
Interplay of Postprandial Triglyceride-Rich Lipoprotein Composition and Adipokines in Obese Adolescents


 , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Baseline Characteristics of Participants
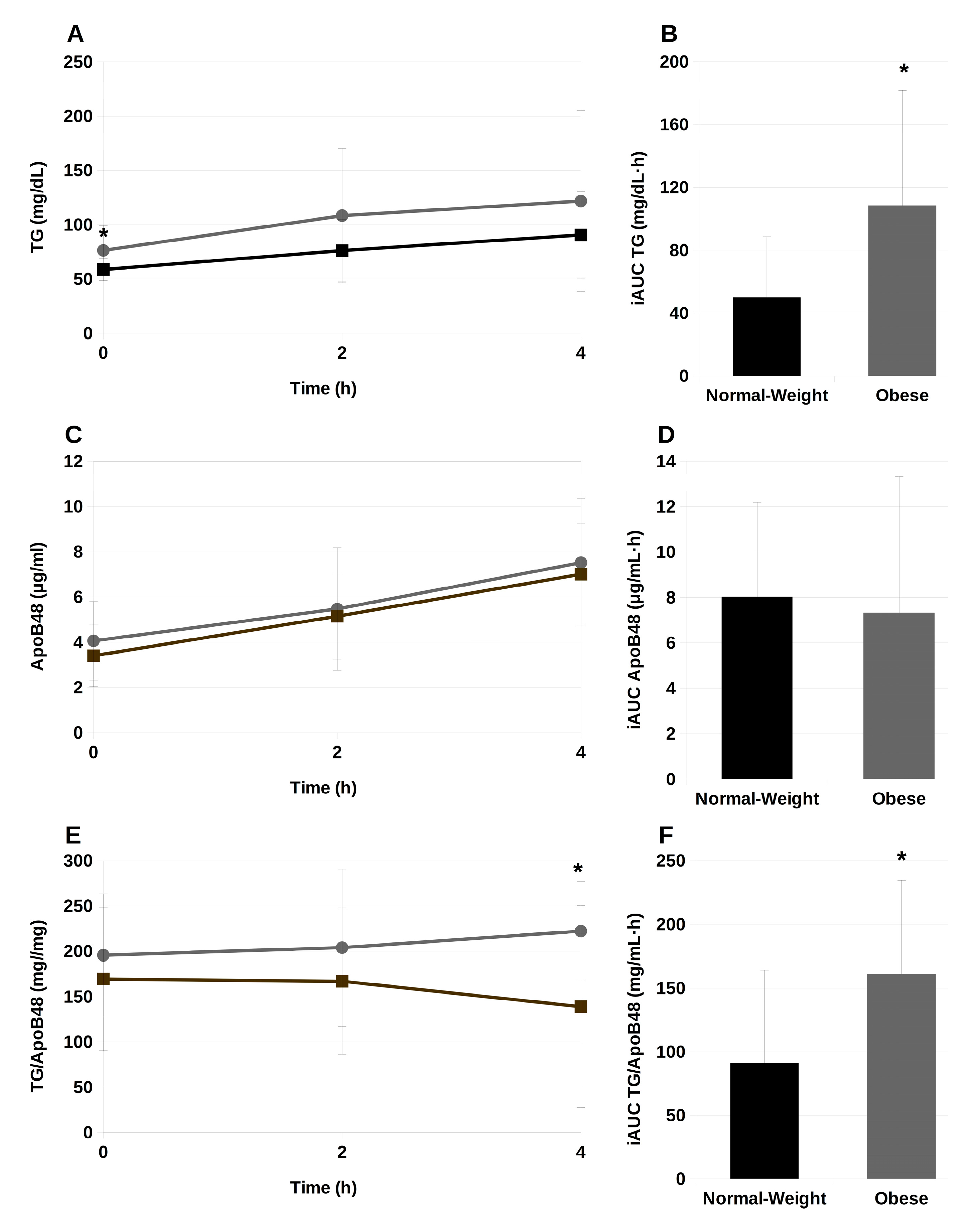
2.2. Serum Triglycerides and ApoB48 during the Postprandial Phase
2.3. Association between Serum TG Concentrations and Biochemical and Anthropometric Parameters
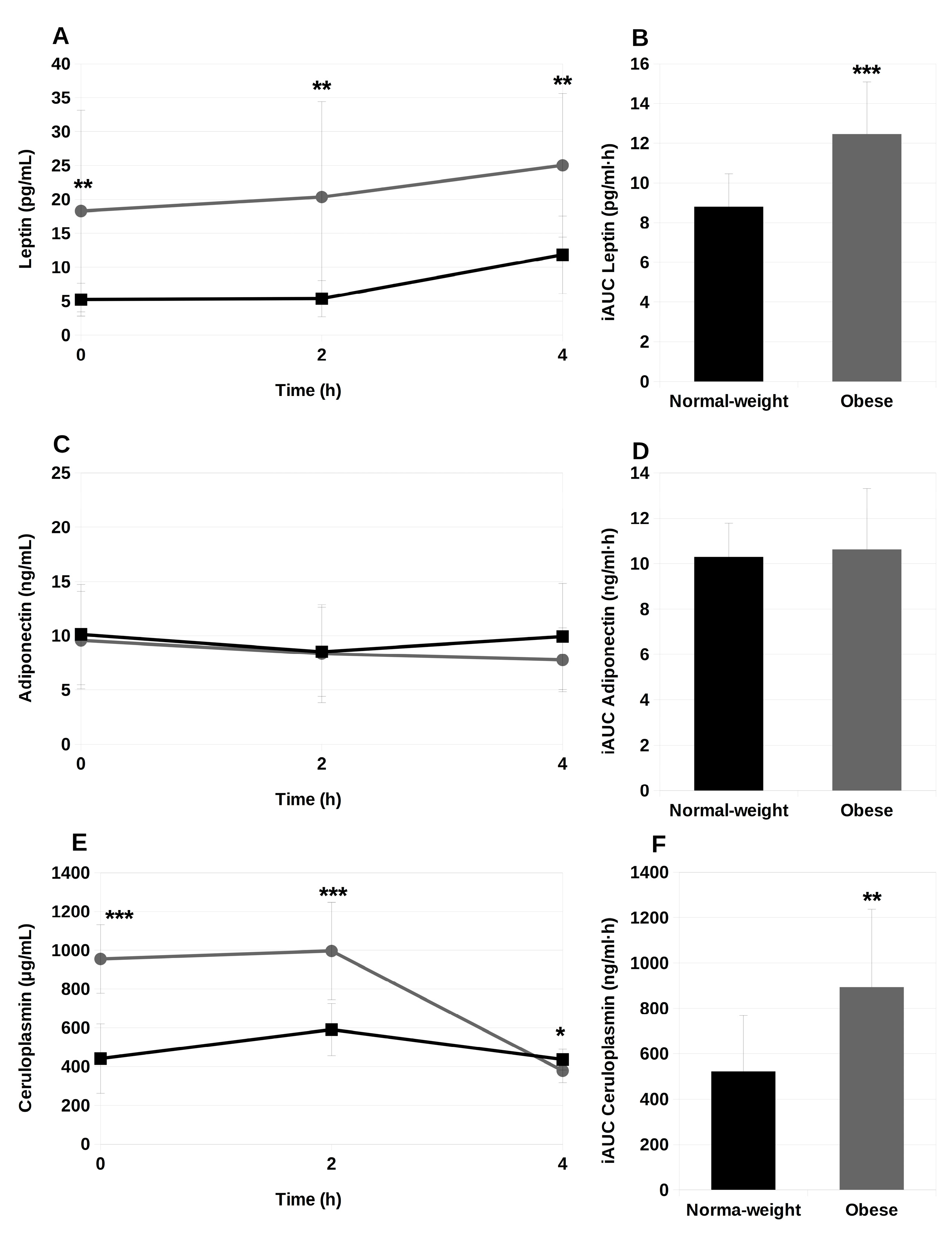
2.4. Circulating Adipokines during the Postprandial Period
2.5. Analysis of Fatty Acids in Triglycerides and Phospholipids Isolated from TRL
3. Discussion
4. Materials and Methods
4.1. Design Overview, Settings and Participants
4.2. Intervention
4.3. Anthropometric Measures of Participants
4.4. Clinical Biochemistry of Participants
4.5. Inflammatory Markers Production
4.6. Isolation and Characterization of Human Triglyceride-Rich Lipoproteins (TRL)
4.7. Lipid Extraction from TRL
4.8. Triglyceride and Phospholipid Fatty Acid Composition
4.9. Analysis of Total ApoB and Apo B48 Content in TRL
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANGPTL3 | angiopoietin-related protein 3 |
| Apo B | Apolipoprotein B |
| Apo CIII | apolipoprotein CIII |
