
Pulsatile Myofilament Activity in Myotrem Myopathy Associated with Myogenic Tremor

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Contractile Properties of Soleus Muscle Fibers
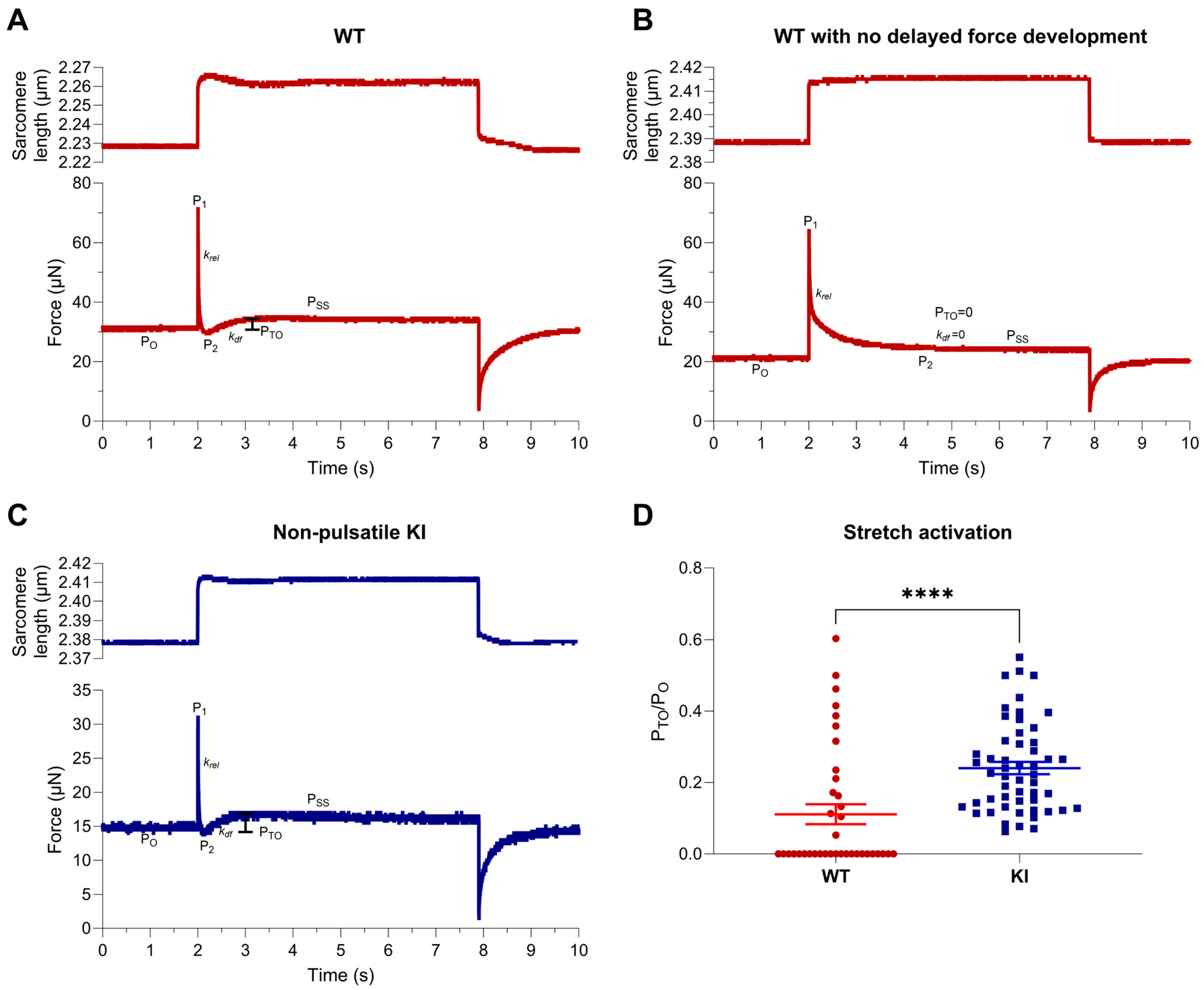
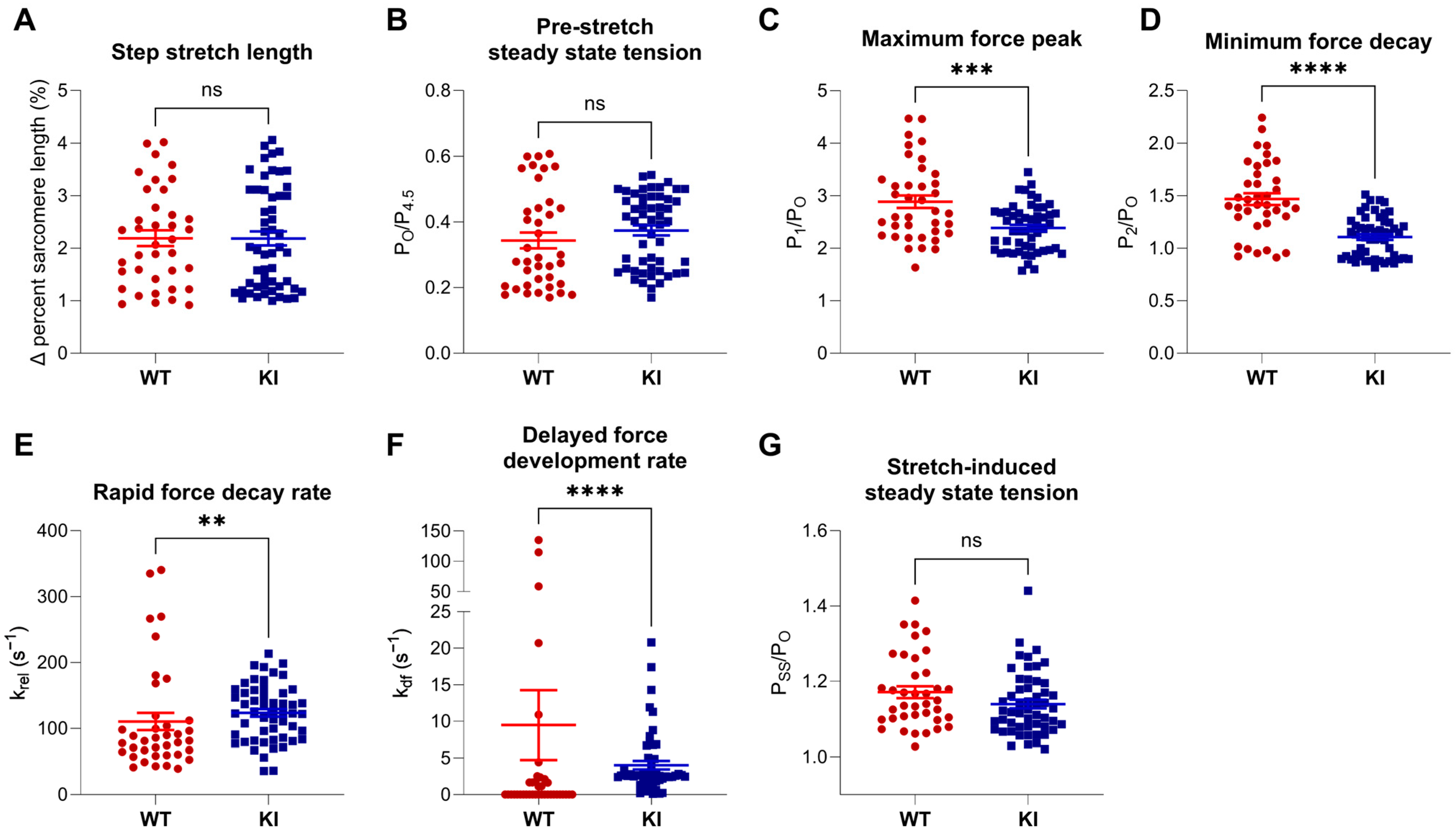
2.2. Stretch Activation Properties of Soleus Muscle Fibers
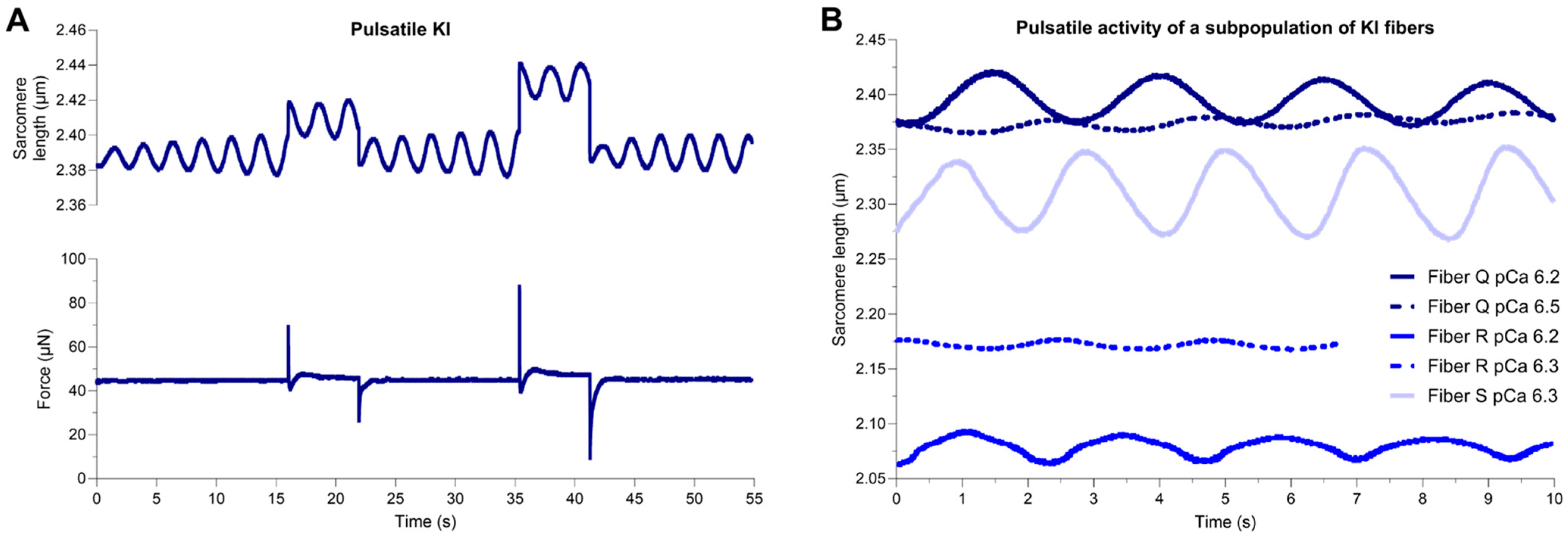
2.3. Pulsatile Activity of a Subpopulation of E248K KI Soleus Muscle Fibers
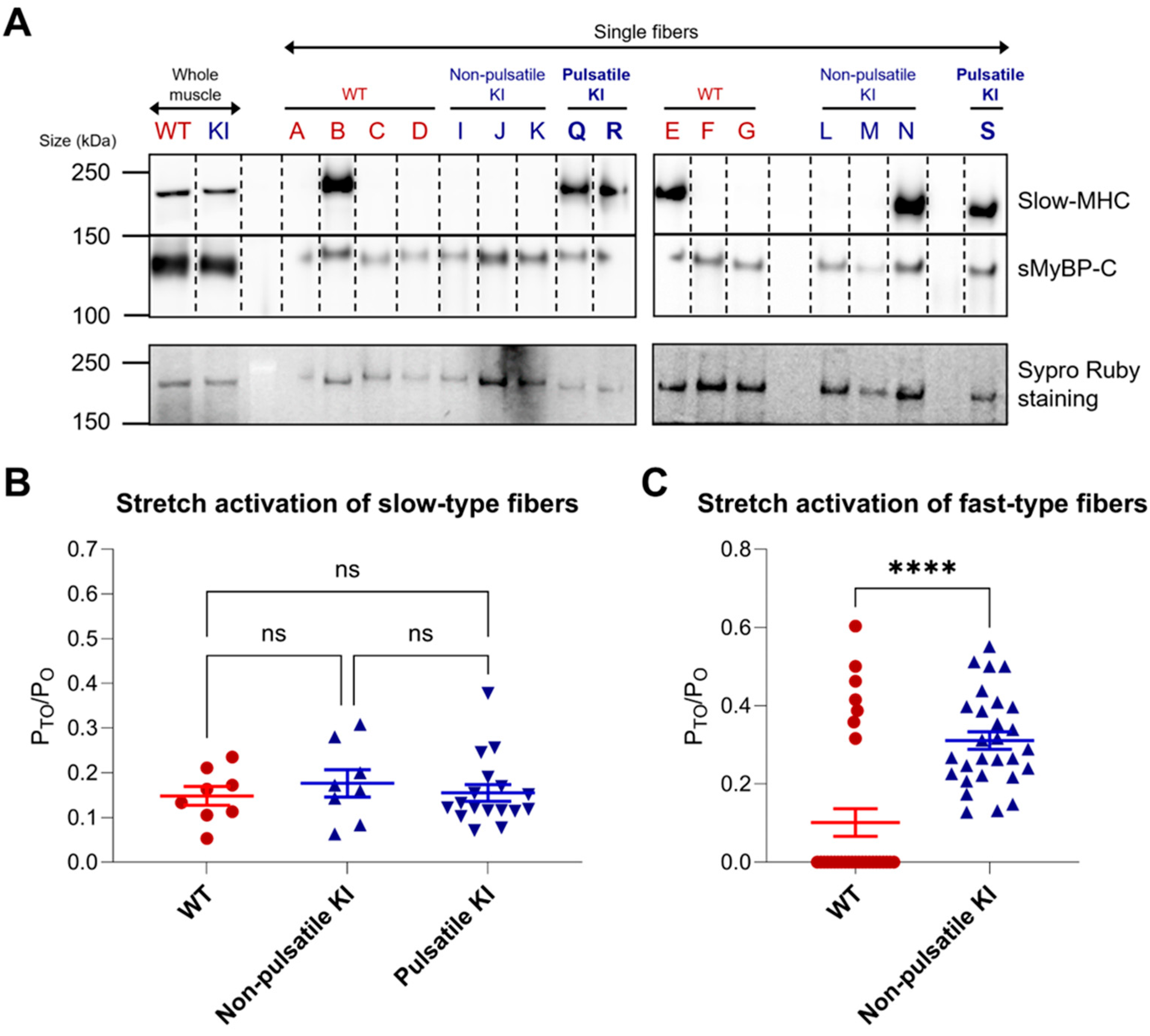
2.4. Fiber-Type Specific Stretch Activation Properties
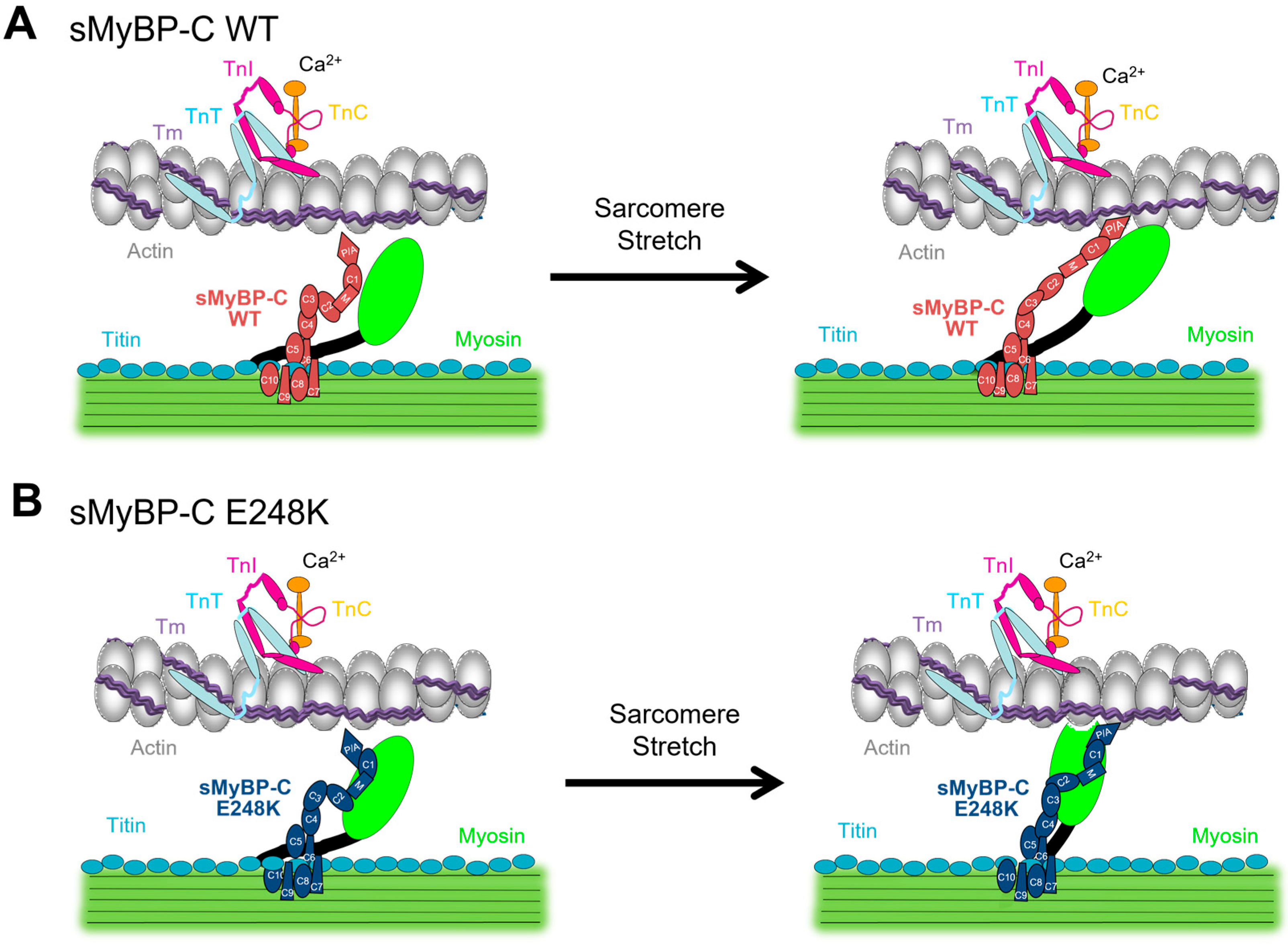
3. Discussion
4. Materials and Methods
4.1. Study Approval
4.2. Solutions
4.3. Fiber Preparation and Experimental Setup
4.4. Slack–Re-Stretch Protocol
4.5. Step–Stretch Protocol and Data Analysis
4.6. Single Fiber Immunoblot Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| EDL | Extensor digitorum longus |
| kdf | Delayed force development rate |
| KI | Knock-In |
| krel | Rapid force decay rate |
| MHC | Myosin Heavy Chain |
| MyBP-C | Myosin binding protein-C |
| PO | Pre-stretch steady state tension |
| PSS | Stretch-induced steady state tension |
| PTO | Stretch activation |
| P1 | Maximum force peak |
| P2 | Minimum force decay |
| sMyBP-C | Slow skeletal myosin binding protein-C |
