
Impact of Sinapic Acid on Bovine Serum Albumin Thermal Stability



Abstract
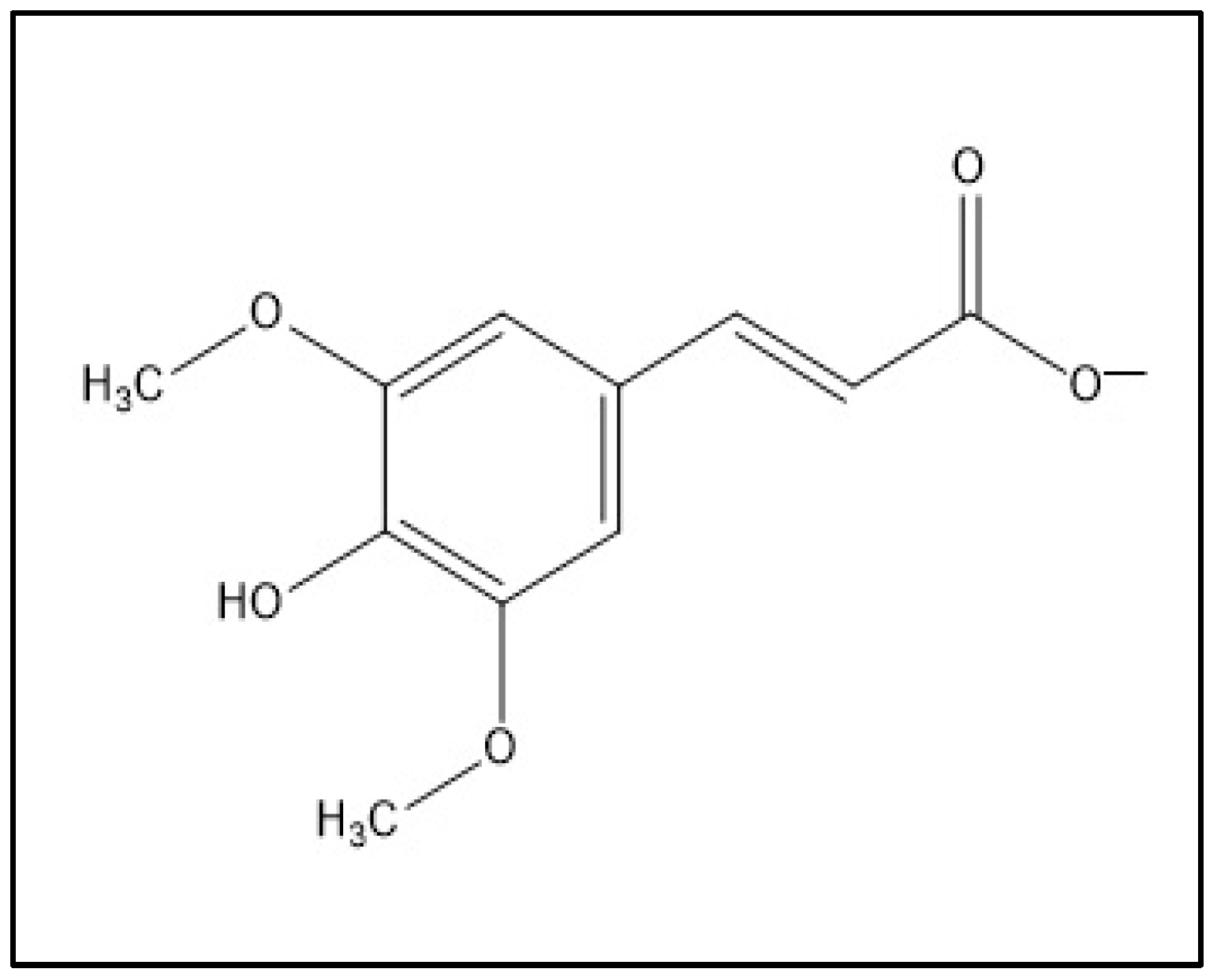
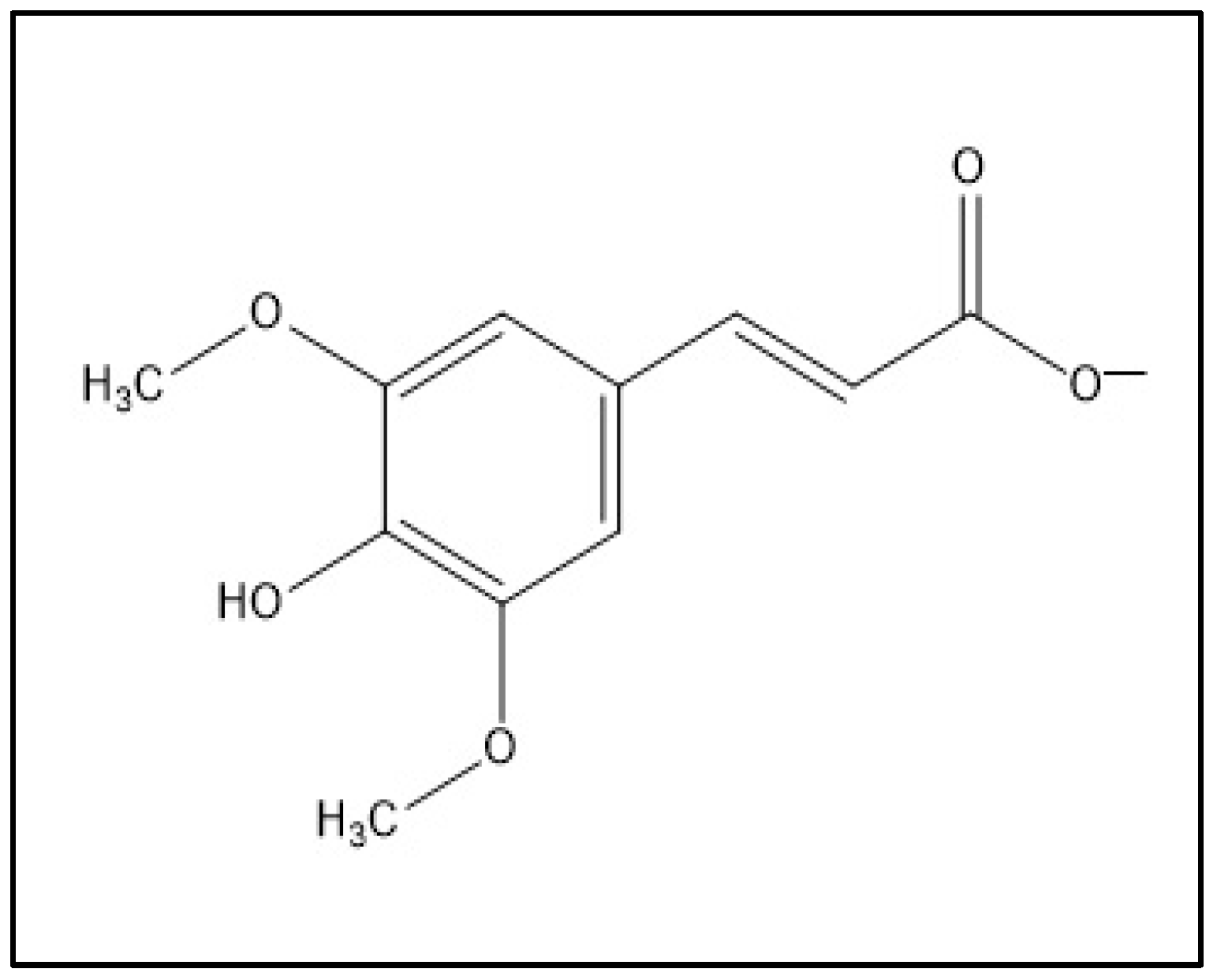
1. Introduction
2. Results and Discussions
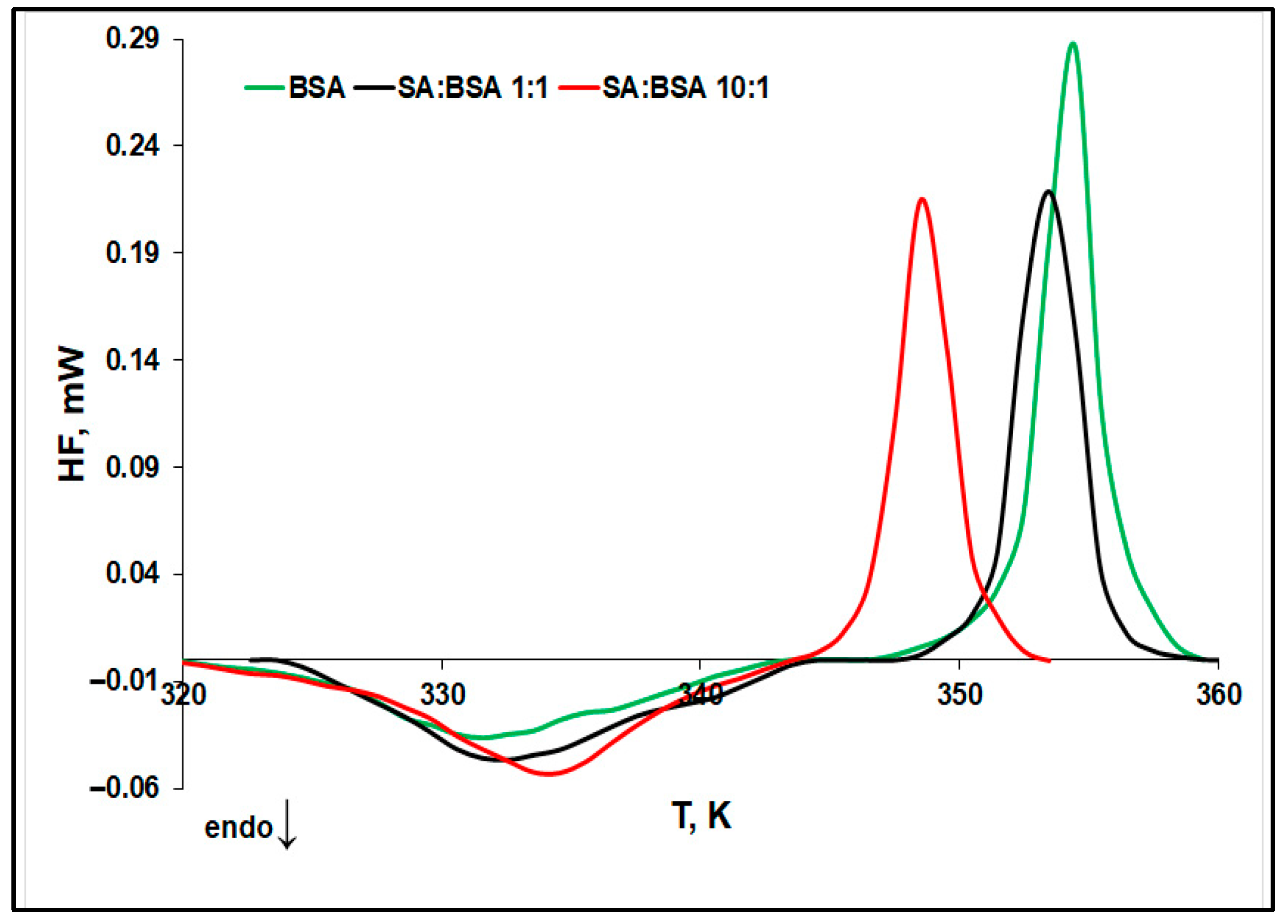
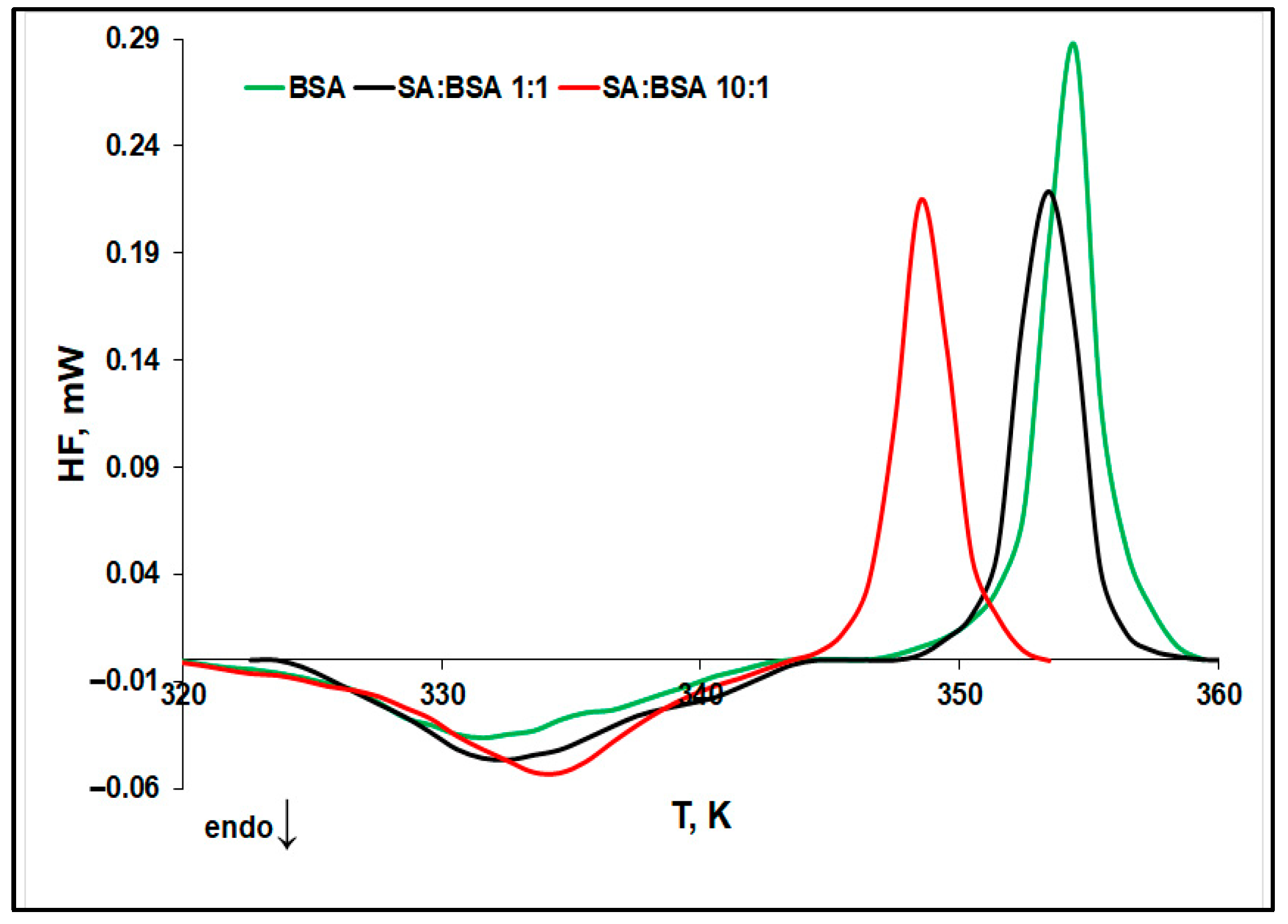
2.1. Evaluation of Protein Thermal Stability by Differential Scanning Microcalorimetry (μDSC)
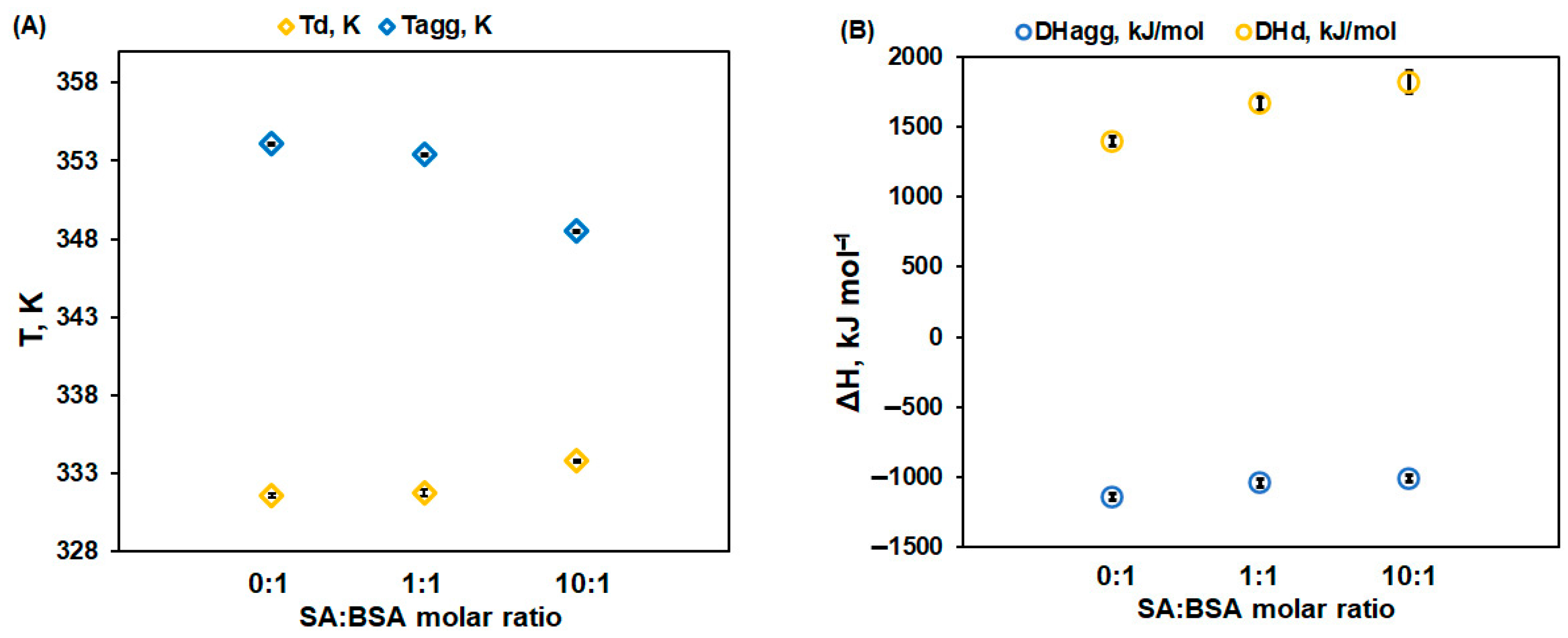