| Apo E | Apolipoprotein E |
| AUC | area under the curve |
| BMI | body mass index |
| CETP | cholesteryl ester transfer protein |
| COX | cyclooxygenase |
| CRP | c-reactive protein |
| DBP | diastolic blood pressure |
| DHA | docosahexaenoic acid |
| FA | fatty acids |
| HbA1c | glycated hemoglobin |
| HDL | high-density lipoprotein |
| HOMA-IR | homeostasic model assessment of insulin resistance |
| iAUC | incremental area under the curve |
| IL | interleukin |
| LDL | low-density lipoprotein |
| LPL | lipoprotein lipase |
| MCP-1 | monocye chemoattractant protein-1 |
| MUFA | monounsaturated fatty acids |
| NO | nitric oxide |
| PGE2 | prostaglandin E2 |
| PL | phospholipids |
| PMA | phorbol 12-myristate 13 acetate |
| PUFA | polyunsaturated fatty acids |
| SBP | systolic blood pressure |
| SFA | saturated fatty acids |
| TG | Triglycerides |
| TNF-α | tumor necrosis factor alpha |
| TRL | Triglyceride-rich lipoprotein |
References
- Vessby, B. Dietary fat, fatty acid composition in plasma and the metabolic syndrome. Curr. Opin. Lipidol. 2003, 14, 15–19. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: https://www.who.int/es/health-topics/obesity#tab=tab_1 (accessed on 3 December 2023).