| SPOC | Spontaneous oscillatory contraction |
| WT | Wild type |
References
- Stavusis, J.; Lace, B.; Schafer, J.; Geist, J.; Inashkina, I.; Kidere, D.; Pajusalu, S.; Wright, N.T.; Saak, A.; Weinhold, M.; et al. Novel Mutations in MYBPC1 Are Associated with Myogenic Tremor and Mild Myopathy. Ann. Neurol. 2019, 86, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.A.; Kontrogianni-Konstantopoulos, A. Myosin Binding Protein-C Slow: A Multifaceted Family of Proteins with a Complex Expression Profile in Fast and Slow Twitch Skeletal Muscles. Front. Physiol. 2013, 4, 391. [Google Scholar] [CrossRef] [PubMed]
- Craig, R.; Offer, G. The Location of C-Protein in Rabbit Skeletal Muscle. Proc. R. Soc. Lond. B Biol. Sci. 1976, 192, 451–461. [Google Scholar] [CrossRef]
- Okagaki, T.; Weber, F.E.; Fischman, D.A.; Vaughan, K.T.; Mikawa, T.; Reinach, F.C. The Major Myosin-Binding Domain of Skeletal Muscle MyBP-C (C Protein) Resides in the COOH-Terminal, Immunoglobulin C2 Motif. J. Cell Biol. 1993, 123, 619–626. [Google Scholar] [CrossRef]
- Gruen, M.; Prinz, H.; Gautel, M. cAPK-Phosphorylation Controls the Interaction of the Regulatory Domain of Cardiac Myosin Binding Protein C with Myosin-S2 in an on-off Fashion. FEBS Lett. 1999, 453, 254–259. [Google Scholar] [CrossRef]
