
Skin Anti-Aging Potential through Whitening and Wrinkle Improvement Using Fermented Oil Derived from Hermetia illucens Larvae


Abstract
1. Introduction
2. Results
2.1. Evaluation of the Lipid Fermentation Abilities in 10 Selected Strains
2.2. Evaluation of the Fermentation Conditions of Hermetia illucens Oil Fermented with Selected Strains
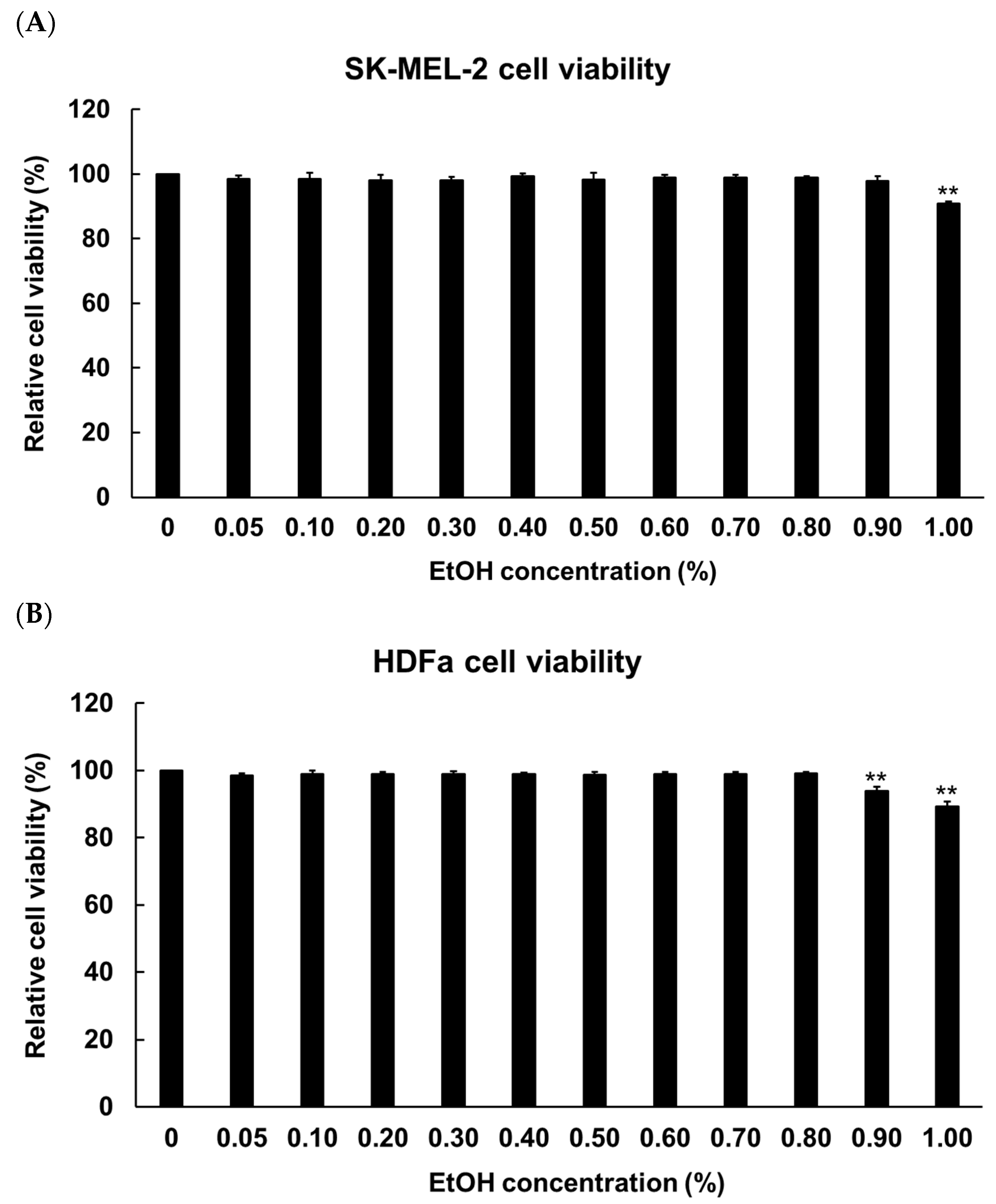
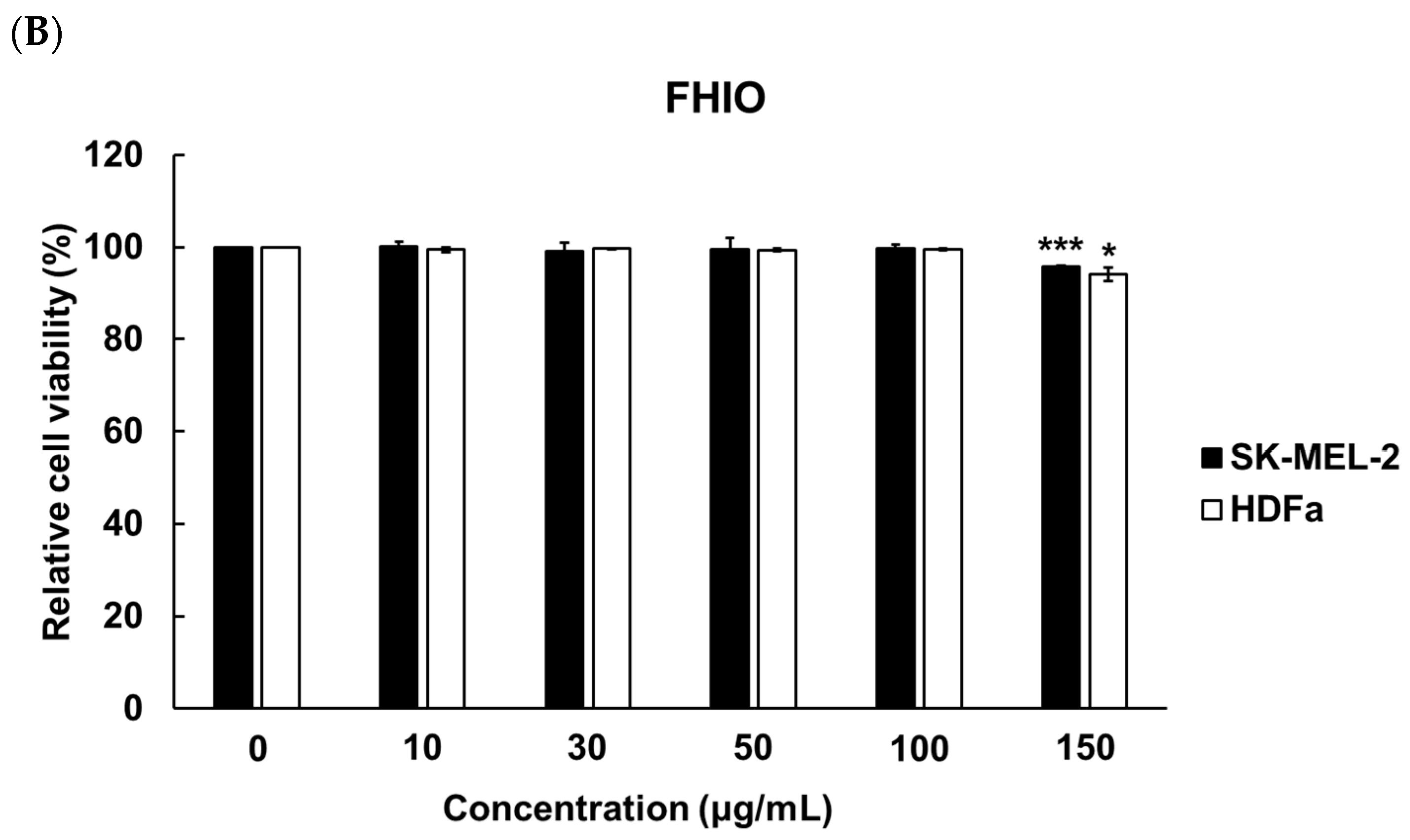
2.3. Evaluation of Cytotoxicity for the Determination of Solvent Concentration and Hermetia illucens Oil Treatment Concentration in Melanoma Cells and Fibroblasts
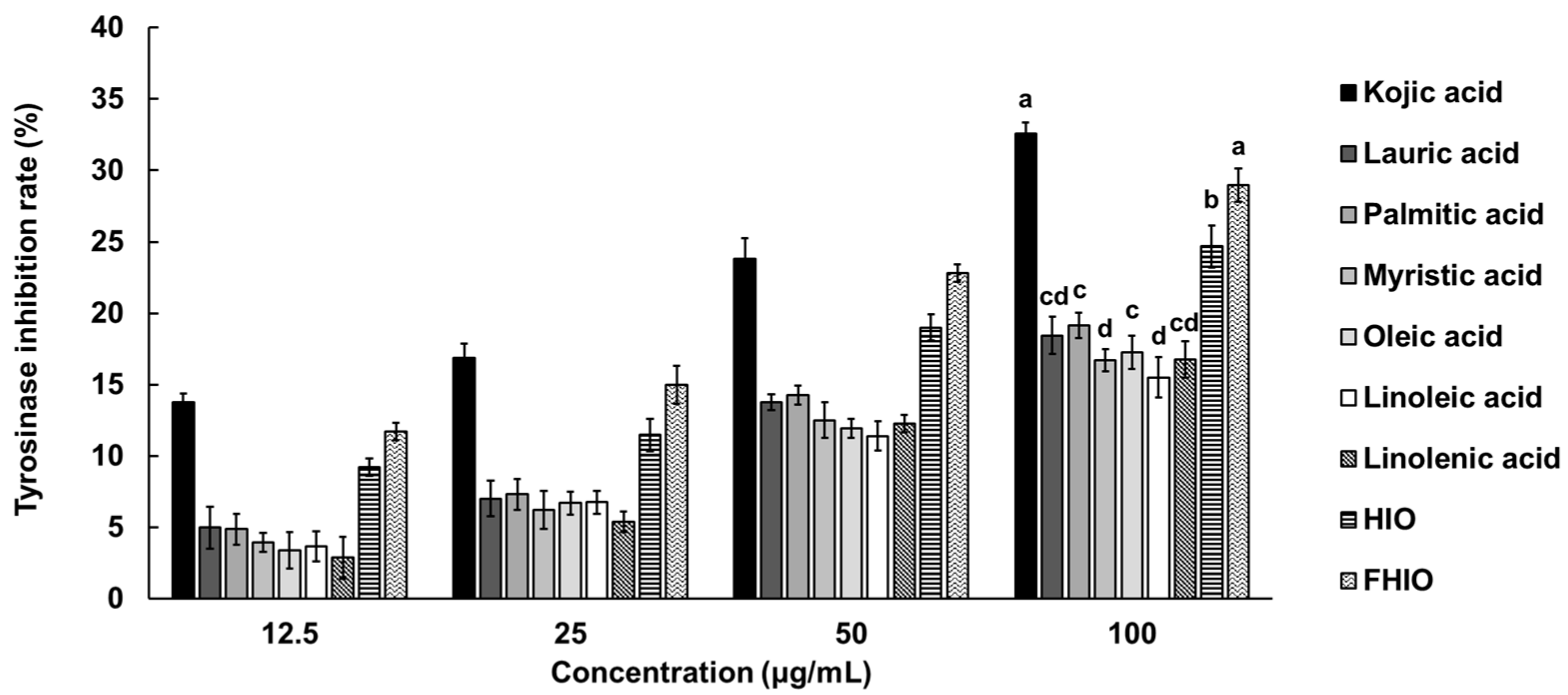
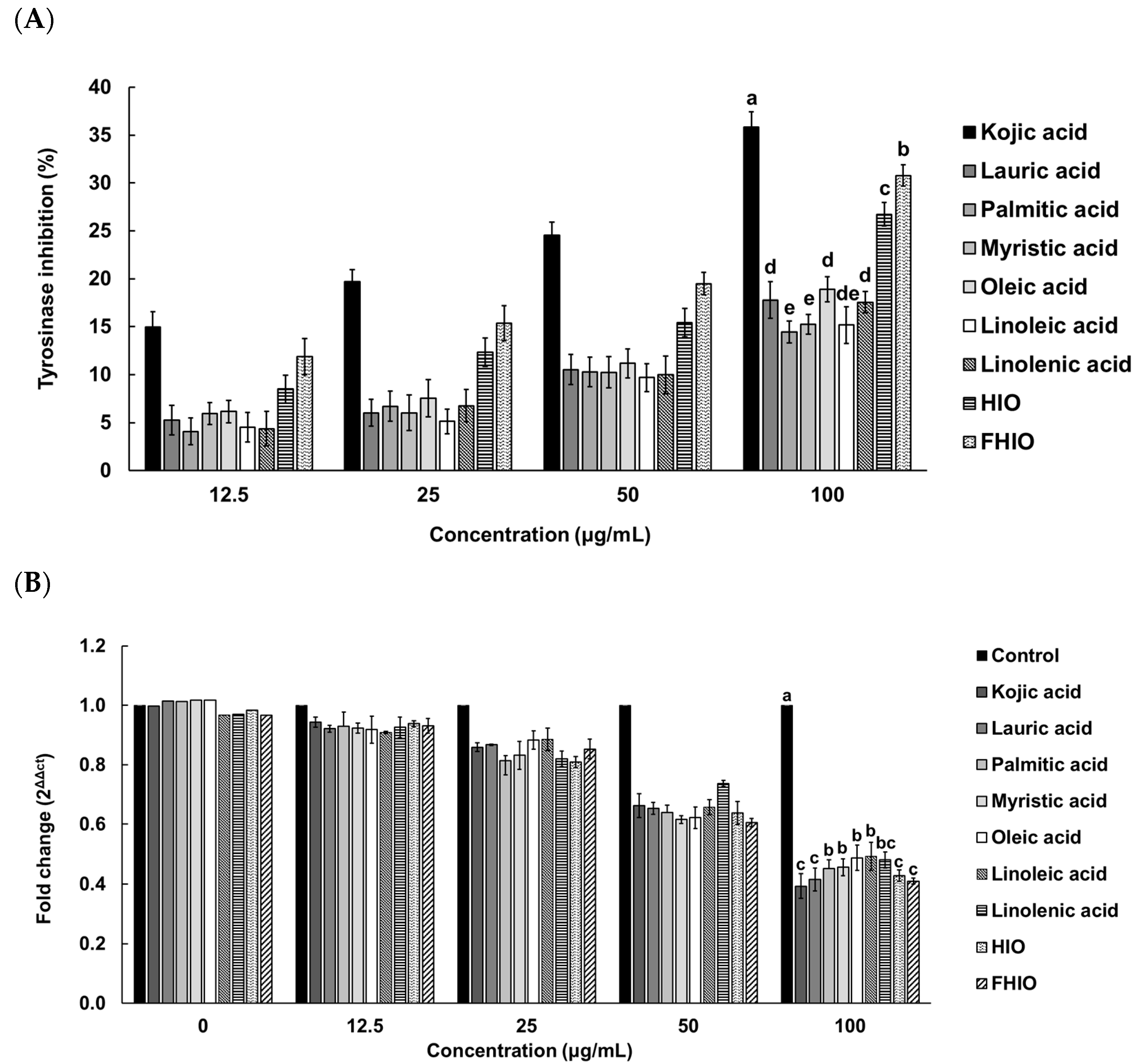
2.4. Evaluation of the Skin-Whitening Potential of Hermetia illucens Oil and Fermented Hermetia illucens Oil in Melanoma Cells
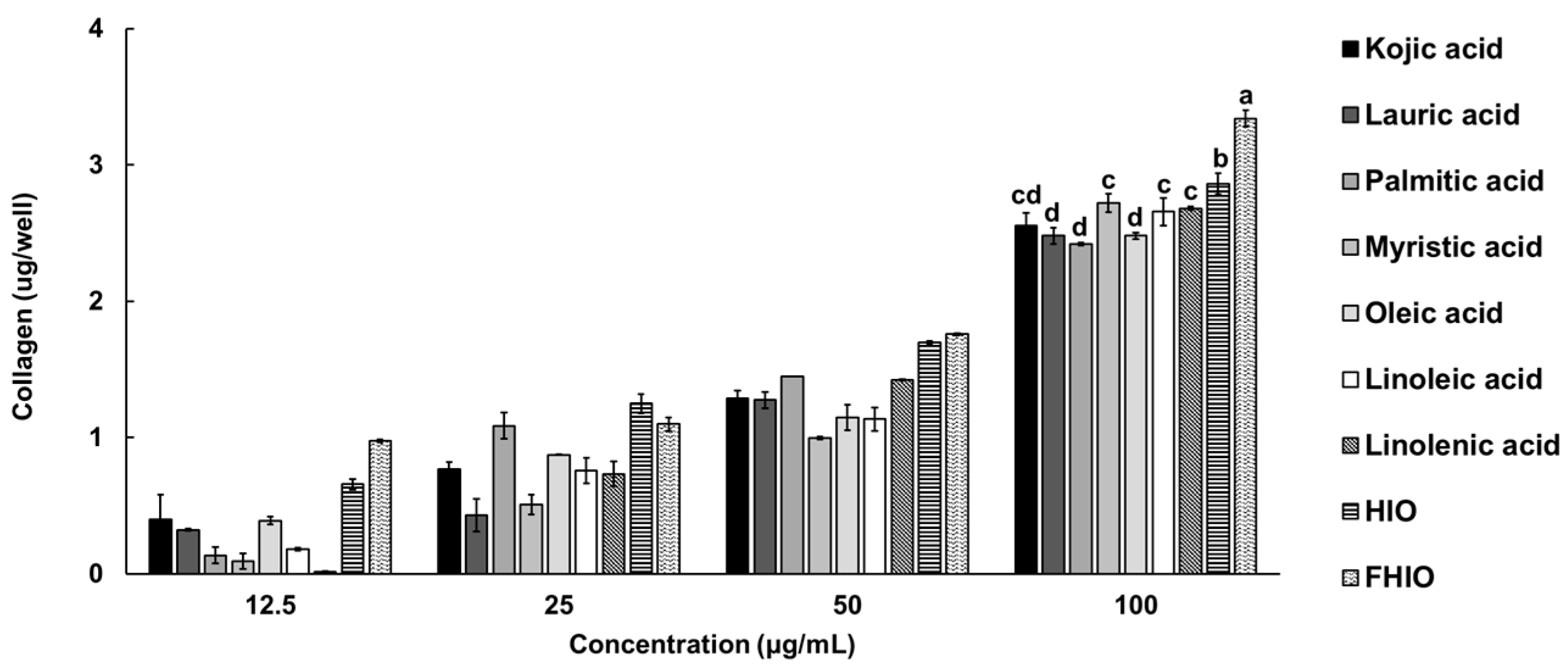
2.5. Evaluation of the Skin Wrinkle Improvement Potential of Hermetia illucens Oil and Fermented Hermetia illucens Oil in Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Hermetia illucens Oil, Microbial Strains, Culture Conditions, and Fermentation Efficacy Assays of Different Strains
4.2. Analysis of Fermentation Efficacy According to Fermentation Period and Fermentation Method for Hermetia illucens Oil
4.3. Cell Culture and Cell Viability Analysis for a Selection of HIO Solvents and HIO Treatment Concentrations
4.4. Analysis of the Skin-Whitening Effects of HIO and FHIO
4.4.1. In Vitro Tyrosinase Inhibition Rate
4.4.2. Tyrosinase Inhibition Rate and mRNA Expression Level in SK-MEL-2 Cells
4.4.3. Melanin Inhibition Rate in SK-MEL-2 Cells
