
MsHDZ23, a Novel Miscanthus HD-ZIP Transcription Factor, Participates in Tolerance to Multiple Abiotic Stresses

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Genome-Wide Identification of HD-ZIP Genes in M. sinensis
2.2. Expression Profiling of MsHD-ZIP Genes in Multiple Abiotic Stresses
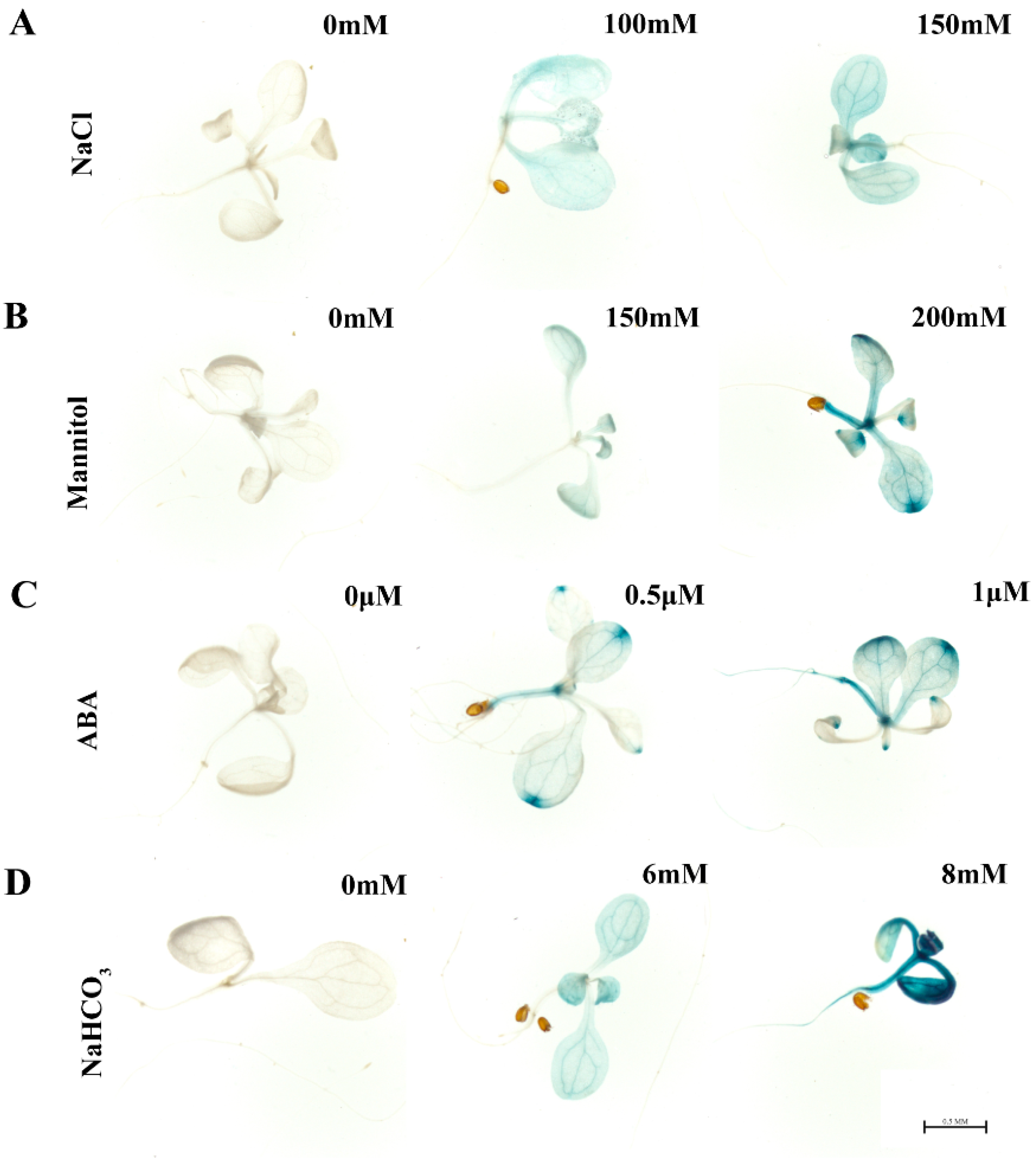
2.3. MsHDZ23 Was Induced by Multiple Abiotic Stresses
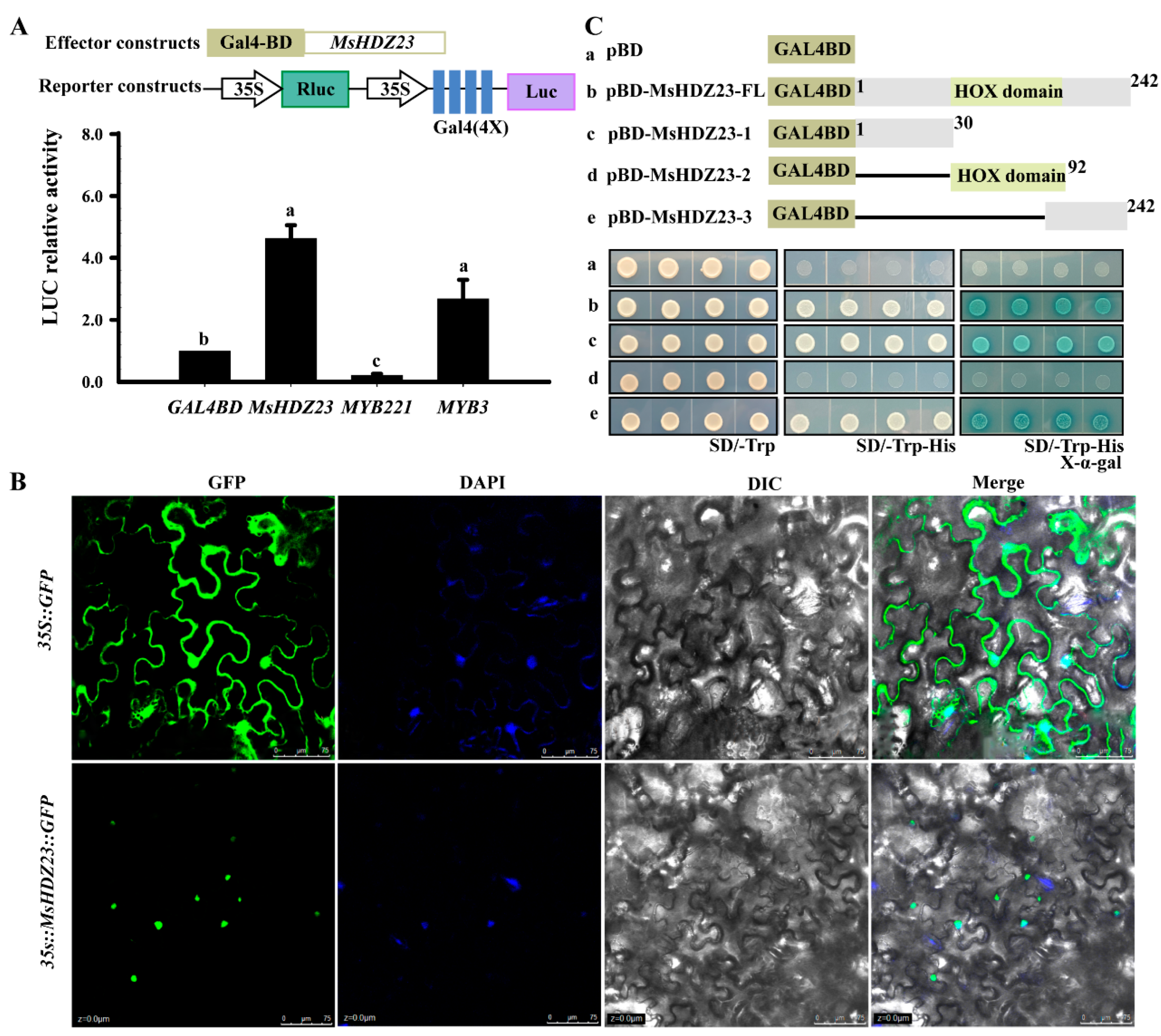
2.4. MsHDZ23 Is a Nucleus-Localized TF with Transactivation Activity
2.5. MsHDZ23 Overexpression Affects the Development of Transgenic Arabidopsis
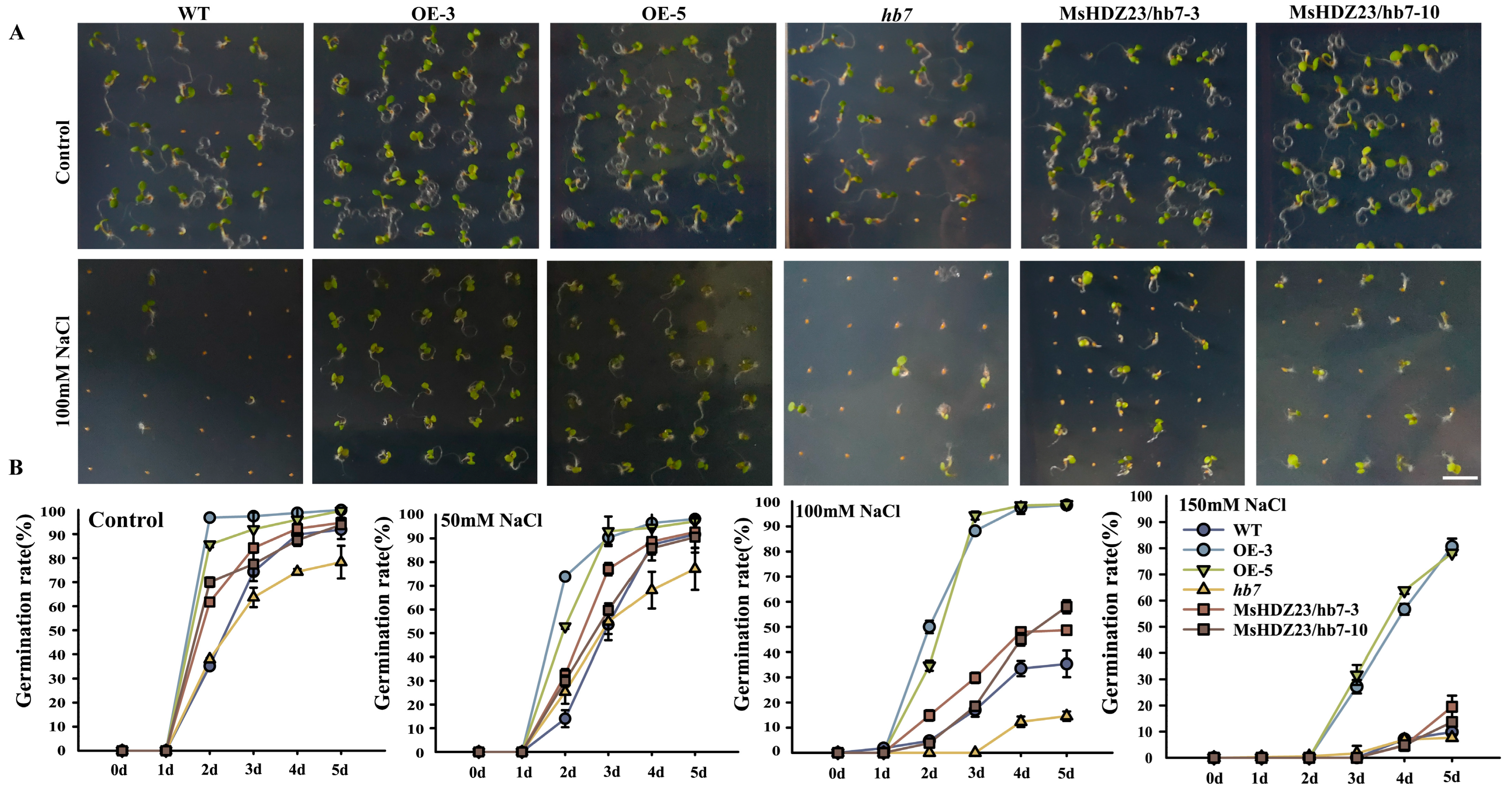
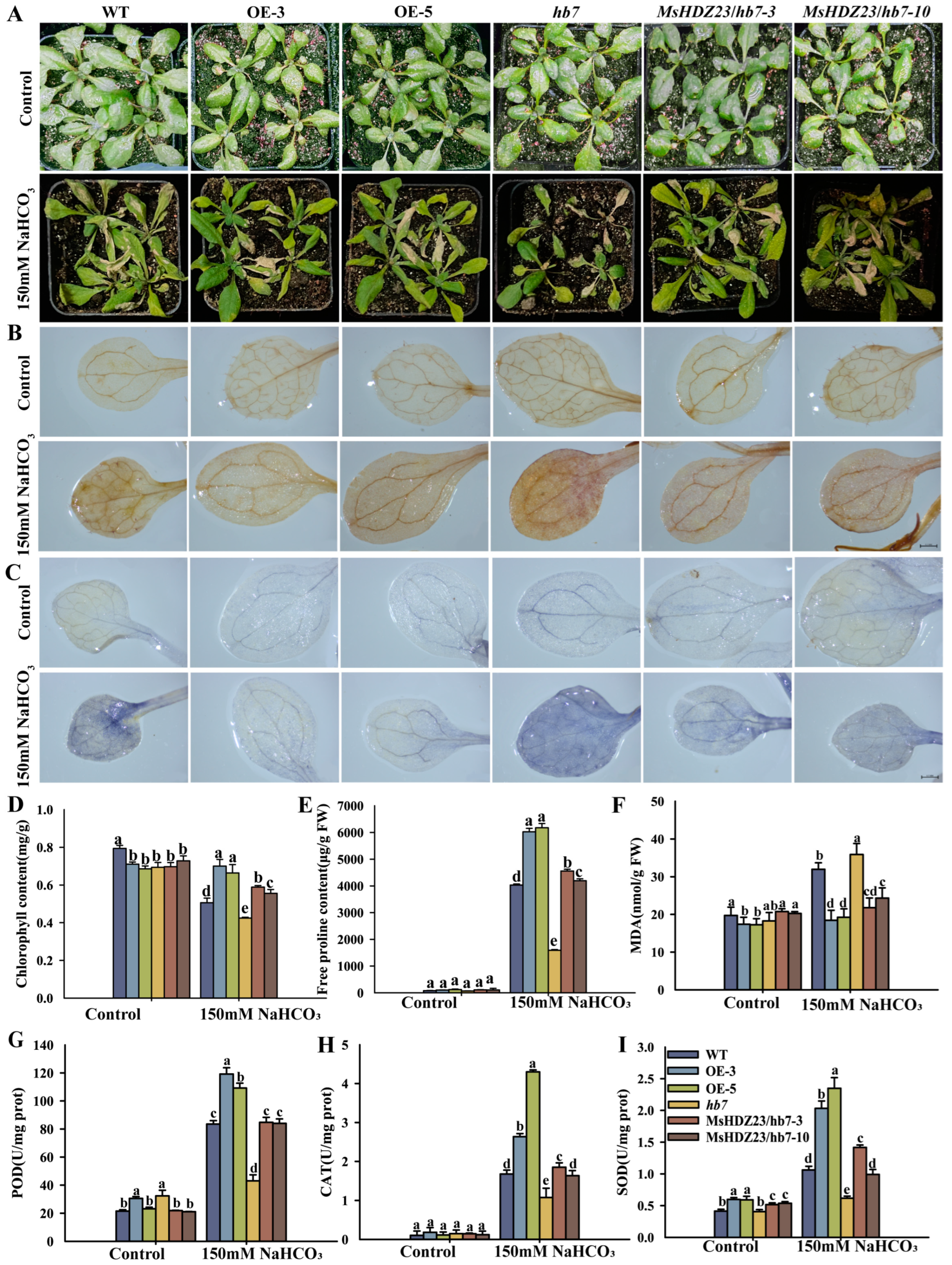
2.6. MsHDZ23 Overexpression Improves Salt Tolerance
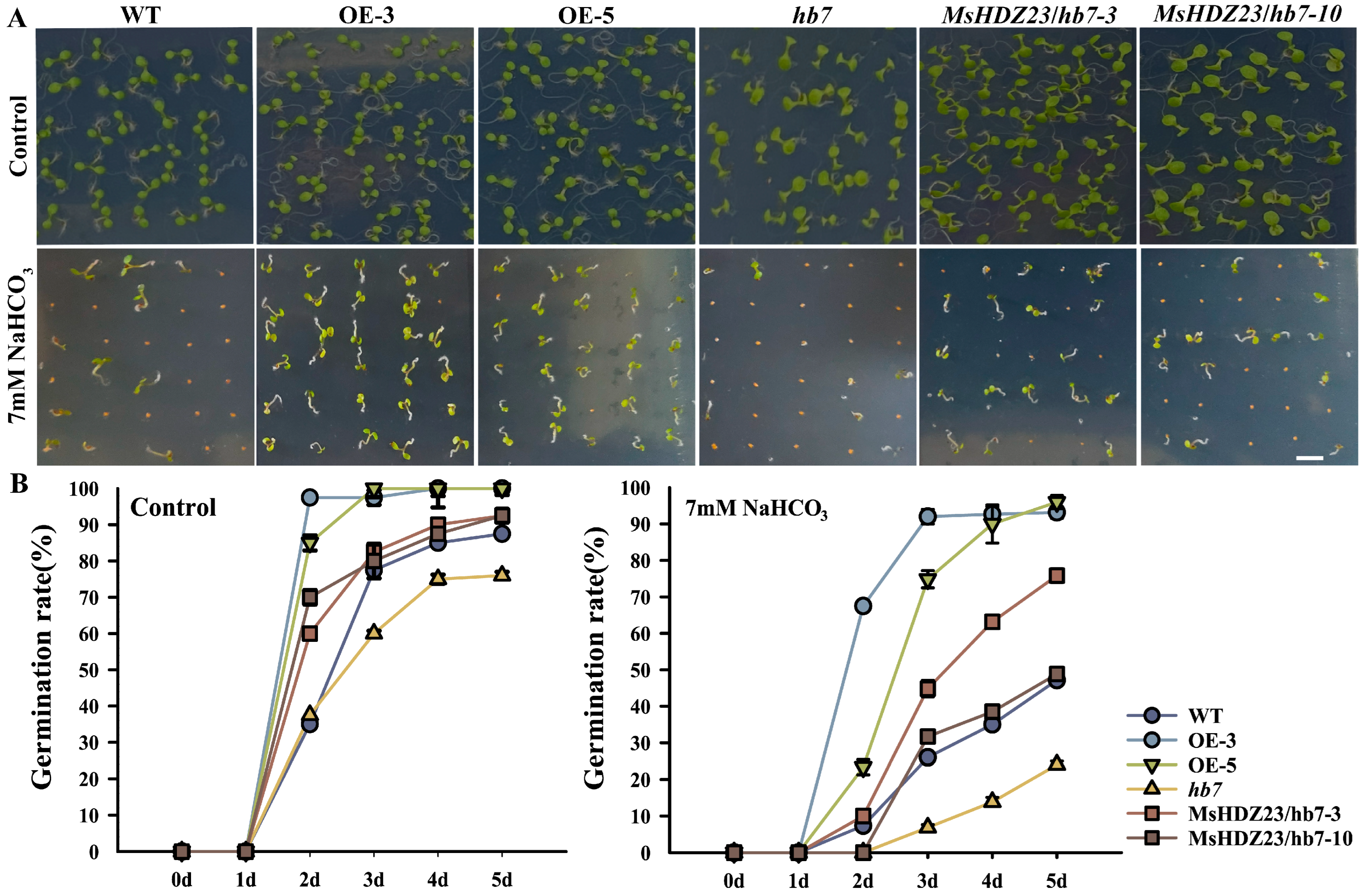
2.7. MsHDZ23 Overexpression Improves Alkali Tolerance
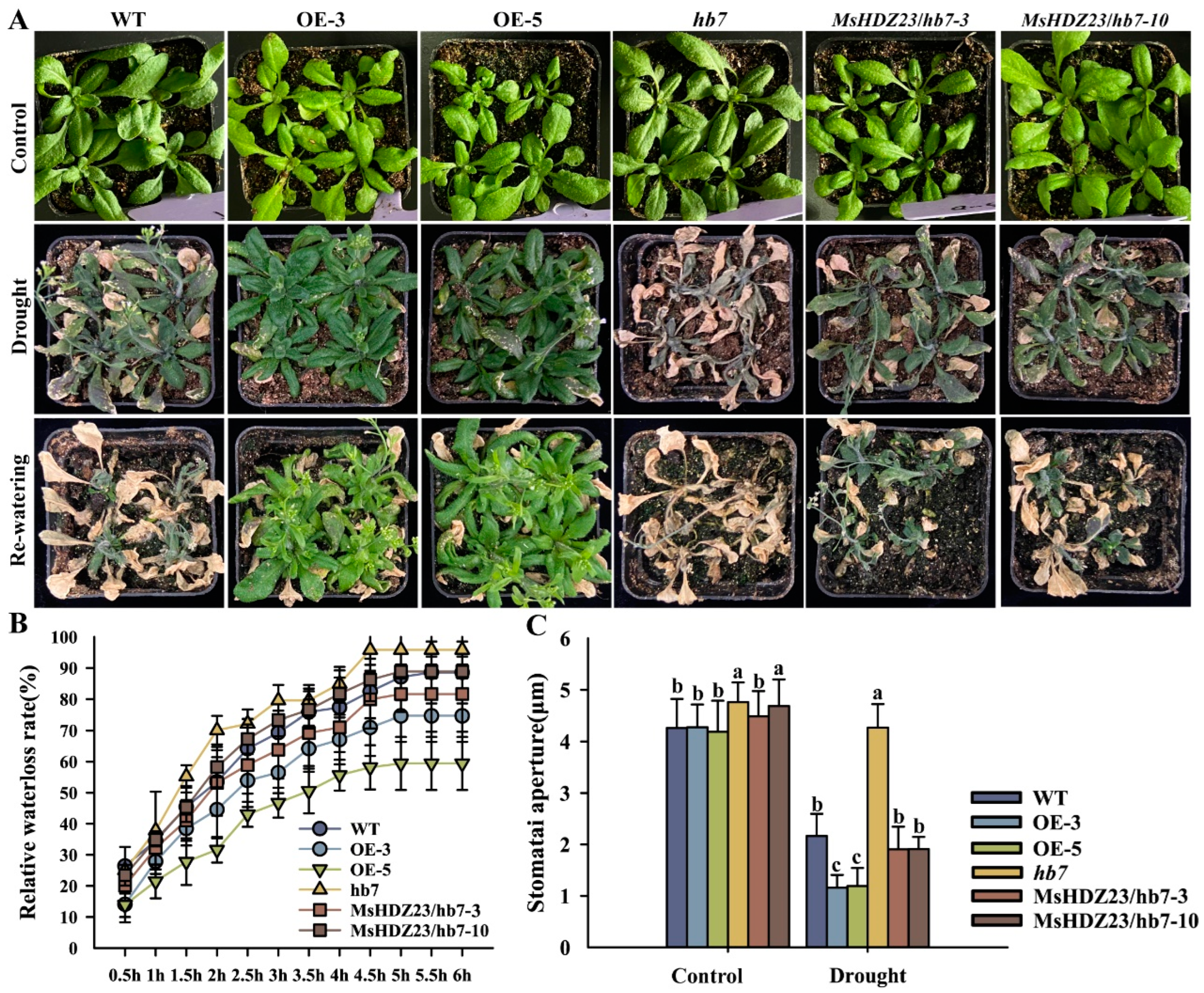
2.8. Overexpression of MsHDZ23 Enhances Drought Tolerance
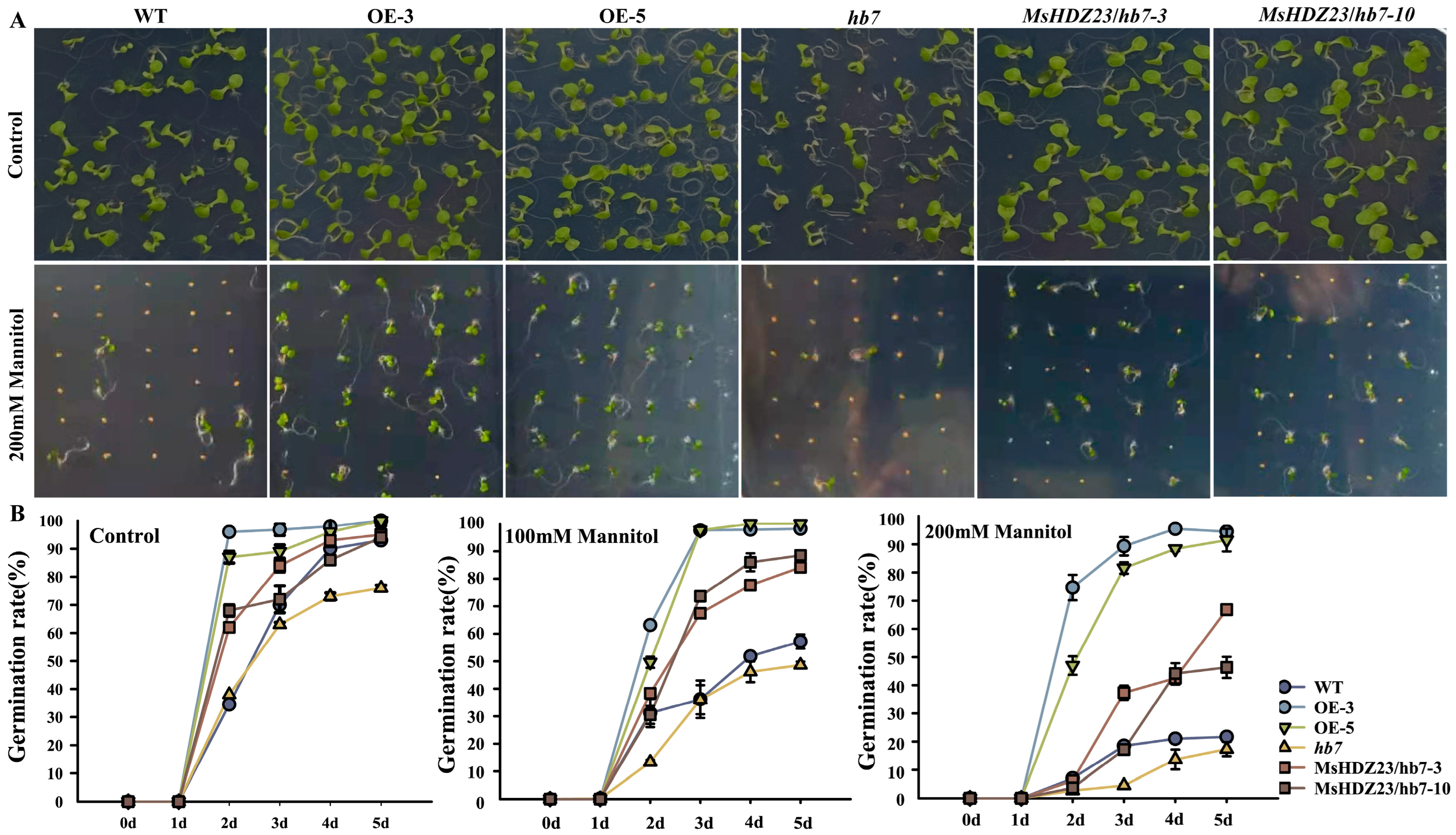
2.9. MsHDZ23 Overexpression Confers ABA Insensitivity
3. Discussion
4. Materials and Methods
4.1. The Identification of HD-ZIP Genes in M. sinensis
4.2. Phylogenetic Analysis
4.3. Plant Materials and Abiotic Stress Treatment
4.4. Plasmid Construction and Transformation of Arabidopsis
4.5. Histochemical GUS Staining
4.6. Subcellular Localization
4.7. Transactivation and Transcriptional Activation Analysis
4.8. Seedling Growth Assays
4.9. Evaluation of Drought, Salt, and Alkali Tolerance
4.10. Physiological Measurements and Histochemical Assays
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Physiol. Plant. 2021, 173, 148–166. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Gong, Q.; Li, P.; Ma, S. Unraveling abiotic stress tolerance mechanisms–getting genomics going. Curr. Opin. Plant Biol. 2006, 9, 180–188. [Google Scholar] [CrossRef]
- Mehrotra, R.; Bhalothia, P.; Bansal, P.; Basantani, M.K.; Bharti, V.; Mehrotra, S. Abscisic acid and abiotic stress tolerance-Different tiers of regulation. J. Plant Physiol. 2014, 171, 486–496. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Sarvajeet, S.G.; Narendra, T. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Lan Thi Hoang, X.; Du Nhi, N.H.; Binh Anh Thu, N.; Phuong Thao, N.; Phan Tran, L.S. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar]
