
Circulating FGF21 and Ketone Bodies Modify the Risk of MASLD and Mortality: Insights from the PREVEND Cohort Study

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Baseline Characteristics of the Study Population
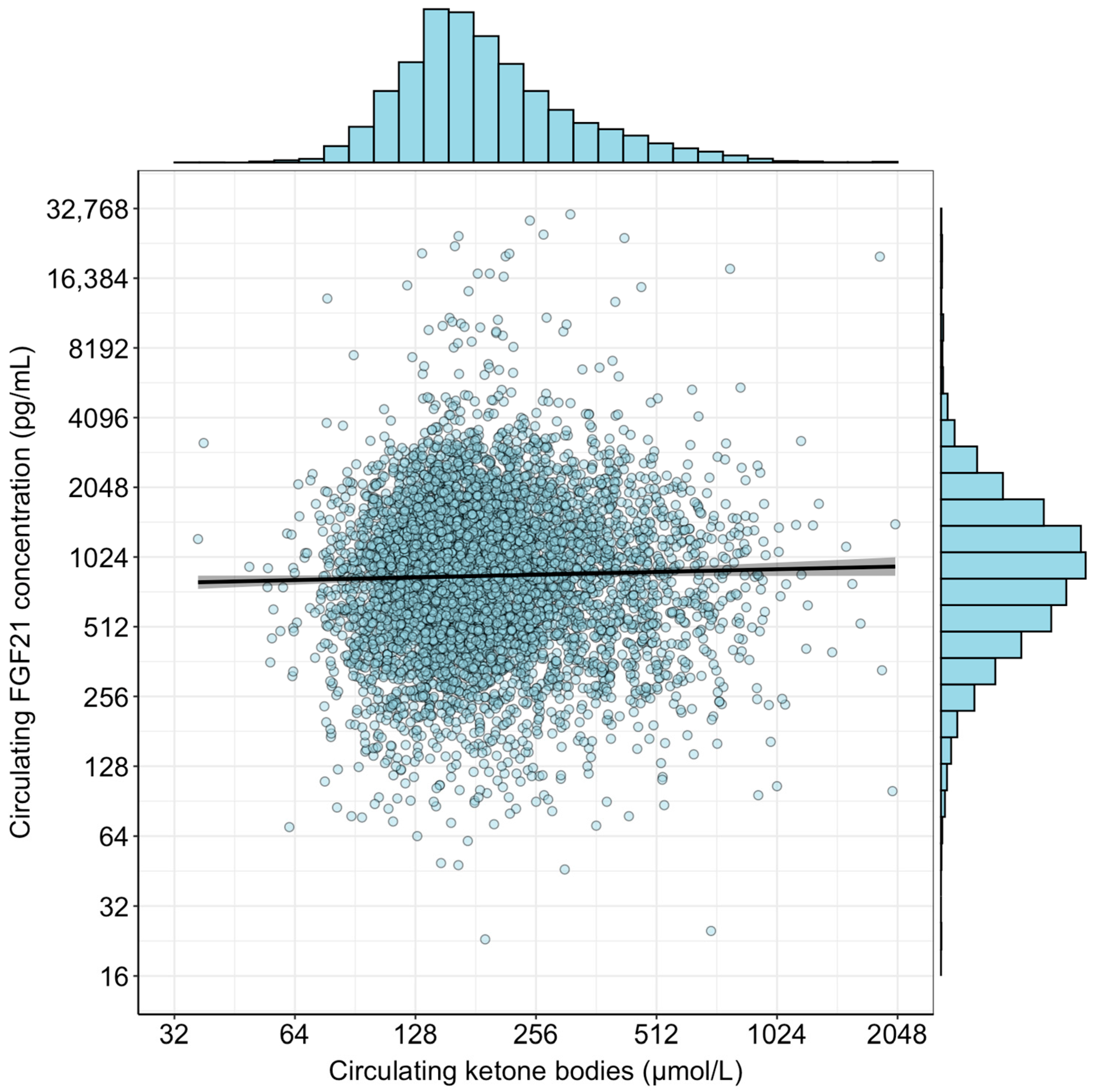
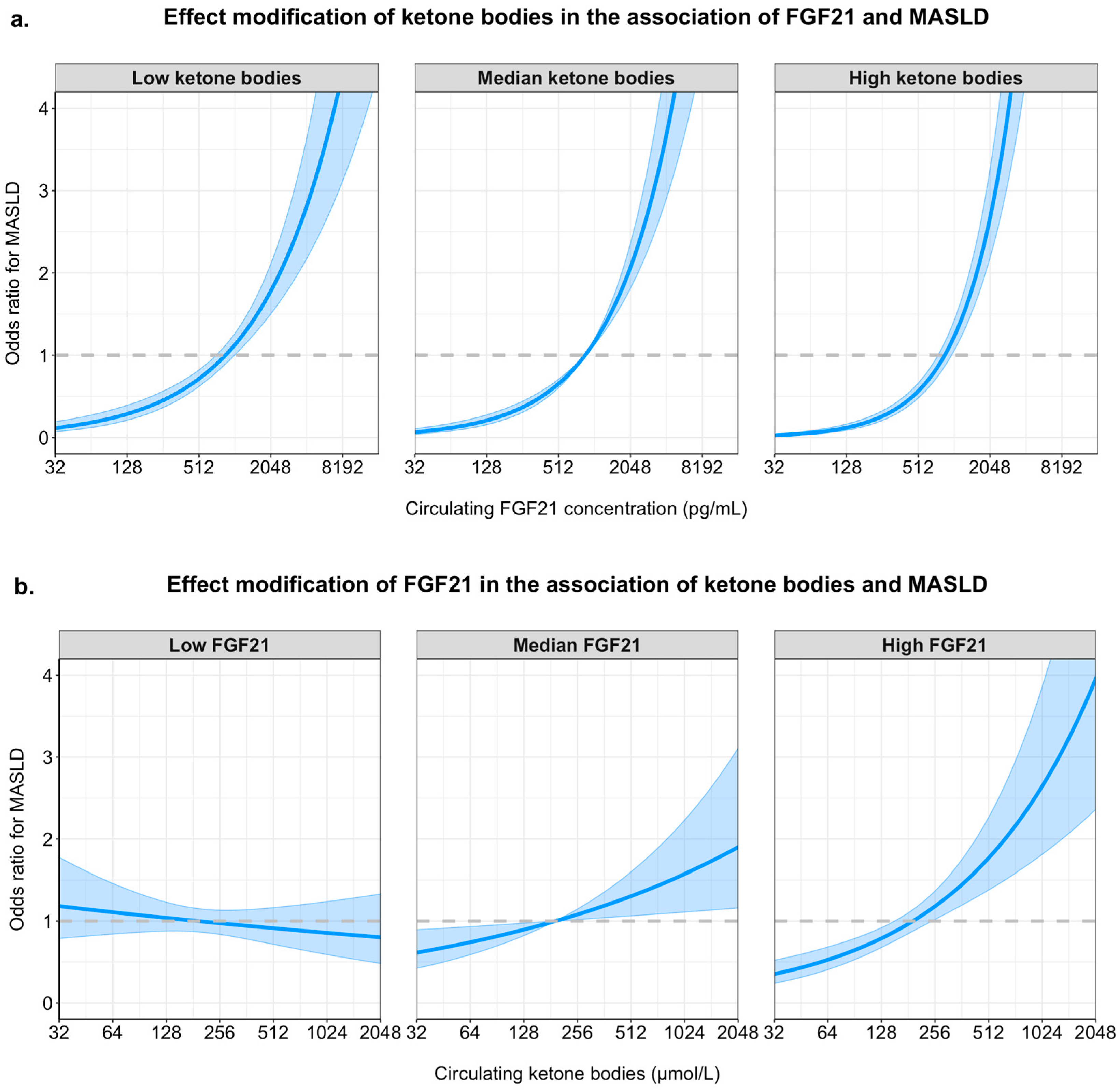
2.2. Logistic Regression Analyses and Interaction Between FGF21 and Ketone Bodies in MASLD
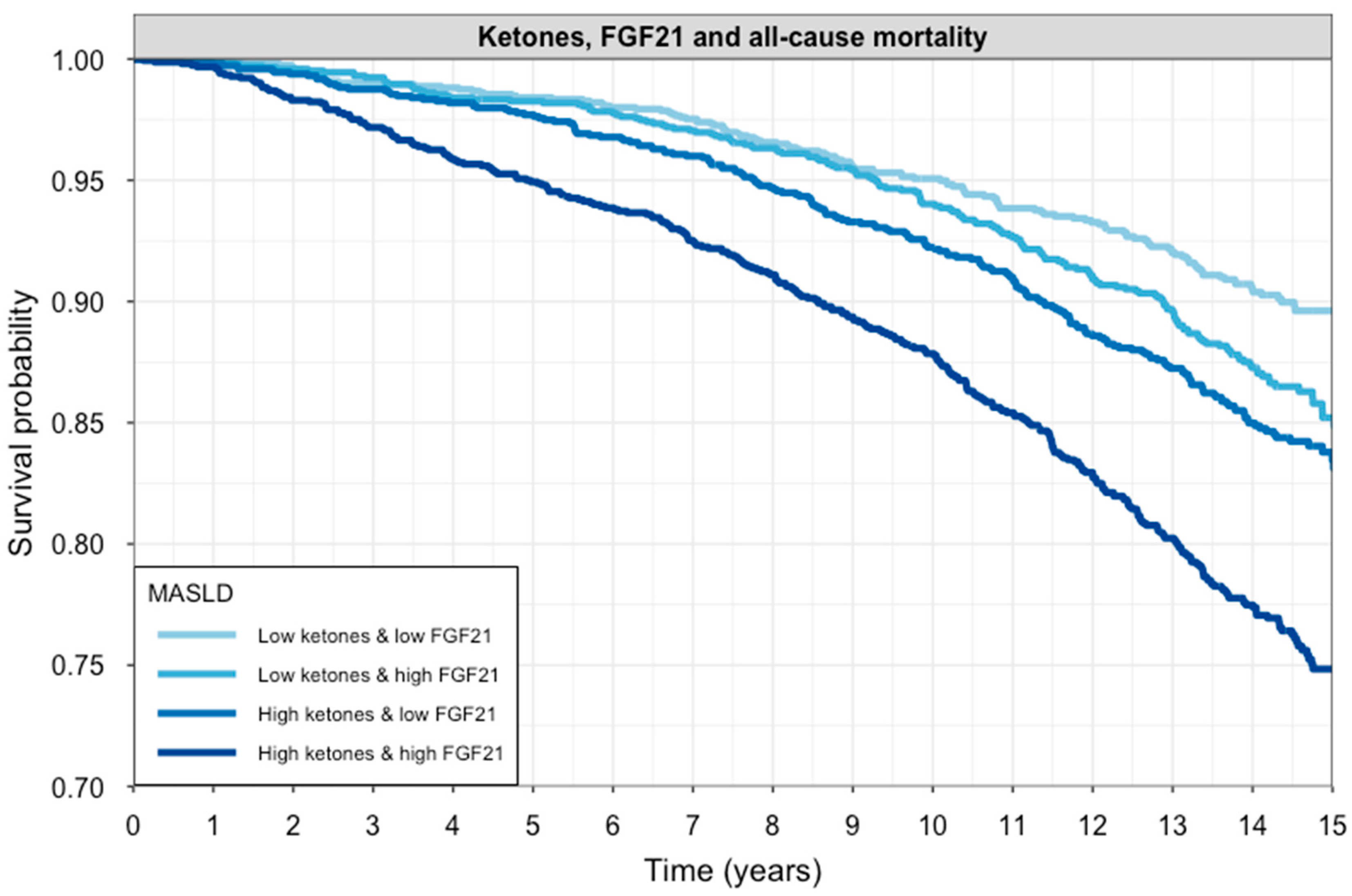
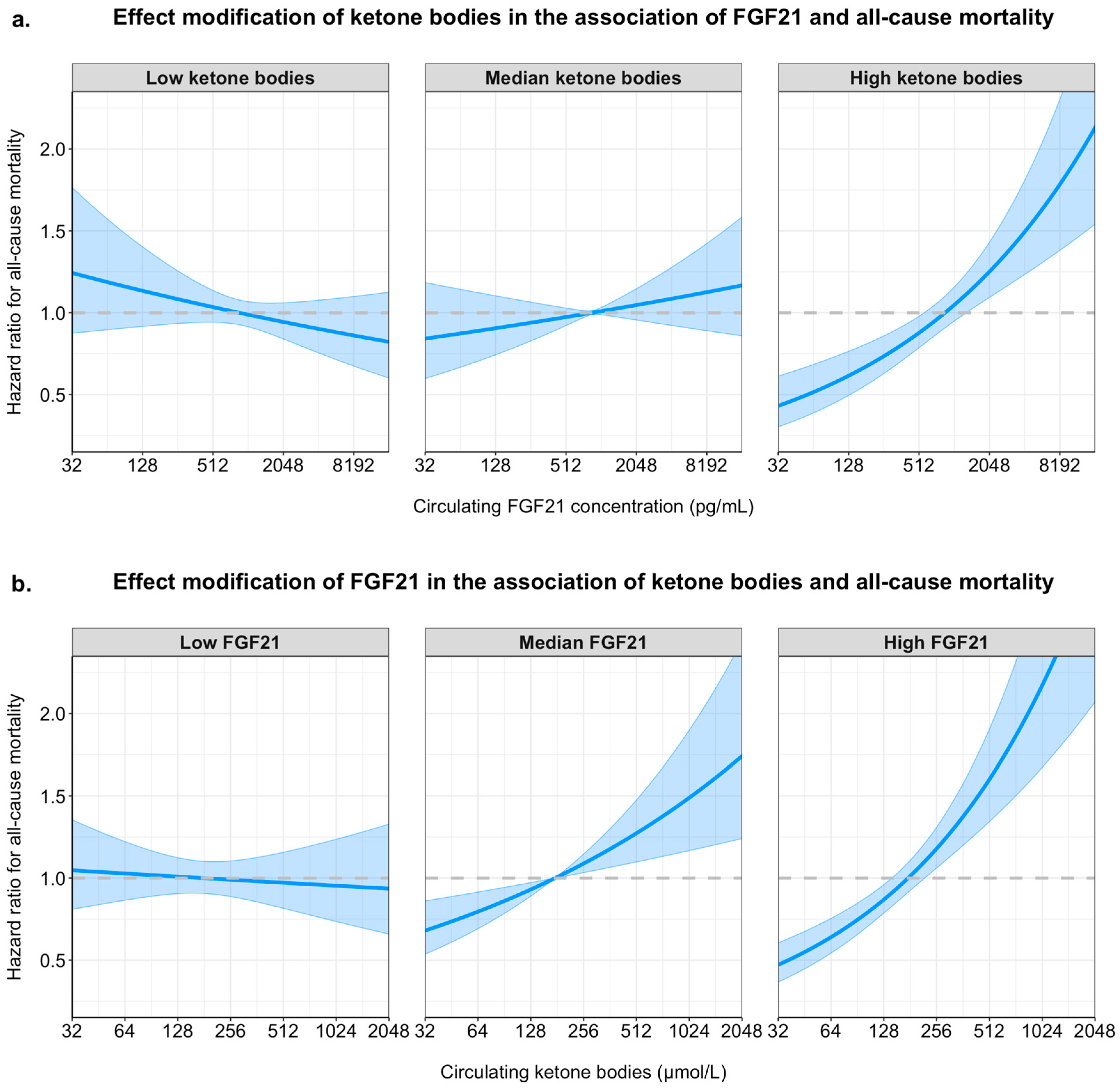
2.3. Longitudinal Analyses and Interaction of FGF21 and Ketone Bodies on All-Cause Mortality
3. Discussion
4. Materials and Methods
4.1. Study Design and Participants
4.2. Data Collection
4.3. Outcome Data
4.4. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AcAc | Acetoacetate |
| AMPK | AMP-activated protein kinase |
| BHB | β-Hydroxybutyrate |
| eGFR | Estimated glomerular filtration rate |
| FGF | Fibroblast growth factor |
| FGF21 | Fibroblast growth factor 21 |
| FLI | Fatty Liver Index |
| GFR | Glomerular filtration rate |
| GGT | Gamma-glutamyltransferase |
| HDL | High-density lipoprotein |
| ICD-10 | International Classification of Diseases, 10th Revision |
| IQ | Interquartile |
| MASLD | Metabolic dysfunction-associated steatotic liver disease |
| MASH | Metabolic dysfunction-associated steatohepatitis |
| NMR | Nuclear magnetic resonance |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| OR | Odds ratio |
| PPARα | Peroxisome proliferator-activated receptor alpha |
| PREVEND | Prevention of Renal and Vascular End-stage Disease |
| SD | Standard deviation |
| T2D | Type 2 diabetes |
References
- Post, A.; Groothof, D.; Schutten, J.C.; Kelly, D.; Swarte, J.C.; Flores-Guerrero, J.L.; van der Veen, Y.; Kema, I.P.; Ozyilmaz, A.; Enya, A.; et al. Fibroblast growth factor 21 and protein energy wasting in hemodialysis patients. Clin. Nutr. 2021, 40, 4216–4224. [Google Scholar] [CrossRef] [PubMed]
- Post, A.; Dam, W.A.; Sokooti, S.; Groothof, D.; Gloerich, J.; van Gool, A.J.; Kremer, D.; Gansevoort, R.T.; Born, J.v.D.; Kema, I.P.; et al. Circulating FGF21 Concentration, Fasting Plasma Glucose, and the Risk of Type 2 Diabetes: Results From the PREVEND Study. J. Clin. Endocrinol. Metab. 2022, 108, 1387–1393. [Google Scholar] [CrossRef]