4.5. Analysis of Skin Wrinkle Improvement Effect of HIO and FHIO
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bilal, M.; Iqbal, H.M.N. New insights on unique features and role of nanostructured materials in cosmetics. Cosmetics 2020, 7, 24. [Google Scholar] [CrossRef]
- Dini, I.; Laneri, S. Nutricosmetics: A brief overview. Phytother. Res. 2019, 33, 3054–3063. [Google Scholar] [CrossRef] [PubMed]
- Arda, O.; Göksügür, N.; Tüzün, Y. Basic histological structure and functions of facial skin. Clin. Dermatol. 2014, 32, 3–13. [Google Scholar] [CrossRef] [PubMed]
- De Macedo, G.M.C.; Nunes, S.; Barreto, T. Skin disorders in diabetes mellitus: An epidemiology and physiopathology review. Diabetol. Metab. Syndr. 2016, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- DiBaise, M.; Tarleton, S.M. Hair, Nails, and Skin: Differentiating Cutaneous Manifestations of Micronutrient Deficiency. Nutr. Clin. Pract. 2019, 34, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Manson Brown, S.; Cross, S.J.; Mehta, R. Defining Skin Quality: Clinical Relevance, Terminology, and Assessment. Dermatol. Surg. 2021, 47, 974–981. [Google Scholar] [CrossRef]
- Costa, E.F.; Magalhães, W.V.; Di Stasi, L.C. Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications. Molecules 2022, 27, 7518. [Google Scholar] [CrossRef]
- Anderson, J.; Anderson, Y.; Diyabalanage, T. Paving the way with actives for skincare. Planta Med. 2015, 8, 23. [Google Scholar] [CrossRef]
- Mir, S.A.; Dar, B.N.; Wani, A.B.; Shah, M.A. Effect of plant extracts on the techno-functional properties of biodegradable packaging films. Trends Food Sci. Technol. 2018, 80, 141–154. [Google Scholar] [CrossRef]
