
Different Kynurenine Pathway Dysregulation in Systemic Sclerosis in Men and Women

 ,
,  ,
,  and
and
Abstract
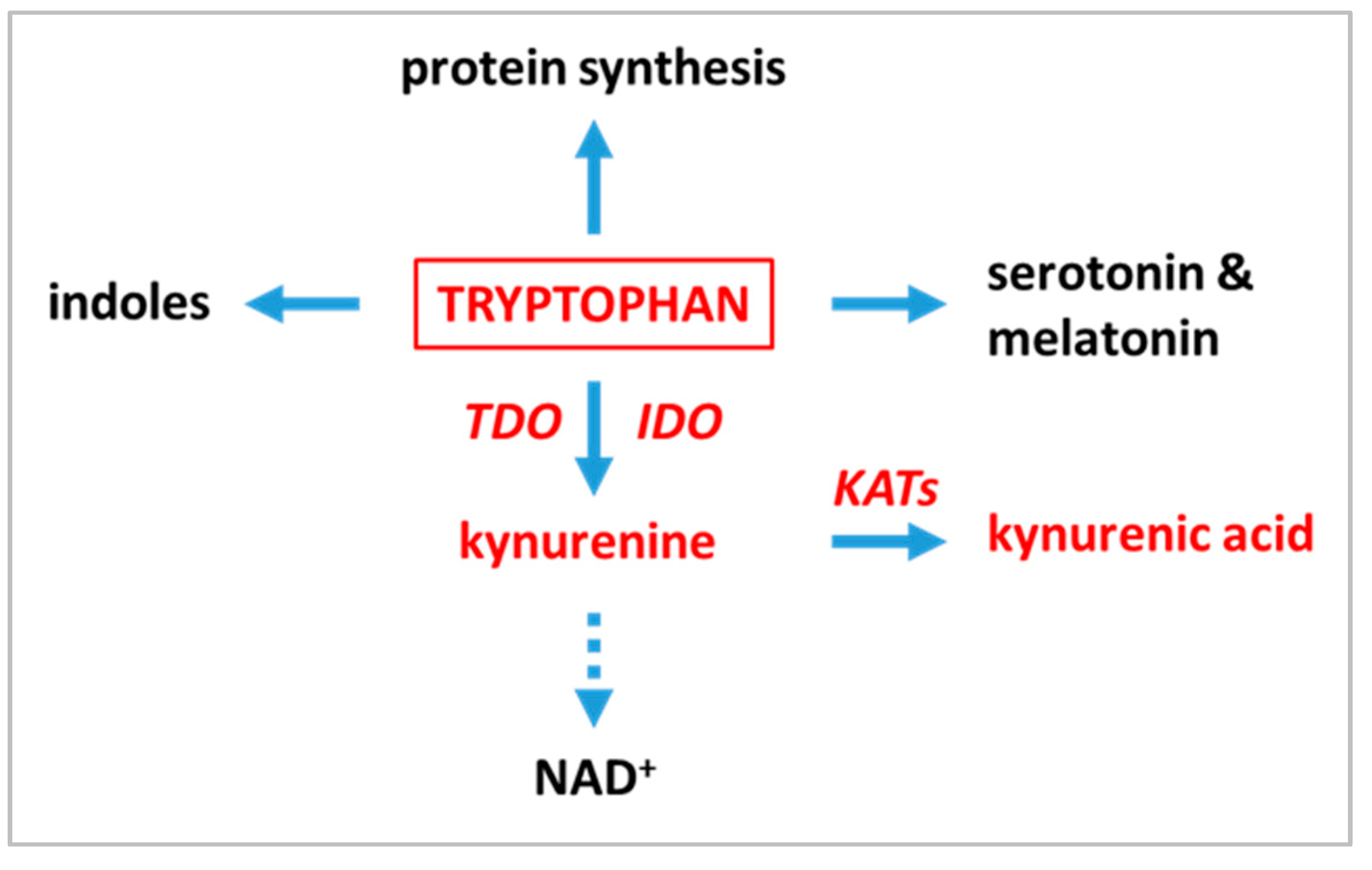
1. Introduction
2. Results
2.1. General Characteristics of the Sample
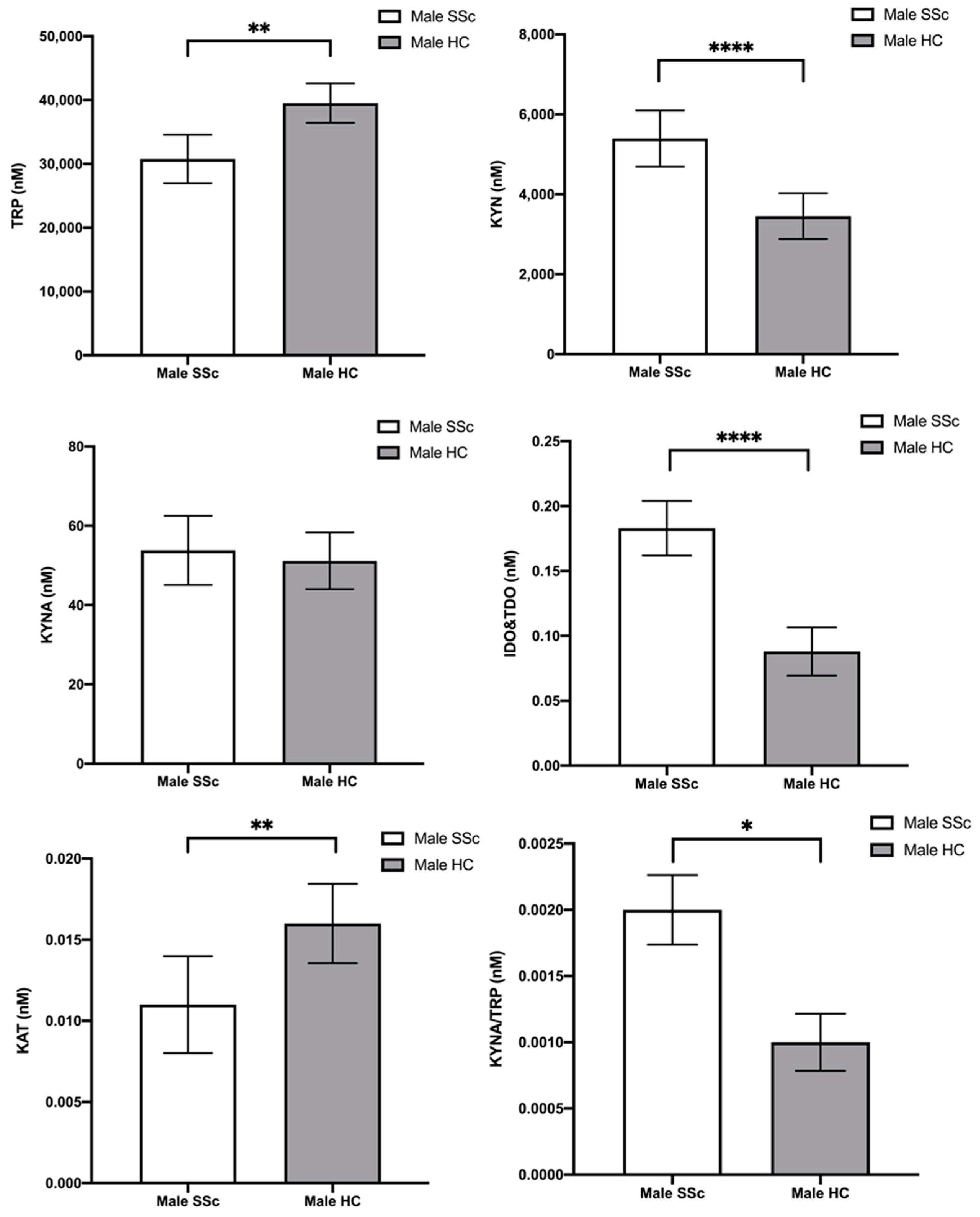
2.2. Tryptophan and Its Metabolites—Differences between Male Scleroderma Patients and Male Healthy Controls
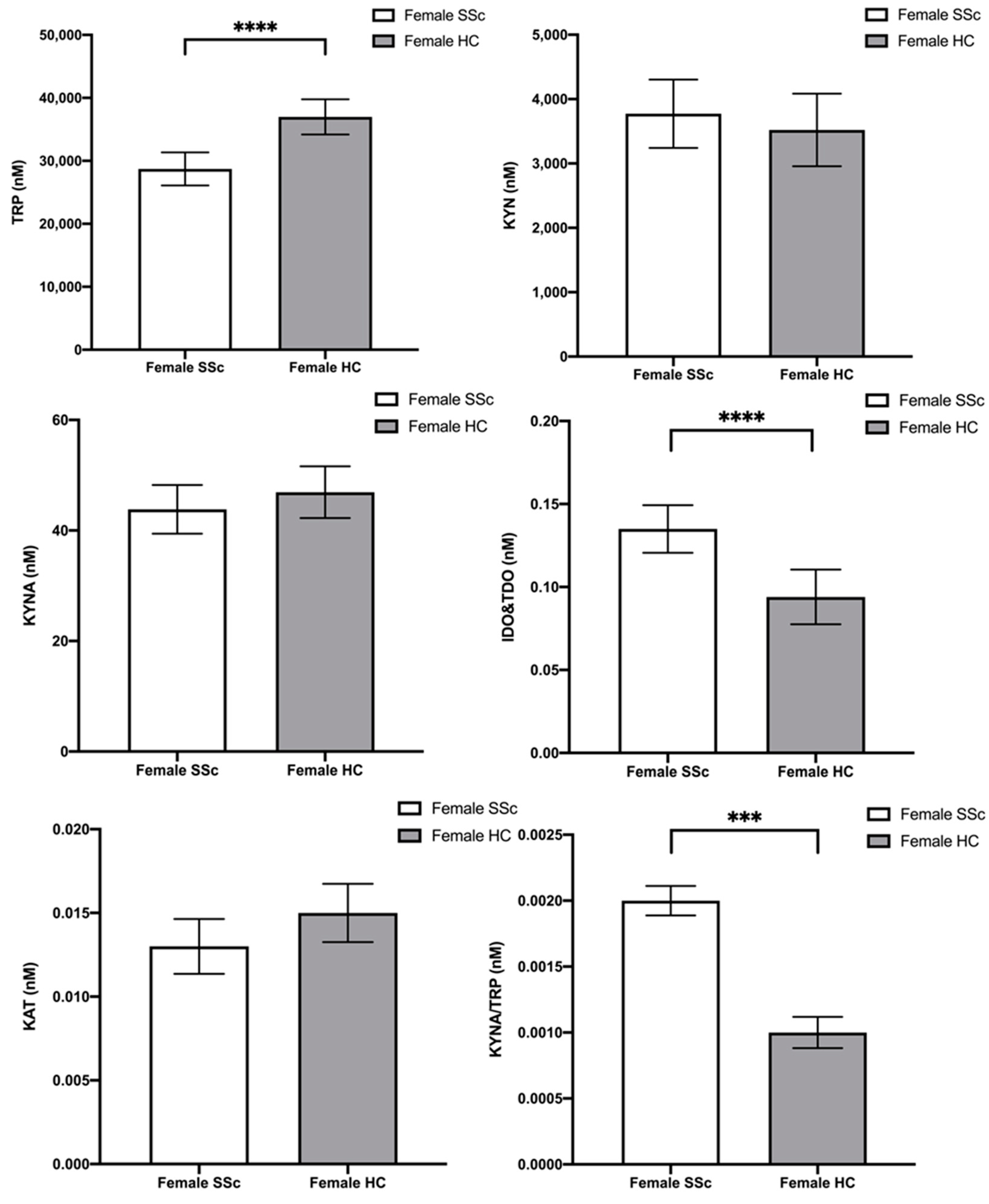
2.3. Tryptophan and Its Metabolites—Differences between Female Scleroderma Patients and Female Healthy Controls
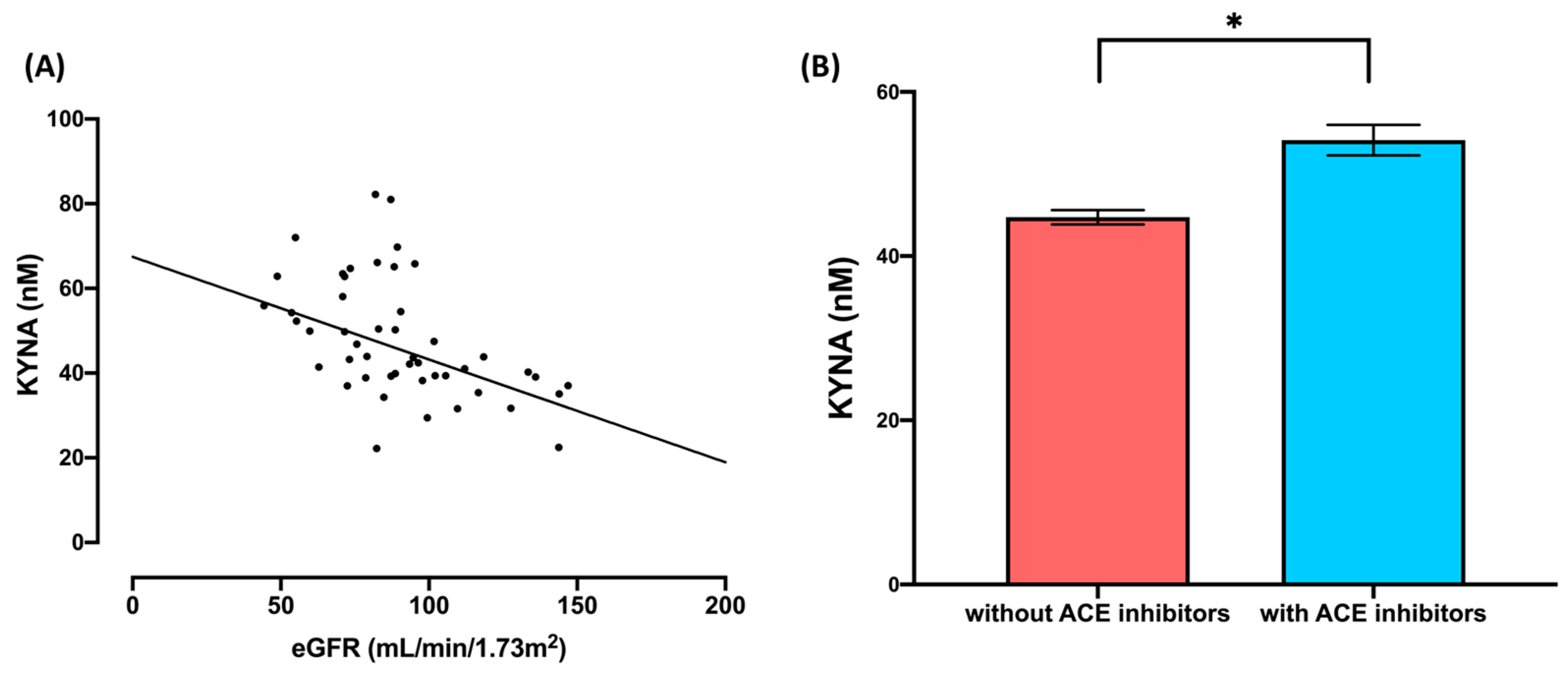
2.4. Tryptophan and Its Metabolites—Effects of the eGFR and Use of ACE Inhibitors
3. Discussion
Study Limitations
4. Materials and Methods
4.1. Participants and Selection Criteria
4.2. Chromatographic Analysis
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bairkdar, M.; Rossides, M.; Westerlind, H.; Hesselstrand, R.; Arkema, E.V.; Holmqvist, M. Incidence and Prevalence of Systemic Sclerosis Globally: A Comprehensive Systematic Review and Meta-Analysis. Rheumatology 2021, 60, 3121–3133. [Google Scholar] [CrossRef] [PubMed]
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic Sclerosis. Nat. Rev. Dis. Primers 2015, 1, 15002. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.; Carreira, P.E.; et al. 2013 Classification Criteria for Systemic Sclerosis: An American College of Rheumatology/European League against Rheumatism Collaborative Initiative. Ann. Rheum. Dis. 2013, 72, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J. The Kynurenine Pathway: A Finger in Every Pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.E.; Ambade, A.S.; Harlan, R.; Roux, A.; Aja, S.; Graham, D.; Shah, A.A.; Hummers, L.K.; Hemnes, A.R.; Leopold, J.A.; et al. Kynurenine Pathway Metabolism Evolves with Development of Preclinical and Scleroderma-Associated Pulmonary Arterial Hypertension. Am. J. Physiol.—Lung Cell Mol. Physiol. 2023, 325, L617–L627. [Google Scholar] [CrossRef] [PubMed]
- Morales-González, V.; Galeano-Sánchez, D.; Covaleda-Vargas, J.E.; Rodriguez, Y.; Monsalve, D.M.; Pardo-Rodriguez, D.; Cala, M.P.; Acosta-Ampudia, Y.; Ramírez-Santana, C. Metabolic Fingerprinting of Systemic Sclerosis: A Systematic Review. Front. Mol. Biosci. 2023, 10, 1215039. [Google Scholar] [CrossRef] [PubMed]
- Bögl, T.; Mlynek, F.; Himmelsbach, M.; Sepp, N.; Buchberger, W.; Geroldinger-Simić, M. Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis. Biomedicines 2022, 10, 607. [Google Scholar] [CrossRef]