- Domouzoglou, E.M.; Naka, K.K.; Vlahos, A.P.; Papafaklis, M.I.; Michalis, L.K.; Tsatsoulis, A.; Maratos-Flier, E. Fibroblast growth factors in cardiovascular disease: The emerging role of FGF21. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1029–H1038. [Google Scholar] [CrossRef] [PubMed]
- Szczepańska, E.; Gietka-Czernel, M. FGF21: A Novel Regulator of Glucose and Lipid Metabolism and Whole-Body Energy Balance. Horm. Metab. Res. 2022, 54, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.E.; Ebling, F.J.; Samms, R.J.; Tsintzas, K. Going Back to the Biology of FGF21: New Insights. Trends Endocrinol. Metab. 2019, 30, 491–504. [Google Scholar] [CrossRef]
- Fisher, F.M.; Maratos-Flier, E. Understanding the Physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef]
- Veneti, S.; Grammatikopoulou, M.G.; Kintiraki, E.; Mintziori, G.; Goulis, D.G. Ketone Bodies in Diabetes Mellitus: Friend or Foe? Nutrients 2023, 15, 4383. [Google Scholar] [CrossRef]
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes/Metabolism Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Szili-Torok, T.; de Borst, M.H.; Garcia, E.; Gansevoort, R.T.; Dullaart, R.P.; Connelly, M.A.; Bakker, S.J.; Tietge, U.J. Fasting Ketone Bodies and Incident Type 2 Diabetes in the General Population. Diabetes 2023, 72, 1187–1192. [Google Scholar] [CrossRef]
- Obokata, M.; Negishi, K.; Sunaga, H.; Ishida, H.; Ito, K.; Ogawa, T.; Iso, T.; Ando, Y.; Kurabayashi, M. Association Between Circulating Ketone Bodies and Worse Outcomes in Hemodialysis Patients. J. Am. Heart Assoc. 2017, 6, e006885. [Google Scholar] [CrossRef]
- Post, A.; Garcia, E.; van den Berg, E.H.; Flores-Guerrero, J.L.; Gruppen, E.G.; Groothof, D.; Westenbrink, B.D.; Connelly, M.A.; Bakker, S.J.; Dullaart, R.P. Nonalcoholic fatty liver disease, circulating ketone bodies and all-cause mortality in a general population-based cohort. Eur. J. Clin. Investig. 2021, 51, e13627. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Shalaurova, I.; Matyus, S.P.; Oskardmay, D.N.; Otvos, J.D.; Dullaart, R.P.; Connelly, M.A. Ketone Bodies Are Mildly Elevated in Subjects with Type 2 Diabetes Mellitus and Are Inversely Associated with Insulin Resistance as Measured by the Lipoprotein Insulin Resistance Index. J. Clin. Med. 2020, 9, 321. [Google Scholar] [CrossRef]