- Wang, H.Y.; Wang, H.L.; Shao, H.B.; Tang, X.L. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 13. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Jha, B. Transcription factors in plants and ABA dependent and independent abiotic stress signalling. Biol. Plant. 2010, 54, 201–212. [Google Scholar] [CrossRef]
- Li, Y.X.; Yang, Z.R.; Zhang, Y.Y.; Guo, J.J.; Liu, L.L.; Wang, C.F.; Wang, B.S.; Han, G.L. The roles of HD-ZIP proteins in plant abiotic stress tolerance. Front. Plant Sci. 2022, 13, 1027071. [Google Scholar] [CrossRef]
- Xie, L.H.; Yan, T.X.; Li, L.; Chen, M.H.; Hassani, D.; Li, Y.P.; Qin, W.; Liu, H.; Chen, T.T.; Fu, X.Q.; et al. An HD-ZIP-MYB complex regulates glandular secretory trichome initiation in Artemisia annua. New Phytol. 2021, 231, 2050–2064. [Google Scholar] [CrossRef]
- Mehmood, M.; Khan, M.J.; Khan, M.J.; Akhtar, N.; Mughal, F.; Shah, S.T.A.; Hyder, M.Z.; Farrakh, S.; Sadiq, I. Systematic analysis of HD-ZIP transcription factors in sesame genome and gene expression profiling of SiHD-ZIP class I entailing drought stress responses at early seedling stage. Mol. Biol. Rep. 2022, 49, 2059–2071. [Google Scholar] [CrossRef]
- Tan, W.; Zhang, D.; Zhou, H.; Zheng, T.; Yin, Y.; Lin, H. Transcription factor HAT1 is a substrate of SnRK2.3 kinase and negatively regulates ABA synthesis and signaling in Arabidopsis responding to drought. PLoS Genet. 2018, 14, e1007336. [Google Scholar] [CrossRef]
- Liu, T.; Longhurst, A.D.; Talavera-Rauh, F.; Hokin, S.A.; Barton, M.K. The Arabidopsis transcription factor ABIG1 relays ABA signaled growth inhibition and drought induced senescence. eLife 2016, 5, 19. [Google Scholar] [CrossRef]
- Li, X.; Hou, Y.Y.; Li, M.N.; Zhang, F.; Yi, F.Y.; Kang, J.M.; Yang, Q.C.; Long, R.C. Overexpression of an ABA-inducible homeodomain-leucine zipper I gene MsHB7 confers salt stress sensitivity to alfalfa. Ind. Crops Prod. 2022, 177, 114463. [Google Scholar] [CrossRef]