- Rodgers, J.; Stone, T.W.; Barrett, M.P.; Bradley, B.; Kennedy, P.G.E. Kynurenine Pathway Inhibition Reduces Central Nervous System Inflammation in a Model of Human African Trypanosomiasis. Brain 2009, 132, 1259–1267. [Google Scholar] [CrossRef]
- Houpt, J.B.; Ogryzlo, M.A.; Hunt, M. Tryptophan Metabolism in Man (with Special Reference to Rheumatoid Arthritis and Scleroderma). Semin. Arthritis Rheum. 1973, 2, 333–353. [Google Scholar] [CrossRef]
- Pinals, R.S. Tryptophan Metabolism in Rheumatic Disease. Arthritis Rheum. 1964, 7, 662–669. [Google Scholar] [CrossRef]
- Binazzi, M.; Calandra, P. Tryptophan to Niacin Pathway in Scleroderma and in Dermatomyositis. Arch. Dermatol. Forsch. 1973, 246, 142–145. [Google Scholar] [CrossRef]
- De Antoni, A.; Muggeo, M.; Costa, C.; Allegri, G.; Crepaldi, G. Tryptophan Metabolism “via” Nicotimic Acid in Patients with Scleroderma. Acta Vitaminol. Enzym. 1976, 30, 134–139. [Google Scholar]
- Poormasjedi-Meibod, M.-S.; Hartwell, R.; Kilani, R.T.; Ghahary, A. Anti-Scarring Properties of Different Tryptophan Derivatives. PLoS ONE 2014, 9, e91955. [Google Scholar] [CrossRef]
- Li, Y.; Kilani, R.T.; Rahmani-Neishaboor, E.; Jalili, R.B.; Ghahary, A. Kynurenine Increases Matrix Metalloproteinase-1 and -3 Expression in Cultured Dermal Fibroblasts and Improves Scarring in Vivo. J. Investig. Dermatol. 2014, 134, 643–650. [Google Scholar] [CrossRef]
- Nestor, M.S.; Berman, B.; Fischer, D.L.; Han, H.; Gade, A.; Arnold, D.; Lawson, A. A Randomized, Double-Blind, Active- and Placebo-Controlled Trial Evaluating a Novel Topical Treatment for Keloid Scars. J. Drugs Dermatol. 2021, 20, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Papp, A.; Hartwell, R.; Evans, M.; Ghahary, A. The Safety and Tolerability of Topically Delivered Kynurenic Acid in Humans. A Phase 1 Randomized Double-Blind Clinical Trial. J. Pharm. Sci. 2018, 107, 1572–1576. [Google Scholar] [CrossRef]
- Sagonas, I.; Daoussis, D. Serotonin and Systemic Sclerosis. An Emerging Player in Pathogenesis. Jt. Bone Spine 2022, 89, 105309. [Google Scholar] [CrossRef] [PubMed]
- Peoples, C.; Medsger, T.A.; Lucas, M.; Rosario, B.L.; Feghali-Bostwick, C.A. Gender Differences in Systemic Sclerosis: Relationship to Clinical Features, Serologic Status and Outcomes. J. Scleroderma Relat. Disord. 2016, 1, 204–212. [Google Scholar] [CrossRef]
- Hughes, M.; Pauling, J.D.; Armstrong-James, L.; Denton, C.P.; Galdas, P.; Flurey, C. Gender-Related Differences in Systemic Sclerosis. Autoimmun. Rev. 2020, 19, 102494. [Google Scholar] [CrossRef]
- Liem, S.I.E.; Ciaffi, J.; Van Leeuwen, N.M.; Boonstra, M.; Ahmed, S.; Beaart-van De Voorde, L.J.J.; Corsel, A.; Dhondai, T.; Ninaber, M.K.; Geelhoed-Veltman, J.J.M.; et al. Step Forward in Early Recognition of Systemic Sclerosis: Data from the Leiden CCISS Cohort. RMD Open 2023, 9, e002971. [Google Scholar] [CrossRef]