- Wellen, K.E.; Hotamisligil, G.S. Obesity-induced inflammatory changes in adipose tissue. J. Clin. Investig. 2003, 112, 1785–1788. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Van Oostrom, A.; Wijk, J.; Cabezas, M. Lipaemia, Inflammation and Atherosclerosis: Novel Opportunities in the Understanding and Treatment of Atherosclerosis. Drugs 2004, 64, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Alipour, A.; Elte, J.W.; van Zaanen, H.C.; Rietveld, A.P.; Castro Cabezas, M. Novel aspects of postprandial lipemia in relation to atherosclerosis. Atheroscler. Suppl. 2008, 9, 39–44. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.A.; Klop, B.; Janssen, H.W.; Njo, T.L.; Westerman, E.M.; Castro Cabezas, M. Postprandial inflammation: Targeting glucose and lipids. Adv. Exp. Med. Biol. 2014, 824, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Moruno, R.; Perona, J.S.; Ruiz-Gutierrez, V. Influence of minor components of olive oils on the composition and size of TRLs and on macrophage receptors involved in foam cell formation. Biochem. Soc. Trans. 2007, 35 Pt 3, 470–471. [Google Scholar] [CrossRef][Green Version]
- Moreno, L.A.; Quintela, I.; Fleta, J.; Sarría, A.; Roda, L.; Giner, A.; Bueno, M. Postprandial triglyceridemia in obese and non-obese adolescents. Importance of body composition and fat distribution. J. Pediatr. Endocrinol. Metab. 2001, 14, 193–202. [Google Scholar] [CrossRef]
- Umpaichitra, V.; Banerji, M.A.; Castells, S. Postprandial hyperlipidemia after a fat loading test in minority adolescents with type 2 diabetes mellitus and obesity. J. Pediatr. Endocrinol. Metab. 2004, 17, 853–864. [Google Scholar] [CrossRef]
- Su, J.W.; Ugo Nzekwu, M.M.; Ball, G.D.; Jetha, M.M.; Proctor, S.D. Postprandial lipemia as an early predictor of cardiovascular complications in childhood obesity. J. Clin. Lipidol. 2009, 3, 78–84. [Google Scholar] [CrossRef]
- Aguilar, M.J.; González-Jiménez, E.; Antelo, A.; Perona, J.S. Insulin resistance and inflammation markers: Correlations in obese adolescents. J. Clin. Nurs. 2013, 22, 2002–2010. [Google Scholar] [CrossRef]
- Zilversmit, D.B. Atherogenesis: A Postprandial Phenomenon. Circulation 1979, 60, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Perona, J.S.; Schmidt-RioValle, J.; Chen, Y.; Jing, J.; González-Jiménez, E. Metabolic Syndrome and Its Associated Early-Life Factors among Chinese and Spanish Adolescents: A Pilot Study. Nutrients 2019, 11, 1568. [Google Scholar] [CrossRef]
- Perona, J.S.; Schmidt-RioValle, J.; Fernández-Aparicio, Á.; Correa-Rodríguez, M.; Ramírez-Vélez, R.; González-Jiménez, E. Waist Circumference and Abdominal Volume Index Can Predict Metabolic Syndrome in Adolescents, but only When the Criteria of the International Diabetes Federation are Employed for the Diagnosis. Nutrients 2019, 11, 1370. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A. Triglyceride-Rich Lipoprotein Metabolism: Key Regulators of Their Flux. J. Clin. Med. 2023, 12, 4399. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.J.; Caslake, M.J.; Griffin, B.A.; Hinnie, J.; Tan, C.E.; Watson, T.D.; Packard, C.J.; Shepherd, J. The effect of smoking on post-heparin lipoprotein and hepatic lipase, cholesteryl ester transfer protein and lecithin:cholesterol acyl transferase activities in human plasma. Eur. J. Clin. Investig. 1998, 28, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Kolovou, G.D.; Anagnostopoulou, K.K.; Daskalopoulou, S.S.; Mikhailidis, D.P.; Cokkinos, D.V. Clinical relevance of postprandial lipaemia. Curr. Med. Chem. 2005, 12, 1931–1945. [Google Scholar] [CrossRef] [PubMed]
- Langsted, A.; Nordestgaard, B.G. Nonfasting versus fasting lipid profile for cardiovascular risk prediction. Pathology 2019, 51, 131–141. [Google Scholar] [CrossRef]
- Thomsen, C.; Storm, H.; Holst, J.J.; Hermansen, K. Differential effects of saturated and monounsaturated fats on postprandial lipemia and glucagon-like peptide 1 responses in patients with type 2 diabetes. Am. J. Clin. Nutr. 2003, 77, 605–611. [Google Scholar] [CrossRef]