- Palm, C.L.; Shalaurova, I.; Connelly, M.A.; Bakker, S.J.L.; Westenbrink, B.D.; Dullaart, R.P.F. Fasting Plasma Ketone Bodies Are Associated with NT-proBNP: A Potential Mechanism to Provide Fuel for the Failing Heart. J. Clin. Med. 2024, 13, 1541. [Google Scholar] [CrossRef]
- de Koning, M.-S.L.; Westenbrink, B.D.; Assa, S.; Garcia, E.; Connelly, M.A.; van Veldhuisen, D.J.; Dullaart, R.P.; Lipsic, E.; van der Harst, P. Association of Circulating Ketone Bodies with Functional Outcomes After ST-Segment Elevation Myocardial Infarction. J. Am. Coll. Cardiol. 2021, 78, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guerrero, J.L.; Westenbrink, B.D.; Connelly, M.A.; Otvos, J.D.; Groothof, D.; Shalaurova, I.; Garcia, E.; Navis, G.; de Boer, R.A.; Bakker, S.J.L.; et al. Association of beta-hydroxybutyrate with development of heart failure: Sex differences in a Dutch population cohort. Eur. J. Clin. Investig. 2021, 51, e13468. [Google Scholar] [CrossRef] [PubMed]
- Tucker, B.; Li, H.; Long, X.; Rye, K.-A.; Ong, K.L. Fibroblast growth factor 21 in non-alcoholic fatty liver disease. Metabolism 2019, 101, 153994. [Google Scholar] [CrossRef]
- Tucker, W.J.; Tucker, B.; Januszewski, A.S.; Jenkins, A.J.; Keech, A.C.; Kestenbaum, B.R.; Allison, M.A.; Rye, K.-A.; Ong, K.L. Association of circulating fibroblast growth factor 21 levels with all-cause and cardiovascular mortality: The multi-ethnic study of atherosclerosis. Clin. Chim. Acta 2024, 555, 117799. [Google Scholar] [CrossRef]
- Yang, M.; Liu, C.; Jiang, N.; Liu, Y.; Luo, S.; Li, C.; Zhao, H.; Han, Y.; Chen, W.; Li, L.; et al. Fibroblast growth factor 21 in metabolic syndrome. Front. Endocrinol. 2023, 14, 1220426. [Google Scholar] [CrossRef]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef]
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as Modulator of Metabolism in Health and Disease. Front. Physiol. 2019, 10, 419. [Google Scholar] [CrossRef]
