
In Vitro Effects of Fentanyl on Aortic Viscoelasticity in a Rat Model of Melatonin Deficiency

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
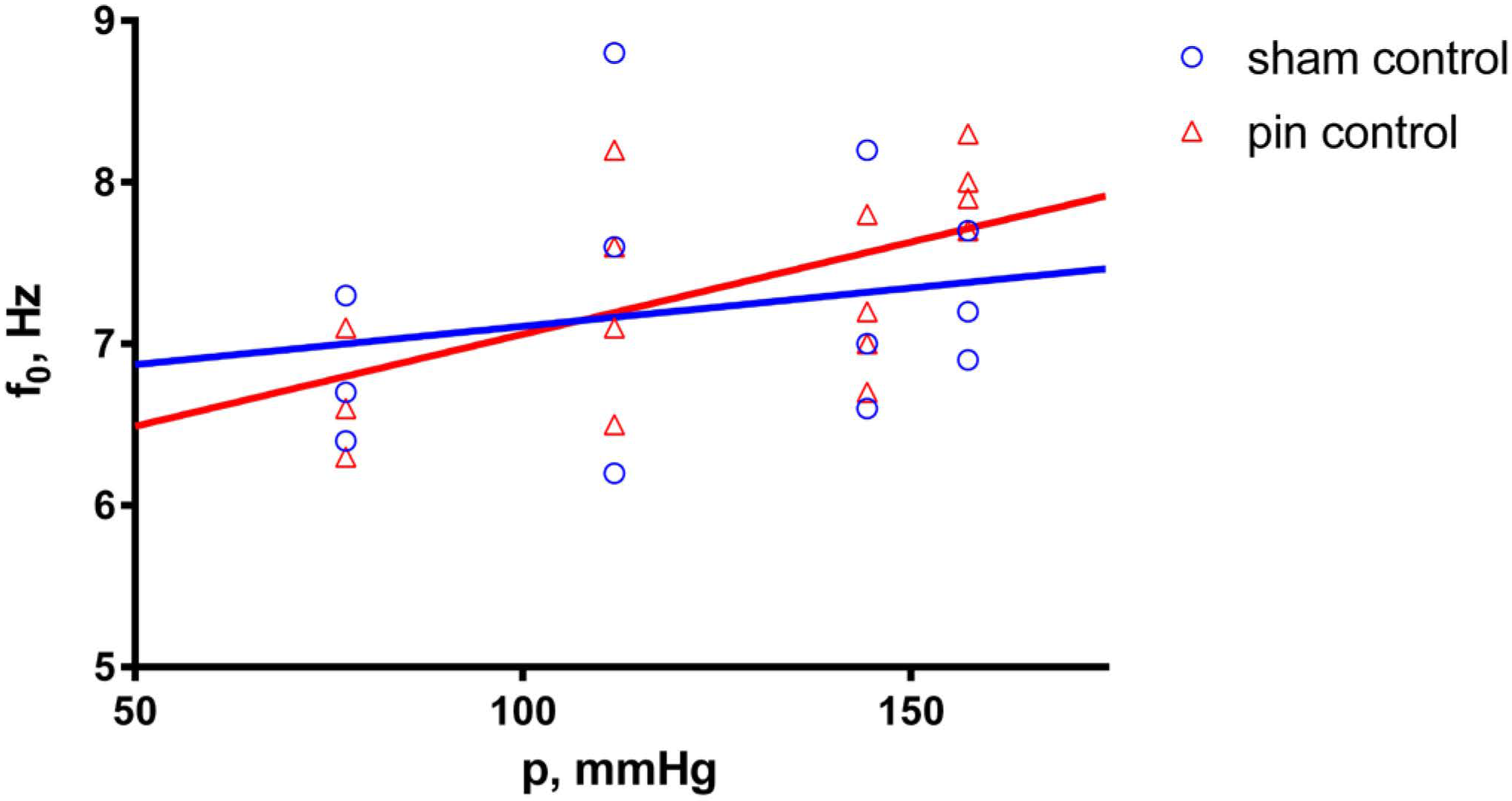
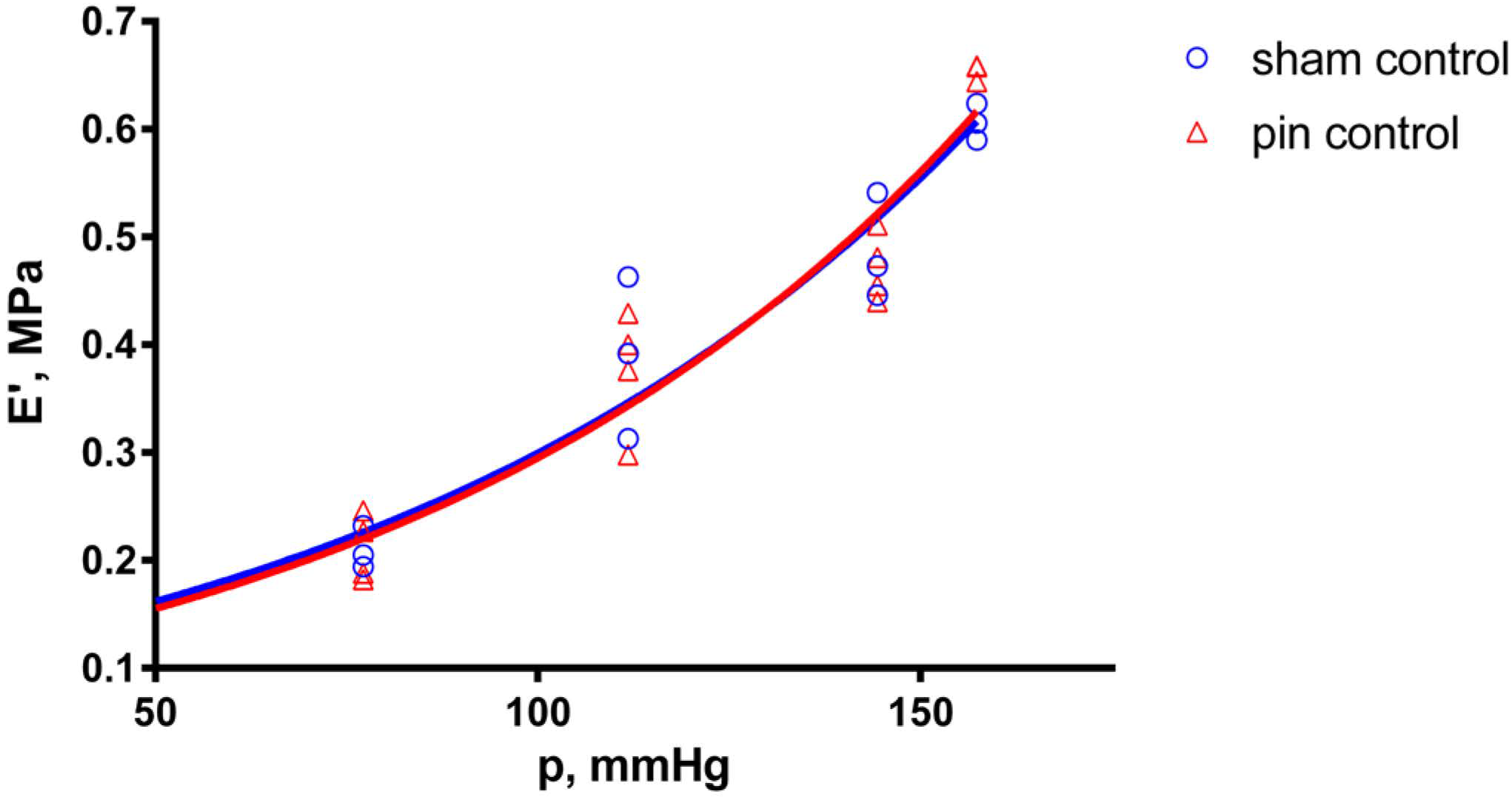
2.1. Effects of Pinealectomy in Untreated Aortic Preparations
2.2. Effects of Pinealectomy in Fentanyl-Treated Aortic Preparations
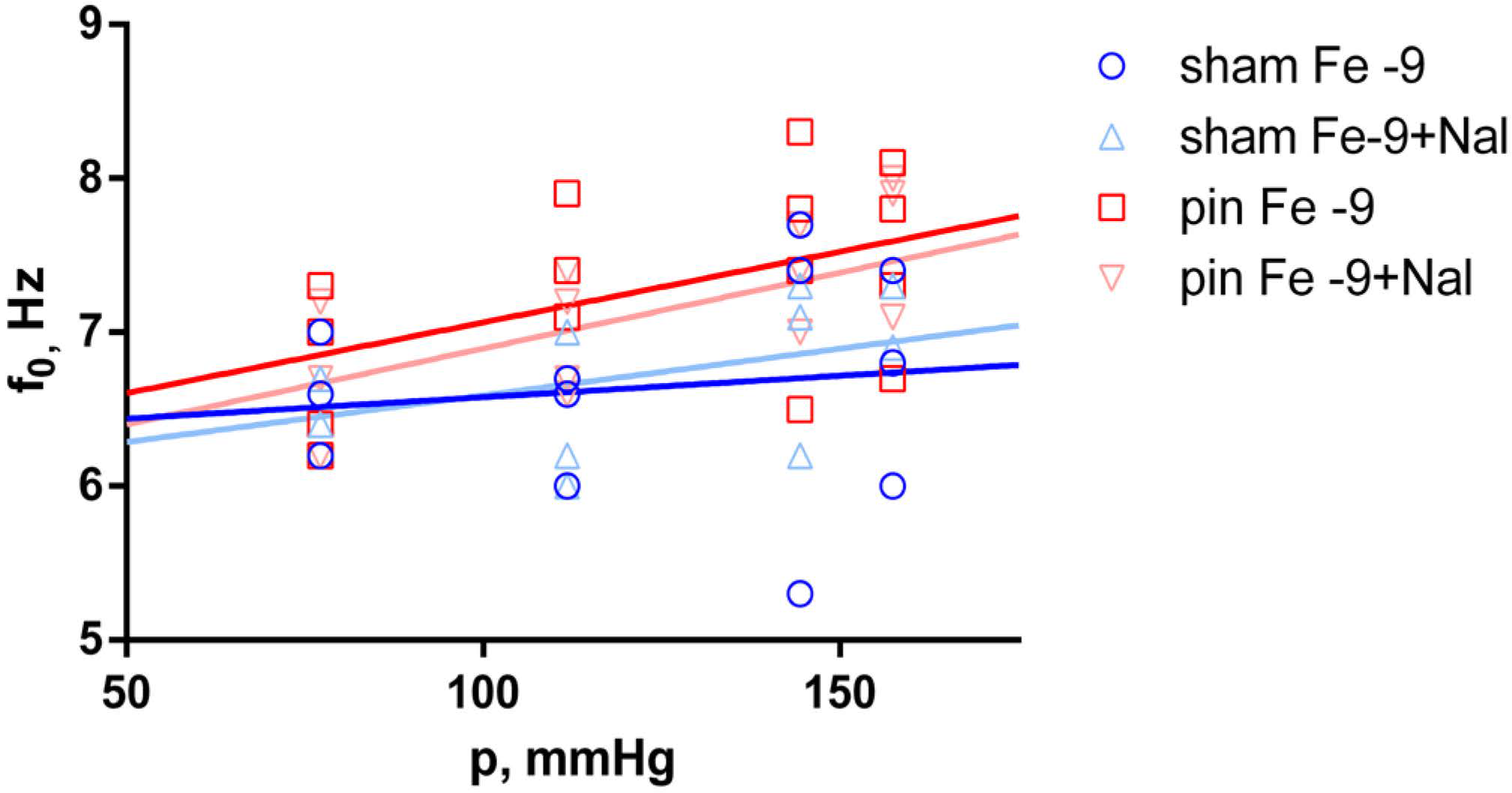
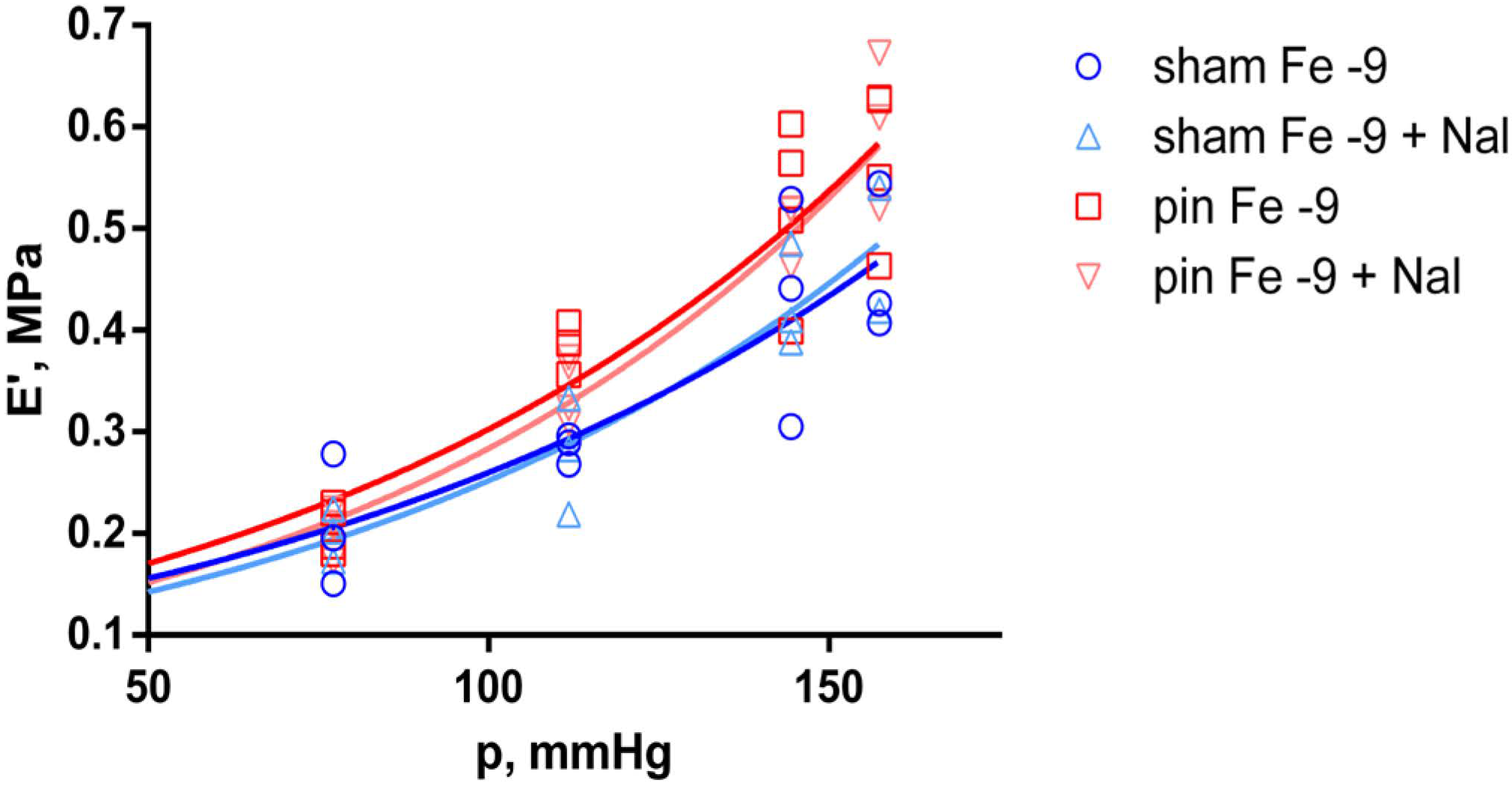
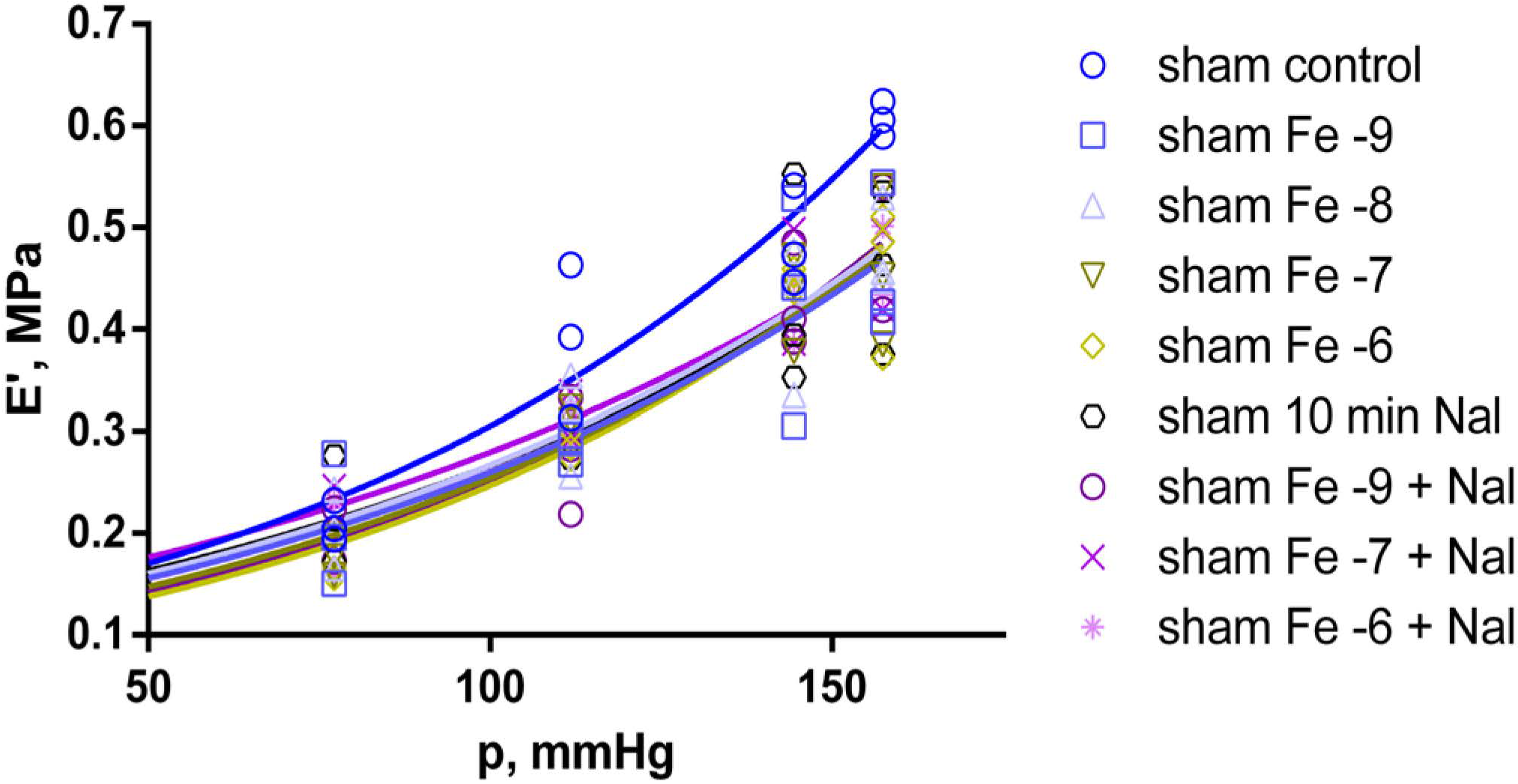