- Nakajima, K.; Nakano, T.; Tokita, Y.; Nagamine, T.; Inazu, A.; Kobayashi, J.; Mabuchi, H.; Stanhope, K.L.; Havel, P.J.; Okazaki, M.; et al. Postprandial Lipoprotein Metabolism: VLDL vs. Chylomicrons. Clin. Chim. Acta 2011, 412, 1306–1318. [Google Scholar] [CrossRef]
- Otokozawa, S.; Ai, M.; Diffenderfer, M.R.; Asztalos, B.F.; Tanaka, A.; Lamon-Fava, S.; Schaefer, E.J. Fasting and postprandial apolipoprotein B-48 levels in healthy, obese, and hyperlipidemic subjects. Metabolism 2009, 58, 1536–1542. [Google Scholar] [CrossRef]
- Krysa, J.A.; Vine, D.F.; Beilin, L.J.; Burrows, S.; Huang, R.C.; Mori, T.A.; Proctor, S.D. ApoB48-remnant lipoproteins are associated with increased cardiometabolic risk in adolescents. Atherosclerosis 2020, 302, 20–26. [Google Scholar] [CrossRef]
- Rodríguez-Mortera, R.; Caccavello, R.; Garay-Sevilla, M.E.; Gugliucci, A. Higher ANGPTL3, apoC-III, and apoB48 dyslipidemia, and lower lipoprotein lipase concentrations are associated with dysfunctional visceral fat in adolescents with obesity. Clin. Chim Acta. 2020, 508, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Mager, R.; Mazurak, V.; Rodriguez-Dimitrescu, C.; Vine, D.; Jetha, M.; Ball, G.; Yap, J. A meal high in saturated fat evokes postprandial dyslipemia, hyperinsulinemia, and altered lipoprotein expression in obese children with and without nonalcoholic fatty liver disease. J. Parenter. Enteral Nutr. 2013, 37, 517–528. [Google Scholar] [CrossRef]
- Wang, Y.; Pendlebury, C.; Dodd, M.M.; Maximova, K.; Vine, D.F.; Jetha, M.M.; Ball, G.D.; Proctor, S.D. Elevated remnant lipoproteins may increase subclinical CVD risk in pre-pubertal children with obesity: A case-control study. Pediatr. Obes. 2013, 8, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Amigo-Benavent, M.; Sinausia, L.; Montero, E.; Perona, J.S. Brief Communication: Discordant ability of the triglyceride to apolipoprotein B ratio to predict triglyceride-rich lipoprotein particle size in normal-weight and obese men. Exp. Biol. Med. 2016, 241, 1772–1775. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Moruno, R.; Martinez-Force, E.; Montero, E.; Perona, J.S. Minor components of olive oil facilitate the triglyceride clearance from postprandial lipoproteins in a polarity-dependent manner in healthy men. Nutr. Res. 2014, 34, 40–47. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Karpe, F.; Olivecrona, T.; Hamsten, A.; Hultin, M. Chylomicron/chylomicron remnant turnover in humans: Evidence for margination of chylomicrons and poor conversion of larger to smaller chylomicron remnants. J. Lipid Res. 1997, 38, 949–961. [Google Scholar] [CrossRef]
- Sato, K.; Takahashi, Y.; Takahashi, T.; Katoh, N.; Akiba, Y. Identification of factors regulating lipoprotein lipase catalyzed hydrolysis in rats with the aid of monoacid-rich lipoprotein preparations. J. Nutr. Biochem. 2002, 13, 528. [Google Scholar] [CrossRef]
- Hussain, M.M. A proposed model for the assembly of chylomicrons. Atherosclerosis 2000, 148, 1–15. [Google Scholar] [CrossRef]
- Teng, K.T.; Chang, C.Y.; Chang, L.F.; Nesaretnam, K. Modulation of obesity-induced inflammation by dietary fats: Mechanisms and clinical evidence. Nutr. J. 2014, 13, 12. [Google Scholar] [CrossRef]
- Liput, K.P.; Lepczyński, A.; Ogłuszka, M.; Nawrocka, A.; Poławska, E.; Grzesiak, A.; Ślaska, B.; Pareek, C.S.; Czarnik, U.; Pierzchała, M. Effects of Dietary n-3 and n-6 Polyunsaturated Fatty Acids in Inflammation and Cancerogenesis. Int. J. Mol. Sci. 2021, 22, 6965. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Berhane, Y.; Botham, K.M.; Elliott, J.; Wheeler-Jones, C.P. Chylomicron-remnant-like particles modify production of vasoactive mediators by endothelial cells. Biochem. Soc. Trans. 2004, 32 Pt 1, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gill, R.; Pedersen, T.L.; Higgins, L.J.; Newman, J.W.; Rutledge, J.C. Triglyceride-rich lipoprotein lipolysis releases neutral and oxidized FFAs that induce endothelial cell inflammation. J. Lipid Res. 2009, 50, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Margioris, A.N. Fatty acids and postprandial inflammation. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.B.; Huang, L.L.; Rong, R.; Tan, R.; Wang, J.; Kang, J.X. Endogenously decreasing tissue n-6/n-3 fatty acid ratio reduces atherosclerotic lesions in apolipoprotein E-deficient mice by inhibiting systemic and vascular inflammation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2487–2494. [Google Scholar] [CrossRef]