- Sun, W.; Nie, T.; Li, K.; Wu, W.; Long, Q.; Feng, T.; Mao, L.; Gao, Y.; Liu, Q.; Gao, X.; et al. Hepatic CPT1A Facilitates Liver-Adipose Cross-Talk via Induction of FGF21 in Mice. Diabetes 2021, 71, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Mo, W.; Feng, J.; Li, J.; Yu, Q.; Li, S.; Zhang, J.; Chen, K.; Ji, J.; Dai, W.; et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br. J. Pharmacol. 2020, 177, 3760–3777. [Google Scholar] [CrossRef]
- Watanabe, M.; Risi, R.; Camajani, E.; Contini, S.; Persichetti, A.; Tuccinardi, D.; Ernesti, I.; Mariani, S.; Lubrano, C.; Genco, A.; et al. Baseline HOMA IR and Circulating FGF21 Levels Predict NAFLD Improvement in Patients Undergoing a Low Carbohydrate Dietary Intervention for Weight Loss: A Prospective Observational Pilot Study. Nutrients 2020, 12, 2141. [Google Scholar] [CrossRef]
- Sarruf, D.A.; Thaler, J.P.; Morton, G.J.; German, J.; Fischer, J.D.; Ogimoto, K.; Schwartz, M.W. Fibroblast growth factor 21 action in the brain increases energy expenditure and insulin sensitivity in obese rats. Diabetes 2010, 59, 1817–1824. [Google Scholar] [CrossRef]
- Gómez-Ambrosi, J.; Gallego-Escuredo, J.M.; Catalán, V.; Rodríguez, A.; Domingo, P.; Moncada, R.; Valentí, V.; Salvador, J.; Giralt, M.; Villarroya, F.; et al. FGF19 and FGF21 serum concentrations in human obesity and type 2 diabetes behave differently after diet- or surgically-induced weight loss. Clin. Nutr. 2017, 36, 861–868. [Google Scholar] [CrossRef]
- Harrison, S.A.; Rolph, T.; Knott, M.; Dubourg, J. FGF21 agonists: An emerging therapeutic for metabolic dysfunction-associated steatohepatitis and beyond. J. Hepatol. 2024, 81, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Sanyal, A.J.; Kowdley, K.V.; Bhatt, D.L.; Alkhouri, N.; Frias, J.P.; Bedossa, P.; Harrison, S.A.; Lazas, D.; Barish, R.; et al. Randomized, Controlled Trial of the FGF21 Analogue Pegozafermin in NASH. N. Engl. J. Med. 2023, 389, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Markan, K.R. Defining “FGF21 Resistance” during obesity: Controversy, criteria and unresolved questions. F1000Research 2018, 7, 289. [Google Scholar] [CrossRef]
