

The Impact of Treadmill Training on Tissue Integrity, Axon Growth, and Astrocyte Modulation

 ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Training Treadmill Stimulates the Motor Function Recovery
2.2. Treadmill Training Maintained Tissue Integrity
2.3. Treadmill Training Stimulates Axonal Growth
2.4. Limited Impact of Treadmill Training on Motoneurons
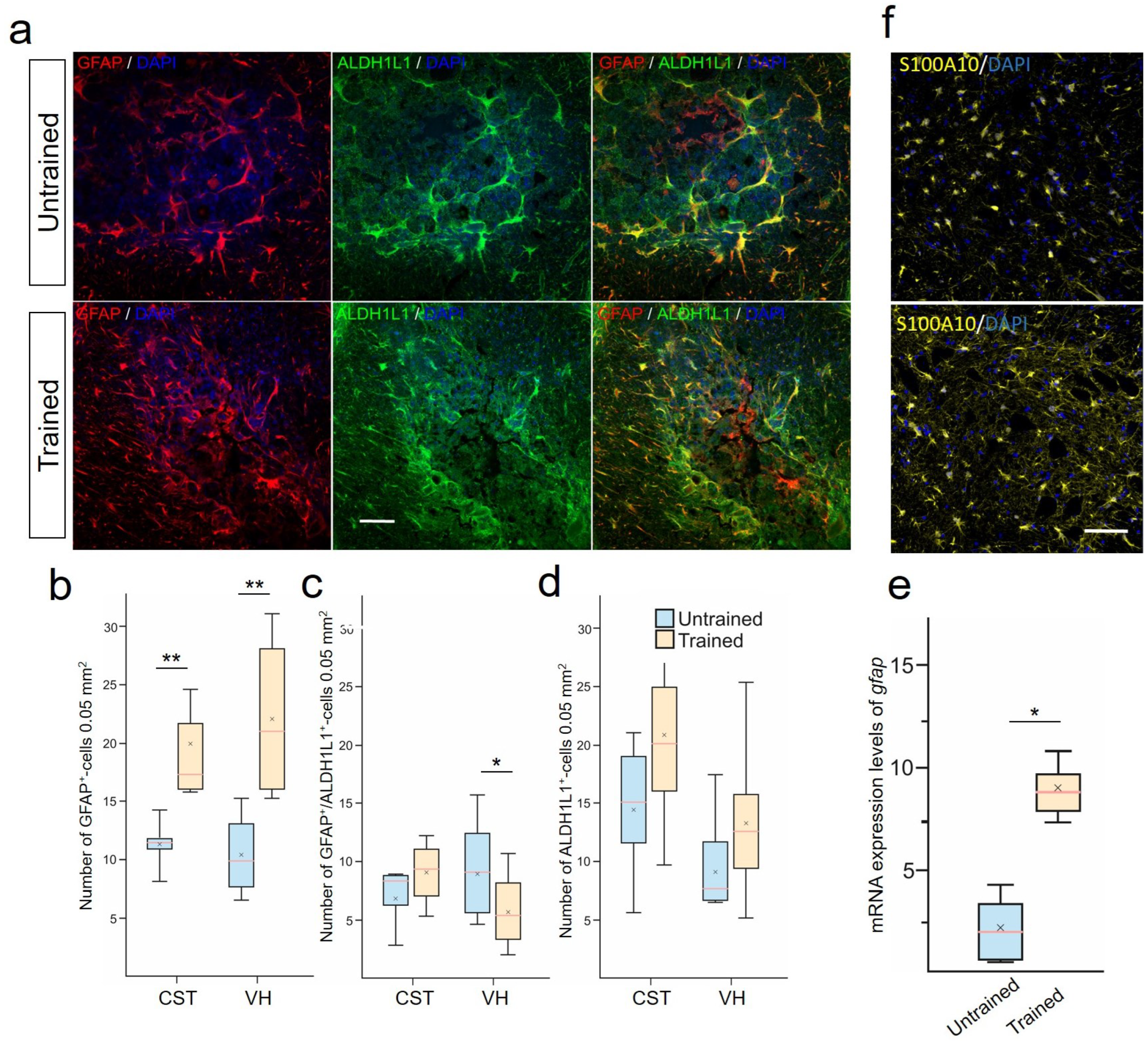
2.5. Treadmill Training Promoted Anti-Inflammatory Reactive Astrocytes
2.6. Stability of NG2 Glia and Limited Astrocytic Response to Treadmill Training Post-SCI
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Rat SCI Model Construction
4.3. Motor Training Protocol
4.4. Behavioral Test
4.5. Electromyography
4.6. Morphometric Analysis
4.7. Immunofluorescent Staining
4.8. RT-qPCR
4.9. Statistical Software and Data Analysis Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Won, S.J.; Choi, B.Y.; Yoo, B.H.; Sohn, M.; Ying, W.; Swanson, R.A.; Suh, S.W. Chronic cervical spinal cord injury: DTI correlates with clinical and electrophysiological measures. J. Neurotrauma 2012, 29, 1556–1566. [Google Scholar] [CrossRef]
- Angeli, C.A.; Edgerton, V.R.; Gerasimenko, Y.P.; Harkema, S.J. Altering spinal cord excitability enables voluntary movements after chronic complete paralysis in humans. Brain 2014, 137, 1394–1409. [Google Scholar] [CrossRef] [PubMed]
- Barthélemy, D.; Willerslev-Olsen, M.; Lundell, H.; Biering-Sørensen, F.; Nielsen, J.B. Assessment of transmission in specific descending pathways in relation to gait and balance following spinal cord injury. Prog. Brain Res. 2015, 218, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Kakulas, B.A. A review of the neuropathology of human spinal cord injury with emphasis on special features. J. Spinal Cord Med. 1999, 22, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Courtine, G.; Gerasimenko, Y.; Van Den Br, R.; Yew, A.; Musienko, P.; Zhong, H.; Song, B.; Ao, Y.; Ichiyama, R.M.; Lavrov, I.; et al. Transformation of nonfunctional spinal circuits into functional states after the loss of brain input. Nat. Neurosci. 2009, 12, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Kiehn, O. Locomotor circuits in the mammalian spinal cord. Annu. Rev. Neurosci. 2006, 29, 279–306. [Google Scholar] [CrossRef] [PubMed]
- James, N.D.; Bartus, K.; Grist, J.; Bennett, D.L.; McMahon, S.B.; Bradbury, E.J. Conduction failure following spinal cord injury: Functional and anatomical changes from acute to chronic stages. J. Neurosci. 2011, 31, 18543–18555. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, L.C.; Matis, A.; Lindau, N.T.; Felder, P.; Gullo, M.; Schwab, M.E. Deep brain stimulation of the midbrain locomotor region improves paretic hindlimb function after spinal cord injury in rats. Sci. Transl. Med. 2013, 5, 208ra146. [Google Scholar] [CrossRef] [PubMed]