- Sen, S.; Chakraborty, J.; Ghosh, P.; Basu, D.; Das, S. Chickpea WRKY70 Regulates the Expression of a Homeodomain-Leucine Zipper (HD-Zip) I Transcription Factor CaHDZ12, which Confers Abiotic Stress Tolerance in Transgenic Tobacco and Chickpea. Plant Cell Physiol. 2017, 58, 1934–1952. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Ida, Y.; Kitajima, S.; Kawazu, T.; Hibino, T.; Hanba, Y.T. Overexpressing the HD-Zip class II transcription factor EcHB1 from Eucalyptus camaldulensis increased the leaf photosynthesis and drought tolerance of Eucalyptus. Sci. Rep. 2019, 9, 14121. [Google Scholar] [CrossRef]
- Kovalchuk, N.; Chew, W.; Sornaraj, P.; Borisjuk, N.; Yang, N.N.; Singh, R.; Bazanova, N.; Shavrukov, Y.; Guendel, A.; Munz, E.; et al. The homeodomain transcription factor TaHDZipI-2 from wheat regulates frost tolerance, flowering time and spike development in transgenic barley. New Phytol. 2016, 211, 671–687. [Google Scholar] [CrossRef]
- Li, S.M.; Chen, N.; Li, F.F.; Mei, F.M.; Wang, Z.X.; Cheng, X.X.; Kang, Z.S.; Mao, H.D. Characterization of wheat homeo-domain-leucine zipper family genes and functional analysis of TaHDZ5-6A in drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2020, 20, 23. [Google Scholar]
- Yang, Y.F.; Luang, S.; Harris, J.; Riboni, M.; Li, Y.; Bazanova, N.; Hrmova, M.; Haefele, S.; Kovalchuk, N.; Lopato, S. Overexpression of the class I homeodomain transcription factor TaHDZipI-5 increases drought and frost tolerance in transgenic wheat. Plant Biotechnol. J. 2018, 16, 1227–1240. [Google Scholar] [CrossRef]
- Depège-Fargeix, N.; Javelle, M.; Chambrier, P.; Frangne, N.; Gerentes, D.; Perez, P.; Rogowsky, P.M.; Vernoud, V. Functional characterization of the HD-ZIP IV transcription factor OCL1 from maize. J. Exp. Bot. 2011, 62, 293–305. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, Q.; Jin, X.L.; Peng, X.J.; Liu, J.Y.; Deng, L.; Yan, H.W.; Sheng, L.; Jiang, H.Y.; Cheng, B.J. A Novel Maize Homeodomain–Leucine Zipper (HD-Zip) I Gene, Zmhdz10, Positively Regulates Drought and Salt Tolerance in Both Rice and Arabidopsis. Plant Cell Physiol. 2014, 55, 1142–1156. [Google Scholar] [CrossRef]