- Ling, X.; Pan, Z.; Zhang, H.; Wu, M.; Gui, Z.; Yuan, Q.; Chen, J.; Peng, J.; Liu, Z.; Tan, Q.; et al. PARP-1 modulates the expression of miR-223 through histone acetylation to involve in the hydroquinone-induced carcinogenesis of TK6 cells. J. Biochem. Mol. Toxicol. 2022, 36, e23142. [Google Scholar] [CrossRef]
- Ahmed, I.A.; Mikail, M.A.; Zamakshshari, N.; Abdullah, A.H. Natural anti-aging skincare: Role and potential. Biogerontology 2020, 21, 293–310. [Google Scholar] [CrossRef]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential Oils as Natural Sources of Fragrance Compounds for Cosmetics and Cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive Compounds from Natural Resources against Skin Aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.J.; Pedrosa, S.S.; Mendes, A.; Azevedo-Silva, J.; Fernandes, J.; Pintado, M.; Oliveira, A.L.S.; Madureira, A.R. Anti-Aging Potential of a Novel Ingredient Derived from Sugarcane Straw Extract (SSE). Int. J. Mol. Sci. 2023, 25, 21. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.J. The Implications of the Nagoya Protocol for the Ethical Sourcing of Biodiversity; Brill Nijhoff: Leiden, The Netherlands, 2013; pp. 369–387. [Google Scholar]
- Atef, M.; Ojagh, S.M.; Latifi, A.M.; Esmaeili, M.; Udenigwe, C.C. Biochemical and structural characterization of sturgeon fish skin collagen (Huso huso). J. Food Biochem. 2020, 44, e13256. [Google Scholar] [CrossRef] [PubMed]
