
New Strategies for Macrophage Re-Education in Cancer: An Update


Abstract
1. Introduction
2. Recruitment of Monocytes/Macrophages into Tumors

{kind=link}
{kind=link}
| Molecule | Tumor | References |
|---|---|---|
| CSF1/CSFR1 | CC, BC, OSCC | [23,24] |
| CCL2/MCP1 | PC, EC, HCC, BC, PDA | [26,27,28,29,30] |
| CCL20 | BC | [31] |
| CXCL1 | Bladder cancer | [32] |
| CXCL12 | Bladder cancer | [33] |
| CCL5/RANTES | Melanoma | [34] |
| RSK4 | ESCC, OvTs, RCC | [35,36,37] |
3. Role of Monocytes/Macrophages in Cancer Progression and Metastasis
3.1. Crosstalk between Macrophages and CAFs
3.2. Crosstalk between Macrophage and Monocyte/Macrophage
3.3. Crosstalk between TAMs and TME
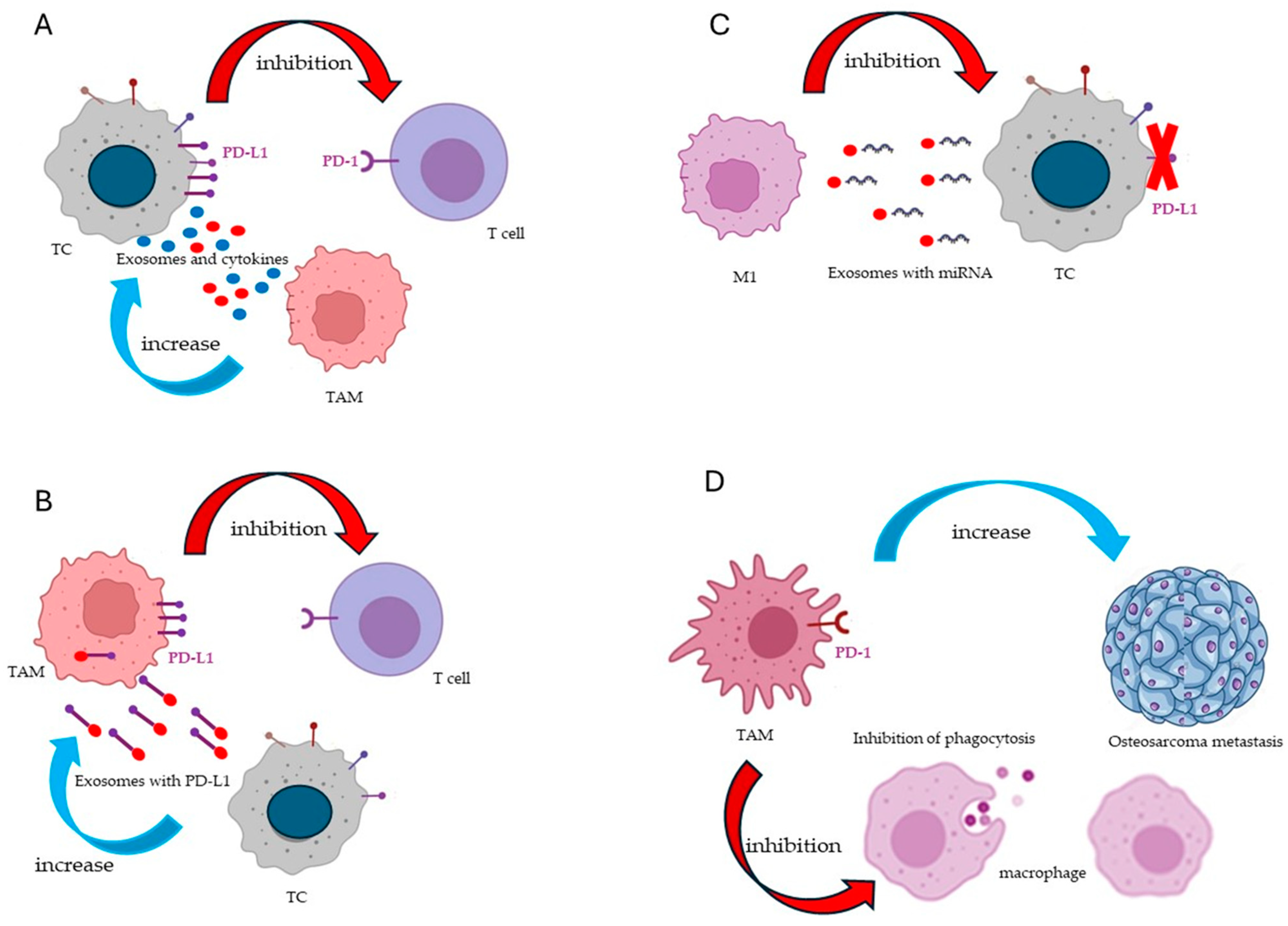
3.4. Crosstalk between TAMs and TCs
4. Re-Education of TAM towards the Anti-Tumor Phenotype
4.1. Immune Checkpoints
4.2. LncRNAs and miRNAs
4.3. JAK/STAT Pathway
4.4. Receptors
4.5. Extracellular Matrix
4.6. Cytokines and Chemokines
5. Artificial Intelligence Can Help Re-Education of TAMs
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue Homeostasis and Inflammation. Annu. Rev. Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and Tumor Progression: Signaling Pathways and Targeted Intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Horn, G.; Moulton, K.; Oza, A.; Byler, S.; Kokolus, S.; Longacre, M. Cancer Development, Progression, and Therapy: An Epigenetic Overview. Int. J. Mol. Sci. 2013, 14, 21087–21113. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as Tools and Targets in Cancer Therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, Z.; Gao, S.; Li, C.; Feng, Y.; Zhou, X. Tumor-Associated Macrophages: Recent Insights and Therapies. Front. Oncol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-Associated Macrophages in Tumor Metastasis: Biological Roles and Clinical Therapeutic Applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Kolesnikoff, N.; Chen, C.H.; Samuel, M.S. Interrelationships between the Extracellular Matrix and the Immune Microenvironment That Govern Epithelial Tumour Progression. Clin. Sci. 2022, 136, 361–377. [Google Scholar] [CrossRef]
- Zhang, Q.; Sioud, M. Tumor-Associated Macrophage Subsets: Shaping Polarization and Targeting. Int. J. Mol. Sci. 2023, 24, 7493. [Google Scholar] [CrossRef] [PubMed]
- Nasir, I.; McGuinness, C.; Poh, A.R.; Ernst, M.; Darcy, P.K.; Britt, K.L. Tumor Macrophage Functional Heterogeneity Can Inform the Development of Novel Cancer Therapies. Trends Immunol. 2023, 44, 971–985. [Google Scholar] [CrossRef]
- Lampiasi, N. Macrophage Polarization: Learning to Manage It 2.0. Int. J. Mol. Sci. 2023, 24, 17409. [Google Scholar] [CrossRef]
- Van Dyken, S.J.; Locksley, R.M. Interleukin-4-and Interleukin-13-Mediated Alternatively Activated Macrophages: Roles in Homeostasis and Disease. Annu. Rev. Immunol. 2013, 31, 317–343. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(Lps+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 451543. [Google Scholar] [CrossRef]
- Wang, L.X.; Zhang, S.X.; Wu, H.J.; Rong, X.L.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Biswas, S.K.; Gangi, L.; Paul, S.; Schioppa, T.; Saccani, A.; Sironi, M.; Bottazzi, B.; Doni, A.; Vincenzo, B.; Pasqualini, F.; et al. A Distinct and Unique Transcriptional Program Expressed by Tumor-Associated Macrophages (Defective NF-B and Enhanced IRF-3/STAT1 Activation). Blood 2006, 107, 2112–2222. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ponce de León, U.; Vázquez-Jiménez, A.; Matadamas-Guzman, M.; Pelayo, R.; Resendis-Antonio, O. Transcriptional and Microenvironmental Landscape of Macrophage Transition in Cancer: A Boolean Analysis. Front. Immunol. 2021, 12, 642842. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-Associated Macrophages as Treatment Targets in Oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Tcyganov, E.; Mastio, J.; Chen, E.; Gabrilovich, D.I. Plasticity of Myeloid-Derived Suppressor Cells in Cancer. Curr. Opin. Immunol. 2018, 51, 76–82. [Google Scholar] [CrossRef]