- Mushahwar, V.K.; Jacobs, P.L.; Normann, R.A.; Triolo, R.J.; Kleitman, N. New functional electrical stimulation approaches to standing and walking. J. Neural Eng. 2007, 4, S181–S197. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, S.; Giroux, N.; Chau, C.; Marcoux, J.; Brustein, E.; Reader, T.A. Pharmacological aids to locomotor training after spinal injury in the cat. J. Physiol. 2001, 533, 65–74. [Google Scholar] [CrossRef]
- Van den Br, R.; Heutschi, J.; Barraud, Q.; DiGiovanna, J.; Bartholdi, K.; Huerlimann, M.; Friedli, L.; Vollenweider, I.; Moraud, E.M.; Duis, S.; et al. Restoring voluntary control of locomotion after paralyzing spinal cord injury. Science 2012, 336, 1182–1185. [Google Scholar] [CrossRef]
- Radhakrishna, M.; Steuer, I.; Prince, F.; Roberts, M.; Mongeon, D.; Kia, M.; Dyck, S.; Matte, G.; Vaillancourt, M.; Guertin, P. Double-blind, placebo-controlled, randomized phase I/IIa study (safety and efficacy) with buspirone/levodopa/carbidopa (SpinalonTM) in subjects with complete AIS A or motor-complete AIS B spinal cord injury. Curr. Pharm. Des. 2017, 23, 1789–1804. [Google Scholar] [CrossRef] [PubMed]
- Dietz, V.; Muller, R.; Colombo, G. Locomotor activity in spinal man: Significance of afferent input from joint and load receptors. Brain 2002, 125, 2626–2634. [Google Scholar] [CrossRef]
- Grasso, R.; Ivanenko, Y.P.; Zago, M.; Molinari, M.; Scivoletto, G.; Castellano, V.; Macellari, V.; Lacquaniti, F. Distributed plasticity of locomotor pattern generators in spinal cord injured patients. Brain 2004, 127, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Varejao, A.S.; Filipe, V.M. Contribution of cutaneous inputs from the hind paw to the control of locomotion in rats. Behav. Brain Res. 2007, 176, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.S.; Tillakaratne, N.J.; de Leon, R.D. Treadmill training stimulates brain-derived neurotrophic factor mRNA expression in motor neurons of the lumbar spinal cord in spinally transected rats. Neuroscience 2012, 224, 135–144. [Google Scholar] [CrossRef]
- Buck, C.R.; Seburn, K.L.; Cope, T.C. Neurotrophin expression by spinal motoneurons in adult and developing rats. J. Comp. Neurol. 2000, 416, 309–318. [Google Scholar] [CrossRef]
- Macias, M.; Dwornik, A.; Ziemlinska, E.; Fehr, S.; Schachner, M.; Czarkowska-Bauch, J.; Skup, M. Locomotor exercise alters expression of pro-brain-derived neurotrophic factor, brain-derived neurotrophic factor and its receptor TrkB in the spinal cord of adult rats. Eur. J. Neurosci. 2007, 25, 2425–2444. [Google Scholar] [CrossRef] [PubMed]
- Krakowiak, J.; Liu, C.; Papudesu, C.; Ward, P.J.; Wilhelm, J.C.; English, A.W. Neuronal BDNF signaling is necessary for the effects of treadmill exercise on synaptic stripping of axotomized motoneurons. Neural Plast. 2015, 2015, 392591. [Google Scholar] [CrossRef]