- Li, H.; Wu, G.; Fang, Q.; Zhang, M.; Hui, X.; Sheng, B.; Wu, L.; Bao, Y.; Li, P.; Xu, A.; et al. Fibroblast growth factor 21 increases insulin sensitivity through specific expansion of subcutaneous fat. Nat. Commun. 2018, 9, 272. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Kolb, H.; Kempf, K.; Röhling, M.; Lenzen-Schulte, M.; Schloot, N.C.; Martin, S. Ketone bodies: From enemy to friend and guardian angel. BMC Med. 2021, 19, 313. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Muhammad, S.; Khan, M.A.; Chen, H.; Ridder, D.A.; Müller-Fielitz, H.; Pokorná, B.; Vollbrandt, T.; Stölting, I.; Nadrowitz, R.; et al. The β-hydroxybutyrate receptor HCA2 activates a neuroprotective subset of macrophages. Nat. Commun. 2014, 5, 3944. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, E.S.; Svart, M.V.; Voss, T.; Møller, N.; Bjerre, M. Impact of Acutely Increased Endogenous- and Exogenous Ketone Bodies on FGF21 Levels in Humans. Endocr. Res. 2021, 46, 20–27. [Google Scholar] [CrossRef]
- Katsu-Jiménez, Y.; Giménez-Cassina, A. Fibroblast growth Factor-21 promotes ketone body utilization in neurons through activation of AMP-dependent kinase. Mol. Cell. Neurosci. 2019, 101, 103415. [Google Scholar] [CrossRef]
- Kawakami, R.; Sunaga, H.; Iso, T.; Kaneko, R.; Koitabashi, N.; Obokata, M.; Harada, T.; Matsui, H.; Yokoyama, T.; Kurabayashi, M. Ketone body and FGF21 coordinately regulate fasting-induced oxidative stress response in the heart. Sci. Rep. 2022, 12, 7338. [Google Scholar] [CrossRef]
- Reitman, M.L. FGF21: A missing link in the biology of fasting. Cell Metab. 2007, 5, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Sietsma, S.-J.; Janssen, W.M.; Hillege, H.L.; Navis, G.; DE Zeeuw, D.; DE Jong, P.E. Urinary albumin excretion is associated with renal functional abnormalities in a nondiabetic population. J. Am. Soc. Nephrol. 2000, 11, 1882–1888. [Google Scholar] [CrossRef]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P.; STROBE Initiative. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. J. Clin. Epidemiol. 2008, 61, 344–349. [Google Scholar] [CrossRef]
