the Equilibrium between Closed and Open Conformations of Annexin A2 by N-Terminal Phosphorylation and S100A4-Binding. Structure 2017, 25, 1195–1207.e5. [Google Scholar] [CrossRef]
- McCulloch, K.M.; Yamakawa, I.; Shifrin, D.A.; McConnell, R.E.; Foegeding, N.J.; Singh, P.K.; Mao, S.; Tyska, M.J.; Iverson, T.M. An alternative N-terminal fold of the intestine-specific annexin A13a induces dimerization and regulates membrane-binding. J. Biol. Chem. 2019, 294, 3454–3463. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Chipot, C.; Scheuring, S. Annexin-V stabilizes membrane defects by inducing lipid phase transition. Nat. Commun. 2020, 11, 230. [Google Scholar] [CrossRef]
- Eden, E.R.; White, I.J.; Tsapara, A.; Futter, C.E. Membrane contacts between endosomes and ER provide sites for PTP1B–epidermal growth factor receptor interaction. Nat. Cell Biol. 2010, 12, 267–272. [Google Scholar] [CrossRef]
- Liao, Y.-C.; Fernandopulle, M.S.; Wang, G.; Choi, H.; Hao, L.; Drerup, C.M.; Patel, R.; Qamar, S.; Nixon-Abell, J.; Shen, Y.; et al. RNA Granules Hitchhike on Lysosomes for Long-Distance Transport, Using Annexin A11 as a Molecular Tether. Cell 2019, 179, 147–164.e20. [Google Scholar] [CrossRef]
- Zhu, J.; Yuan, S.; Wei, G.; Qian, D.; Wu, X.; Jia, H.; Gui, M.; Liu, W.; An, L.; Xiang, Y. Annexin5 Is Essential for Pollen Development in Arabidopsis. Mol. Plant 2014, 7, 751–754. [Google Scholar] [CrossRef]
- Shirakawa, T.; Nakamura, A.; Kohama, K.; Hirakata, M.; Ogihara, S. Class-Specific Binding of Two Aminoacyl-tRNA Synthetases to Annexin, a Ca2+- and Phospholipid-Binding Protein. Cell Struct. Funct. 2005, 29, 159–164. [Google Scholar] [CrossRef]
- Arur, S.; Uche, U.E.; Rezaul, K.; Fong, M.; Scranton, V.; Cowan, A.E.; Mohler, W.; Han, D.K. Annexin I Is an Endogenous Ligand that Mediates Apoptotic Cell Engulfment. Dev. Cell 2003, 4, 587–598. [Google Scholar] [CrossRef]
- Hayes, M.J.; Shao, D.; Bailly, M.; Moss, S.E. Regulation of actin dynamics by annexin 2. EMBO J. 2006, 25, 1816–1826. [Google Scholar] [CrossRef] [PubMed]
- Andrawis, A.; Solomon, M.; Delmer, D.P. Cotton fiber annexins: A potential role in the regulation of callose synthase. Plant J. 1993, 3, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Kuchler, K.; Thorner, J. Secretion of Peptides and Proteins Lacking Hydrophobic Signal Sequences: The Role of Adenosine Triphosphate-Driven Membrane Translocators*. Endocr. Rev. 1992, 13, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef]
- Morgan, J.L.W.; Acheson, J.F.; Zimmer, J. Structure of a Type-1 Secretion System ABC Transporter. Structure 2017, 25, 522–529. [Google Scholar] [CrossRef]
- Akhtar, A.A.; Turner, D.P.J. The role of bacterial ATP-binding cassette (ABC) transporters in pathogenesis and virulence: Therapeutic and vaccine potential. Microb. Pathog. 2022, 171, 105734. [Google Scholar] [CrossRef]
- Chapman, L.P.; Epton, M.J.; Buckingham, J.C.; Morris, J.F.; Christian, H.C. Evidence for a Role of the Adenosine 5′-Triphosphate-Binding Cassette Transporter A1 in the Externalization of Annexin I from Pituitary Folliculo-Stellate Cells. Endocrinology 2003, 144, 1062–1073. [Google Scholar] [CrossRef]