2.3. Effects of Fentanyl and Fentanyl + Naloxone in the Sham Group
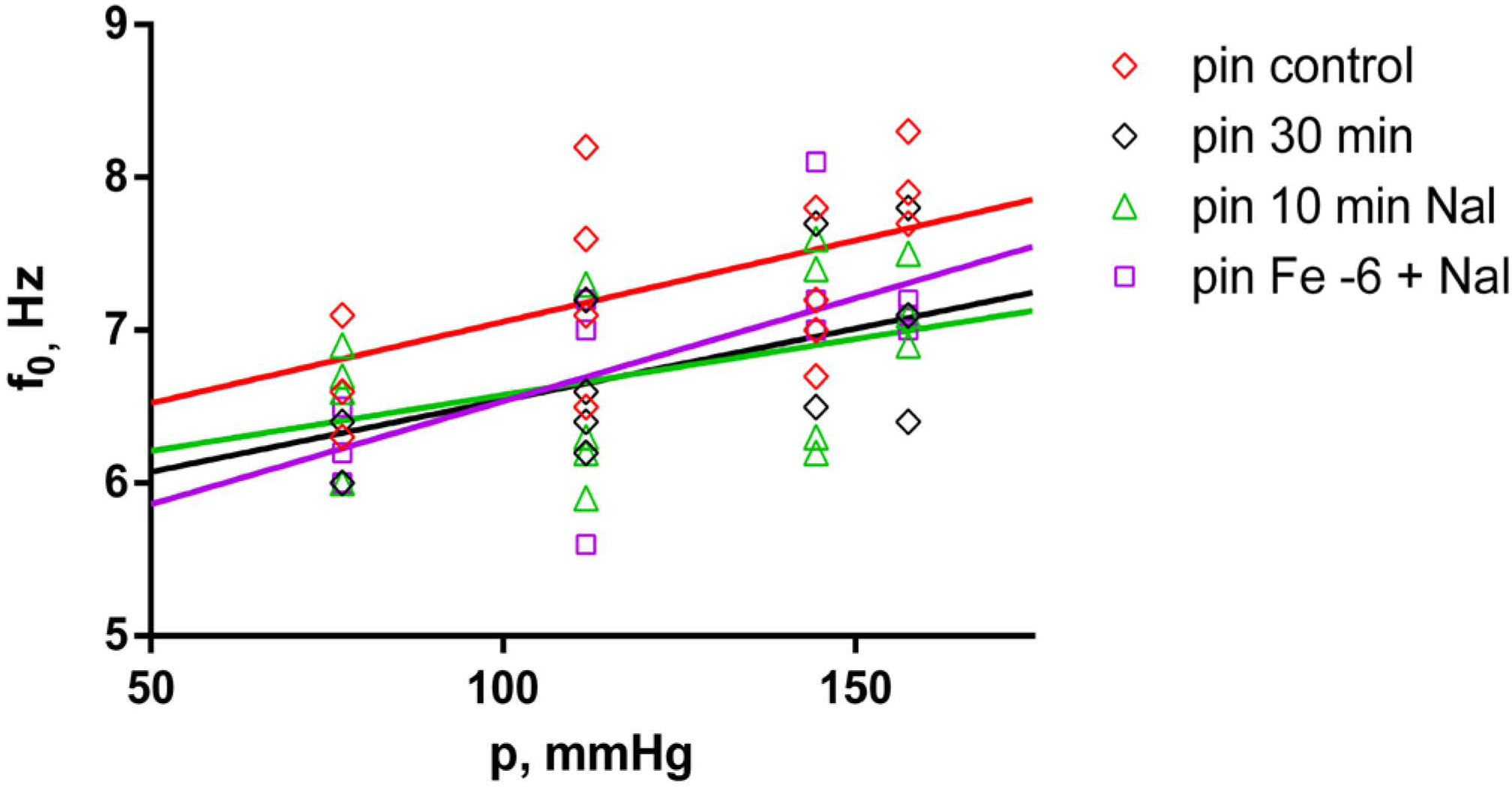
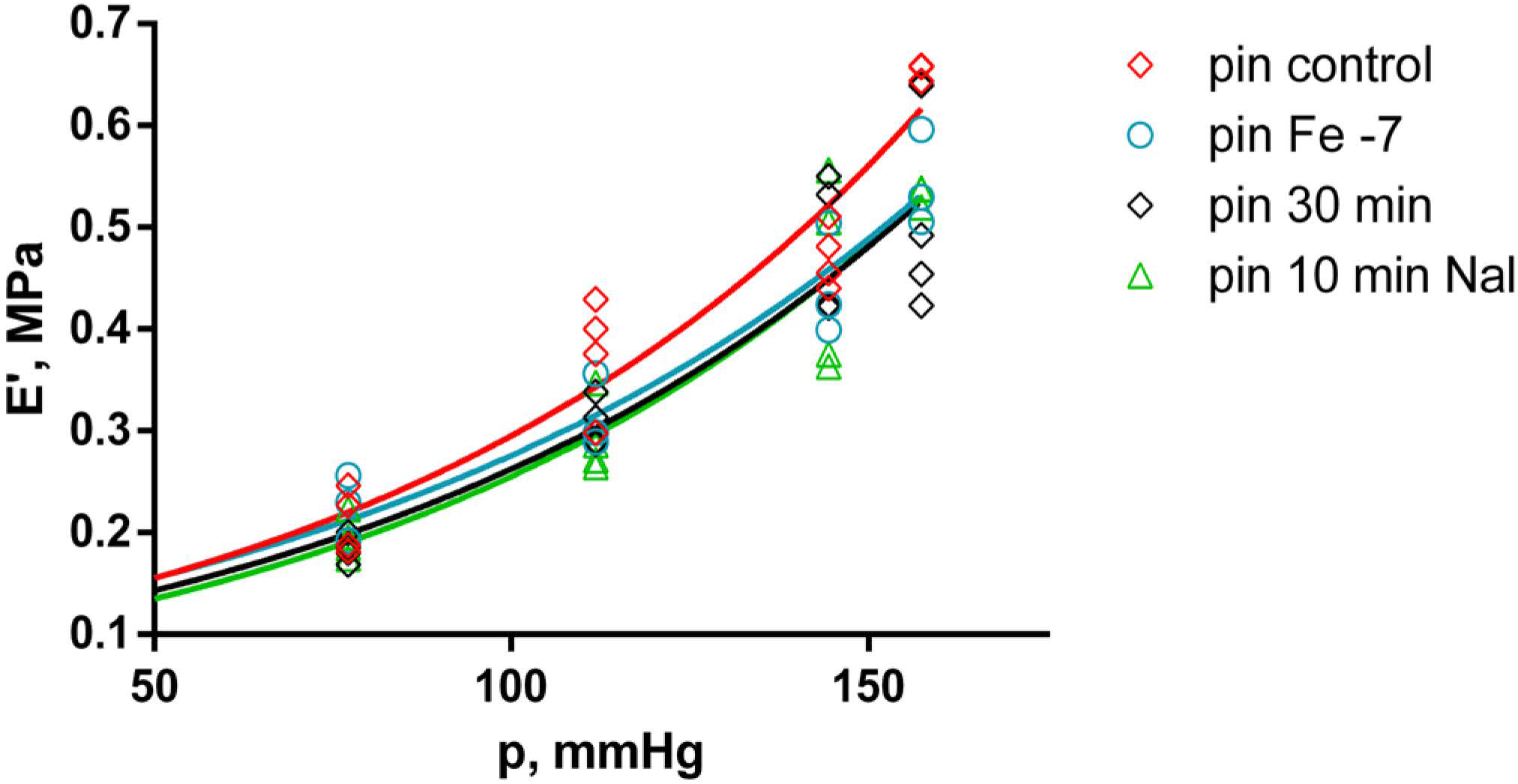
2.4. Effects of Fentanyl and Fentanyl + Naloxone in the Pin Group
3. Discussion
3.1. Effects of Pinealectomy
3.2. Effects of Fentanyl
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Surgical Procedure
4.4. Preparation of Aortic Strips
4.5. Nutrient Medium
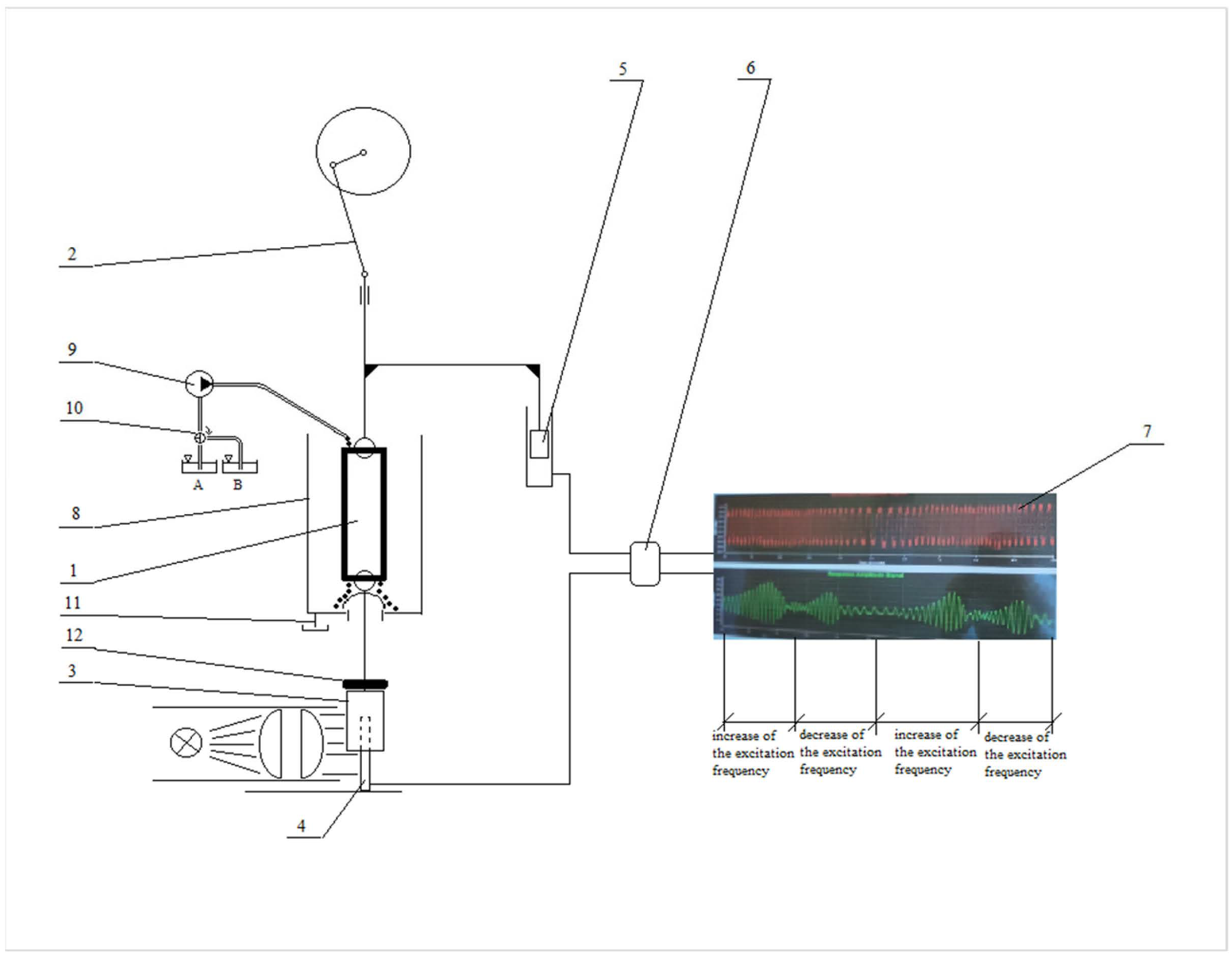
4.6. Forced Oscillations Method
4.7. Experimental Protocol