- Qiu, X.; Wang, G.R.; Abou-Elwafa, S.F.; Fu, J.X.; Liu, Z.X.; Zhang, P.Y.; Xie, X.W.; Ku, L.X.; Ma, Y.; Guan, X.K.; et al. Genome-wide identification of HD-ZIP transcription factors in maize and their regulatory roles in promoting drought tolerance. Physiol. Mol. Biol. Plants 2022, 28, 425–437. [Google Scholar] [CrossRef]
- Zhang, S.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.H.; Schluepmann, H.; Liu, C.-M.; Ouwerkerk, P.B.F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Ding, Y.F.; Gong, S.H.; Wang, Y.; Wang, F.J.; Bao, H.; Sun, J.W.; Cai, C.; Yi, K.K.; Chen, Z.X.; Zhu, C. MicroRNA166 Modulates Cadmium Tolerance and Accumulation in Rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef]
- Bang, S.W.; Lee, D.K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Suh, J.W.; Kim, J.K. Overexpression of OsTF1L, a rice HD-Zip transcription factor, promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant Biotechnol. J. 2018, 17, 118–131. [Google Scholar] [CrossRef]
- Dong, N.Q.; Sun, Y.W.; Guo, T.; Shi, C.L.; Zhang, Y.M.; Kan, Y.; Xiang, Y.H.; Zhang, H.; Yang, Y.B.; Li, Y.C.; et al. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 16. [Google Scholar] [CrossRef]
- Kaur, R.; Das, S.; Bansal, S.; Singh, G.; Sardar, S.; Dhar, H.; Ram, H. Heavy metal stress in rice: Uptake, transport, signaling, and tolerance mechanisms. Physiol. Plant. 2021, 173, 430–448. [Google Scholar] [CrossRef]
- Brosse, N.; Dufour, A.; Meng, X.; Sun, Q.; Ragauskas, A. Miscanthus: A fast-growing crop for biofuels and chemicals production. Biofuels, Bioprod. Biorefining 2012, 6, 580–598. [Google Scholar] [CrossRef]
- Lee, W.C.; Kuan, W.C. Miscanthus as cellulosic biomass for bioethanol production. Biotechnol. J. 2015, 10, 840–854. [Google Scholar] [CrossRef]
- Hu, S.-W.; Wu, L.-M.; Persson, S.; Peng, L.-C.; Feng, S.-Q. Sweet sorghum and Miscanthus: Two potential dedicated bioenergy crops in China. J. Integr. Agric. 2017, 16, 1236–1243. [Google Scholar] [CrossRef]