- Kwak, T.; Wang, F.; Deng, H.; Condamine, T.; Kumar, V.; Perego, M.; Kossenkov, A.; Montaner, L.J.; Xu, X.; Xu, W.; et al. Distinct Populations of Immune-Suppressive Macrophages Differentiate from Monocytic Myeloid-Derived Suppressor Cells in Cancer. Cell Rep. 2020, 33, 108571. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 Recruits Inflammatory Monocytes to Facilitate Breast-Tumour Metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wu, F.; Zhou, Y.; Bao, Z.; Li, H.; Zheng, P.; Zhao, S. Gastric Cancer-Derived Mesenchymal Stromal Cells Trigger M2 Macrophage Polarization That Promotes Metastasis and EMT in Gastric Cancer. Cell Death Dis. 2019, 10, 918. [Google Scholar] [CrossRef]
- Maldonado, M.D.M.; Schlom, J.; Hamilton, D.H. Blockade of Tumor-Derived Colony-Stimulating Factor 1 (CSF1) Promotes an Immune-Permissive Tumor Microenvironment. Cancer Immunol. Immunother. 2023, 72, 3349–3362. [Google Scholar] [CrossRef]
- da Silva, S.D.; Marchi, F.A.; Su, J.; Yang, L.; Valverde, L.; Hier, J.; Bijian, K.; Hier, M.; Mlynarek, A.; Kowalski, L.P.; et al. Co-Overexpression of Twist1-Csf1 Is a Common Event in Metastatic Oral Cancer and Drives Biologically Aggressive Phenotype. Cancers 2021, 13, 153. [Google Scholar] [CrossRef]
- Kazakova, A.; Sudarskikh, T.; Kovalev, O.; Kzhyshkowska, J.; Larionova, I. Interaction of Tumor-associated Macrophages with Stromal and Immune Components in Solid Tumors: Research Progress (Review). Int. J. Oncol. 2023, 62, 32. [Google Scholar] [CrossRef]
- Li, X.; He, G.; Liu, J.; Yan, M.; Shen, M.; Xu, L.; An, M.; Huang, J.; Gao, Z. CCL2-Mediated Monocytes Regulate Immune Checkpoint Blockade Resistance in Pancreatic Cancer. Int. Immunopharmacol. 2022, 106, 108598. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Q.; Xu, M.; Wang, L.; Chen, X.; Feng, Y.; Li, Y.; Zhang, X.; Cui, W.; Jia, X. CCL2-CCR2 Axis Recruits Tumor Associated Macrophages to Induce Immune Evasion through PD-1 Signaling in Esophageal Carcinogenesis. Mol. Cancer 2020, 19, 41. [Google Scholar] [CrossRef]
- Li, X.; Yao, W.; Yan, Y.; Chen, P.; Li, B.; Li, J. Targeting of Tumor Infiltrating Macrophages via CCL2/CCR2 Signaling as a Therapeutic Strategy against Hepatocellular Carcinoma. Gut 2017, 66, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.L.; Angela Murphy, E. Importance of Chemokine (CC-Motif) Ligand 2 in Breast Cancer. Int. J. Biol. Markers 2012, 27, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Li, C.; Wang, Y.; Matsukawa, A. The Chemokine Monocyte Chemoattractant Protein-1/CCL2 Is a Promoter of Breast Cancer Metastasis. Cell. Mol. Immunol. 2023, 20, 714–738. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Zhang, N.; Mo, D.; Qin, T. CCL20 Is a Potential Therapeutic Target Associated with Immune Infiltration in Breast Cancer. J. Int. Med. Res. 2023, 51, 3000605231171762. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Hori, S.; Morizawa, Y.; Tatsumi, Y.; Nakai, Y.; Anai, S.; Torimoto, K.; Aoki, K.; Tanaka, N.; Shimada, K.; et al. CXCL1-Mediated Interaction of Cancer Cells with Tumor-Associated Macrophages and Cancer-Associated Fibroblasts Promotes Tumor Progression in Human Bladder Cancer. Neoplasia 2016, 18, 636–646. [Google Scholar] [CrossRef]
- Lu, G.; Qiu, Y. SPI1-Mediated CXCL12 Expression in Bladder Cancer Affects the Recruitment of Tumor-Associated Macrophages. Mol. Carcinog. 2023, 63, 448–460. [Google Scholar] [CrossRef]
- Lucianò, A.M.; Di Martile, M.; Pérez-Oliva, A.B.; Di Caprio, M.; Foddai, M.L.; Buglioni, S.; Mulero, V.; Del Bufalo, D. Exploring Association of Melanoma-Specific Bcl-XL with Tumor Immune Microenvironment. J. Exp. Clin. Cancer Res. 2023, 42, 178. [Google Scholar] [CrossRef]
- He, S.; Lu, M.; Zhang, L.; Wang, Z. RSK4 Promotes the Macrophage Recruitment and M2 Polarization in Esophageal Squamous Cell Carcinoma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2023, 1870, 166996. [Google Scholar] [CrossRef]
- Nagumo, Y.; Villareal, M.O.; Isoda, H.; Usui, T. RSK4 Confers Paclitaxel Resistance to Ovarian Cancer Cells, Which Is Resensitized by Its Inhibitor BI-D1870. Biochem. Biophys. Res. Commun. 2023, 679, 23–30. [Google Scholar] [CrossRef]
- Ma, J.; Wang, K.; Chai, J.; Xu, T.; Wei, J.; Liu, Y.; Wang, Y.; Xu, J.; Li, M.; Fan, L. High RSK4 Expression Constitutes a Predictor of Poor Prognosis for Patients with Clear Cell Renal Carcinoma. Pathol. Res. Pract. 2021, 227, 153642. [Google Scholar] [CrossRef]