- da Costa, R.O.; Gadelha-Filho, C.V.J.; da Costa, A.E.M.; Feitosa, M.L.; de Araújo, D.P.; de Lucena, J.D.; de Aquino, P.E.A.; Lima, F.A.V.; Neves, K.R.T.; de Barros Viana, G.S. The Treadmill Exercise Protects against Dopaminergic Neuron Loss and Brain Oxidative Stress in Parkinsonian Rats. Oxid. Med. Cell. Longev. 2017, 2017, 2138169. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, R.; Theisen, C.C.; Ninan, V.; Twiss, J.L.; Houlé, J.D. Exercise dependent increase in axon regeneration into peripheral nerve grafts by propriospinal but not sensory neurons after spinal cord injury is associated with modulation of regeneration-associated genes. Exp. Neurol. 2016, 276, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.M.; Dong, Z.Q.; Li, Q.; Chen, X. Treadmill training improves neurological deficits and suppresses neuronal apoptosis in cerebral ischemic stroke rats. Neural Regen Res. 2019, 14, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, M.J.; Redmon, N.; Schwartz, G.; English, A.W. Treadmill training promotes axon regeneration in injured peripheral nerves. Exp. Neurol. 2008, 211, 489–493. [Google Scholar] [CrossRef] [PubMed]
- English, A.W.; Wilhelm, J.C.; Sabatier, M.J. Enhancing recovery from peripheral nerve injury using treadmill training. Ann. Anat. 2011, 193, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Yu, X.; Zhu, J.; Li, X.; Zheng, Y.; Xie, Q.; Wu, Q.; Li, S.; Yue, J.; Zhou, Y.; et al. Water Treadmill Training Ameliorates Neurite Outgrowth Inhibition Associated with NGR/RhoA/ROCK by Inhibiting Astrocyte Activation following Spinal Cord Injury. Oxid. Med. Cell. Longev. 2022, 2022, 1724362. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.M.; Gull, K.; Dooley, D. Motor rehabilitation as a therapeutic tool for spinal cord injury: New perspectives in immunomodulation. Cytokine Growth Factor Rev. 2023, 69, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, N.K.; Zhang, Y.P.; Deng, L.; Lu, Q.B.; Shields, C.B.; Walker, M.J.; Li, J.; Xu, X.M. Treadmill training induced lumbar motoneuron dendritic plasticity and behavior recovery in adult rats after a thoracic contusive spinal cord injury. Exp. Neurol. 2015, 271, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Cao, Y.; Dong, C.; Wang, H.; Wang, Q.; Tong, W.; Li, X.; Shan, C.; Wang, T. Neuromuscular interaction is required for neurotrophins-mediated locomotor recovery following treadmill training in rat spinal cord injury. PeerJ 2016, 4, e2025. [Google Scholar] [CrossRef] [PubMed]