- Kunutsor, S.K.; Bakker, S.J.; Kootstra-Ros, J.E.; Blokzijl, H.; Gansevoort, R.T.; Dullaart, R.P. Inverse linear associations between liver aminotransferases and incident cardiovascular disease risk: The PREVEND study. Atherosclerosis 2015, 243, 138–147. [Google Scholar] [CrossRef]
- Dullaart, R.P.; Perton, F.; van der Klauw, M.M.; Hillege, H.L.; Sluiter, W.J.; PREVEND Study Group. High plasma lecithin:cholesterol acyltransferase activity does not predict low incidence of cardiovascular events: Possible attenuation of cardioprotection asso-ciated with high HDL cholesterol. Atherosclerosis 2010, 208, 537–542. [Google Scholar] [CrossRef]
- Kouvari, M.; Valenzuela-Vallejo, L.; Guatibonza-Garcia, V.; Polyzos, S.A.; Deng, Y.; Kokkorakis, M.; Agraz, M.; Mylonakis, S.C.; Katsarou, A.; Verrastro, O.; et al. Liver biopsy-based validation, confirmation and comparison of the diagnostic performance of established and novel non-invasive steatotic liver disease indexes: Results from a large multi-center study. Metabolism 2023, 147, 155666. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef] [PubMed]
- Bedogni, G.; Bellentani, S.; Miglioli, L.; Masutti, F.; Passalacqua, M.; Castiglione, A.; Tiribelli, C. The Fatty Liver Index: A simple and accurate predictor of hepatic steatosis in the general population. BMC Gastroenterol. 2006, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Crudele, L.; De Matteis, C.; Novielli, F.; Di Buduo, E.; Petruzzelli, S.; De Giorgi, A.; Antonica, G.; Berardi, E.; Moschetta, A. Fatty Liver Index (FLI) is the best score to predict MASLD with 50% lower cut-off value in women than in men. Biol. Sex Differ. 2024, 15, 43. [Google Scholar] [CrossRef]
- Harel, O.; Zhou, X. Multiple imputation: Review of theory, implementation and software. Stat. Med. 2007, 26, 3057–3077. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; White, I.R.; Carlin, J.B.; Spratt, M.; Royston, P.; Kenward, M.G.; Wood, A.M.; Carpenter, J.R. Multiple imputation for missing data in epidemiological and clinical research: Potential and pitfalls. BMJ 2009, 338, b2393. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Value |