- Maeng, Y.; Kim, K.T.; Zhou, X.; Jin, L.; Kim, K.S.; Kim, Y.H.; Lee, S.; Park, J.H.; Chen, X.; Kong, M.; et al. A novel microbial technique for producing high-quality sophorolipids from horse oil suitable for cosmetic applications. Microb. Biotechnol. 2018, 11, 917–929. [Google Scholar] [CrossRef]
- Rabasco Álvarez, A.M.; González Rodríguez, M.L. Lipids in pharmaceutical and cosmetic preparations. Grasas. Aceites 2000, 51, 74–96. [Google Scholar] [CrossRef]
- Lee, C.; Eom, Y.A.; Yang, H.; Jang, M.; Jung, S.U.; Park, Y.O.; Lee, S.E.; Jung, H. Skin Barrier Restoration and Moisturization Using Horse Oil-Loaded Dissolving Microneedle Patches. Skin Pharmaco. Physiol. 2018, 31, 163–171. [Google Scholar] [CrossRef]
- Almeida, C.; Murta, D.; Nunes, R.; Baby, A.R.; Fernandes, Â.; Barros, L.; Rijo, P.; Rosado, C. Characterization of lipid extracts from the Hermetia illucens larvae and their bioactivities for potential use as pharmaceutical and cosmetic ingredients. Heliyon 2022, 8, e09455. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Meijer, N.; Nijkamp, M.M.; Schmitt, E.; Van Loon, J.J.A. Chemical food safety of using former foodstuffs for rearing black soldier fly larvae (Hermetia illucens) for feed and food use. J. Insects Food Feed 2020, 6, 475–488. [Google Scholar] [CrossRef]
- Franco, A.; Salvia, R.; Scieuzo, C.; Schmitt, E.; Russo, A.; Falabella, P. Lipids from insects in cosmetics and for personal care products. Insects 2021, 13, 41. [Google Scholar] [CrossRef]
- Ushakova, N.A.; Brodskii, E.S.; Kovalenko, A.A.; Bastrakov, A.I.; Kozlova, A.A.; Pavlov, D.S. Characteristics of lipid fractions of larvae of the black soldier fly Hermetia illucens. Dokl. Biochem. Biophys. 2016, 468, 209–212. [Google Scholar] [CrossRef]