- Younsi, A.; Zheng, G.; Scherer, M.; Riemann, L.; Zhang, H.; Tail, M.; Hatami, M.; Skutella, T.; Unterberg, A.; Zweckberger, K. Treadmill training improves survival and differentiation of transplanted neural precursor cells after cervical spinal cord injury. Stem Cell Res. 2020, 45, 101812. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.J.; Li, X.; Li, Y.; Bennett, D.J. 5-HT2 receptor activation facilitates a persistent sodium current and repetitive firing in spinal motoneurons of rats with and without chronic spinal cord injury. J. Neurophysiol. 2006, 96, 1158–1170. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.C.; Nakae, A.; Stephens, M.J.; Rank, M.; D’Amico, J.; Harvey, P.J.; Li, X.; Harris, R.L.; Ballou, E.W.; Anelli, R.; et al. Recovery of motoneuron and locomotor function after spinal cord injury depends on constitutive activity in 5-HT2C receptors. Nat. Med. 2010, 16, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Fouad, K.; Rank, M.M.; Vavrek, R.; Murray, K.C.; Sanelli, L.; Bennett, D.J. Locomotion after spinal cord injury depends on constitutive activity in serotonin receptors. J. Neurophysiol. 2010, 104, 2975–2984. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef]
- Hu, X.; Xu, W.; Ren, Y.; Wang, Z.; He, X.; Huang, R.; Ma, B.; Zhao, J.; Zhu, R.; Cheng, L. Spinal cord injury: Molecular mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Xie, Q.; Li, S.; Yu, X.; Zhou, K.; Yue, J.; Chen, X.; Tu, W.; Yang, G.; Jiang, S. Water treadmill training attenuates blood-spinal cord barrier disruption in rats by promoting angiogenesis and inhibiting matrix metalloproteinase-2/9 expression following spinal cord injury. Fluids Barriers CNS 2020, 17, 70. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, H.; Zhong, J.; Zhan, Z.; Zhao, Q.; Liu, Y.; Li, S.; Wang, H.; Yang, C.; Yu, L.; et al. Osteopontin enhances the effect of treadmill training and promotes functional recovery after spinal cord injury. Mol. Biomed. 2023, 4, 44. [Google Scholar] [CrossRef]
- Chen, K.; Zheng, Y.; Wei, J.A.; Ouyang, H.; Huang, X.; Zhang, F.; Lai, C.S.W.; Ren, C.; So, K.F.; Zhang, L. Exercise training improves motor skill learning via selective activation of mTOR. Sci. Adv. 2019, 5, eaaw1888. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, X.; Zhang, Y.; Chu, G.; Liang, J.; Zhang, B.; Lu, Y.; Steward, O.; Luo, J. Synergistic effect of chemogenetic activation of corticospinal motoneurons and physical exercise in promoting functional recovery after spinal cord injury. Exp. Neurol. 2023, 370, 114549. [Google Scholar] [CrossRef] [PubMed]
- Cantoria, M.J.; See, P.A.; Singh, H.; de Leon, R.D. Adaptations in glutamate and glycine content within the lumbar spinal cord are associated with the generation of novel gait patterns in rats following neonatal spinal cord transection. J. Neurosci. 2011, 31, 18598–18605. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Tashiro, S.; Shinozaki, M.; Hashimoto, S.; Matsumoto, M.; Nakamura, M.; Okano, H.; Nagoshi, N. Treadmill training based on the overload principle promotes locomotor recovery in a mouse model of chronic spinal cord injury. Exp. Neurol. 2021, 345, 113834. [Google Scholar] [CrossRef] [PubMed]