- Moos, C. Fluorescence Microscope Study of the Binding of Added C Protein to Skeletal Muscle Myofibrils. J. Cell Biol. 1981, 90, 25–31. [Google Scholar] [CrossRef]
- Kulikovskaya, I.; McClellan, G.; Flavigny, J.; Carrier, L.; Winegrad, S. Effect of MyBP-C Binding to Actin on Contractility in Heart Muscle. J. Gen. Physiol. 2003, 122, 761–774. [Google Scholar] [CrossRef]
- Geist, J.; Ward, C.W.; Kontrogianni-Konstantopoulos, A. Structure before Function: Myosin Binding Protein-C Slow Is a Structural Protein with Regulatory Properties. FASEB J. 2018, 32, fj201800624R. [Google Scholar] [CrossRef]
- Campbell, K.B.; Chandra, M. Functions of Stretch Activation in Heart Muscle. J. Gen. Physiol. 2006, 127, 89–94. [Google Scholar] [CrossRef]
- Linari, M.; Reedy, M.K.; Reedy, M.C.; Lombardi, V.; Piazzesi, G. Ca-Activation and Stretch-Activation in Insect Flight Muscle. Biophys. J. 2004, 87, 1101–1111. [Google Scholar] [CrossRef]
- Josephson, R.K.; Malamud, J.G.; Stokes, D.R. Asynchronous Muscle: A Primer. J. Exp. Biol. 2000, 203, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.P. Making waves: A Proposed New Role for Myosin-Binding Protein C in Regulating Oscillatory Contractions in Vertebrate Striated Muscle. J. Gen. Physiol. 2021, 153, e202012729. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, J.E.; Dunning, S.B.; Moss, R.L. Ablation of Cardiac Myosin-Binding Protein-C Accelerates Stretch Activation in Murine Skinned Myocardium. Circ. Res. 2006, 98, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Straight, C.R.; Bell, K.M.; Slosberg, J.N.; Miller, M.S.; Swank, D.M. A Myosin-Based Mechanism for Stretch Activation and Its Possible Role Revealed by Varying Phosphate Concentration in Fast and Slow Mouse Skeletal Muscle Fibers. Am. J. Physiol. Cell Physiol. 2019, 317, C1143–C1152. [Google Scholar] [CrossRef]
- Robinett, J.C.; Hanft, L.M.; Biesiadecki, B.; McDonald, K.S. Molecular Regulation of Stretch Activation. Am. J. Physiol. Cell Physiol. 2022, 323, C1728–C1739. [Google Scholar] [CrossRef]
- Stelzer, J.E.; Moss, R.L. Contributions of Stretch Activation to Length-Dependent Contraction in Murine Myocardium. J. Gen. Physiol. 2006, 128, 461–471. [Google Scholar] [CrossRef]
- Geist Hauserman, J.; Stavusis, J.; Joca, H.C.; Robinett, J.C.; Hanft, L.; Vandermeulen, J.; Zhao, R.; Stains, J.P.; Konstantopoulos, K.; McDonald, K.S.; et al. Sarcomeric Deficits Underlie MYBPC1-Associated Myopathy with Myogenic Tremor. JCI Insight 2021, 6, e147612. [Google Scholar] [CrossRef]