- Liu, W.; Mi, J.; Song, Z.; Yan, J.; Li, J.; Sang, T. Long-term water balance and sustainable production of Miscanthus energy crops in the Loess Plateau of China. Biomass-Bioenergy 2014, 62, 47–57. [Google Scholar] [CrossRef]
- Xue, S.; Lewandowski, I.; Wang, X.Y.; Yi, Z.L. Assessment of the production potentials of Miscanthus on marginal land in China. Renew. Sustain. Energy Rev. 2016, 54, 932–943. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, C.; Liang, L.; Yi, Z.L.; Xue, S. Quantitative assessment of the potential for soil improvement by planting Miscanthus on saline-alkaline soil and the underlying microbial mechanism. Glob. Change Biol. Bioenergy 2021, 13, 1191–1205. [Google Scholar] [CrossRef]
- Bevan, M.W.; Uauy, C.; Wulff, B.B.H.; Zhou, J.; Krasileva, K.; Clark, M.D. Genomic innovation for crop improvement. Nature 2017, 543, 346–354. [Google Scholar] [CrossRef]
- Chai, G.H.; Qi, G.; Cao, Y.P.; Wang, Z.G.; Yu, L.; Tang, X.F.; Yu, Y.C.; Wang, D.; Kong, Y.Z.; Zhou, G.K. Poplar PdC3H17 and PdC3H18 are direct targets of PdMYB3 and PdMYB21, and positively regulate secondary wall formation in Arabidopsis and poplar. New Phytol. 2014, 203, 520–534. [Google Scholar] [CrossRef]
- Tang, X.; Zhuang, Y.; Qi, G.; Wang, D.; Liu, H.; Wang, K.; Chai, G.; Zhou, G. Poplar PdMYB221 is involved in the direct and indirect regulation of secondary wall biosynthesis during wood formation. Sci. Rep. 2015, 5, 12240. [Google Scholar] [CrossRef]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; Kirch, H.-H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ. 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and powerful stress gene expression in Arabidopsis in response to malondialdehyde. Plant J. 2004, 37, 877–888. [Google Scholar] [CrossRef]
- Huang, X.-S.; Liu, J.-H.; Chen, X.-J. Overexpression of PtrABF gene, a bZIP transcription factor isolated from Poncirus trifoliata, enhances dehydration and drought tolerance in tobacco via scavenging ROS and modulating expression of stress-responsive genes. BMC Plant Biol. 2010, 10, 230. [Google Scholar] [CrossRef]