- Krauss, R.M.; King, S.M. Remnant Lipoprotein Particles and Cardiovascular Disease Risk. Best Pr. Res. Clin. Endocrinol. Metab. 2022, 37, 101682. [Google Scholar] [CrossRef]
- Siri-Tarino, P.W.; Krauss, R.M. The Early Years of Lipoprotein Research: From Discovery to Clinical Application. J. Lipid Res. 2016, 57, 1771–1777. [Google Scholar] [CrossRef]
- Botham, K.M.; Moore, E.H.; De Pascale, C.; Bejta, F. The induction of macrophage foam cell formation by chylomicron remnants. Biochem. Soc. Trans. 2007, 35 Pt 3, 454–458. [Google Scholar] [CrossRef]
- Napolitano, M.; Avella, M.; Botham, K.M.; Bravo, E. Chylomicron remnant induction of lipid accumulation in J774 macrophages is associated with up-regulation of triacylglycerol synthesis which is not dependent on oxidation of the particles. Biochim. Biophys. Acta. 2003, 1631, 255–264. [Google Scholar] [CrossRef]
- Quintero-Flórez, A.; Sinausia Nieva, L.; Sánchez-Ortíz, A.; Beltrán, G.; Perona, J.S. The Fatty Acid Composition of Virgin Olive Oil from Different Cultivars Is Determinant for Foam Cell Formation by Macrophages. J. Agric. Food Chem. 2015, 63, 6731–6738. [Google Scholar] [CrossRef]
- Song, Z.; Xia, H.; Yang, L.; Wang, S.; Sun, G. Lowering the n-6/n-3 PUFAs ratio inhibits the formation of THP-1 macrophage-derived foam cell. Lipids Health Dis. 2018, 17, 125. [Google Scholar] [CrossRef]
- Ninla-Aesong, P.; Puangsri, P.; Kietdumrongwong, P.; Jongkrijak, H.; Noipha, K. Being overweight and obese increases suicide risk, the severity of depression, and the inflammatory response in adolescents with major depressive disorders. Front. Immunol. 2023, 14, 1197775. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Jian, D.Y.; Lin, M.W.; Zhao, J.Z.; Ho, L.T.; Juan, C.C. Evidence in obese children: Contribution of hyperlipidemia, obesity-inflammation, and insulin sensitivity. PLoS ONE 2015, 10, e0125935. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Becerril, S.; Salvador, J.; Portincasa, P.; Colina, I.; Gómez-Ambrosi, J. Involvement of the leptin-adiponectin axis in inflammation and oxidative stress in the metabolic syndrome. Sci. Rep. 2017, 7, 6619. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Riejos, P.; Najib, S.; Santos-Alvarez, J.; Martín-Romero, C.; Pérez-Pérez, A.; González-Yanes, C.; Sánchez-Margalet, V. Role of leptin in the activation of immune cells. Mediat. Inflamm. 2010, 2010, 568343. [Google Scholar] [CrossRef]
- Rubin, D.; Helwig, U.; Nothnagel, M.; Lemke, N.; Schreiber, S.; Fölsch, U.R.; Döring, F.; Schrezenmeir, J. Postprandial plasma adiponectin decreases after glucose and high fat meal and is independently associated with postprandial triacylglycerols but not with—11388 promoter polymorphism. Br. J. Nutr. 2008, 99, 76–82. [Google Scholar] [CrossRef] [PubMed]
- English, P.J.; Coughlin, S.R.; Hayden, K.; Malik, I.A.; Wilding, J.P. Plasma adiponectin increases postprandially in obese, but not in lean, subjects. Obes. Res. 2003, 11, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Poppitt, S.D.; Keogh, G.F.; Lithander, F.E.; Wang, Y.; Mulvey, T.B.; Chan, Y.K.; McArdle, B.H.; Cooper, G.J. Postprandial response of adiponectin, interleukin-6, tumor necrosis factor-alpha, and C-reactive protein to a high-fat dietary load. Nutrition 2008, 24, 22–29. [Google Scholar] [CrossRef]
- Gil-Campos, M.; Ramírez Tortosa, M.C.; Aguilera, C.M.; Cañete, R.; Gil, A. Fasting and postprandial adiponectin alterations anticipate NEFA and TNF-α changes in prepubertal obese children. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 62–68. [Google Scholar] [CrossRef]
- Annuzzi, G.; Bozzetto, L.; Patti, L.; Santangelo, C.; Giacco, R.; Di Marino, L.; De Natale, C.; Masella, R.; Riccardi, G.; Rivellese, A.A. Type 2 diabetes mellitus is characterized by reduced postprandial adiponectin response: A possible link with diabetic postprandial dyslipidemia. Metab. Clin. Exp. 2010, 59, 567–574. [Google Scholar] [CrossRef]