- Vinciguerra, C.; Iacono, S.; Bevilacqua, L.; Landolfi, A.; Piscosquito, G.; Ginanneschi, F.; Schiro, G.; Di Stefano, V.; Brighina, F.; Barone, P.; et al. Sex Differences in Neuromuscular Disorders. Mech. Ageing Dev. 2023, 211, 111793. [Google Scholar] [CrossRef]
- Yasuda, K.; Shindo, Y.; Ishiwata, S. Synchronous Behavior of Spontaneous Oscillations of Sarcomeres in Skeletal Myofibrils under Isotonic Conditions. Biophys. J. 1996, 70, 1823–1829. [Google Scholar] [CrossRef]
- Schaefer, J.; Saak, A.; Bonnemann, C.G.; Jackson, S. Myogenic Tremor—A Novel Tremor Entity. Curr. Opin. Neurol. 2021, 34, 706–713. [Google Scholar] [CrossRef]
- Semmler, J.G.; Kornatz, K.W.; Dinenno, D.V.; Zhou, S.; Enoka, R.M. Motor Unit Synchronisation Is Enhanced during Slow Lengthening Contractions of a Hand Muscle. J. Physiol. 2002, 545, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Napierski, N.C.; Granger, K.; Langlais, P.R.; Moran, H.R.; Strom, J.; Touma, K.; Harris, S.P. A Novel “Cut and Paste” Method for in situ Replacement of cMyBP-C Reveals a New Role for cMyBP-C in the Regulation of Contractile Oscillations. Circ. Res. 2020, 126, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, J.E.; Patel, J.R.; Moss, R.L. Protein Kinase A-Mediated Acceleration of the Stretch Activation Response in Murine Skinned Myocardium Is Eliminated by Ablation of cMyBP-C. Circ. Res. 2006, 99, 884–890. [Google Scholar] [CrossRef]