- Almeida, C.; Murta, D.; Nunes, R.; Baby, A.R.; Rijo, P.; Rosado, C. Preliminary evaluation of the antimicrobial activity of different Hermetia illucens larvae extracts for application as a cosmetic ingredient. J. Biomed. Biopharmaceut. Res. 2020, 17, 327–336. [Google Scholar] [CrossRef]
- Park, J.Y.; Jung, S.; Na, Y.G.; Jeon, C.H.; Cheon, H.Y.; Yoon, E.Y.; Lee, S.H.; Kwon, E.E.; Kim, J.K. Biodiesel production from the black soldier fly larvae grown on food waste and its fuel property characterization as a potential transportation fuel. Environ. Eng. Res. 2022, 27, 200704. [Google Scholar] [CrossRef]
- Erskine, E.; Ozkan, G.; Lu, B.; Capanoglu, E. Effects of fermentation process on the antioxidant capacity of fruit byproducts. ACS Omega 2023, 8, 4543–4553. [Google Scholar] [CrossRef]
- Mitri, S.; Koubaa, M.; Maroun, R.G.; Rossignol, T.; Nicaud, J.M.; Louka, N. Bioproduction of 2-phenylethanol through yeast fermentation on synthetic media and on agro-industrial waste and by-broducts: A review. Foods 2022, 11, 109. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, J.J. The melanocyte and the epidermal melanin unit: An expanded concept. Dermatol. Clin. 2007, 25, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Ngoc, L.T.N.; Tran, V.V.; Moon, J.-Y.; Chae, M.; Park, D.; Lee, Y.-C. Recent trends of sunscreen cosmetic: An update review. Cosmetics 2019, 6, 64. [Google Scholar] [CrossRef]
- Solano, F.; García-Borrón, J.C. Enzymology of melanin formation. In The Pigmentary System: Physiology and Pathophysiology; Blackwell Pub. Ltd.: Oxford, UK, 2006; pp. 261–281. [Google Scholar]