- De Souza, M.D.G.C.; Maranhão, P.A.; Panazzolo, D.G.; Nogueira Neto, J.F.; Bouskela, E.; Kraemer-Aguiar, L.G. Effects of a high-fat meal on inflammatory and endothelial injury biomarkers in accordance with adiposity status: A cross-sectional study. Nutr. J. 2022, 21, 65. [Google Scholar] [CrossRef]
- Larsen, M.A.; Isaksen, V.T.; Paulssen, E.J.; Goll, R.; Florholmen, J.R. Postprandial leptin and adiponectin in response to sugar and fat in obese and normal weight individuals. Endocrine 2019, 66, 517–525. [Google Scholar] [CrossRef]
- Holland, W.L.; Adams, A.C.; Brozinick, J.T.; Bui, H.H.; Miyauchi, Y.; Kusminski, C.M.; Bauer, S.M.; Wade, M.; Singhal, E.; Cheng, C.C.; et al. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell Metab. 2013, 17, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Iroz, A.; Montagner, A.; Benhamed, F.; Levavasseur, F.; Polizzi, A.; Anthony, E.; Régnier, M.; Fouché, E.; Lukowicz, C.; Cauzac, M.; et al. A Specific ChREBP and PPARα Cross-Talk Is Required for the Glucose-Mediated FGF21 Response. Cell Rep. 2017, 21, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Korek, E.; Krauss, H.; Gibas-Dorna, M.; Kupsz, J.; Piątek, M.; Piątek, J. Fasting and postprandial levels of ghrelin, leptin and insulin in lean, obese and anorexic subjects. Prz. Gastroenterol. 2013, 8, 383–389. [Google Scholar] [CrossRef]
- Stirban, A.; Negrean, M.; Götting, C.; Stratmann, B.; Gawlowski, T.; Mueller-Roesel, M.; Kleesiek, K.; Koschinsky, T.; Tschoepe, D. Leptin decreases postprandially in people with type 2 diabetes, an effect reduced by the cooking method. Horm. Metab. Res. 2008, 40, 896–900. [Google Scholar] [CrossRef] [PubMed]
- González-Jiménez, E.; Schmidt-Riovalle, J.; Sinausía, L.; Carmen Valenza, M.; Perona, J.S. Predictive value of ceruloplasmin for metabolic syndrome in adolescents. Biofactors 2016, 42, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, Á.; Perona, J.S.; Schmidt-RioValle, J.; Montero-Alonso, M.A.; Navarro-Pérez, C.F.; González-Jiménez, E. cMetS Based on Z-Scores as an Accurate and Efficient Scoring System to Determine Metabolic Syndrome in Spanish Adolescents. J. Pers. Med. 2022, 13, 10. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Normal-Weight | Obese | |||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Glucose (mg/dL) | 59.8 | 6.8 | 60.0 | 12.2 |
| Insulin (µIU/mL) | 7.3 | 3.2 | 13.6 | 5.7 ** |
| HOMA-IR | 0.8 | 0.4 | 1.6 | 0.6 ** |
| McAuley index | 9.6 | 1.2 | 7.3 | 1.1 *** |
| HbA1c (%) | 5.3 | 0.2 | 5.6 | 1.7 |
| Cholesterol (mg/dL) | 164.4 | 20.7 | 160.0 | 26.4 |
| LDL-cholesterol (mg/dL) | 83.0 | 18.6 | 87.8 | 20.3 |
| HDL-cholesterol (mg/dL) | 57.0 | 7.17 | 45.2 | 8.78 ** |
| Lipoprotein (a) | 57.3 | 37.3 | 76.9 | 53.2 |
| Triglycerides (mg/dL) | 58.7 | 10.2 | 76.3 | 22.8 * |
| Apolipoprotein A-I (mg/dL) | 159.8 | 13.3 | 137.1 | 14.4 ** |
| Apolipoprotein B (mg/dL) | 53.8 | 8.3 | 60.5 | 10.8 1 |
| Weight (kg) | 57.9 | 11.9 | 90.3 | 14.2 *** |
| Height (cm) | 163.8 | 8.0 | 164.0 | 7.8 |
| BMI (kg/m2) | 21.4 | 3.0 | 33.4 | 3.0 *** |
| Systolic pressure (mmHg) | 112.8 | 7.9 | 141.4 | 4.2 *** |
| Diastolic pressure (mmHg) | 60.4 | 2.7 | 76.7 | 2.6 *** |
| Upper Arm Circumference (cm) | 26.6 | 4.2 | 34.1 | 1.9 *** |
| Thigh Circumference (cm) | 52.5 | 9.2 | 63.9 | 10.5 *** |
| Waist Circumference (cm) | 79.7 | 14.2 | 100.7 | 8.6 ** |
| Hip Circumference (cm) | 91.7 | 10.5 | 107.8 | 7.8 *** |
| Tricipital fold (mm) | 20.8 | 9.3 | 31.8 | 7.2 ** |
| Bicipital fold (mm) | 13.5 | 8.8 | 21.8 | 5.3 * |
| Subscapular fold (mm) | 17.1 | 10.3 | 34.5 | 6.0 *** |
| Suprailiac fold (mm) | 20.3 | 12.5 | 36.2 | 7.6 ** |
| Calf Fold (mm) | 18.9 | 10.2 | 31.4 | 8.6 ** |
| Thigh Fold (mm) | 31.1 | 13.34 | 43.4 | 6.2 * |
| C-reactive protein (mg/dL) | 0.16 | 0.10 | 0.27 | 0.02 *** |
| MCP-1 (pg/mL) | 140.4 | 40.5 | 146.7 | 65.3 |
| IL-6 (pg/mL) | 82.0 | 57.9 | 229.4 | 167.2 ** |
| IL-1β (pg/mL) | 261.2 | 182.8 | 426.6 | 167.1 * |
| TNF-α (pg/mL) | 18.3 | 9.1 | 47.2 | 15.4 *** |