- Minegishi, Y.; Nishimoto, J.; Uto, M.; Ozone, K.; Oka, Y.; Kokubun, T.; Murata, K.; Takemoto, H.; Kanemura, N. Effects of exercise on muscle reinnervation and plasticity of spinal circuits in rat sciatic nerve crush injury models with different numbers of crushes. Muscle Nerve 2022, 65, 612–620. [Google Scholar] [CrossRef] [PubMed]
- He, L.-W.; Guo, X.-J.; Zhao, C.; Rao, J.-S. Rehabilitation Training after Spinal Cord Injury Affects Brain Structure and Function: From Mechanisms to Methods. Biomedicines 2024, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Garraway, S.M.; Huie, J.R. Spinal Plasticity and Behavior: BDNF-Induced Neuromodulation in Uninjured and Injured Spinal Cord. Neural Plast. 2016, 2016, 9857201. [Google Scholar] [CrossRef] [PubMed]
- Tillakaratne, N.J.; De Leon, R.D.; Hoang, T.X.; Roy, R.R.; Edgerton, V.R.; Tobin, A.J. Use-Dependent Modulation of Inhibitory Capacity in the Feline Lumbar Spinal Cord. J. Neurosci. 2002, 22, 3130–3143. [Google Scholar] [CrossRef] [PubMed]
- Khristy, W.; Ali, N.J.; Bravo, A.B.; De Leon, R.; Roy, R.R.; Zhong, H.; London, N.J.; Edgerton, V.R.; Tillakaratne, N.J. Changes in GABA(A) Receptor Subunit Gamma 2 in Extensor and Flexor Motoneurons and Astrocytes after Spinal Cord Transection and Motor Training. Brain Res. 2009, 1273, 9–17. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bilchak, J.N.; Caron, G.; Côté, M.-P. Exercise-Induced Plasticity in Signaling Pathways Involved in Motor Recovery after Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 4858. [Google Scholar] [CrossRef] [PubMed]
- Burda, J.E.; Bernstein, A.M.; Sofroniew, M.V. Astrocyte Roles in Traumatic Brain Injury. Exp. Neurol. 2016, 275, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wu, M.; Peng, C.; Zhao, G.; Gu, R. GFAP Expression in Injured Astrocytes in Rats. Exp. Ther. Med. 2017, 14, 1905–1908. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, C.; Tramontina, A.C.; Nardin, P.; Biasibetti, R.; Costa, A.P.; Vizueti, A.F.; Batassini, C.; Tortorelli, L.S.; Wartchow, K.M.; Dutra, M.F.; et al. Treadmill Exercise Induces Hippocampal Astroglial Alterations in Rats. Neural Plast. 2013, 2013, 709732. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Ushiba, J.; Soekadar, S.R. Neurorehabilitation: Neural Plasticity and Functional Recovery 2018. Neural Plast. 2019, 2019, 7812148. [Google Scholar] [CrossRef] [PubMed]
- Saur, L.; Baptista, P.P.; de Senna, P.N.; Paim, M.F.; do Nascimento, P.; Ilha, J.; Bagatini, P.B.; Achaval, M.; Xavier, L.L. Physical Exercise Increases GFAP Expression and Induces Morphological Changes in Hippocampal Astrocytes. Brain Struct. Funct. 2014, 219, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Gómez-Isla, T.; Growdon, J.H.; Frosch, M.P.; Hyman, B.T. A phenotypic change but not proliferation underlies glial responses in Alzheimer disease. Am. J. Pathol. 