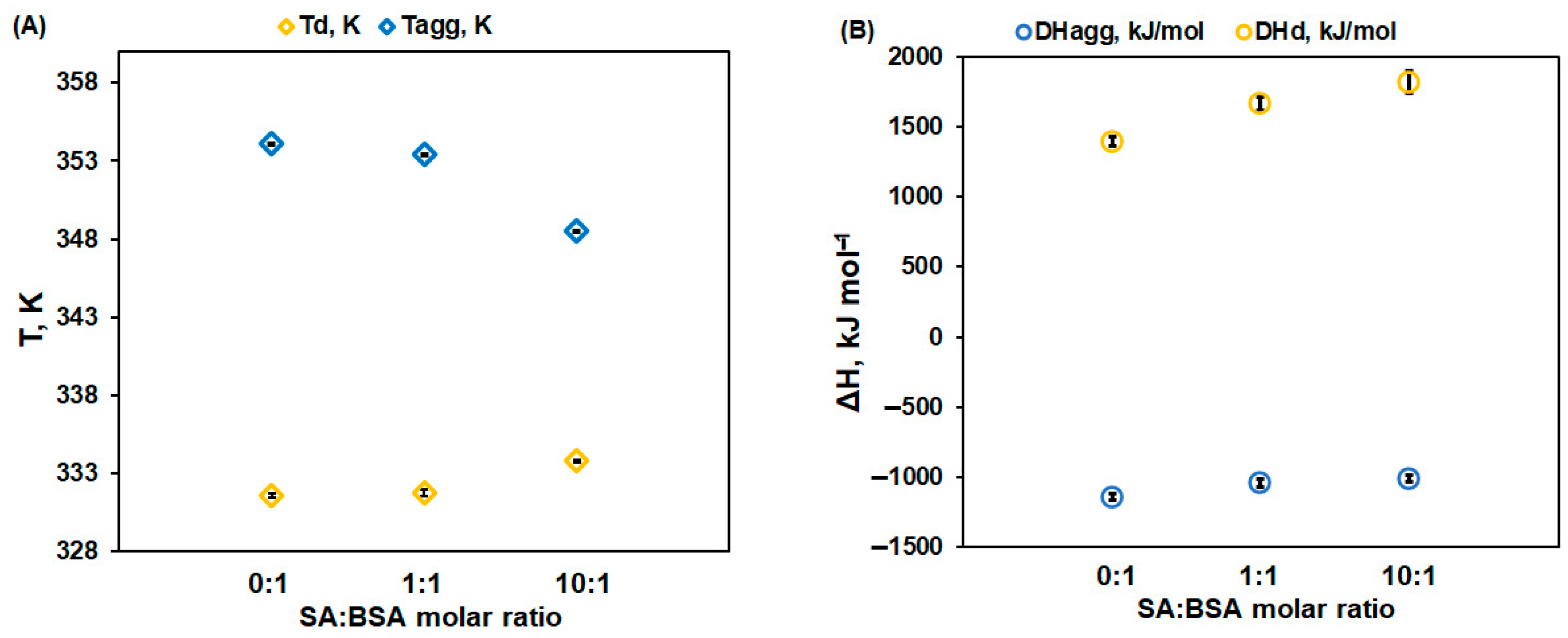
- The denaturation temperature (Td) of protein increases with the SA concentration, pointing to a stabilization effect of SA on the protein native structure. Moreover, Figure 3 displays an increase in the denaturation enthalpy change (ΔHd) with an increasing in the ligand concentration. This may be ascribed to a lower exposure to the solvent molecules of the native protein’s buried hydrophobic regions, during the unfolding process in the presence of SA [32].
- The aggregation temperature (Tagg) and the absolute value of the enthalpy change (|ΔHagg|) decrease with SA concentrations: the ligand binding weakens the unfolded structure of protein. For a 10:1 SA:BSA molar ratio, the denaturation process and the aggregation one slightly overlap, producing a supplementary decrease in the calculated |ΔHagg| value.