- Jaspers, P.; KangasjcTrvi, J. Reactive oxygen species in abiotic stress signaling. Physiol. Plant. 2010, 138, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, T.; Shen, C.-Y.; Xu, G.-H.; Chen, S.-X.; Song, L.-Z.; Li, M.-J.; Wang, L.-L.; Zhu, Y.; Lv, W.-T.; et al. Identification of a Retroelement from the Resurrection Plant Boea hygrometrica That Confers Osmotic and Alkaline Tolerance in Arabidopsis thaliana. PLoS ONE 2014, 9, e98098. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Shi, L.; Yang, Y. Germination, Growth, Osmotic Adjustment and Ionic Balance of Wheat in Response to Saline and Alkaline Stresses; Taylor & Francis Group: Abingdon, UK, 2009. [Google Scholar]
- Cao, L.; Yu, Y.; DuanMu, H.Z.; Chen, C.; Duan, X.B.; Zhu, P.H.; Chen, R.R.; Li, Q.; Zhu, Y.M.; Ding, X.D. A novel Glycine soja homeodomain-leucine zipper (HD-Zip) I gene, Gshdz4, positively regulates bicarbonate tolerance and responds to osmotic stress in Arabidopsis. BMC Plant Biol. 2016, 16, 14. [Google Scholar] [CrossRef]
- Zhu, D.; Bai, X.; Chen, C.; Chen, Q.; Cai, H.; Li, Y.; Ji, W.; Zhai, H.; Lv, D.; Luo, X.; et al. GsTIFY10, a novel positive regulator of plant tolerance to bicarbonate stress and a repressor of jasmonate signaling. Plant Mol. Biol. 2011, 77, 285–297. [Google Scholar] [CrossRef]
- Nie, G.; Liao, Z.; Zhong, M.; Zhou, J.; Cai, J.; Liu, A.; Wang, X.; Zhang, X. MicroRNA-Mediated Responses to Chromium Stress Provide Insight Into Tolerance Characteristics of Miscanthus sinensis. Front. Plant Sci. 2021, 12, 666117. [Google Scholar] [CrossRef]
- Mitros, T.; Session, A.M.; James, B.T.; Wu, G.A.; Belaffif, M.B.; Clark, L.V.; Shu, S.; Dong, H.; Barling, A.; Holmes, J.R.; et al. Genome biology of the paleotetraploid perennial biomass crop Miscanthus. Nat. Commun. 2020, 11, 5442. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, H.; Zhang, K.; Li, F.; Wu, M.; Xiang, Y. A moso bamboo transcription factor, Phehdz1, positively regulates the drought stress response of transgenic rice. Plant Cell Rep. 2021, 40, 187–204. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Li, Y.; Zhu, Y.-M.; Bai, X.; Lv, D.-K.; Guo, D.; Ji, W.; Cai, H. Global transcriptome profiling of wild soybean (Glycine soja) roots under NaHCO3treatment. BMC Plant Biol. 2010, 10, 153. [Google Scholar] [CrossRef]