- De Angelis, R.; Giuggioli, D.; Bajocchi, G.; Dagna, L.; Zanframundo, G.; Foti, R.; Cacciapaglia, F.; Cuomo, G.; Ariani, A.; Rosato, E.; et al. Sex-Related Differences in Systemic Sclerosis: A Multicenter Cross-Sectional Study From the National Registry of the Italian Society for Rheumatology. J. Rheumatol. 2022, 49, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Volkmann, E.R.; Andréasson, K.; Smith, V. Systemic Sclerosis. Lancet 2023, 401, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.; Herrick, A.L. Systemic Sclerosis. Br. J. Hosp. Med. 2019, 80, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Csipô, I.; Czirják, L.; Szántó, S.; Szerafin, L.; Sipka, S.; Szegedi, G. Decreased Serum Tryptophan and Elevated Neopterin Levels in Systemic Sclerosis. Clin. Exp. Rheumatol. 1995, 13, 269–270. [Google Scholar] [PubMed]
- Bengtsson, A.A.; Trygg, J.; Wuttge, D.M.; Sturfelt, G.; Theander, E.; Donten, M.; Moritz, T.; Sennbro, C.-J.; Torell, F.; Lood, C.; et al. Metabolic Profiling of Systemic Lupus Erythematosus and Comparison with Primary Sjögren’s Syndrome and Systemic Sclerosis. PLoS ONE 2016, 11, e0159384. [Google Scholar] [CrossRef] [PubMed]
- Campochiaro, C.; Lytton, S.; Nihtyanova, S.; Fuchs, D.; Ong, V.H.; Denton, C.P. Elevated Kynurenine Levels in Diffuse Cutaneous and Anti-RNA Polymerase III Positive Systemic Sclerosis. Clin. Immunol. 2019, 199, 18–24. [Google Scholar] [CrossRef]
- Meier, C.; Freiburghaus, K.; Bovet, C.; Schniering, J.; Allanore, Y.; Distler, O.; Nakas, C.; Maurer, B. Serum Metabolites as Biomarkers in Systemic Sclerosis-Associated Interstitial Lung Disease. Sci. Rep. 2020, 10, 21912. [Google Scholar] [CrossRef] [PubMed]
- Smolenska, Z.; Zabielska-Kaczorowska, M.; Wojteczek, A.; Kutryb-Zajac, B.; Zdrojewski, Z. Metabolic Pattern of Systemic Sclerosis: Association of Changes in Plasma Concentrations of Amino Acid-Related Compounds with Disease Presentation. Front. Mol. Biosci. 2020, 7, 585161. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, D.; Jiang, Y.; Chen, W.; Zhao, L.; Bao, D.; Li, Y.; Distler, J.H.W.; Zhu, H. Serum Metabolomic Profiling Reveals Potential Biomarkers in Systemic Sclerosis. Metabolism 2023, 144, 155587. [Google Scholar] [CrossRef]
- Pellicano, C.; Vaiarello, V.; Colalillo, A.; Gigante, A.; Iannazzo, F.; Rosato, E. Role of Kinurenic Acid in the Systemic Sclerosis Renal Involvement. Clin. Exp. Med. 2022, 23, 1713–1719. [Google Scholar] [CrossRef]