|---|---|
| Circulating FGF21 (pg/mL) | 888 [534–1374] |
| Ketone bodies (µmol/L) | 176 [139–247] |
| Age (years) | 53 ± 12 |
| Sex (Male, %) | 2968 (49) |
| Waist circumference (cm) | 91 [82–100] |
| eGFR (mL/min/1.73 m2) | 97 ± 17 |
| Urinary albumin excretion (mg/day) | 9 [6–15] |
| Alcohol intake (>1 drink per day/%) | 1558 (26) |
| Smoking status (n/%) | 1689 (28) |
| Total cholesterol (mmol/L) | 5.4 ± 1.0 |
| HDL cholesterol (mmol/L) | 1.3 ± 0.3 |
| Systolic blood pressure (mmHg) | 126 ± 19 |
| Diabetes (n/%) | 324 (5) |
| Fatty Liver Index | 36 [14–67] |
| MASLD (n/%) | 1864 (31) |
| Circulating FGF21 Concentration | Circulating Ketone Bodies | Interaction Between FGF21 and Ketone Bodies | |||
|---|---|---|---|---|---|
| Model | OR (95% CI) | p-Value | OR (95% CI) | p-Value | p-Value |
| Model 1 | 1.93 (1.81–2.05) | <0.001 | 1.29 (1.19–2.05) | <0.001 | 0.004 |
| Model 2 | 1.98 (1.86–2.12) | <0.001 | 1.21 (1.11–2.12) | <0.001 | 0.03 |
| Model 3 | 1.91 (1.79–2.05) | <0.001 | 1.17 (1.07–1.28) | <0.001 | 0.03 |
| Model 4 | 1.82 (1.64–2.02) | <0.001 | 1.19 (1.03–1.38) | 0.01 | 0.01 |
| Events, n (%) | 1864 (31%) | 1864 (31%) | |||
| Analyses of FGF21 with MASLD in Participants with Ketone Bodies Below or Above the Median | |||
| Events, n (%) | OR (95% CI) per Doubling | p-Value | |
| Subgroup | |||
| Ketone bodies < 176 µM | 768 (26%) | 1.58 (1.35–1.85) | <0.001 |
| Ketone bodies > 176 µM | 1096 (36%) | 2.05 (1.78–2.37) | <0.001 |
| Analyses of Ketone Bodies with MASLD in Participants with Ketone Bodies Below or Above the Median | |||
| Events, n (%) | OR (95% CI) per Doubling | p-Value | |
| Subgroup | |||
| FGF21 < 888 pg/mL | 580 (19%) | 0.98 (0.78–1.23) | 0.86 |
| FGF21 > 888 pg/mL | 1283 (43%) | 1.41 (1.73–1.70) | <0.001 |
| Circulating FGF21 Concentration | Circulating Ketone Bodies | Interaction FGF21 and Ketone Bodies | |||
|---|---|---|---|---|---|
| Model | HR (95% CI) | p-Value | HR (95% CI) | p-Value | p-Value |
| Model 1 | 1.24 (1.16–1.32) | <0.001 | 1.46 (1.34–1.59) | <0.001 | 0.038 |
| Model 2 | 1.13 (1.08–1.30) | <0.001 | 1.18 (1.08–1.30) | <0.001 | 0.044 |