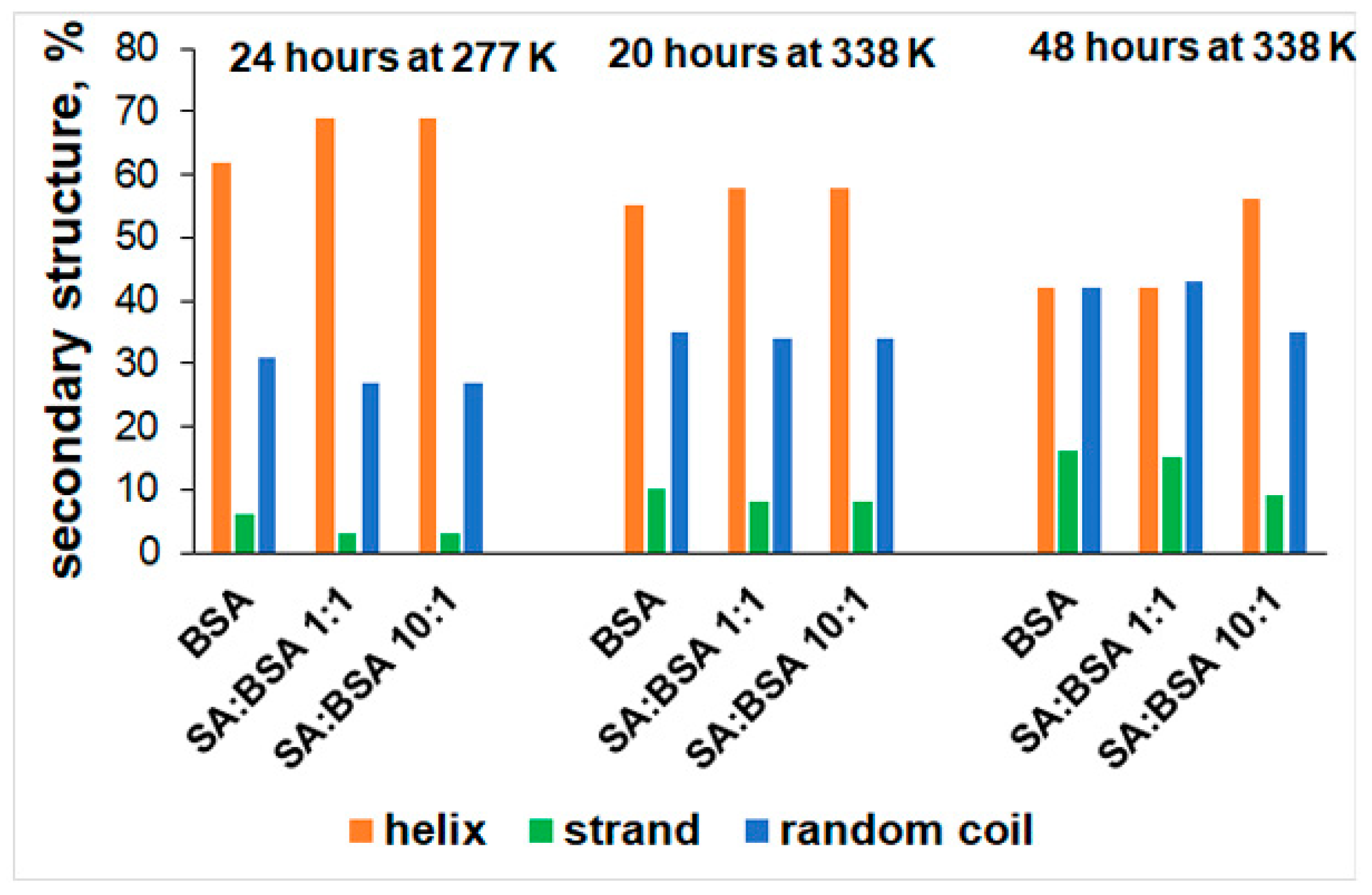
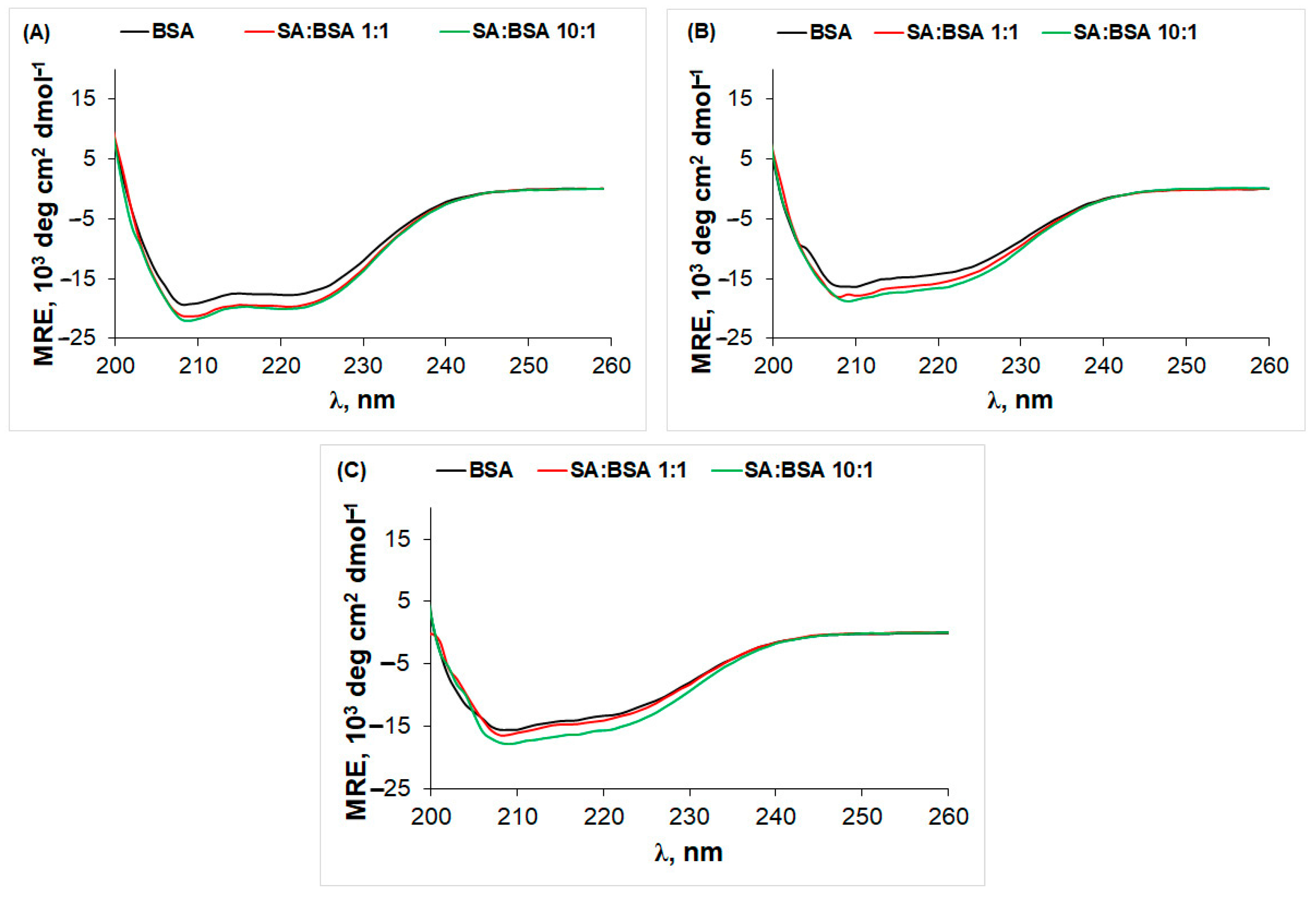
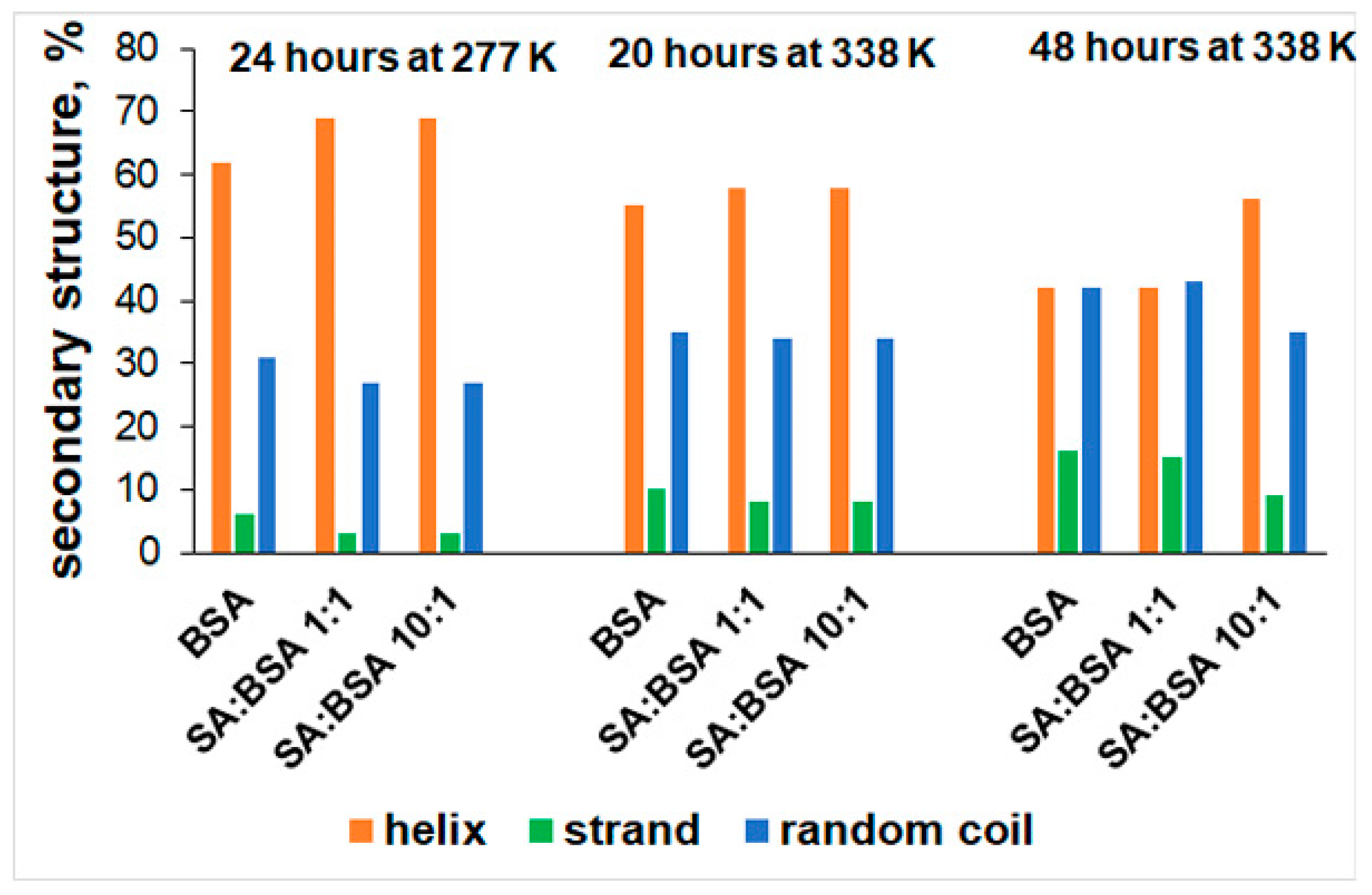
2.2. Circular Dichroism (CD) Spectra
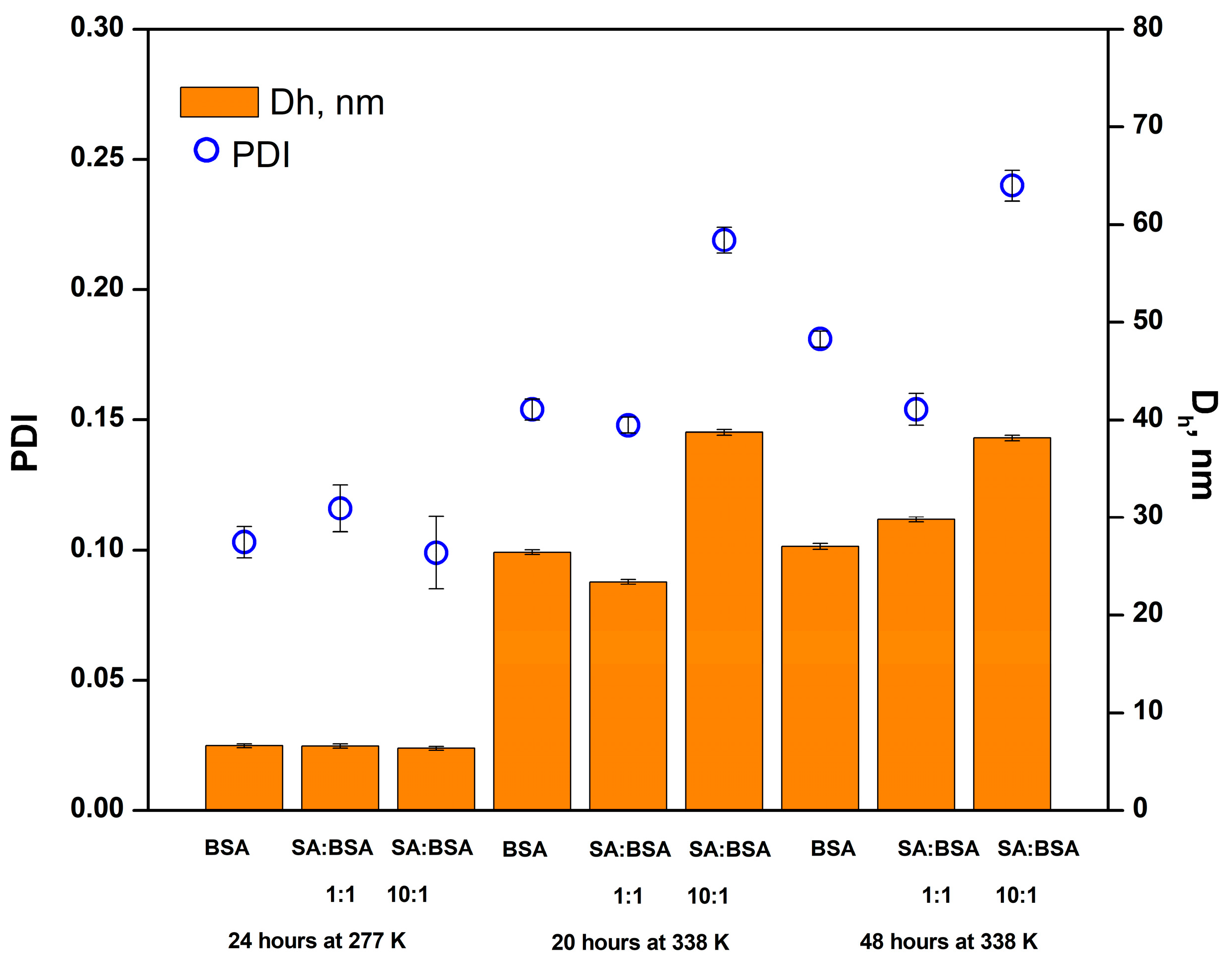
2.3. Changes in the Hydrodynamic Dimeter of BSA Aggregates Investigated by Dynamic Light Scattering (DLS)
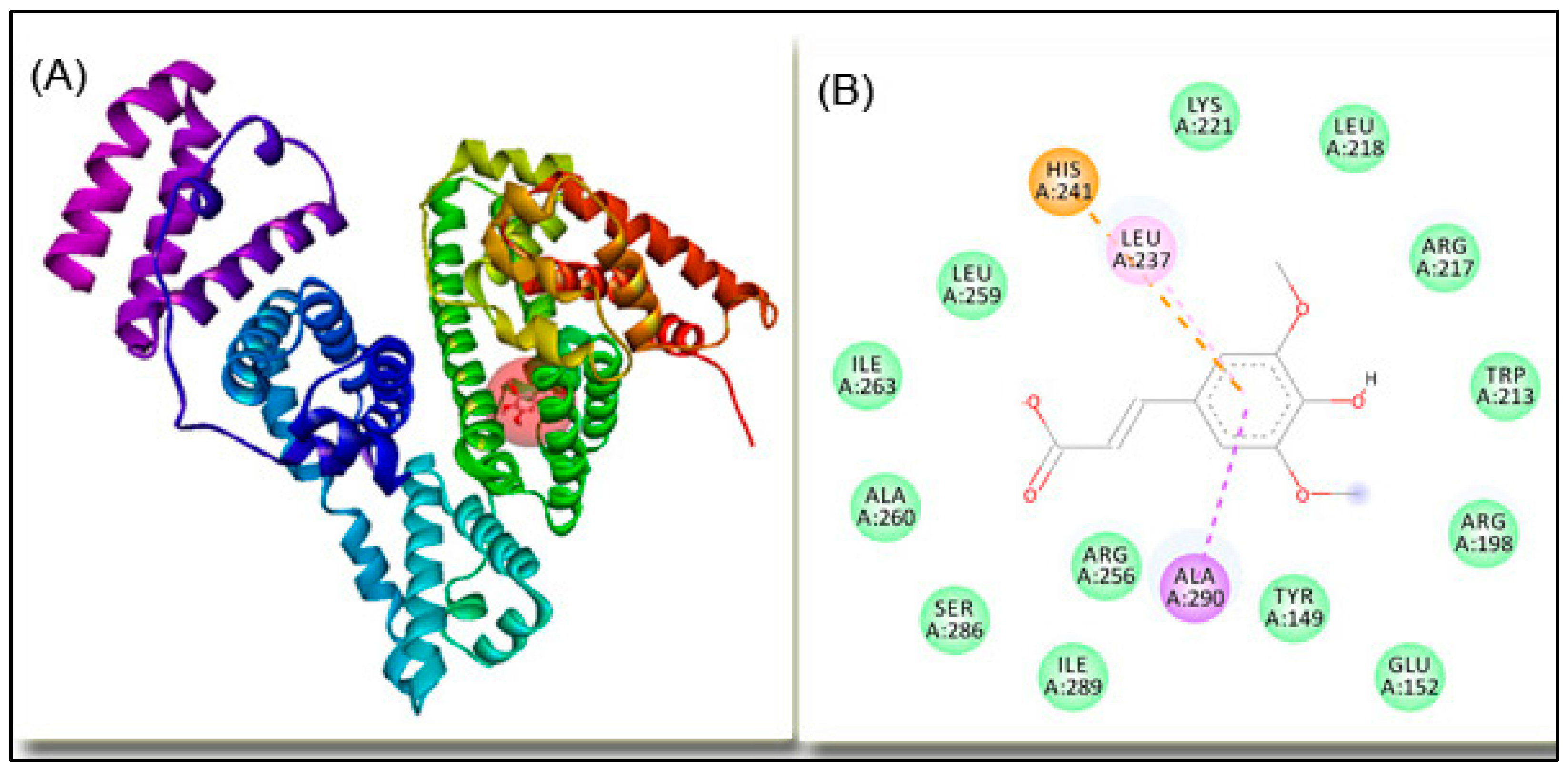
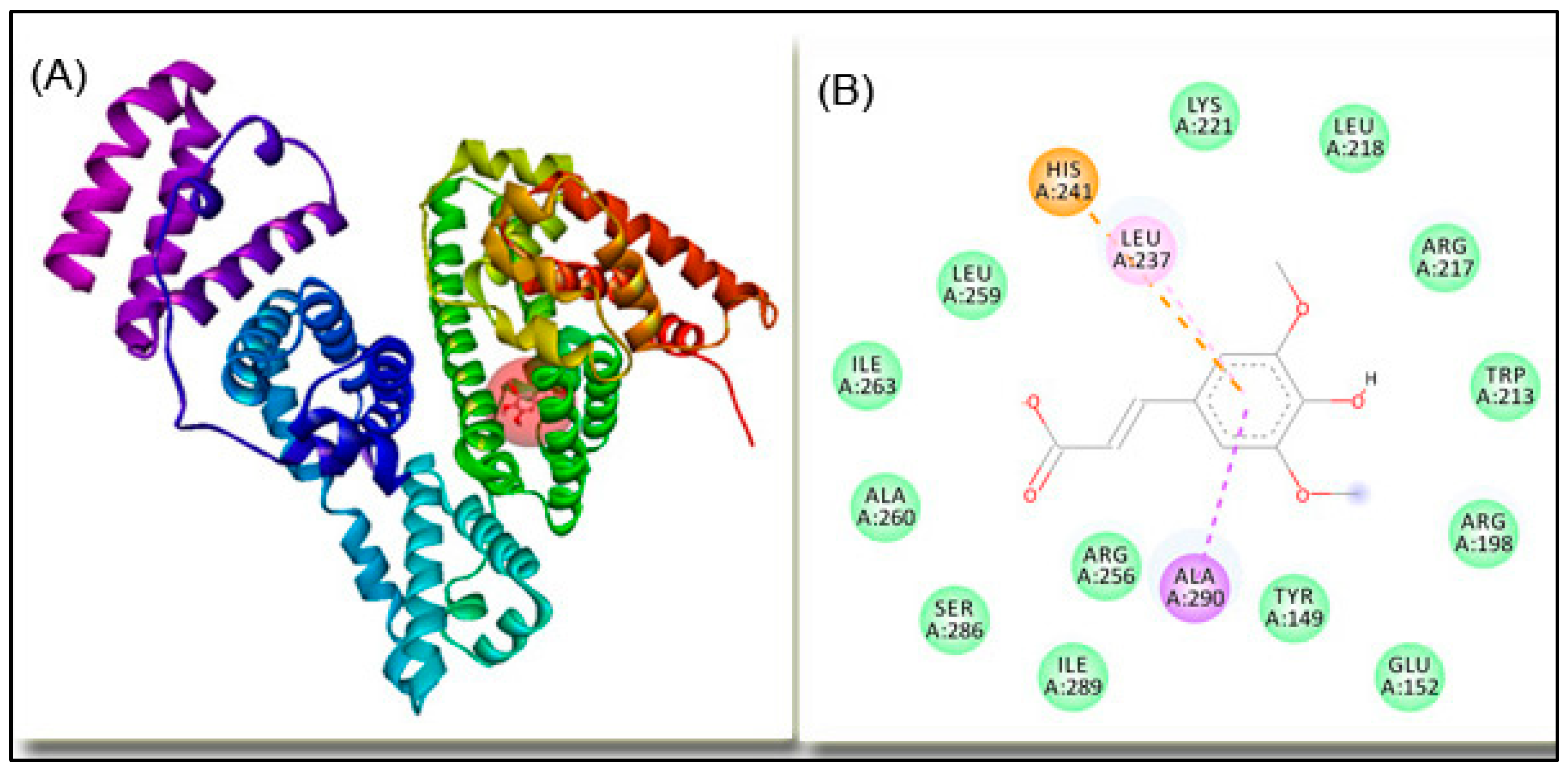
2.4. Molecular Docking
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Differential Scanning Microcalorimetry (μDSC)
3.2.2. Circular Dichroism (CD)
3.2.3. Dynamic Light Scattering (DLS)
3.2.4. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreno, D.A.; Pérez-Balibrea, S.; Ferreres, F.; Gil-Izquierdo, A.; García-Viguera, C. Acylated anthocyanins in broccoli sprouts. J. Food Chem. 2010, 123, 358–363. [Google Scholar] [CrossRef]
- Andreasen, M.F.; Landbo, A.-K.; Christensen, L.P.; Hansen, A.; Meyer, A.S. Antioxidant effects of phenolic rye (Secale cereale L.) extracts, monomeric hydroxycinnamates, and ferulic acid dehydrodimers on human low-density lipoproteins. J. Agric. Food Chem. 2001, 49, 4090–4096. [Google Scholar] [CrossRef]
- Niciforovic, N.; Abramovic, H. Sinapic acid and its derivatives: Natural sources and bioactivity. Compr. Rev. Food Sci. Food Saf. 2014, 13, 34–51. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Schieber, A.; Gänzle, M.G. Sinapic acid derivatives in defatted oriental mustard (Brassica juncea L.) seed meal extracts using UHPLC-DADESI-MSn and identification of compounds with antibacterial activity. Eur. Food Res. Technol. 2012, 234, 535–542. [Google Scholar] [CrossRef]
- Yun, K.J.; Koh, D.J.; Kim, S.H.; Park, S.J.; Ryu, J.H.; Kim, D.G.; Lee, J.Y.; Lee, K.T. Anti-inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expressions via nuclear factor-kappaB inactivation. J. Agric. Food Chem. 2008, 56, 10265–10272. [Google Scholar] [CrossRef] [PubMed]
- Cherng, Y.G.; Tsai, C.C.; Chung, H.H.; Lai, Y.W.; Kuo, S.C.; Cheng, J.T. Antihyperglycemic action of sinapic acid in diabetic rats. J. Agric. Food Chem. 2013, 61, 12053–12059. [Google Scholar] [CrossRef] [PubMed]
- Hudson, E.A.; Dinh, P.A.; Kokubun, T.; Simmonds, M.S.J.; Gescher, A. Characterization of potentially hemopreventive phenols in extracts of brown rice that inhibit the growth of human breast and colon cancer cells. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1163–1170. [Google Scholar]
- Kikuzaki, H.; Hisamoto, M.; Hirose, K.; Akiyama, K.; Taniguchi, H. Antioxidant properties of ferulic acid and its related compounds. J. Agric. Food Chem. 2002, 50, 2161–2168. [Google Scholar] [CrossRef]
- Maddox, C.E.; Laur, L.M.; Tian, L. Antibacterial activity of phenolic compounds against the phytopathogen Xylella fastidiosa. Curr. Microbiol. 2010, 60, 53–58. [Google Scholar] [CrossRef]