- Ackermann, M.A.; Kontrogianni-Konstantopoulos, A. Myosin Binding Protein-C Slow Is a Novel Substrate for Protein Kinase A (PKA) and C (PKC) in Skeletal Muscle. J. Proteome Res. 2011, 10, 4547–4555. [Google Scholar] [CrossRef]
- Proske, U.; Gandevia, S.C. The Proprioceptive Senses: Their Roles in Signaling Body Shape, Body Position and Movement, and Muscle Force. Physiol. Rev. 2012, 92, 1651–1697. [Google Scholar] [CrossRef]
- Kistemaker, D.A.; Van Soest, A.J.; Wong, J.D.; Kurtzer, I.; Gribble, P.L. Control of Position and Movement Is Simplified by Combined Muscle Spindle and Golgi Tendon Organ Feedback. J. Neurophysiol. 2013, 109, 1126–1139. [Google Scholar] [CrossRef]
- Koller, W.C.; Rueda, M.G. Mechanism of Action of Dopaminergic Agents in Parkinson’s Disease. Neurology 1998, 50, S11–S14, discussion S44–S48. [Google Scholar] [CrossRef]
- McDonald, K.S.; Fitts, R.H. Effect of Hindlimb Unweighting on Single Soleus Fiber Maximal Shortening Velocity and ATPase Activity. J. Appl. Physiol. 1993, 74, 2949–2957. [Google Scholar] [CrossRef]
- McDonald, K.S.; Fitts, R.H. Effect of Hindlimb Unloading on Rat Soleus Fiber Force, Stiffness, and Calcium Sensitivity. J. Appl. Physiol. 1995, 79, 1796–1802. [Google Scholar] [CrossRef]