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antonova, M.L.; Antonov, P.S.; Marinov, G.R.; Vlaskovska, M.V.; Kasakov, L.N. Viscoelastic Characteristics of in Vitro Vital and Devitalized Rat Aorta and Human Arterial Prostheses. Ann. Biomed. Eng. 2008, 36, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Bergel, D.H. The Dynamic Elastic Properties of the Arterial Wall. J. Physiol. 1961, 156, 458. [Google Scholar] [CrossRef] [PubMed]
- Antonov, P.; Antonova, M.; Nikolova, N.; Antonova, N.; Vlaskovska, M.; Kasakov, L. Age Dependent Changes of Arterial Wall Viscoelasticity. Clin. Hemorheol. Microcirc. 2008, 39, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.X.; Tomioka, J.; Debes, J.C.; Fung, Y.C. New Experiments on Shear Modulus of Elasticity of Arteries. Am. J. Physiol. Hear. Circ. Physiol. 1994, 266, H1–H10. [Google Scholar] [CrossRef] [PubMed]
- Franchini, G.; Breslavsky, I.D.; Holzapfel, G.A.; Amabili, M. Viscoelastic Characterization of Human Descending Thoracic Aortas under Cyclic Load. Acta Biomater. 2021, 130, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Apter, J.T.; Marquez, E. A Relation between Hysteresis and Other Visco Elastic Properties of Some Biomaterials. Biorheology 1968, 5, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Bergel, D.H. The Static Elastic Properties of the Arterial Wall. J. Physiol. 1961, 156, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Hardung, V. Dynamic Elasticity and Interior Friction of Muscular Blood Vessels during Varying Wall Stress Caused by Stretching and Tonic Contration. Arch. Kreislaufforsch. 