| Adiponectin (ng/mL) | 10.3 | 1.5 | 10.6 | 2.7 |
| Leptin (pg/mL) | 8.8 | 1.7 | 12.5 | 2.6 ** |
| Ceruloplasmin (μg/mL) | 440.8 | 179.5 | 594.8 | 176.9 * 1 |
| Normal-Weight | Obese | |||||
|---|---|---|---|---|---|---|
| r2 | Crude p | Corrected p | r2 | Crude p | Corrected p | |
| Glucose (mg/dL) | −0.373 | 0.161 | 0.572 | −0.116 | 0.360 | 0.411 |
| Insulin (µIU/mL) | 0.484 | 0.136 | 0.544 | 0.609 | 0.040 | 0.213 |
| HOMA-IR | −0.266 | 0.245 | 0.490 | 0.341 | 0.139 | 0.262 |
| McAuley index | −0.498 | 0.128 | 0.585 | −0.452 | 0.130 | 0.260 |
| HbA1c (%) | 0.388 | 0.171 | 0.547 | 0.233 | 0.233 | 0.324 |
| Cholesterol (mg/dL) | −0.515 | 0.066 | 0.528 | 0.337 | 0.142 | 0.252 |
| LDL-cholesterol (mg/dL) | −0.541 | 0.078 | 0.499 | 0.481 | 0.057 | 0.182 |
| HDL-cholesterol (mg/dL) | −0.525 | 0.060 | 0.640 | −0.405 | 0.026 | 0.208 |
| Lipoprotein (a) (mg/dL) | 0.110 | 0.393 | 0.434 | 0.869 | 0.006 | 0.192 |
| Apolipoprotein A-I (mg/dL) | −0.563 | 0.045 | 0.720 | 0.110 | 0.292 | 0.346 |
| Apolipoprotein B (mg/dL) | −0.162 | 0.327 | 0.419 | 0.503 | 0.048 | 0.171 |
| BMI (kg/m2) | 0.001 | 0.928 | 0.928 | 0.666 | 0.013 | 0.208 |
| Systolic pressure (mmHg) | 0.198 | 0.292 | 0.467 | −0.464 | 0.065 | 0.189 |
| Diastolic pressure (mmHg) | 0.116 | 0.375 | 0.429 | −0.436 | 0.078 | 0.192 |
| Upper Arm Circumference (cm) | −0.123 | 0.368 | 0.436 | −0.444 | 0.074 | 0.197 |
| Thigh Circumference (cm) | −0.217 | 0.274 | 0.516 | −0.239 | 0.227 | 0.363 |
| Waist Circumference (cm) | 0.209 | 0.281 | 0.500 | 0.506 | 0.047 | 0.188 |
| Hip Circumference (cm) | 0.206 | 0.284 | 0.478 | 0.617 | 0.016 | 0.171 |
| Tricipital fold (mm) | −0.279 | 0.217 | 0.534 | 0.220 | 0.246 | 0.328 |
| Bicipital fold (mm) | −0.009 | 0.490 | 0.506 | 0.206 | 0.260 | 0.333 |
| Subscapular fold (mm) | −0.106 | 0.324 | 0.432 | −0.190 | 0.277 | 0.341 |
| Suprailiac fold (mm) | −0.179 | 0.310 | 0.431 | 0.027 | 0.467 | 0.482 |
| Calf Fold (mm) | −0.130 | 0.360 | 0.443 | −0.113 | 0.363 | 0.401 |
| Thigh Fold (mm) | −0.264 | 0.230 | 0.491 | 0.371 | 0.118 | 0.270 |
| C-reactive protein (mg/dL) | 0.444 | 0.189 | 0.550 | −0.291 | 0.231 | 0.336 |
| MCP-1 (pg/mL) | 0.068 | 0.426 | 0.454 | 0.411 | 0.119 | 0.254 |
| IL-6 (pg/mL) | 0.740 | 0.103 | 0.549 | 0.757 | 0.227 | 0.346 |
| IL-1β (pg/mL) | 0.242 | 0.301 | 0.459 | 0.023 | 0.481 | 0.481 |
| TNF-α (pg/mL) | 0.570 | 0.215 | 0.573 | 0.233 | 0.425 | 0.453 |
| Adiponectin (ng/mL) | 0.181 | 0.308 | 0.448 | 0.377 | 0.142 | 0.239 |
| Leptin (pg/mL) | 0.349 | 0.222 | 0.507 | 0.584 | 0.030 | 0.192 |
| Ceruloplasmin (mg/mL) | 0.747 | 0.017 | 0.544 | 0.572 | 0.042 | 0.192 1 |
| 0 h | 2 h | 4 h | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal-Weight | Obese | Normal-Weight | Obese | Normal-Weight | Obese | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| 14:0 | 1.41 | 0.91 | 1.25 | 0.60 | 1.13 | 0.57 | 1.15 | 0.36 | 1.59 | 0.17 | 1.47 | 0.26 |
| 14:1 n-5 | 0.46 | 0.32 | 0.22 | 0.25 | 0.59 | 0.73 | 0.26 | 0.23 | 0.48 | 0.13 | 0.24 | 0.08 *** |
| 16:0 | 20.33 | 2.38 | 21.57 | 2.87 | 20.60 | 2.13 | 21.04 | 3.10 | 19.55 | 1.47 | 21.20 | 2.83 |
| 16:1 n-7 | 4.66 | 1.60 | 4.14 | 0.88 | 3.76 | 1.14 | 3.92 | 0.81 | 3.51 | 0.71 | 3.87 | 0.62 |
| 18:0 | 6.86 | 3.64 | 4.25 | 1.53 * | 7.87 | 5.16 | 4.39 | 1.45 * | 5.79 | 1.30 | 4.16 | 0.86 ** |
| 18:1 n-9 | 38.57 | 6.61 | 38.09 | 3.64 | 40.52 | 7.34 | 40.70 | 4.48 | 44.28 | 3.53 | 40.49 | 3.98 * |
| 18:2 n-6 | 19.28 | 4.62 | 23.73 | 4.94 * | 18.26 | 3.52 | 22.01 | 3.25 * | 18.17 | 1.77 | 20.91 | 2.23 ** |
| 18:3 n-6 | 0.66 | 0.71 | 0.79 | 0.28 | 0.49 | 0.30 | 0.65 | 0.26 | 0.54 | 0.15 | 0.77 | 0.22 |
| 18:3 n-3 | 0.74 | 0.33 | 0.66 | 0.27 | 0.83 | 0.57 | 0.56 | 0.18 | 0.58 | 0.13 | 0.73 | 0.22 |
| 20:0 | 0.60 | 0.52 | 0.39 | 0.19 | 0.68 | 0.63 | 0.42 | 0.20 | 0.35 | 0.08 | 0.38 | 0.13 |
| 20:1 n-9 | 1.19 | 0.78 | 0.54 | 0.62 * | 1.38 | 1.73 | 0.54 | 0.66 | 0.27 | 0.08 | 0.34 | 0.10 |
| 20:2 n-6 | 0.32 | 0.14 | 0.49 | 0.16 * | 0.26 | 0.11 | 0.44 | 0.11 | 0.17 | 0.05 | 0.30 | 0.09 |
| 20:4 n-6 | 2.07 | 0.50 | 2.59 | 0.50 | 1.92 | 0.55 | 2.20 | 0.33 | 1.57 | 0.23 | 1.49 | 0.18 |