- Dric Blanpain, C.; Migeotte, I.; Lee, B.; Vakili, J.; Doranz, B.J.; Dric Govaerts, C.; Vassart, G.; Doms, R.W.; Parmentier, M. CCR5 Binds Multiple CC-Chemokines: MCP-3 Acts as a Natural Antagonist. Blood J. Am. Soc. Hematol. 1999, 94, 1899–1905. [Google Scholar]
- Parody, T.R.; Stone, M.J. High Level Expression, Activation, and Antagonism of CC Chemokine Receptors CCR2 and CCR3 in Chinese Hamster Ovary Cells. Cytokine 2004, 27, 38–46. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interferon Cytokine Res. 2009, 29, 313–325. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior Beyond Chemotaxis. Front. Immunol. 2019, 10, 491100. [Google Scholar] [CrossRef]
- Fei, L.; Ren, X.; Yu, H.; Zhan, Y. Targeting the CCL2/CCR2 Axis in Cancer Immunotherapy: One Stone, Three Birds? Front. Immunol. 2021, 12, 771210. [Google Scholar] [CrossRef]
- Franklin, R.A.; Liao, W.; Sarkar, A.; Kim, M.V.; Bivona, M.R.; Liu, K.; Pamer, E.G.; Li, M.O. The Cellular and Molecular Origin of Tumor-Associated Macrophages. Science 2014, 344, 921–925. [Google Scholar] [CrossRef]
- Shand, F.H.W.; Ueha, S.; Otsuji, M.; Koid, S.S.; Shichino, S.; Tsukui, T.; Kosugi-Kanaya, M.; Abe, J.; Tomura, M.; Ziogas, J.; et al. Tracking of Intertissue Migration Reveals the Origins of Tumor-Infiltrating Monocytes. Proc. Natl. Acad. Sci. USA 2014, 111, 7771–7776. [Google Scholar] [CrossRef]
- Yoshie, O.; Matsushima, K. CCR4 and Its Ligands: From Bench to Bedside. Int. Immunol. 2015, 27, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 438073. [Google Scholar]
- Sung, G.H.; Chang, H.; Lee, J.Y.; Song, S.Y.; Kim, H.S. Pancreatic-Cancer-Cell-Derived Trefoil Factor 2 Impairs Maturation and Migration of Human Monocyte-Derived Dendritic Cells In Vitro. Anim. Cells Syst. 2018, 22, 368–381. [Google Scholar] [CrossRef]
- Khabipov, A.; Trung, D.N.; van der Linde, J.; Miebach, L.; Lenz, M.; Erne, F.; von Bernstorff, W.; Schulze, T.; Kersting, S.; Bekeschus, S.; et al. CCR4 Blockade Diminishes Intratumoral Macrophage Recruitment and Augments Survival of Syngeneic Pancreatic Cancer-Bearing Mice. Biomedicines 2023, 11, 1517. [Google Scholar] [CrossRef] [PubMed]
- Storz, P. Roles of Differently Polarized Macrophages in the Initiation and Progressionof Pancreatic Cancer. Front. Immunol. 2023, 14, 1237711. [Google Scholar] [CrossRef]
- Zaynagetdinov, R.; Sherrill, T.P.; Polosukhin, V.V.; Han, W.; Ausborn, J.A.; McLoed, A.G.; McMahon, F.B.; Gleaves, L.A.; Degryse, A.L.; Stathopoulos, G.T.; et al. A Critical Role for Macrophages in Promotion of Urethane-Induced Lung Carcinogenesis. J. Immunol. 2011, 187, 5703–5711. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Z.; Bertran, T.; Finetti, P.; Lohmann, E.; Mamessier, E.; Bidaut, G.; Bertucci, F.; Rego, M.; Tomasini, R. Macrophages Reprogramming Driven by Cancer-Associated Fibroblasts under FOLFIRINOX Treatment Correlates with Shorter Survival in Pancreatic Cancer. Cell Commun. Signal. 2024, 22, 1. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, R.; Chatterjee, S.; Hembram, K.C.; Sethy, C.; Mandal, M.; Kundu, C.N. Nano Formulated Resveratrol Inhibits Metastasis and Angiogenesis by Reducing Inflammatory Cytokines in Oral Cancer Cells by Targeting Tumor Associated Macrophages. J. Nutr. Biochem. 2021, 92, 108624. [Google Scholar] [CrossRef]
- Pradhan, R.; Paul, S.; Acharya, S.S.; Sinha, S.; Dash, S.R.; Kundu, C.N. Nano Formulated Resveratrol Inhibits PD-L1 in Oral Cancer Cells by Deregulating the Association between Tumor Associated Macrophages and Cancer Associated Fibroblasts through IL-6/JAK2/STAT3 Signaling Axis. J. Nutr. Biochem. 2024, 125, 109568. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, L.; Zhao, L.; Shao, S.; Ning, Q.; Jing, X.; Zhang, Y.; Zhao, F.; Liu, X.; Gu, S.; et al. VEGFA/NRP-1/GAPVD1 Axis Promotes Progression and Cancer Stemness of Triple-Negative Breast Cancer by Enhancing Tumor Cell-Macrophage Crosstalk. Int. J. Biol. Sci. 2024, 20, 446–463. [Google Scholar] [CrossRef]
- Zhang, Q.; Tsui, Y.M.; Zhang, V.X.; Lu, A.J.; Lee, J.M.F.; Lee, E.; Cheung, G.C.H.; Li, P.M.; Cheung, E.T.Y.; Chia, N.H.; et al. Reciprocal Interactions between Malignant Cells and Macrophages Enhance Cancer Stemness and M2 Polarization in HBV-Associated Hepatocellular Carcinoma. Theranostics 2024, 14, 892–910. [Google Scholar] [CrossRef]