- DuanMu, H.; Wang, Y.; Bai, X.; Cheng, S.; Deyholos, M.K.; Wong, G.K.-S.; Li, D.; Zhu, D.; Li, R.; Yu, Y.; et al. Wild soybean roots depend on specific transcription factors and oxidation reduction related genesin response to alkaline stress. Funct. Integr. Genom. 2015, 15, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Koropatkin, N.M.; Koppenaal, D.W.; Pakrasi, H.B.; Smith, T.J. The structure of a cyanobacterial bicarbonate transport protein, CmpA. J. Biol. Chem. 2007, 282, 2606–2614. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, C.; Liu, G.; Jiang, J. Development of a cDNA microarray to identify gene expression of Puccinellia tenuiflora under saline–alkali stress. Plant Physiol. Biochem. 2007, 45, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Chew, W.; Hrmova, M.; Lopato, S. Role of Homeodomain leucine zipper (HD-Zip) IV transcription factors in plant development and plant protection from deleterious environmental factors. Int. J. Mol. Sci. 2013, 14, 8122–8147. [Google Scholar] [CrossRef] [PubMed]
- Khosla, A.; Paper, J.M.; Boehler, A.P.; Bradley, A.M.; Neumann, T.R.; Schrick, K. HD-Zip Proteins GL2 and HDG11 Have Redundant Functions in Arabidopsis Trichomes, and GL2 Activates a Positive Feedback Loop via MYB23. Plant Cell 2014, 26, 2184–2200. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, B.; Xu, X.; Chen, H.; Zou, L.; Chen, G.; Cao, B.; Chen, C.; Lei, J. Overexpression of AtEDT1/HDG11 in Chinese Kale (Brassica oleracea var. alboglabra) Enhances Drought and Osmotic Stress Tolerance. Front. Plant Sci. 2016, 7, 1285. [Google Scholar]
- Zhao, X.; Xu, Y.; He, G.; He, K.; Xiao, L.; Hu, R.; Li, S. Genome-Wide Characterization and Expression Profiling of the GRAS Gene Family in Salt and Alkali Stresses in Miscanthus sinensis. Int. J. Mol. Sci. 2022, 23, 14521. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-C.; Liao, P.-M.; Kuo, W.-W.; Lin, T.-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 Regulates Abiotic Stress-Responsive Gene Expression by Binding to Different cis-Acting Elements in Response to Different Stress Signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef]