- Ansari, M.A.; Raish, M.; Ahmad, A.; Ahmad, S.F.; Mudassar, S.; Mohsin, K.; Shakeel, F.; Korashy, H.M.; Bakheet, S.A. Sinapic acid mitigates gentamicin-induced nephrotoxicity and associated oxidative/nitrosative stress, apoptosis, and inflammation in rats. Life Sci. 2016, 165, 1–8. [Google Scholar] [CrossRef]
- Sun, X.L.; Ito, H.; Masuoka, T.; Kamei, C.; Hatano, T. Effect of Polygala tenuifolia root extract on scopolamine-induced impairment of rat spatial cognition in an eight-arm radial maze task. Biol. Pharm. Bull. 2007, 30, 1727–1731. [Google Scholar] [CrossRef] [PubMed]
- Raish, M.; Ahmad, A.; Ansari, M.A.; Alkharfy, K.M.; Ahad, A.; Khan, A.; Aljenobi, F.I.; Ali, N.; Al-Mohizea, A.M. Effect of sinapic acid on aripiprazole pharmacokinetics in rats: Possible food drug interaction. J. Food Drug Anal. 2019, 27, 332–338. [Google Scholar] [CrossRef]
- Smyk, B. Fluorescence study of sinapic acid interaction with bovine serum albumin and egg albumin. J. Fluoresc. 2003, 13, 349–356. [Google Scholar] [CrossRef]
- Otagiri, M.A. Molecular functional study on the interactions of drugs with plasma proteins. Drug Metabol. Pharmacokinet. 2005, 20, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Sudlow, G.; Birkett, D.J.; Wade, D.N. The Characterization of Two Specific Drug Binding Sites on Human Serum Albumin. Mol. Pharmacol. 1975, 11, 824–832. [Google Scholar]
- Nnyigide, O.S.; Hyun, K. The protection of bovine serum albumin against thermal denaturation and gelation by sodium dodecyl sulfate studied by rheology and molecular dynamics simulation. Food Hydrocoll. 2020, 103, 105656. [Google Scholar] [CrossRef]
- Tipping, K.W.; van Oosten-Hawle, P.; Hewitt, E.W.; Radford, S.E. Amyloid fibres: Inert end-stage aggregates or key players in disease? Trends Biochem. Sci. 2015, 40, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, B.V.; Janczy, J.R.; Hatlevik, Ø.; Wolfson, G.; Anseth, K.S.; Tibbitt, M.W. Thermal stabilization of biologics with photoresponsive hydrogels. Biomacromolecules 2018, 19, 740–747. [Google Scholar] [CrossRef]
- Aricov, L.; Precupas, A.; Tudose, M.; Baltag, D.; Trică, B.; Sandu, R.; Leonties, A.R. Trametes versicolor laccase activity modulated by the interaction with gold nanoparticles. Environ. Res. 2023, 237, 116920. [Google Scholar] [CrossRef]
- Banu, A.; Khan, R.H.; Qashqoosh, M.T.A.; Manea, Y.K.; Furkan, M.; Naqvi, S. Multispectroscopic and computational studies of interaction of bovine serum albumin, human serum albumin and bovine hemoglobin with bisacodyl. J. Mol. Struct. 2022, 1249, 131550. [Google Scholar] [CrossRef]