- Shang, C.; Sun, Y.; Wang, Y.; Shi, H.; Han, X.; Mo, Y.; Wang, D.; Ke, Y.; Zeng, X. CXCL10 Conditions Alveolar Macrophages within the Premetastatic Niche to Promote Metastasis. Cancer Lett. 2022, 537, 215667. [Google Scholar] [CrossRef]
- Cao, X.; Song, Y.; Wu, H.; Ren, X.; Sun, Q.; Liang, Z. C-X-C Motif Chemokine Ligand 9 Correlates with Favorable Prognosis in Triple-Negative Breast Cancer by Promoting Immune Cells Infiltration. Mol. Cancer Ther. 2023, 22, 1493–1502. [Google Scholar] [CrossRef]
- Bill, R.; Wirapati, P.; Messemaker, M.; Roh, W.; Zitti, B.; Duval, F.; Kiss, M.; Park, J.C.; Saal, T.M.; Hoelzl, J.; et al. CXCL9:SPP1 Macrophage Polarity Identifies a Network of Cellular Programs That Control Human Cancers. Science 2023, 381, 515–524. [Google Scholar] [CrossRef]
- Liu, K.; Wang, H.; Zhou, J.; Zhu, S.; Ma, M.; Xiao, H.; Ding, Y. HMGB1 in Exosomes Derived from Gastric Cancer Cells Induces M2-like Macrophage Polarization by Inhibiting the NF-ΚB Signaling Pathway. Cell Biol. Int. 2023, 48, 334–346. [Google Scholar] [CrossRef]
- Yin, C.; Li, J.; Li, S.; Yang, X.; Lu, Y.; Wang, C.; Liu, B. LncRNA-HOXC-AS2 Regulates Tumor-Associated Macrophage Polarization through the STAT1/SOCS1 and STAT1/CIITA Pathways to Promote the Progression of Non-Small Cell Lung Cancer. Cell. Signal. 2024, 115, 111031. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Zhang, X.; Xu, W. The Promising Role of Tumor-Associated Macrophages in the Treatment of Cancer. Drug Resist. Updates 2024, 73, 101041. [Google Scholar] [CrossRef]
- Lin, B.; Zhang, T.; Ye, X.; Yang, H. High Expression of EMP1 Predicts a Poor Prognosis and Correlates with Immune Infiltrates in Bladder Urothelial Carcinoma. Oncol. Lett. 2020, 20, 2840–2854. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Wang, H.; Ma, J.; Wang, Y.; Xu, W. Single-Cell Analyses EMP1 as a Marker of the Ratio of M1/M2 Macrophages Is Associated with EMT, Immune Infiltration, and Prognosis in Bladder Cancer. Bladder 2023, 10, e21200011. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In Vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Cao, J.; Dong, R.; Jiang, L.; Gong, Y.; Yuan, M.; You, J.; Meng, W.; Chen, Z.; Zhang, N.; Weng, Q.; et al. LncRNA-Mm2p Identified as a Modulator of Macrophage M2 Polarization. Cancer Immunol. Res. 2019, 7, 292–305. [Google Scholar] [CrossRef]
- Yang, X.; Luo, Y.; Li, M.; Jin, Z.; Chen, G.; Gan, C. Long Non-Coding RNA NBR2 Suppresses the Progression of Colorectal Cancer by Downregulating MiR-19a to Regulate M2 Macrophage Polarization. Chin. J. Physiol. 2023, 66, 546. [Google Scholar] [CrossRef]
- Yang, J.; Liu, F.; Wang, Y.; Qu, L.; Lin, A. LncRNAs in Tumor Metabolic Reprogramming and Immune Microenvironment Remodeling. Cancer Lett. 2022, 543, 215798. [Google Scholar] [CrossRef]
- Dong, N.; Guo, J.; Han, S.; Bao, L.; Diao, Y.; Lin, Z. Positive Feedback Loop of LncRNA HOXC-AS2/ MiR-876-5p/ZEB1 to Regulate EMT in Glioma. OncoTargets Ther. 2019, 12, 7601–7609. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Ji, X.; Bu, Z.; Zhang, J.; Wu, X.; Zong, X.; Fan, B.; Jia, Z.; Ji, J. Identification of Key Long Non Coding RNAs in Gastric Adnocarcinoma. Cancer Biomark. 2020, 27, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, P.; Zhou, G. Long Noncoding RNAs in the Prediction of Survival of Patients with Digestive Cancers. Turk. J. Gastroenterol. 2023, 34, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-Associated Macrophages Are a Distinct M2 Polarised Population Promoting Tumour Progression: Potential Targets of Anti-Cancer Therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef]
- Pittet, M.J.; Michielin, O.; Migliorini, D. Clinical Relevance of Tumor-Associated Macrophages. Nat. Rev. Clin. Oncol. 2022, 19, 402–421. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, Y.; Wang, M.; Liu, K.; Hoover, A.R.; Li, M.; Towner, R.A.; Mukherjee, P.; Zhou, F.; Qu, J.; et al. Synergistic Interventional Photothermal Therapy and Immunotherapy Using an Iron Oxide Nanoplatform for the Treatment of Pancreatic Cancer. Acta Biomater. 2022, 138, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, J.; Luo, Z.; Shi, Q.; Liu, G.; Wu, F.; Wang, Z.; Huang, Y.; Zhou, D. Engineering Endogenous Tumor-Associated Macrophage-Targeted Biomimetic Nano-RBC to Reprogram Tumor Immunosuppressive Microenvironment for Enhanced Chemo-Immunotherapy. Adv. Mater. 2021, 33, 2103497. [Google Scholar] [CrossRef]