| 20:5 n-3 | 0.55 | 0.22 | 0.47 | 0.20 | 0.72 | 0.61 | 0.36 | 0.22 | 2.24 | 0.23 | 2.41 | 0.28 |
| 22:6 n-3 | 2.30 | 0.86 | 1.36 | 0.17 ** | 1.32 | 0.23 | 1.53 | 0.21 | 0.90 | 0.18 | 1.23 | 0.26 ** |
| SFA | 29.20 | 5.48 | 27.46 | 4.24 | 30.28 | 7.25 | 27.00 | 4.23 | 27.29 | 2.16 | 27.21 | 3.22 |
| MUFA | 44.98 | 5.19 | 42.99 | 2.99 | 46.28 | 5.29 | 45.39 | 4.12 | 48.55 | 3.07 | 44.95 | 3.68 * |
| PUFA n-6 | 22.33 | 5.32 | 27.59 | 4.99 * | 20.93 | 3.88 | 25.30 | 3.41 ** | 20.45 | 1.79 | 23.47 | 2.22 ** |
| PUFA n-3 | 3.59 | 1.28 | 2.49 | 0.70 * | 2.87 | 1.41 | 2.44 | 0.51 | 3.72 | 0.22 | 4.37 | 0.32 *** 1 |
| 0 h | 2 h | 4 h | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal-Weight | Obese | Normal-Weight | Obese | Normal-Weight | Obese | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| 14:0 | 1.96 | 1.21 | 1.09 | 0.62 * | 1.36 | 1.15 | 0.94 | 0.54 | 2.14 | 0.30 | 1.99 | 0.52 |
| 14:1 n-5 | 1.54 | 1.11 | 0.74 | 0.72 * | 0.32 | 0.21 | 0.50 | 0.36 | 0.37 | 0.16 | 0.35 | 0.08 |
| 16:0 | 25.02 | 6.10 | 25.33 | 6.07 | 23.82 | 3.36 | 24.10 | 2.39 | 21.92 | 1.01 | 20.54 | 2.03 |
| 16:1 n-7 | 3.03 | 2.31 | 2.62 | 1.88 | 4.21 | 2.63 | 2.48 | 1.43 ** | 1.88 | 0.18 | 1.97 | 0.18 |
| 18:0 | 18.59 | 5.41 | 19.16 | 2.56 | 18.46 | 1.72 | 17.73 | 3.37 | 15.59 | 1.50 | 15.05 | 1.65 |
| 18:1 n-9 | 17.31 | 6.03 | 15.17 | 2.92 | 17.11 | 6.05 | 16.84 | 6.41 | 25.60 | 1.46 | 26.11 | 1.84 |
| 18:2 n-6 | 11.95 | 3.06 | 15.68 | 3.60 * | 13.99 | 5.44 | 16.36 | 3.62 | 19.25 | 1.36 | 18.19 | 1.32 |
| 18:3 n-6 | 0.39 | 0.23 | 0.50 | 0.25 | 0.47 | 0.27 | 0.35 | 0.16 | 1.17 | 0.13 | 1.08 | 0.10 |
| 18:3 n-3 | 2.61 | 1.99 | 1.86 | 1.32 | 3.48 | 2.31 | 1.83 | 1.33 * | 0.28 | 0.09 | 0.35 | 0.10 |
| 20:0 | 1.58 | 1.32 | 0.69 | 0.66 * | 0.43 | 0.21 | 0.79 | 0.76 | 0.18 | 0.06 | 0.26 | 0.08 * |
| 20:1 n-9 | 2.48 | 1.69 | 1.16 | 0.73 * | 1.00 | 0.74 | 1.41 | 1.04 | 1.28 | 0.29 | 1.21 | 0.24 |
| 20:2 n-6 | 1.79 | 0.79 | 2.90 | 1.23 * | 3.53 | 2.03 | 2.70 | 1.11 | 2.03 | 1.15 | 2.13 | 0.63 |
| 20:4 n-6 | 6.28 | 2.50 | 8.37 | 2.79 * | 7.24 | 3.53 | 7.73 | 2.11 | 5.48 | 1.04 | 7.52 | 0.68 *** |
| 20:5 n-3 | 1.81 | 1.63 | 1.11 | 0.84 | 0.90 | 0.81 | 1.28 | 0.95 | 1.28 | 0.21 | 1.51 | 0.39 |
| 22:6 n-3 | 3.69 | 1.65 | 4.29 | 1.54 | 3.77 | 1.36 | 4.96 | 1.85 | 1.56 | 0.27 | 1.75 | 0.30 |
| SFA | 47.16 | 10.40 | 45.49 | 7.12 | 44.08 | 3.23 | 43.56 | 3.33 | 39.82 | 1.47 | 37.85 | 2.08 |
| MUFA | 24.32 | 5.80 | 19.31 | 3.85 * | 22.55 | 6.59 | 21.23 | 5.06 | 29.13 | 1.48 | 29.64 | 1.82 |
| PUFA n-6 | 20.41 | 5.24 | 27.91 | 7.31 ** | 25.23 | 8.13 | 27.14 | 5.31 | 27.93 | 1.21 | 28.91 | 1.10 |
| PUFA n-3 | 8.22 | 4.53 | 5.11 | 2.52 * | 8.14 | 2.68 | 8.07 | 2.60 | 3.12 | 0.25 | 3.61 | 0.41 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, S.; Espinosa-Cabello, J.M.; García-González, A.; González-Jiménez, E.; Aguilar-Cordero, M.J.; Castellano, J.M.; Perona, J.S. Interplay of Postprandial Triglyceride-Rich Lipoprotein Composition and Adipokines in Obese Adolescents. Int. J. Mol. Sci. 2024, 25, 1112. https://doi.org/10.3390/ijms25021112
García-Rodríguez S, Espinosa-Cabello JM, García-González A, González-Jiménez E, Aguilar-Cordero MJ, Castellano JM, Perona JS. Interplay of Postprandial Triglyceride-Rich Lipoprotein Composition and Adipokines in Obese Adolescents. International Journal of Molecular Sciences. 2024; 25(2):1112. https://doi.org/10.3390/ijms25021112
Chicago/Turabian StyleGarcía-Rodríguez, Silvia, Juan M. Espinosa-Cabello, Aída García-González, Emilio González-Jiménez, María J. Aguilar-Cordero, José M. Castellano, and Javier S. Perona. 2024. "Interplay of Postprandial Triglyceride-Rich Lipoprotein Composition and Adipokines in Obese Adolescents" International Journal of Molecular Sciences 25, no. 2: 1112. https://doi.org/10.3390/ijms25021112
APA StyleGarcía-Rodríguez, S., Espinosa-Cabello, J. M., García-González, A., González-Jiménez, E., Aguilar-Cordero, M. J., Castellano, J. M., & Perona, J. S. (2024). Interplay of Postprandial Triglyceride-Rich Lipoprotein Composition and Adipokines in Obese Adolescents. International Journal of Molecular Sciences, 25(2), 1112. https://doi.org/10.3390/ijms25021112