- Solano, F. Photoprotection and skin pigmentation: Melanin-related molecules and some other new agents obtained from natural sources. Molecules 2020, 5, 137. [Google Scholar] [CrossRef] [PubMed]
- Suto, M.; Masutomi, H.; Ishihara, K.; Masaki, H. A potato peel extract stimulates type I collagen synthesis via Akt and ERK signaling in normal human dermal fibroblasts. Biol. Pharm. Bull. 2019, 42, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Pientaweeratch, S.; Panapisal, V.; Tansirikongkol, A. Antioxidant, anti-collagenase and anti-elastase activities of Phyllanthus emblica, Manilkara zapota and sylimarin: An in vitro comparative study for anti-aging applications. Pharmac. Biol. 2016, 54, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Kim, J. Synthesis and biological evaluation of the anti-melanogenesis effect of coumaric and caffeic acid-conjugated peptides in human melanocytes. Front. Phamacol. 2020, 11, 922. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content (%) | |
|---|---|
| Capric acid (C10:0) | 0.7 |
| Lauric acid (C12:0) | 23.6 |
| Myristic acid (C14:0) | 6 |
| Myristoleic acid (C14:1) | 0.1 |
| Palmitic acid (C16:0) | 20.3 |
| Palmitoleic acid (C16:1) | 2.7 |
| Margaroleic acid (C17:0) | 0.5 |
| Stearic acid (C18:0) | 4.6 |
| Oleic acid (C18:1) | 23.2 |
| Linoleic acid (C18:2) | 15.9 |
| Linolenic acid (C18:3) | 2.2 |
| Arachidic acid (C20:0) | 0.2 |
| Saturated fatty acids | 55.9 |
| Monounsaturated fatty acid | 26.0 |
| Polyunsaturated fatty acid | 18.1 |
| Microorganisms | Lipase (mU/mL) |
|---|---|
| L. gasseri | 3.879 ± 0.397 a |
| L. pentosus | 3.557 ± 0.031 b |
| L. plantarum | 3.240 ± 0.329 c |