- Amobi, A.; Qian, F.; Lugade, A.A.; Odunsi, K. Tryptophan Catabolism and Cancer Immunotherapy Targeting IDO Mediated Immune Suppression. In Tumor Immune Microenvironment in Cancer Progression and Cancer Therapy; Kalinski, P., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 1036, pp. 129–144. ISBN 978-3-319-67575-6. [Google Scholar]
- Stone, T.W.; Williams, R.O. Modulation of T Cells by Tryptophan Metabolites in the Kynurenine Pathway. Trends Pharmacol. Sci. 2023, 44, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Wang, Y.; Wang, Y.; Cheng, M.; Dong, W.; Wei, W.; Zhao, Y.; Chang, Y. TDO2-Overexpressed Dendritic Cells Possess Tolerogenicity and Ameliorate Collagen-Induced Arthritis by Modulating the Th17/Regulatory T Cell Balance. J. Immunol. 2024, 212, 941–950. [Google Scholar] [CrossRef]
- Klaessens, S.; Stroobant, V.; De Plaen, E.; Van den Eynde, B.J. Systemic Tryptophan Homeostasis. Front. Mol. Biosci. 2022, 9, 897929. [Google Scholar] [CrossRef]
- Liu, C.; Bonaventure, P.; Lee, G.; Nepomuceno, D.; Kuei, C.; Wu, J.; Li, Q.; Joseph, V.; Sutton, S.W.; Eckert, W.; et al. GPR139, an Orphan Receptor Highly Enriched in the Habenula and Septum, Is Activated by the Essential Amino Acids l-Tryptophan and l-Phenylalanine. Mol. Pharmacol. 2015, 88, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Murakoshi, M.; Kuwabara, H.; Nagasaki, M.; Xiong, Y.M.; Reagan, J.D.; Maeda, H.; Nara, F. Discovery and Pharmacological Effects of a Novel GPR142 Antagonist. J. Recept. Signal Transduct. 2017, 37, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Solvay, M.; Holfelder, P.; Klaessens, S.; Pilotte, L.; Stroobant, V.; Lamy, J.; Naulaerts, S.; Spillier, Q.; Frédérick, R.; De Plaen, E.; et al. Tryptophan Depletion Sensitizes the AHR Pathway by Increasing AHR Expression and GCN2/LAT1-Mediated Kynurenine Uptake, and Potentiates Induction of Regulatory T Lymphocytes. J. Immunother. Cancer 2023, 11, e006728. [Google Scholar] [CrossRef]
- Jin, W.; Zheng, Y.; Zhu, P. T Cell Abnormalities in Systemic Sclerosis. Autoimmun. Rev. 2022, 21, 103185. [Google Scholar] [CrossRef]
- Mellor, A.L.; Munn, D.H. Tryptophan Catabolism and T-Cell Tolerance: Immunosuppression by Starvation? Immunol. Today 1999, 20, 469–473. [Google Scholar] [CrossRef]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An Endogenous Tumour-Promoting Ligand of the Human Aryl Hydrocarbon Receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef]
- Seo, S.-K.; Kwon, B. Immune Regulation through Tryptophan Metabolism. Exp. Mol. Med. 2023, 55, 1371–1379. [Google Scholar] [CrossRef]