- Li, M.; He, L.; Zhu, J.; Zhang, P.; Liang, S. Targeting Tumor-Associated Macrophages for Cancer Treatment. Cell Biosci. 2022, 12, 85. [Google Scholar] [CrossRef]
- Pathria, P.; Louis, T.L.; Varner, J.A. Targeting Tumor-Associated Macrophages in Cancer. Trends Immunol. 2019, 40, 310–327. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Li, L.; He, D.; Chi, J.; Li, Q.; Wu, Y.; Zhao, Y.; Zhang, S.; Wang, L.; et al. Engineered Extracellular Vesicles and Their Mimetics for Cancer Immunotherapy. J. Control. Release 2022, 349, 679–698. [Google Scholar] [CrossRef]
- Nocini, R.; Vianini, M.; Girolami, I.; Calabrese, L.; Scarpa, A.; Martini, M.; Morbini, P.; Marletta, S.; Brunelli, M.; Molteni, G.; et al. PD-L1 in Oral Squamous Cell Carcinoma: A Key Biomarker from the Laboratory to the Bedside. Clin. Exp. Dent. Res. 2022, 8, 690–698. [Google Scholar] [CrossRef]
- Pu, Y.; Ji, Q. Tumor-Associated Macrophages Regulate PD-1/PD-L1 Immunosuppression. Front. Immunol. 2022, 13, 874589. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Cheng, D.; Ma, D.; Chen, R.; Li, D.; Zhao, W.; Fang, C.; Ji, M. Mutual Regulation of PD-L1 Immunosuppression between Tumor-Associated Macrophages and Tumor Cells: A Critical Role for Exosomes. Cell Commun. Signal. 2024, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Tao, L.; Yu, Y.; Wang, Q.; Ye, P.; Sun, Y.; Zhou, J. Oral Squamous Cell Carcinoma Cell-Derived GM-CSF Regulates PD-L1 Expression in Tumor-Associated Macrophages through the JAK2/STAT3 Signaling Pathway. Am. J. Cancer Res. 2023, 13, 589. [Google Scholar]
- Zheng, C.; Li, H.; Zhao, X.; Yang, S.; Zhan, J.; Liu, H.; Jiang, Y.; Shi, L.; Song, Y.; Lei, Y.; et al. Expression of PD-1 Mitigates Phagocytic Activities TAM in Osteosarcoma. Heliyon 2024, 10, e23498. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gong, L.; Cao, Y.; Liu, Z.; Wang, Y.; Cheng, H.; Feng, Y.; Yao, S.; Yin, Y.; Wu, Z.; et al. Reprogramming Tumor-Associated Macrophages by a Dually Targeted Milk Exosome System as a Potent Monotherapy for Cancer. J. Control. Release 2024, 366, 395–409. [Google Scholar] [CrossRef]
- Arora, L.; Patra, D.; Roy, S.; Nanda, S.; Singh, N.; Verma, A.K.; Chakraborti, A.; Dasgupta, S.; Pal, D. Hypoxia-Induced MiR-210-3p Expression in Lung Adenocarcinoma Potentiates Tumor Development by Regulating CCL2-Mediated Monocyte Infiltration. Mol. Oncol. 2023. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Fang, P.; Li, W.J.; Zhang, J.; Wang, G.P.; Jiang, D.F.; Chen, F.P. LncRNA NEAT1 Sponges MiR-214 to Regulate M2 Macrophage Polarization by Regulation of B7-H3 in Multiple Myeloma. Mol. Immunol. 2020, 117, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, S.; Liu, Y.; Han, H.; Gong, M.; Song, Y. The Role of Exosomal MiR-181b in the Crosstalk between NSCLC Cells and Tumor-Associated Macrophages. Genes Genom. 2022, 44, 1243–1258. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Yu, G. Hypoxic Lung Cancer Cell-Derived Exosomal MiR-21 Mediates Macrophage M2 Polarization and Promotes Cancer Cell Proliferation through Targeting IRF1. World J. Surg. Oncol. 2022, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, M.B.; Tang, W.S.; Wang, D.; Kaniowski, D.; Kang, E.; Dizman, N.; Chehrazi-Raffle, A.; Meza, L.; Zengin, Z.; Hall, J.; et al. Targeting STAT3 in Tumor-Associated Antigen-Presenting Cells as a Strategy for Kidney and Bladder Cancer Immunotherapy. Front. Immunol. 2023, 14, 1274781. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yu, H.; Bai, M.; Lu, S.; Wang, C.; Ke, S.; Huang, J.; Li, Z.; Xu, Y.; Yin, B.; et al. IRG1 Restrains M2 Macrophage Polarization and Suppresses Intrahepatic Cholangiocarcinoma Progression via the CCL18/STAT3 Pathway. Cancer Sci. 2024, 115, 777–790. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, D.; Su, N.; Lou, W.; Chen, Y.; Yang, H.; Chen, C.; Xi, F.; Chen, Y.; Deng, L.; et al. TRIM65 Knockout Inhibits the Development of HCC by Polarization Tumor-Associated Macrophages towards M1 Phenotype via JAK1/STAT1 Signaling Pathway. Int. Immunopharmacol. 2024, 128, 111494. [Google Scholar] [CrossRef]