| L. curvatus | 3.135 ± 0.003 c |
| B. subtillis | 2.910 ± 0.048 e |
| P. pseudoalcaligenes | 3.065 ± 0.310 cd |
| P. acidiloctici | 3.037 ± 0.062 cd |
| S. warneri | 3.077 ± 0.021 d |
| P. pentosaceus | 3.122 ± 0.061 c |
| S. cerevisiae | 3.036 ± 0.046 d |
| Free Glycerol (μmol/mL) | Free Fatty Acid (μmol/mL) | |
|---|---|---|
| L. gasseri | 1.330 ± 0.075 a | 2.915 ± 0.175 a |
| L. pentosus | 1.094 ± 0.054 b | 2.396 ± 0.136 cd |
| L. plantarum | 1.297 ± 0.069 a | 2.508 ± 0.145 b |
| L. curvatus | 1.083 ± 0.062 b | 2.378 ± 0.027 c |
| B. subtillis | 1.014 ± 0.009 c | 2.403 ± 0.009 c |
| P. pseudoalcaligenes | 1.063 ± 0.022 b | 2.297 ± 0.021 d |
| P. acidiloctici | 1.047 ± 0.010 c | 2.377 ± 0.021 c |
| S. warneri | 1.077 ± 0.001 b | 2.440 ± 0.001 b |
| P. pentosaceus | 1.019 ± 0.009 d | 2.710 ± 0.013 ab |
| S. cerevisiae | 0.951 ± 0.011 e | 2.365 ± 0.053 c |
| Free Glycerol (μmol/mL) | Free Fatty Acid (μmol/mL) | ||||
|---|---|---|---|---|---|
| Fermentation Method | Strains with Culture Medium | Culture Medium Only | Strains with Culture Medium | Culture Medium Only | |
| Fermentation period (days) | 0 | 2.678 ± 0.030 d | 2.780 ± 0.032 d | 3.138 ± 0.069 d | 3.122 ± 0.071 d |
| 0.5 | 2.705 ± 0.143 d | 2.807 ± 0.040 cd | 3.148 ± 0.287 d | 3.133 ± 0.040 d | |
| 1 | 2.860 ± 0.023 d | 2.864 ± 0.019 cd | 3.170 ± 0.052 d | 3.255 ± 0.019 cd | |
| 2 | 2.795 ± 0.051 cd | 2.889 ± 0.041 cd | 3.211 ± 0.048 d | 3.295 ± 0.041 cd | |
| 3 | 2.863 ± 0.053 cd | 2.967 ± 0.091 cd | 3.307 ± 0.057 cd | 3.292 ± 0.091 cd | |
| 4 | 3.041 ± 0.169 cd | 3.145 ± 0.075 c | 3.429 ± 0.039 c | 3.314 ± 0.074 cd | |