- Shi, Y.; Tang, B.; Yu, J.; Luo, Y.; Xiao, Y.; Pi, Z.; Tang, R.; Wang, Y.; Kanekura, T.; Zeng, Z.; et al. Aryl Hydrocarbon Receptor Signaling Activation in Systemic Sclerosis Attenuates Collagen Production and Is a Potential Antifibrotic Target. Int. Immunopharmacol. 2020, 88, 106886. [Google Scholar] [CrossRef]
- Shi, Y.; Xiao, Y.; Yu, J.; Liu, J.; Liu, L.; Ding, Y.; Qiu, X.; Zhan, Y.; Tang, R.; Zeng, Z.; et al. UVA1 Irradiation Attenuates Collagen Production via Ficz/AhR/MAPK Signaling Activation in Scleroderma. Int. Immunopharmacol. 2023, 116, 109764. [Google Scholar] [CrossRef]
- DiNatale, B.C.; Murray, I.A.; Schroeder, J.C.; Flaveny, C.A.; Lahoti, T.S.; Laurenzana, E.M.; Omiecinski, C.J.; Perdew, G.H. Kynurenic Acid Is a Potent Endogenous Aryl Hydrocarbon Receptor Ligand That Synergistically Induces Interleukin-6 in the Presence of Inflammatory Signaling. Toxicol. Sci. 2010, 115, 89–97. [Google Scholar] [CrossRef]
- Burckhardt, G. Drug Transport by Organic Anion Transporters (OATs). Pharm. Ther. 2012, 136, 106–130. [Google Scholar] [CrossRef]
- Wu, W.; Bush, K.T.; Nigam, S.K. Key Role for the Organic Anion Transporters, OAT1 and OAT3, in the in Vivo Handling of Uremic Toxins and Solutes. Sci. Rep. 2017, 7, 4939. [Google Scholar] [CrossRef]
- Kaszaki, J.; Palásthy, Z.; Erczes, D.; Rácz, A.; Torday, C.; Varga, G.; Vécsei, L.; Boros, M. Kynurenic Acid Inhibits Intestinal Hypermotility and Xanthine Oxidase Activity during Experimental Colon Obstruction in Dogs. Neurogastroenterol. Motil. 2008, 20, 53–62. [Google Scholar] [CrossRef]
- Varga, G.; Erces, D.; Fazekas, B.; Fülöp, M.; Kovács, T.; Kaszaki, J.; Fülöp, F.; Vécsei, L.; Boros, M. N-Methyl-D-Aspartate Receptor Antagonism Decreases Motility and Inflammatory Activation in the Early Phase of Acute Experimental Colitis in the Rat. Neurogastroenterol. Motil. 2010, 22, 217–225.e68. [Google Scholar] [CrossRef]
- Ramírez Ortega, D.; Ugalde Muñiz, P.E.; Blanco Ayala, T.; Vázquez Cervantes, G.I.; Lugo Huitrón, R.; Pineda, B.; González Esquivel, D.F.; Pérez de la Cruz, G.; Pedraza Chaverrí, J.; Sánchez Chapul, L.; et al. On the Antioxidant Properties of L-Kynurenine: An Efficient ROS Scavenger and Enhancer of Rat Brain Antioxidant Defense. Antioxidants 2021, 11, 31. [Google Scholar] [CrossRef]
- Lugo-Huitrón, R.; Blanco-Ayala, T.; Ugalde-Muñiz, P.; Carrillo-Mora, P.; Pedraza-Chaverrí, J.; Silva-Adaya, D.; Maldonado, P.D.; Torres, I.; Pinzón, E.; Ortiz-Islas, E.; et al. On the Antioxidant Properties of Kynurenic Acid: Free Radical Scavenging Activity and Inhibition of Oxidative Stress. Neurotoxicol. Teratol. 2011, 33, 538–547. [Google Scholar] [CrossRef]