- Sallam, M.A.; Wyatt Shields, I.V.C.; Prakash, S.; Kim, J.; Pan, D.C.; Mitragotri, S. A Dual Macrophage Polarizer Conjugate for Synergistic Melanoma Therapy. J. Control. Release 2021, 335, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Jiang, X.; Sun, T.; Zeng, F.; Huang, G.; Liang, C.; Nie, L. In Vivo Visualization of Tumor-Associated Macrophages Re-Education by Photoacoustic/Fluorescence Dual-Modal Imaging with a Metal-Organic Frames-Based Caspase-1 Nanoreporter. J. Colloid Interface Sci. 2024, 659, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Anfray, C.; Varela, C.F.; Ummarino, A.; Maeda, A.; Sironi, M.; Gandoy, S.; Brea, J.; Loza, M.I.; León, S.; Calvo, A.; et al. Polymeric Nanocapsules Loaded with Poly(I:C) and Resiquimod to Reprogram Tumor-Associated Macrophages for the Treatment of Solid Tumors. Front. Immunol. 2023, 14, 1334800. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, C.; Wang, Y.; Hao, J.; Liu, Y. GSH/PH Dual Responsive Chitosan Nanoparticles for Reprogramming M2 Macrophages and Overcoming Cancer Chemoresistance. Biomater. Sci. 2023, 12, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhou, G.; Shi, Z.; Yu, L.; Zhou, X. TREM1 Facilitates the Development of Gastric Cancer through Regulating Neutrophil Extracellular Traps-Mediated Macrophage Polarization. Dig. Liver Dis. 2023, in press. [CrossRef]
- Kim, J.; Kim, M.; Yong, S.B.; Han, H.; Kang, S.; Lahiji, S.F.; Kim, S.; Hong, J.; Seo, Y.; Kim, Y.H. Engineering TGF-β Inhibitor-Encapsulated Macrophage-Inspired Multi-Functional Nanoparticles for Combination Cancer Immunotherapy. Biomater. Res. 2023, 27, 136. [Google Scholar] [CrossRef]
- Huang, C.; Ou, R.; Chen, X.; Zhang, Y.; Li, J.; Liang, Y.; Zhu, X.; Liu, L.; Li, M.; Lin, D.; et al. Tumor Cell-Derived SPON2 Promotes M2-Polarized Tumor-Associated Macrophage Infiltration and Cancer Progression by Activating PYK2 in CRC. J. Exp. Clin. Cancer Res. 2021, 40, 304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Li, Q.; Yang, X.M.; Fang, F.; Li, J.; Wang, Y.H.; Yang, Q.; Zhu, L.; Nie, H.Z.; Zhang, X.L.; et al. Spon2 Promotes M1-like Macrophage Recruitment and Inhibits Hepatocellular Carcinoma Metastasis by Distinct Integrin–Rho Gtpase–Hippo Pathways. Cancer Res. 2018, 78, 2305–2317. [Google Scholar] [CrossRef]
- Ihle, C.L.; Straign, D.M.; Canari, J.A.; Torkko, K.C.; Zolman, K.L.; Smith, E.E.; Owens, P. Unique Macrophage Phenotypes Activated by BMP Signaling in Breast Cancer Bone Metastases. JCI Insight 2024, 9, e168517. [Google Scholar] [CrossRef]
- Mazzoni, M.; Mauro, G.; Erreni, M.; Romeo, P.; Minna, E.; Vizioli, M.G.; Belgiovine, C.; Rizzetti, M.G.; Pagliardini, S.; Avigni, R.; et al. Senescent Thyrocytes and Thyroid Tumor Cells Induce M2-like Macrophage Polarization of Human Monocytes via a PGE2-Dependent Mechanism. J. Exp. Clin. Cancer Res. 2019, 38, 208. [Google Scholar] [CrossRef]
- Mazzoni, M.; Mauro, G.; Minoli, L.; Cleris, L.; Anania, M.C.; Di Marco, T.; Minna, E.; Pagliardini, S.; Rizzetti, M.G.; Manenti, G.; et al. Senescent Thyrocytes, Similarly to Thyroid Tumor Cells, Elicit M2-like Macrophage Polarization In Vivo. Biology 2021, 10, 985. [Google Scholar] [CrossRef]
- Hu, Y.; Nie, W.; Lyu, L.; Zhang, X.; Wang, W.; Zhang, Y.; He, S.; Guo, A.; Liu, F.; Wang, B.; et al. Tumor-Microenvironment-Activatable Nanoparticle Mediating Immunogene Therapy and M2 Macrophage-Targeted Inhibitor for Synergistic Cancer Immunotherapy. ACS Nano 2023, 18, 3295–3312. [Google Scholar] [CrossRef]
- Chen, S.; Morine, Y.; Tokuda, K.; Yamada, S.; Saito, Y.; Nishi, M.; Ikemoto, T.; Shimada, M. Cancer-Associated Fibroblast-Induced M2-Polarized Macrophages Promote Hepatocellular Carcinoma Progression via the Plasminogen Activator Inhibitor-1 Pathway. Int. J. Oncol. 2021, 59, 59. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Suo, B.; Long, G.; Gao, Y.; Song, J.; Zhang, M.; Feng, B.; Shang, C.; Wang, D. Exosomal MiRNA-16-5p Derived From M1 Macrophages Enhances T Cell-Dependent Immune Response by Regulating PD-L1 in Gastric Cancer. Front. Cell Dev. Biol. 2020, 8, 572689. [Google Scholar] [CrossRef] [PubMed]
- Binnemars-Postma, K.; Bansal, R.; Storm, G.; Prakash, J. Targeting the Stat6 Pathway in Tumor-Associated Macrophages Reduces Tumor Growth and Metastatic Niche Formation in Breast Cancer. FASEB J. 2018, 32, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Iriki, T.; Ohnishi, K.; Fujiwara, Y.; Horlad, H.; Saito, Y.; Pan, C.; Ikeda, K.; Mori, T.; Suzuki, M.; Ichiyasu, H.; et al. The Cell-Cell Interaction between Tumor-Associated Macrophages and Small Cell Lung Cancer Cells Is Involved in Tumor Progression via STAT3 Activation. Lung Cancer 2017, 106, 22–32. [Google Scholar] [CrossRef]