| 5 | 3.387 ± 0.280 cd | 3.294 ± 0.159 c | 3.441 ± 0.048 c | 3.526 ± 0.159 c | |
| 7 | 3.685 ± 0.114 c | 3.990 ± 0.124 b | 3.632 ± 0.027 b | 3.917 ± 0.124 b | |
| 14 | 3.668 ± 0.280 b | 4.071 ± 0.480 ab | 4.215 ± 0.065 a | 4.799 ± 0.480 a | |
| 21 | 3.745 ± 0.114 a | 4.091 ± 0.231 a | 4.240 ± 0.048 a | 4.824 ± 0.231 a | |
| 28 | 3.816 ± 0.098 a | 4.098 ± 0.058 a | 4.251 ± 0.027 a | 4.855 ± 0.058 a | |
| 35 | 3.728 ± 0.120 a | 4.031 ± 0.069 a | 4.214 ± 0.065 a | 4.798 ± 0.069 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.; Goo, T.-W.; Lee, S.H.; Yun, E.-Y. Skin Anti-Aging Potential through Whitening and Wrinkle Improvement Using Fermented Oil Derived from Hermetia illucens Larvae. Int. J. Mol. Sci. 2024, 25, 2736. https://doi.org/10.3390/ijms25052736
Hwang D, Goo T-W, Lee SH, Yun E-Y. Skin Anti-Aging Potential through Whitening and Wrinkle Improvement Using Fermented Oil Derived from Hermetia illucens Larvae. International Journal of Molecular Sciences. 2024; 25(5):2736. https://doi.org/10.3390/ijms25052736
Chicago/Turabian StyleHwang, Dooseon, Tae-Won Goo, Seung Hun Lee, and Eun-Young Yun. 2024. "Skin Anti-Aging Potential through Whitening and Wrinkle Improvement Using Fermented Oil Derived from Hermetia illucens Larvae" International Journal of Molecular Sciences 25, no. 5: 2736. https://doi.org/10.3390/ijms25052736
APA StyleHwang, D., Goo, T.-W., Lee, S. H., & Yun, E.-Y. (2024). Skin Anti-Aging Potential through Whitening and Wrinkle Improvement Using Fermented Oil Derived from Hermetia illucens Larvae. International Journal of Molecular Sciences, 25(5), 2736. https://doi.org/10.3390/ijms25052736