- McDonald, K.S.; Wolff, M.R.; Moss, R.L. Sarcomere Length Dependence of the Rate of Tension Redevelopment and Submaximal Tension in Rat and Rabbit Skinned Skeletal Muscle Fibers. J. Physiol. 1997, 501, 607–621. [Google Scholar] [CrossRef]
- McDonald, K.S. Ca2+ Dependence of Loaded Shortening in Rat Skinned Cardiac Myocytes and Skeletal Muscle Fibers. J. Physiol. 2000, 525, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Hanft, L.M.; McDonald, K.S. Length Dependence of Force Generation Exhibit Similarities between Rat Cardiac Myocytes and Skeletal Muscle Fibres. J. Physiol. 2010, 588, 2891–2903. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B. An Indirect Proof of Stretch-Induced Ca++ Release from the Sarcoplasmic Reticulum in Glycerinated Skeletal and Heart Muscle Preparations (Author’s Transl). Basic. Res. Cardiol. 1979, 74, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Hinken, A.C.; McDonald, K.S. Inorganic Phosphate Speeds Loaded Shortening in Rat Skinned Cardiac Myocytes. Am. J. Physiol. Cell Physiol. 2004, 287, C500–C507. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fiber Length (µm) | Fiber Widthair (µm) | Sarcomere Length (µm) | Passive Tension (kN/m2) | Maximal Tension (kN/m2) | pCa 4.5 ktr (s−1) | ||
|---|---|---|---|---|---|---|---|
| WT | n′ = 8 | 911 ±71 | 39 ± 2 | 2.36 ± 0.03 | 3.29 ± 0.49 | 93 ± 5 | 10.5 ± 1.1 |