- Jill, N.; Bhootra, S.; Kannanthodi, S.; Shanmugam, G.; Rakshit, S.; Rajak, R.; Thakkar, V.; Sarkar, K. Interplay between Signal Transducers and Activators of Transcription (STAT) Proteins and Cancer: Involvement, Therapeutic and Prognostic Perspective. Clin. Exp. Med. 2023, 23, 4323–4339. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhang, M.F.; Tian, Q.H.; Zhang, C.Z. TRIM65 Triggers β-Catenin Signaling via Ubiquitylation of Axin1 to Promote Hepatocellular Carcinoma. J. Cell Sci. 2017, 130, 3108–3115. [Google Scholar] [CrossRef]
- Li, Y.; Ma, C.; Zhou, T.; Liu, Y.; Sun, L.; Yu, Z. TRIM65 Negatively Regulates P53 through Ubiquitination. Biochem. Biophys. Res. Commun. 2016, 473, 278–282. [Google Scholar] [CrossRef]
- Tang, T.; Li, P.; Zhou, X.; Wang, R.; Fan, X.; Yang, M.; Qi, K. The E3 Ubiquitin Ligase TRIM65 Negatively Regulates Inflammasome Activation Through Promoting Ubiquitination of NLRP3. Front. Immunol. 2021, 12, 741839. [Google Scholar] [CrossRef]
- Rodell, C.B.; Arlauckas, S.P.; Cuccarese, M.F.; Garris, C.S.; Li, R.; Ahmed, M.S.; Kohler, R.H.; Pittet, M.J.; Weissleder, R. TLR7/8-Agonist-Loaded Nanoparticles Promote the Polarization of Tumour-Associated Macrophages to Enhance Cancer Immunotherapy. Nat. Biomed. Eng. 2018, 2, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Kakwere, H.; Zhang, H.; Ingham, E.S.; Nura-Raie, M.; Tumbale, S.K.; Allen, R.; Tam, S.M.; Wu, B.; Liu, C.; Kheirolomoom, A.; et al. Systemic Immunotherapy with Micellar Resiquimod–Polymer Conjugates Triggers a Robust Antitumor Response in a Breast Cancer Model. Adv. Healthc. Mater. 2021, 10, e2100008. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Liu, H.; Xue, Y.; Lin, J.; Fu, Y.; Xia, Z.; Pan, D.; Zhang, J.; Qiao, K.; Zhang, Z.; et al. Reversing Cold Tumors to Hot: An Immunoadjuvant-Functionalized Metal-Organic Framework for Multimodal Imaging-Guided Synergistic Photo-Immunotherapy. Bioact. Mater. 2021, 6, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Raggi, F.; Bosco, M.C. Targeting Mononuclear Phagocyte Receptors in Cancer Immunotherapy: New Perspectives of the Triggering Receptor Expressed on Myeloid Cells (Trem-1). Cancers 2020, 12, 1337. [Google Scholar] [CrossRef] [PubMed]
- Cescon, M.; Rampazzo, E.; Bresolin, S.; Da Ros, F.; Manfreda, L.; Cani, A.; Della Puppa, A.; Braghetta, P.; Bonaldo, P.; Persano, L. Collagen VI Sustains Cell Stemness and Chemotherapy Resistance in Glioblastoma. Cell. Mol. Life Sci. 2023, 80, 233. [Google Scholar] [CrossRef] [PubMed]
- Patras, L.; Paul, D.; Matei, I.R. Weaving the Nest: Extracellular Matrix Roles in Pre-Metastatic Niche Formation. Front. Oncol. 2023, 13, 1163786. [Google Scholar] [CrossRef]
- Lu, H.; Feng, Y.; Hu, Y.; Guo, Y.; Liu, Y.; Mao, Q.; Xue, W. Spondin 2 Promotes the Proliferation, Migration and Invasion of Gastric Cancer Cells. J. Cell. Mol. Med. 2020, 24, 98–113. [Google Scholar] [CrossRef]
- Tang, J.; Huang, Q.; Li, X.; Gu, S. Comprehensive Analysis of the Oncogenic and Immunological Role of SPON2 in Human Tumors. Medicine 2023, 102, E35122. [Google Scholar] [CrossRef]
- Owens, P.; Pickup, M.W.; Novitskiy, S.V.; Giltnane, J.M.; Gorska, A.E.; Hopkins, C.R.; Hong, C.C.; Moses, H.L. Inhibition of BMP Signaling Suppresses Metastasis in Mammary Cancer. Oncogene 2015, 34, 2437–2449. [Google Scholar] [CrossRef]
- Sawa-Wejksza, K.; Dudek, A.; Lemieszek, M.; Kaławaj, K.; Kandefer-Szerszeń, M. Colon Cancer–Derived Conditioned Medium Induces Differentiation of THP-1 Monocytes into a Mixed Population of M1/M2 Cells. Tumor Biol. 2018, 40, 1010428318797880. [Google Scholar] [CrossRef]
- Khabipov, A.; Käding, A.; Liedtke, K.R.; Freund, E.; Partecke, L.I.; Bekeschus, S. RAW 264.7 Macrophage Polarization by Pancreatic Cancer Cells—A Model for Studying Tumour-Promoting Macrophages. Anticancer Res. 2019, 39, 2871–2882. [Google Scholar] [CrossRef]
- Li, P.; Ma, C.; Li, J.; You, S.; Dang, L.; Wu, J.; Hao, Z.; Li, J.; Zhi, Y.; Chen, L.; et al. Proteomic Characterization of Four Subtypes of M2 Macrophages Derived from Human THP-1 Cells. J. Zhejiang Univ. Sci. B 2022, 23, 407–422. [Google Scholar] [CrossRef]