- Wein, S.; Fauroux, M.; Laffitte, J.; de Nadaï, P.; Guaïni, C.; Pons, F.; Coméra, C. Mediation of annexin 1 secretion by a probenecid-sensitive ABC-transporter in rat inflamed mucosa. Biochem. Pharmacol. 2004, 67, 1195–1202. [Google Scholar] [CrossRef]
- Alam, A.; Locher, K.P. Structure and Mechanism of Human ABC Transporters. Annu. Rev. Biophys. 2023, 52, 275–300. [Google Scholar] [CrossRef]
- Thomas, C.; Tampé, R. Structural and Mechanistic Principles of ABC Transporters. Annu. Rev. Biochem. 2020, 89, 605–636. [Google Scholar] [CrossRef]
- Fernández, M.B.; Pagano, M.R.; Daleo, G.R.; Guevara, M.G. Hydrophobic proteins secreted into the apoplast may contribute to resistance against Phytophthora infestans in potato. Plant Physiol. Biochem. 2012, 60, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Engelbrecht, S.; Bakker-Grunwald, T.; Scholze, H. Functional identification of α-giardin as an annexin of Giardia lamblia. FEMS Microbiol. Lett. 1999, 173, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Davids, B.J.; Palm, J.E.D.; Housley, M.P.; Smith, J.R.; Andersen, Y.S.; Martin, M.G.; Hendrickson, B.A.; Johansen, F.-E.; Svärd, S.G.; Gillin, F.D.; et al. Polymeric Immunoglobulin Receptor in Intestinal Immune Defense against the Lumen-Dwelling Protozoan Parasite Giardia. J. Immunol. 2006, 177, 6281–6290. [Google Scholar] [CrossRef] [PubMed]
- Dubourg, A.; Xia, D.; Winpenny, J.P.; Al Naimi, S.; Bouzid, M.; Sexton, D.W.; Wastling, J.M.; Hunter, P.R.; Tyler, K.M. Giardia secretome highlights secreted tenascins as a key component of pathogenesis. Gigascience 2018, 7, giy003. [Google Scholar] [CrossRef]
- Ma’ayeh, S.Y.; Liu, J.; Peirasmaki, D.; Hörnaeus, K.; Bergström Lind, S.; Grabherr, M.; Bergquist, J.; Svärd, S.G. Characterization of the Giardia intestinalis secretome during interaction with human intestinal epithelial cells: The impact on host cells. PLoS Negl. Trop. Dis. 2017, 11, e0006120. [Google Scholar] [CrossRef]
- Moyano, S.; Musso, J.; Feliziani, C.; Zamponi, N.; Frontera, L.S.; Ropolo, A.S.; Lanfredi-Rangel, A.; Lalle, M.; Touz, M.C. Exosome Biogenesis in the Protozoa Parasite Giardia lamblia: A Model of Reduced Interorganellar Crosstalk. Cells 2019, 8, 1600. [Google Scholar] [CrossRef]
- Midlej, V.; de Souza, W.; Benchimol, M. The peripheral vesicles gather multivesicular bodies with different behavior during the Giardia intestinalis life cycle. J. Struct. Biol. 2019, 207, 301–311. [Google Scholar] [CrossRef]
- Chander, A.; Naidu, D.G.; Chen, X.-L. A ten-residue domain (Y11–A20) in the NH2-terminus modulates membrane association of annexin A7. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2006, 1761, 775–784. [Google Scholar] [CrossRef]
- Rintala-Dempsey, A.C.; Rezvanpour, A.; Shaw, G.S. S100–annexin complexes—Structural insights. FEBS J. 2008, 275, 4956–4966. [Google Scholar] [CrossRef]
- Patel, D.R.; Isas, J.M.; Ladokhin, A.S.; Jao, C.C.; Kim, Y.E.; Kirsch, T.; Langen, R.; Haigler, H.T. The conserved core domains of annexins A1, A2, A5, and B12 can be divided into two groups with different Ca2+-dependent membrane-binding properties. Biochemistry 2005, 44, 2833–2844. [Google Scholar] [CrossRef]