- Dombi, G.; Horváth, P.; Fiser, B.; Mirzahosseini, A.; Dobó, M.; Szabó, Z.-I.; Tóth, G. Enantioselective Human Serum Albumin Binding of Apremilast: Liquid Chromatographic, Fluorescence and Molecular Docking Study. Int. J. Mol. Sci. 2023, 24, 2168. [Google Scholar] [CrossRef] [PubMed]
- Toma, C.-M.; Imre, S.; Farczadi, L.; Ion, V.; Marc, G. Enantioselective binding of carvedilol to human serum albumin and alpha-1-acid glycoprotein. Chirality 2023, 35, 779–792. [Google Scholar] [CrossRef]
- Sengupta, P.; Pal, U.; Mondal, P.; Bose, A. Multi-spectroscopic and computational evaluation on the binding of sinapic acid and its Cu(II) complex with bovine serum albumin. Food Chem. 2019, 301, 125254. [Google Scholar] [CrossRef]
- Barone, G.; Giancola, C.; Verdoliva, A. DSC studies on the denaturation and aggregation of serum albumins. Thermochim. Acta 1992, 199, 197–205. [Google Scholar] [CrossRef]
- Precupas, A.; Sandu, R.; Neculae, A.V.F.; Neacsu, A.; Popa, V.T. Calorimetric, spectroscopic and computational investigation of morin binding effect on bovine serum albumin stability. J. Mol. Liq. 2021, 333, 115953. [Google Scholar] [CrossRef]
- Precupas, A.; Sandu, R.; Leonties, A.R.; Anghel, D.F.; Popa, V.T. Complex interaction of caffeic acid with bovine serum albumin: Calorimetric, spectroscopic and molecular docking evidence. New J. Chem. 2017, 41, 15003–15015. [Google Scholar] [CrossRef]
- Precupas, A.; Leonties, A.R.; Neacsu, A.; Sandu, R.; Popa, V.T. Gallic acid influence on bovine serum albumin thermal stability. New J. Chem. 2019, 43, 3891–3898. [Google Scholar] [CrossRef]
- Borzova, V.A.; Markossian, K.A.; Chebotareva, N.A.; Kleymenov, S.Y.; Poliansky, N.B.; Muranov, K.O.; Stein-Margolina, V.A.; Shubin, V.V.; Markov, D.I.; Kurganov, B.I. Kinetics of Thermal Denaturation and Aggregation of Bovine Serum Albumin. PLoS ONE 2016, 11, e0153495. [Google Scholar] [CrossRef]
- Satish, L.; Millan, S.; Das, S.; Jena, S.; Sahoo, H. Thermal Aggregation of Bovine Serum Albumin in Conventional Buffers: An Insight into Molecular Level Interactions. J. Solution Chem. 2017, 46, 831–848. [Google Scholar] [CrossRef]
- Samuelsen, L.; Holm, R.; Lathuile, A.; Schönbeck. Buffer solutions in drug formulation and processing: How pKa values depend on temperature, pressure and ionic strength. Int. J. Pharm. 2019, 560, 357–364. [Google Scholar] [CrossRef]
- Shrake, A.; Ross, P.D. Origins and consequences of ligand induced multiphasic thermal protein denaturation. Biopolymers 1992, 32, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Nishi, K.; Anraku, M.; Taguchi, K.; Maruyama, T.; Otagiri, M. Metal-catalyzed oxidation of human serum albumin does not alter the interactive binding to the two principal drug binding sites. Biochem. Biophys. Rep. 2018, 14, 155–160. [Google Scholar] [CrossRef]