2013, 182, 2332–2344. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Gu, Y.; Han, X.; Luan, C.; Liu, C.; Wang, X.; Sun, Y.; Zheng, M.; Fang, M.; Yang, S.; et al. Spatiotemporal Dynamics of the Molecular Expression Pattern and Intercellular Interactions in the Glial Scar Response to Spinal Cord Injury. Neurosci. Bull. 2023, 39, 213–244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Ren, Y.L.; Zhu, Y.J.; Huang, R.Q.; Zhu, R.R.; Cheng, L.M.; Xie, N. The origins and dynamic changes of C3- and S100A10-positive reactive astrocytes after spinal cord injury. Front. Cell. Neurosci. 2023, 17, 1276506. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.R.; Malcangio, M. Astrocytes–multitaskers in chronic pain. Eur. J. Pharmacol. 2013, 716, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Levine, J. The reactions and role of NG2 glia in spinal cord injury. Brain Res. 2016, 1638, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Bai, X.; Stopper, L.; Catalin, B.; Cartarozzi, L.P.; Scheller, A.; Kirchhoff, F. During Development NG2 Glial Cells of the Spinal Cord are Restricted to the Oligodendrocyte Lineage, but Generate Astrocytes upon Acute Injury. Neuroscience 2018, 385, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Komitova, M.; Serwanski, D.R.; Lu, Q.R.; Nishiyama, A. NG2 cells are not a major source of reactive astrocytes after neocortical stab wound injury. Glia 2011, 59, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Hayashibe, M.; Homma, T.; Fujimoto, K.; Oi, T.; Yagi, N.; Kashihara, M.; Nishikawa, N.; Ishizumi, Y.; Abe, S.; Hashimoto, H.; et al. Locomotor improvement of spinal cord-injured rats through treadmill training by forced plantar placement of hind paws. Spinal Cord 2016, 54, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Shulman, I.; Ageeva, T.; Kostennikov, A.; Ogurcov, S.; Tazetdinova, L.; Kabdesh, I.; Rogozhin, A.; Ganiev, I.; Rizvanov, A.; Mukhamedshina, Y. Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs. Int. J. Mol. Sci. 2023, 24, 8240. [Google Scholar] [CrossRef] [PubMed]
- Kostennikov, A.; Kabdesh, I.; Sabirov, D.; Timofeeva, A.; Rogozhin, A.; Shulman, I.; Rizvanov, A.; Mukhamedshina, Y. A Comparative Study of Mesenchymal Stem Cell-Derived Extracellular Vesicles’ Local and Systemic Dose-Dependent Administration in Rat Spinal Cord Injury. Biology 2022, 11, 1853. [Google Scholar] [CrossRef] [PubMed]
- Povysheva, T.; Shmarov, M.; Logunov, D.; Naroditsky, B.; Shulman, I.; Ogurcov, S.; Kolesnikov, P.; Islamov, R.; Chelyshev, Y. Post-spinal cord injury astrocyte-mediated functional recovery in rats after intraspinal injection of the recombinant adenoviral vectors Ad5-VEGF and Ad5-ANG. J. Neurosurg. 2017, 27, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Chelyshev, Y.A.; Muhamedshina, Y.O.; Povysheva, T.V.; Shaymardanova, G.F.; Rizvanov, A.A.; Nigmetzyanova, M.V.; Tiapkina, O.V.; Bondarenko, N.I.; Nikolskiy, E.E.; Islamov, R.R. Characterization of spinal cord glial cells in a model of hindlimb unloading in mice. Neuroscience 2014, 280, 328–339. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host Species | Dilution | Manufacturer |