| KI | n′ = 11 | 939 ± 60 | 44 ± 1 | 2.32 ± 0.03 | 2.56 ± 0.32 | 64 ± 6 * | 10.3 ± 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariano, J.M.; Hanft, L.M.; Cho, S.; Ward, C.W.; McDonald, K.S.; Kontrogianni-Konstantopoulos, A. Pulsatile Myofilament Activity in Myotrem Myopathy Associated with Myogenic Tremor. Int. J. Mol. Sci. 2025, 26, 5252. https://doi.org/10.3390/ijms26115252
Mariano JM, Hanft LM, Cho S, Ward CW, McDonald KS, Kontrogianni-Konstantopoulos A. Pulsatile Myofilament Activity in Myotrem Myopathy Associated with Myogenic Tremor. International Journal of Molecular Sciences. 2025; 26(11):5252. https://doi.org/10.3390/ijms26115252
Chicago/Turabian StyleMariano, Jennifer Megan, Laurin M. Hanft, Suhan Cho, Christopher W. Ward, Kerry S. McDonald, and Aikaterini Kontrogianni-Konstantopoulos. 2025. "Pulsatile Myofilament Activity in Myotrem Myopathy Associated with Myogenic Tremor" International Journal of Molecular Sciences 26, no. 11: 5252. https://doi.org/10.3390/ijms26115252
APA StyleMariano, J. M., Hanft, L. M., Cho, S., Ward, C. W., McDonald, K. S., & Kontrogianni-Konstantopoulos, A. (2025). Pulsatile Myofilament Activity in Myotrem Myopathy Associated with Myogenic Tremor. International Journal of Molecular Sciences, 26(11), 5252. https://doi.org/10.3390/ijms26115252