- Preisz, Z.; Kunsági-Máté, S. Effect of methotrexate and its photodegradation products on the temperature induced denaturation of human serum albumin. Spectrochim. Acta A 2021, 245, 118905. [Google Scholar] [CrossRef] [PubMed]
- Lohner, K.; Sen, A.C.; Prankerd, R.; Esser, A.F.; Perrin, J.H. Effects of drug-binding on the thermal denaturation of human serum albumin. J. Pharm. Biomed. Anal. 1994, 12, 1501–1505. [Google Scholar] [CrossRef] [PubMed]
- Labieniec, M.; Gabryelak, T. Interactions of tannic acid and its derivatives (ellagic and gallic acid) with calf thymus DNA and bovine serum albumin using spectroscopic method. J. Photochem. Photobiol. B 2006, 82, 72–78. [Google Scholar] [CrossRef]
- Qishu, J.; Wei, Z.; Yanyan, J.; Lijuan, J.; Xiangyang, C.; Bin, L. Study on the interactions between caffeoylquinic acids with Bovine Serum Albumin: Spectroscopy, antioxidant activity, LC-MSn, and molecular docking approach. Front. Chem. 2019, 7, 840. [Google Scholar] [CrossRef]
- Basu, A.; Bhayye, S.; Kundu, S.; Dasa, A.; Mukherjee, A. Andrographolide inhibits human serum albumin fibril formations through site-specific molecular interactions. RSC Adv. 2018, 8, 30717. [Google Scholar] [CrossRef]
- Jachimska, B.; Wasilewska, M.; Adamczyk, Z. Characterization of globular protein solutions by Dynamic Light Scattering, electrophoretic mobility, and viscosity measurements. Langmuir 2008, 24, 6866–6872. [Google Scholar] [CrossRef]
- Khan, J.M.; Chaturvedi, S.K.; Rahman, S.K.; Ishtikhar, M.; Qadeer, A.; Ahmad, E.; Khan, R.H. Protonation favors aggregation of lysozyme with SDS. Soft Matt. 2014, 10, 2591–2599. [Google Scholar] [CrossRef]
- Jin, X.L.; Wei, X.; Qi, F.-M.; Yu, S.-S.; Zhou, B.; Bai, S. Characterization of hydroxycinnamic acid derivatives binding to bovine serum albumin. Org. Biomol. Chem. 2012, 10, 3424–3431. [Google Scholar] [CrossRef]
- Skrt, M.; Benedik, E.; Podlipnik, C.; Poklar Ulrih, N. Interactions of different polyphenols with bovine serum albumin using fluorescence quenching and molecular docking. Food Chem. 2012, 135, 2418–2424. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.A.; Chacón, P.; Merelo, J.J.; Morán, F. Evaluation of secondary structure of proteins from UV circular dichroism using an unsupervised learning neural network. Prot. Eng. 1993, 6, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and Reference Databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Mao, D.; Wachter, E.; Wallace, B.A. Folding of the mitochondrial proton adenosine triphosphatase proteolipid channel in phospholipid vesicles. Biochemistry 1982, 21, 4960–4968. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Wallace, B.A. Methods of Analysis for Circular Dichroism Spectroscopy of Proteins and the DichroWeb Server. In Advances in Biomedical Spectroscopy, Vol. 1, Modern Techniques for Circular Dichroism and Synchrotron Radiation Circular Dichroism Spectroscopy; Wallace, B.A., Janes, R.W., Eds.; IOS Press: Amsterdam, The Netherlands, 2009; pp. 165–182. [Google Scholar]