|---|---|---|---|
| ACAN | Rabbit | 1:200 | Sigma-Aldrich, St. Louis, MO, USA |
| ALDH1L1 | Rabbit | 1:250 | Abcam, Cambridge, UK |
| bTubIII | Mouse | 1:100 | Abcam |
| ChAt | Goat | 1:100 | Abcam |
| CSPG4 | Mouse | 1:100 | Invitrogen, Waltham, MA, USA |
| GAP43 | Rabbit | 1:200 | Santa Cruz Biotechnology, Dallas, TX, USA |
| GFAP | Mouse | 1:200 | Santa Cruz Biotechnology |
| NeuN | Rabbit | 1:100 | Sigma-Aldrich |
| OPN | Rabbit | 1:200 | Cloud-Clone Corp., Houston, TX, USA |
| S100A10 | Rabbit | 1:100 | Abcam |
| Secondary antibodies (anti-rabbit 488, anti-mouse 555, anti-rabbit 647, anti-goat 647, anti-mouse 488) | - | 1:200 | Invitrogen |
| Gene Name Primer | Probe Sequences |
|---|---|
| rGapdh For | ATGACTCTACCCACGGCAAG |
| rGapdh Rev | TGGAGGATGGTGATGGGTTT |
| rGfap For | CCAAAGCCTCAAGGAGGAGA |
| rGfap Rev | CGATGTCCAGGGCTAGCTTA |
| rChat For | GAGCCAATCGCTGGTATGAC |
| rChat Rev | CCCTGACGAGCTTCTTGTTG |
| rSyp For | CTCCTTCCTCCTCTCCCTCT |
| rSyp Rev | AGCCTCCTCCACTCAGTCTA |
| rNg2 For | TCCTGGAGAGAGGTGGAAGA |
| rNg2 Rev | CAAGCCTGTGTTTGTGGTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ageeva, T.; Sabirov, D.; Sufianov, A.; Davletshin, E.; Plotnikova, E.; Shigapova, R.; Sufianova, G.; Timofeeva, A.; Chelyshev, Y.; Rizvanov, A.; et al. The Impact of Treadmill Training on Tissue Integrity, Axon Growth, and Astrocyte Modulation. Int. J. Mol. Sci. 2024, 25, 3772. https://doi.org/10.3390/ijms25073772
Ageeva T, Sabirov D, Sufianov A, Davletshin E, Plotnikova E, Shigapova R, Sufianova G, Timofeeva A, Chelyshev Y, Rizvanov A, et al. The Impact of Treadmill Training on Tissue Integrity, Axon Growth, and Astrocyte Modulation. International Journal of Molecular Sciences. 2024; 25(7):3772. https://doi.org/10.3390/ijms25073772
Chicago/Turabian StyleAgeeva, Tatyana, Davran Sabirov, Albert Sufianov, Eldar Davletshin, Elizaveta Plotnikova, Rezeda Shigapova, Galina Sufianova, Anna Timofeeva, Yuri Chelyshev, Albert Rizvanov, and et al. 2024. "The Impact of Treadmill Training on Tissue Integrity, Axon Growth, and Astrocyte Modulation" International Journal of Molecular Sciences 25, no. 7: 3772. https://doi.org/10.3390/ijms25073772
APA StyleAgeeva, T., Sabirov, D., Sufianov, A., Davletshin, E., Plotnikova, E., Shigapova, R., Sufianova, G., Timofeeva, A., Chelyshev, Y., Rizvanov, A., & Mukhamedshina, Y. (2024). The Impact of Treadmill Training on Tissue Integrity, Axon Growth, and Astrocyte Modulation. International Journal of Molecular Sciences, 25(7), 3772. https://doi.org/10.3390/ijms25073772