- Chen, J.M.; Sheldon, A.; Pincus, M.R. Structure-Function Correlations of Calcium Binding and Calcium Channel Activities Based on 3-Dimensional Models of Human Annexins I, II, III, V and VII. J. Biomol. Struct. Dyn. 1993, 10, 1067–1089. [Google Scholar] [CrossRef] [PubMed]
- Trotter, P.J.; Orchard, M.A.; Walker, J.H. Ca2+ concentration during binding determines the manner in which annexin V binds to membranes. Biochem. J. 1995, 308, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Raynal, P.; Pollard, H.B. Annexins: The problem of assessing the biological role for a gene family of multifunctional calcium- and phospholipid-binding proteins. Biochim. Biophys. Acta—Rev. Biomembr. 1994, 1197, 63–93. [Google Scholar] [CrossRef] [PubMed]
- Turnay, J.; Lecona, E.; Fernández-Lizarbe, S.; Guzmán-Aránguez, A.; Fernández, M.P.; Olmo, N.; Lizarbe, M.A. Structure–function relationship in annexin A13, the founder member of the vertebrate family of annexins. Biochem. J. 2005, 389, 899–911. [Google Scholar] [CrossRef]
- Sopkova, J.; Vincent, M.; Takahashi, M.; Lewit-Bentley, A.; Gallay, J. Conformational Flexibility of Domain III of Annexin V Studied by Fluorescence of Tryptophan 187 and Circular Dichroism: The Effect of PH. Biochemistry 1998, 37, 11962–11970. [Google Scholar] [CrossRef]
- Zschörnig, O.; Opitz, F.; Müller, M. Annexin A4 binding to anionic phospholipid vesicles modulated by pH and calcium. Eur. Biophys. J. 2007, 36, 415–424. [Google Scholar] [CrossRef]
- Dabitz, N.; Hu, N.-J.; Yusof, A.M.; Tranter, N.; Winter, A.; Daley, M.; Zschörnig, O.; Brisson, A.; Hofmann, A. Structural Determinants for Plant Annexin−Membrane Interactions. Biochemistry 2005, 44, 16292–16300. [Google Scholar] [CrossRef]
- Kaetzel, M.A.; Mo, Y.D.; Mealy, T.R.; Campos, B.; Bergsma-Schutter, W.; Brisson, A.; Dedman, J.R.; Seaton, B.A. Phosphorylation Mutants Elucidate the Mechanism of Annexin IV-Mediated Membrane Aggregation. Biochemistry 2001, 40, 4192–4199. [Google Scholar] [CrossRef]
- Matos, A.L.L.; Kudruk, S.; Moratz, J.; Heflik, M.; Grill, D.; Ravoo, B.J.; Gerke, V. Membrane Binding Promotes Annexin A2 Oligomerization. Cells 2020, 9, 1169. [Google Scholar] [CrossRef]
- Grill, D.; Matos, A.L.L.; de Vries, W.C.; Kudruk, S.; Heflik, M.; Dörner, W.; Mootz, H.D.; Ravoo, B.J.; Galla, H.J.; Gerke, V. Bridging of membrane surfaces by annexin A2. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warmus, D.; Balmer, E.A.; Faso, C. Annexin–Membrane Interactions Across Eukaryotic Domains of Life—A Comparative Approach. Int. J. Mol. Sci. 2025, 26, 6517. https://doi.org/10.3390/ijms26136517
Warmus D, Balmer EA, Faso C. Annexin–Membrane Interactions Across Eukaryotic Domains of Life—A Comparative Approach. International Journal of Molecular Sciences. 2025; 26(13):6517. https://doi.org/10.3390/ijms26136517
Chicago/Turabian StyleWarmus, Dawid, Erina Alexandra Balmer, and Carmen Faso. 2025. "Annexin–Membrane Interactions Across Eukaryotic Domains of Life—A Comparative Approach" International Journal of Molecular Sciences 26, no. 13: 6517. https://doi.org/10.3390/ijms26136517
APA StyleWarmus, D., Balmer, E. A., & Faso, C. (2025). Annexin–Membrane Interactions Across Eukaryotic Domains of Life—A Comparative Approach. International Journal of Molecular Sciences, 26(13), 6517. https://doi.org/10.3390/ijms26136517