- Precupas, A.; Leonties, A.R.; Neacsu, A.; Angelescu, D.G.; Popa, V.T. Bovine hemoglobin thermal stability in the presence of naringenin: Calorimetric, spectroscopic and molecular modeling studies. J. Mol. Liq. 2022, 361, 119617. [Google Scholar] [CrossRef]
- Bujacz, A. Structures of bovine, equine and leporine serum albumin. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, D68, 1278–1289. [Google Scholar] [CrossRef]
- RSCB Protein Data Bank. Available online: https://www.rcsb.org/pdb (accessed on 15 March 2023).
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Montgomery, J.A., Jr.; Vreven, T.; Kudin, K.N.; Burant, J.C.; et al. Gaussian 03, Revision C.01; Gaussian, Inc.: Wallingford, CT, USA, 2004. [Google Scholar]
- Varlan, A.; Hillebrand, M. Exploring the capabilities of TDDFT calculations to explain the induced chirality upon a binding process: A simple case, 3-carboxycoumarin. J. Mol. Struct. 2013, 1036, 341–349. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 45–461. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef]
- Dassault Systemes BIOVIA, Discovery Studio Modeling Environment, Release 2019, San Diego, Dassault Systemes. Available online: http://accelrys.com/products/discovery-studio (accessed on 20 March 2023).
- Chong, X.; Sun, L.; Sun, Y.; Chang, L.; Chang, A.K.; Lu, X.; Zhou, X.; Liu, J.; Zhang, B.; Jones, G.W.; et al. Insights into the mechanism of how Morin suppresses amyloid fibrillation of hen egg white lysozyme. Int. J. Biol. Macromol. 2017, 101, 321–325. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino acid Residue | Distance, Å | Type of Interaction |
|---|---|---|
| HIS241 | 4.75 | electrostatic |
| ALA290 | 3.69 | hydrophobic |
| HIS241 | 5.50 | hydrophobic |
| LEU237 | 4.80 | hydrophobic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Precupas, A.; Popa, V.T. Impact of Sinapic Acid on Bovine Serum Albumin Thermal Stability. Int. J. Mol. Sci. 2024, 25, 936. https://doi.org/10.3390/ijms25020936
Precupas A, Popa VT. Impact of Sinapic Acid on Bovine Serum Albumin Thermal Stability. International Journal of Molecular Sciences. 2024; 25(2):936. https://doi.org/10.3390/ijms25020936
Chicago/Turabian StylePrecupas, Aurica, and Vlad Tudor Popa. 2024. "Impact of Sinapic Acid on Bovine Serum Albumin Thermal Stability" International Journal of Molecular Sciences 25, no. 2: 936. https://doi.org/10.3390/ijms25020936
APA StylePrecupas, A., & Popa, V. T. (2024). Impact of Sinapic Acid on Bovine Serum Albumin Thermal Stability. International Journal of Molecular Sciences, 25(2), 936. https://doi.org/10.3390/ijms25020936