- Pe, K.C.S.; Saetung, R.; Yodsurang, V.; Chaotham, C.; Suppipat, K.; Chanvorachote, P.; Tawinwung, S. Triple-Negative Breast Cancer Influences a Mixed M1/M2 Macrophage Phenotype Associated with Tumor Aggressiveness. PLoS ONE 2022, 17, e0273044. [Google Scholar] [CrossRef] [PubMed]
- de León, U.A.P.; Vázquez-Jiménez, A.; Matadamas-Guzmán, M.; Resendis-Antonio, O. Boolean Modeling Reveals That Cyclic Attractors in Macrophage Polarization Serve as Reservoirs of States to Balance External Perturbations from the Tumor Microenvironment. Front. Immunol. 2022, 13, 1012730. [Google Scholar] [CrossRef] [PubMed]
- Selig, M.; Poehlman, L.; Lang, N.C.; Völker, M.; Rolauffs, B.; Hart, M.L. Prediction of Six Macrophage Phenotypes and Their IL-10 Content Based on Single-Cell Morphology Using Artificial Intelligence. Front. Immunol. 2023, 14, 1336393. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.; Jarrah, A.S.; Tieri, P.; Cesareni, G.; Castiglione, F. Gene Regulatory Network Modeling of Macrophage Differentiation Corroborates the Continuum Hypothesis of Polarization States. Front. Physiol. 2018, 9, 1659. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.A.; Byrne, H.M. Quantification of Spatial and Phenotypic Heterogeneity in an Agent-Based Model of Tumour-Macrophage Interactions. PLoS Comput. Biol. 2023, 19, e1010994. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Medeiros, T.X.; Sové, R.J.; Annex, B.H.; Popel, A.S. A Data-Driven Computational Model Enables Integrative and Mechanistic Characterization of Dynamic Macrophage Polarization. iScience 2021, 24, 102112. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, C.; Santa-Maria, C.A.; Emens, L.A.; Popel, A.S. Dynamics of Tumor-Associated Macrophages in a Quantitative Systems Pharmacology Model of Immunotherapy in Triple-Negative Breast Cancer. iScience 2022, 25, 104702. [Google Scholar] [CrossRef]
- Ruiz-Martinez, A.; Gong, C.; Wang, H.; Sove, R.J.; Mi, H.; Kimko, H.; Popel, A.S. Simulations of Tumor Growth and Response to Immunotherapy by Coupling a Spatial Agent-Based Model with a Whole-Patient Quantitative Systems Pharmacology Model. PLoS Comput. Biol. 2022, 18, e1010254. [Google Scholar] [CrossRef]
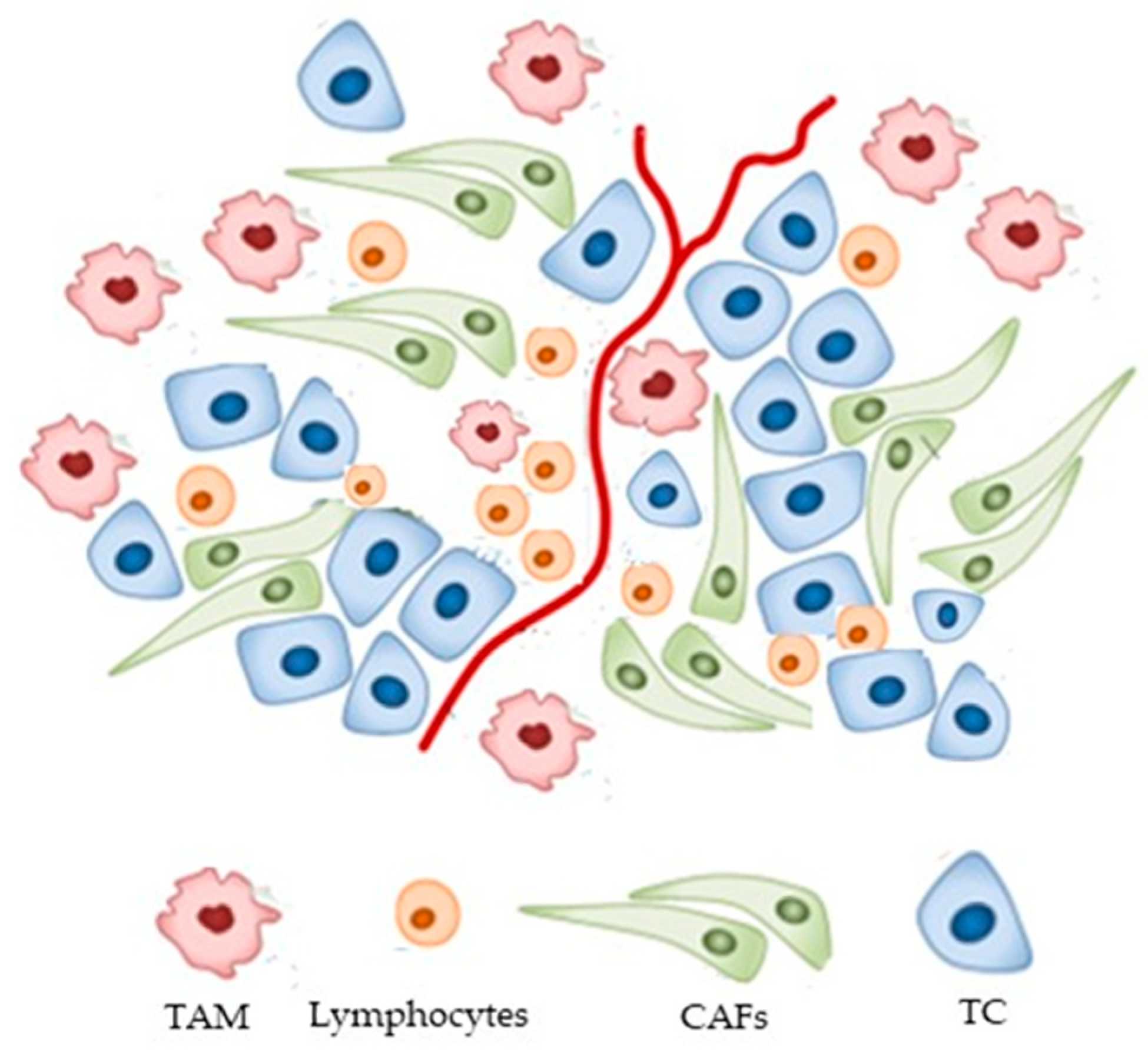
| Phenotype | Cytokines/Chemokines/TFs | References |
|---|---|---|
| M0 | None | |
| M1 | IL-6, TNF-α, IL-1β, NF-κB, AP-1, | [5] |
| M2 | IL-10, TGF-β, IL-6 | [5] |
| M2a | STAT6, JAK, FN, IGF, TGF-β, IL-10, CCL-17, CCL18, CCL22 | [13] |
| M2b | AP-1, ERK, IRFs, IL-10, TNF-α, IL1 | [14,15] |
| M2c | STAT3, IL-10, TGF-β | [14] |
| M2d | HIF-1α, VEGF, IL-10, TNF-α, IL-12 | [16] |
| Phenotypes | Molecules Involved | References |
|---|---|---|
| M1-CAFs | CD200R, CD209 | [51] |
| am(M1)-CAFs | IL-6 | [52,53] |
| TAM-CSCs | VEGFA, ITGB3BP, ADAM9 | [54,55] |
| CSCs-TAM | GAS6, ADAM9, ANXA1c | [55] |
| AM-moMDSC | CCL12 | [56] |
| TAM-TME cells | CXCL9 | [57,58] |
| TCs-TAM | HMGB1, LncRNA-HOXC-AS2 | [59,60] |
| Nodes | Molecule Involved | References |
|---|---|---|
| Immune checkpoints | PDL-1/PD-1 | [78,79,80,81,82,83] |
| LncRNAs | HOXC-AS2, NBR2 | [60] |
| miRNAs | miRNA-19a, miRNA-210-3p, miR-181b, miRNA-214, miRNA-21 | [66,84,85,86,87] |
| Signaling pathway | JAK/STAT | [22,85,86,88,89,90] |
| Receptors | TLR7/8, TREM1, TGF-βR1 | [91,92,93,94,95,96] |
| Extracellular matrix | SPON2, BMP2 | [97,98,99] |
| Cytokine | CSF-1/CSFR-1 | [100,101,102] |
| Chemokine | CXCL12 | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lampiasi, N. New Strategies for Macrophage Re-Education in Cancer: An Update. Int. J. Mol. Sci. 2024, 25, 3414. https://doi.org/10.3390/ijms25063414
Lampiasi N. New Strategies for Macrophage Re-Education in Cancer: An Update. International Journal of Molecular Sciences. 2024; 25(6):3414. https://doi.org/10.3390/ijms25063414
Chicago/Turabian StyleLampiasi, Nadia. 2024. "New Strategies for Macrophage Re-Education in Cancer: An Update" International Journal of Molecular Sciences 25, no. 6: 3414. https://doi.org/10.3390/ijms25063414
APA StyleLampiasi, N. (2024). New Strategies for Macrophage Re-Education in Cancer: An Update. International Journal of Molecular Sciences, 25(6), 3414. https://doi.org/10.3390/ijms25063414