- Jayawickrama, G.S.; Nematollahi, A.; Sun, G.; Gorrell, M.D.; Church, W.B. Inhibition of Human Kynurenine Aminotransferase Isozymes by Estrogen and Its Derivatives. Sci. Rep. 2017, 7, 17559. [Google Scholar] [CrossRef]
- Mason, M.; Gullekson, E.H. Estrogen-Enzyme Interactions: Inhibition and Protection of Kynurenine Transaminase by the Sulfate Esters of Diethylstilbestrol, Estradiol, and Estrone. JBC 1960, 235, 1312–1316. [Google Scholar] [CrossRef]
- De Bie, J.; Lim, C.K.; Guillemin, G.J. Progesterone Alters Kynurenine Pathway Activation in IFN-γ-Activated Macrophages—Relevance for Neuroinflammatory Diseases. Int. J. Tryptophan Res. 2016, 9, IJTR.S40332. [Google Scholar] [CrossRef]
- Ciaffi, J.; Morabito, M.F.; Ruscitti, P.; D’Angelo, S.; Mancarella, L.; Brusi, V.; Abignano, G.; Pucino, V.; Giacomelli, R.; Meliconi, R.; et al. Incidence, Prevalence and Mortality of Systemic Sclerosis in Italy: A Nationwide Population-Based Study Using Administrative Health Data. Rheumatol. Int. 2021, 41, 129–137. [Google Scholar] [CrossRef]
- Jutiviboonsuk, A.; Salang, L.; Eamudomkarn, N.; Mahakkanukrauh, A.; Suwannaroj, S.; Foocharoen, C. Prevalence and Clinical Associations with Premature Ovarian Insufficiency, Early Menopause, and Low Ovarian Reserve in Systemic Sclerosis. Clin. Rheumatol. 2021, 40, 2267–2275. [Google Scholar] [CrossRef]
- Zhao, J.; Gao, P.; Zhu, D. Optimization of Zn2+-Containing Mobile Phase for Simultaneous Determination of Kynurenine, Kynurenic Acid and Tryptophan in Human Plasma by High Performance Liquid Chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 603–608. [Google Scholar] [CrossRef]
- Inker, L.A.; Eneanya, N.D.; Coresh, J.; Tighiouart, H.; Wang, D.; Sang, Y.; Crews, D.C.; Doria, A.; Estrella, M.M.; Froissart, M.; et al. New Creatinine- and Cystatin C–Based Equations to Estimate GFR without Race. N. Engl. J. Med. 2021, 385, 1737–1749. [Google Scholar] [CrossRef]
- Morris, P.E.; Fritz, C.O. Effect Sizes in Memory Research. Memory 2013, 21, 832–842. [Google Scholar] [CrossRef]
- University of Cambridge. Rules of Thumb on Magnitudes of Effect Sizes. Available online: http://imaging.mrc-cbu.cam.ac.uk/statswiki/FAQ/effectSize (accessed on 21 February 2020).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSc Patients (n = 48) | HC (n = 53) | t- or χ2-Test | p < | |
|---|---|---|---|---|
| Age | 51.17 (13.15) | 46.77 (13.36) | 1.66 | 0.100 |
| Sex (m/f) | 19/29 | 27/26 | 1.31 | 0.252 |
| Plasma content | ||||
| [TRP] | 29,524.88 (7282.10) | 38,284.19 (7240.42) | −6.06 | <0.001 |
| [KYN] | 4452.07 (1673.82) | 3453.23 (1359.69) | 3.30 | 0.001 |