- Yang, X.; He, K.; Chi, X.; Chai, G.; Wang, Y.; Jia, C.; Zhang, H.; Zhou, G.; Hu, R. Miscanthus NAC transcription factor MlNAC12 positively mediates abiotic stress tolerance in transgenic Arabidopsis. Plant Sci. 2018, 277, 229–241. [Google Scholar] [CrossRef]
- Liu, H.; Guo, S.; Xu, Y.; Li, C.; Zhang, Z.; Zhang, D.; Xu, S.; Zhang, C.; Chong, K. OsmiR396d-Regulated OsGRFs Function in Floral Organogenesis in Rice through Binding to Their Targets OsJMJ706 and OsCR4. Plant Physiol. 2014, 165, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free. Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, N.; Yu, R.; Deng, W.; Hu, R.; He, G.; He, K.; Kong, Y.; Tang, X.; Zhou, G.; Wang, C. MsHDZ23, a Novel Miscanthus HD-ZIP Transcription Factor, Participates in Tolerance to Multiple Abiotic Stresses. Int. J. Mol. Sci. 2024, 25, 3253. https://doi.org/10.3390/ijms25063253
Liu N, Yu R, Deng W, Hu R, He G, He K, Kong Y, Tang X, Zhou G, Wang C. MsHDZ23, a Novel Miscanthus HD-ZIP Transcription Factor, Participates in Tolerance to Multiple Abiotic Stresses. International Journal of Molecular Sciences. 2024; 25(6):3253. https://doi.org/10.3390/ijms25063253
Chicago/Turabian StyleLiu, Naixu, Ruikang Yu, Wendi Deng, Ruibo Hu, Guo He, Kang He, Yingzhen Kong, Xianfeng Tang, Gongke Zhou, and Congpeng Wang. 2024. "MsHDZ23, a Novel Miscanthus HD-ZIP Transcription Factor, Participates in Tolerance to Multiple Abiotic Stresses" International Journal of Molecular Sciences 25, no. 6: 3253. https://doi.org/10.3390/ijms25063253
APA StyleLiu, N., Yu, R., Deng, W., Hu, R., He, G., He, K., Kong, Y., Tang, X., Zhou, G., & Wang, C. (2024). MsHDZ23, a Novel Miscanthus HD-ZIP Transcription Factor, Participates in Tolerance to Multiple Abiotic Stresses. International Journal of Molecular Sciences, 25(6), 3253. https://doi.org/10.3390/ijms25063253