| Model 3 | 1.08 (1.01–1.16) | 0.029 | 1.17 (1.06–1.29) | 0.001 | 0.003 |
| Model 4 | 1.05 (0.97–1.13) | 0.17 | 1.16 (1.06–1.28) | 0.002 | 0.005 |
| Events, n (%) | 843 (14%) | 843 (14%) | |||
| Analyses of FGF21 with Mortality in Participants with Ketone Bodies Below or Above the Median | |||
| Events, n (%) | HR (95% CI) per Doubling | p-Value | |
| Subgroup | |||
| Ketone bodies < 176 µM | 306 (10%) | 1.03 (0.91–1.16) | 0.67 |
| Ketone bodies > 176 µM | 537 (18%) | 1.11 (1.02–1.21) | 0.022 |
| Analyses of Ketone Bodies with Mortality in Participants with Ketone Bodies Below or Above the Median | |||
| Events, n (%) | HR (95% CI) per Doubling | p-Value | |
| Subgroup | |||
| FGF21 < 888 pg/mL | 334 (11%) | 1.11 (0.96–1.28) | 0.17 |
| FGF21 > 888 pg/mL | 508 (17%) | 1.26 (1.11–1.43) | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chvatal-Medina, M.; Li, Y.; Dam, W.A.; Connelly, M.A.; Moshage, H.; Bakker, S.J.L.; Dullaart, R.P.F.; Post, A. Circulating FGF21 and Ketone Bodies Modify the Risk of MASLD and Mortality: Insights from the PREVEND Cohort Study. Int. J. Mol. Sci. 2025, 26, 5059. https://doi.org/10.3390/ijms26115059
Chvatal-Medina M, Li Y, Dam WA, Connelly MA, Moshage H, Bakker SJL, Dullaart RPF, Post A. Circulating FGF21 and Ketone Bodies Modify the Risk of MASLD and Mortality: Insights from the PREVEND Cohort Study. International Journal of Molecular Sciences. 2025; 26(11):5059. https://doi.org/10.3390/ijms26115059
Chicago/Turabian StyleChvatal-Medina, Mateo, Yakun Li, Wendy A. Dam, Margery A. Connelly, Han Moshage, Stephan J. L. Bakker, Robin P. F. Dullaart, and Adrian Post. 2025. "Circulating FGF21 and Ketone Bodies Modify the Risk of MASLD and Mortality: Insights from the PREVEND Cohort Study" International Journal of Molecular Sciences 26, no. 11: 5059. https://doi.org/10.3390/ijms26115059
APA StyleChvatal-Medina, M., Li, Y., Dam, W. A., Connelly, M. A., Moshage, H., Bakker, S. J. L., Dullaart, R. P. F., & Post, A. (2025). Circulating FGF21 and Ketone Bodies Modify the Risk of MASLD and Mortality: Insights from the PREVEND Cohort Study. International Journal of Molecular Sciences, 26(11), 5059. https://doi.org/10.3390/ijms26115059