| [KYNA] | 47.87 (14.05) | 49.00 (15.93) | −0.38 | 0.706 |
| Enzyme activity * | ||||
| IDO and TDO | 0.16 (0.06) | 0.09 (0.03) | 7.28 | <0.001 |
| KATs | 0.01 (0.004) | 0.02 (0.006) | −3.64 | <0.001 |
| KYNA/TRP | 0.002 (0.0004) | 0.0013 (0.0005) | 3.93 | <0.001 |
| Multivariate Tests | |||||||
| Pillai’s Trace ** | F | df1 | df2 | p | η2p | ||
| Age | 0.134 | 0.983 | 6 | 38 | 0.450 | 0.134 | |
| Status | 0.649 | 11.701 | 6 | 38 | * <0.001 | 0.649 | |
| Univariate Tests | |||||||
| DV | Type III SS | df | MS | F | p | η2p | |
| Age | [TRP] | 6.00 × 104 | 1 | 6.00 × 104 | 0.001 | 0.975 | <0.01 |
| [KYN] | 3.87 × 106 | 1 | 3.87 × 106 | 1.853 | 0.181 | 0.04 | |
| [KYNA] | 45.60 | 1 | 45.60 | 0.141 | 0.709 | 0.003 | |
| IDO and TDO *** | 0.01 | 1 | 0.01 | 3.321 | 0.075 | 0.07 | |
| KATs *** | 5.43 × 10−5 | 1 | 5.43 × 10−5 | 1.428 | 0.239 | 0.03 | |
| [KYNA]/[TRP] | 5.49 × 10−9 | 1 | 5.49 × 10−9 | 0.019 | 0.892 | <0.01 | |
| Status | [TRP] | 8.36 × 108 | 1 | 8.36 × 108 | 13.715 | * 0.001 | 0.242 |
| [KYN] | 4.10 × 107 | 1 | 4.10 × 107 | 19.637 | * <0.001 | 0.314 | |
| [KYNA] | 74.58 | 1 | 74.58 | 0.231 | 0.633 | 0.005 | |
| IDO and TDO *** | 0.10 | 1 | 0.10 | 50.391 | * <0.001 | 0.540 | |
| KATs *** | 3.61 × 10−4 | 1 | 3.61 × 10−4 | 9.512 | * 0.004 | 0.181 | |
| [KYNA]/[TRP] | 1.89 × 10−6 | 1 | 1.89 × 10−6 | 6.418 | * 0.015 | 0.130 | |
| Residuals | [TRP] | 2.62 × 109 | 43 | 6.10 × 107 | |||
| [KYN] | 8.99 × 107 | 43 | 2.09 × 106 | ||||
| [KYNA] | 1.39 × 104 | 43 | 322.79 | ||||
| IDO and TDO *** | 0.08 | 43 | 0.002 | ||||
| KATs *** | 0.002 | 43 | 3.799 × 10−5 | ||||
| [KYNA]/[TRP] | 1.267 × 10−5 | 43 | 2.946 × 10−7 | ||||
| Multivariate Tests | |||||||
| Pillai’s Trace ** | F | df1 | df2 | p | η2p | ||
| Age | 0.175 | 1.660 | 6.000 | 47.000 | 0.152 | 0.175 | |
| Status | 0.421 | 5.707 | 6.000 | 47.000 | * <0.001 | 0.421 | |
| Univariate Tests | |||||||
| DV | Type III SS | df | MS | F | p | η2p | |
| Age | [TRP] | 4.60 × 105 | 1 | 4.60 × 105 | 0.010 | 0.922 | <0.01 |
| [KYN] | 2.01 × 106 | 1 | 2.01 × 106 | 1.042 | 0.312 | 0.02 | |
| [KYNA] | 287.36 | 1 | 287.36 | 2.157 | 0.148 | 0.04 | |
| IDO and TDO *** | 0.002 | 1 | 0.002 | 1.542 | 0.220 | 0.03 | |
| KATs *** | 1.51 × 10−7 | 1 | 1.51 × 10−7 | 0.008 | 0.928 | <0.01 | |
| [KYNA]/[TRP] | 5.19 × 10−7 | 1 | 5.19 × 10−7 | 6.076 | 0.017 | 0.11 | |
| Status | [TRP] | 9.20 × 108 | 1 | 9.20 × 108 | 19.414 | * <0.001 | 0.272 |
| [KYN] | 8.65 × 105 | 1 | 8.65 × 105 | 0.447 | 0.507 | 0.009 | |
| [KYNA] | 129.68 | 1 | 129.68 | 0.974 | 0.328 | 0.018 | |
| IDO and TDO *** | 0.02 | 1 | 0.02 | 14.284 | * <0.001 | 0.215 | |
| KATs *** | 5.17 × 10−5 | 1 | 5.17 × 10−5 | 2.816 | 0.099 | 0.051 | |
| [KYNA]/[TRP] | 9.75 × 10−7 | 1 | 9.75 × 10−7 | 11.420 | * 0.001 | 0.180 | |
| Residuals | [TRP] | 2.46 × 109 | 52 | 4.74 × 107 | |||
| [KYN] | 1.01 × 108 | 52 | 1.93 × 106 | ||||
| [KYNA] | 6926.54 | 52 | 133.20 | ||||
| IDO and TDO *** | 0.08 | 52 | 0.002 | ||||
| KATs *** | 0.001 | 52 | 1.84 × 10−5 | ||||
| [KYNA]/[TRP] | 4.44 × 10−6 | 52 | 8.54 × 10−8 | ||||
| Multivariate Tests | |||||||
| Wilks’ Lambda ** | F | df1 | df2 | p | η2p | ||
| eGFR | 0.76 | 4.07 | 3 | 38 | * 0.01 | 0.24 | |
| BMI | 0.98 | 0.29 | 3 | 38 | 0.83 | 0.02 | |
| Use of ACEi | 0.78 | 3.57 | 3 | 38 | * 0.02 | 0.22 | |
| Univariate Tests | |||||||
| DV | Type III SS | df | MS | F | p | η2p | |
| eGFR *** | [TRP] | 3.32 × 106 | 1 | 3.32 × 106 | 0.07 | 0.80 | <0.01 |
| [KYN] | 8.81 × 106 | 1 | 8.81 × 106 | 3.03 | 0.09 | 0.07 | |
| [KYNA] | 822.95 | 1 | 822.95 | 6.06 | * 0.02 | 0.13 | |
| BMI | [TRP] | 1.39 × 106 | 1 | 1.39 × 106 | 0.03 | 0.87 | <0.01 |
| [KYN] | 1.41 × 106 | 1 | 1.41 × 106 | 0.48 | 0.49 | 0.01 | |
| [KYNA] | 0.04 | 1 | 0.04 | 0.00 | 0.99 | <0.01 | |
| Use of ACEi | [TRP] | 4.75 × 107 | 1 | 4.75 × 107 | 0.97 | 0.33 | 0.02 |
| [KYN] | 19,159.33 | 1 | 19,159.33 | 0.01 | 0.94 | 0.00 | |
| [KYNA] | 762.73 | 1 | 762.73 | 5.62 | * 0.02 | 0.12 | |
| Residuals | [TRP] | 1.95 × 109 | 40 | 4.87 × 107 | |||
| [KYN] | 1.16 × 108 | 40 | 2.91 × 106 | ||||
| [KYNA] | 5430.12 | 40 | 135.75 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turska-Kozłowska, M.; Pedraz-Petrozzi, B.; Paluszkiewicz, P.; Parada-Turska, J. Different Kynurenine Pathway Dysregulation in Systemic Sclerosis in Men and Women. Int. J. Mol. Sci. 2024, 25, 3842. https://doi.org/10.3390/ijms25073842
Turska-Kozłowska M, Pedraz-Petrozzi B, Paluszkiewicz P, Parada-Turska J. Different Kynurenine Pathway Dysregulation in Systemic Sclerosis in Men and Women. International Journal of Molecular Sciences. 2024; 25(7):3842. https://doi.org/10.3390/ijms25073842
Chicago/Turabian StyleTurska-Kozłowska, Monika, Bruno Pedraz-Petrozzi, Piotr Paluszkiewicz, and Jolanta Parada-Turska. 2024. "Different Kynurenine Pathway Dysregulation in Systemic Sclerosis in Men and Women" International Journal of Molecular Sciences 25, no. 7: 3842. https://doi.org/10.3390/ijms25073842
APA StyleTurska-Kozłowska, M., Pedraz-Petrozzi, B., Paluszkiewicz, P., & Parada-Turska, J. (2024). Different Kynurenine Pathway Dysregulation in Systemic Sclerosis in Men and Women. International Journal of Molecular Sciences, 25(7), 3842. https://doi.org/10.3390/ijms25073842