1970, 61, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Petkov, S. In Vitro Investigation of the Mechanic and Magnet-Resonance Properties of Human Blood Vessels; Technical University Gabrovo: Gabrovo, Bulgaria, 1998. [Google Scholar]
- Mammoto, A.; Matus, K.; Mammoto, T. Extracellular Matrix in Aging Aorta. Front. Cell Dev. Biol. 2022, 10, 367. [Google Scholar] [CrossRef]
- Nagai, Y.; Metter, E.J.; Earley, C.J.; Kemper, M.K.; Becker, L.C.; Lakatta, E.G.; Fleg, J.L. Increased Carotid Artery Intimal-Medial Thickness in Asymptomatic Older Subjects with Exercise-Induced Myocardial Ischemia. Circulation 1998, 98, 1504–1509. [Google Scholar] [CrossRef]
- Oishi, Y.; Miyoshi, H.; Mizuguchi, Y.; Iuchi, A.; Nagase, N.; Oki, T. Aortic Stiffness Is Strikingly Increased with Age ≥ 50 Years in Clinically Normal Individuals and Preclinical Patients with Cardiovascular Risk Factors: Assessment by the New Technique of 2D Strain Echocardiography. J. Cardiol. 2011, 57, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Makino, A.; Kakoi, D. Remodeling of Arterial Wall: Response to Changes in Both Blood Flow and Blood Pressure. J. Mech. Behav. Biomed. Mater. 2018, 77, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, G.A.; Ogden, R.W. Biomechanical Relevance of the Microstructure in Artery Walls with a Focus on Passive and Active Components. Am. J. Physiol. Hear. Circ. Physiol. 2018, 315, H540–H549. [Google Scholar] [CrossRef] [PubMed]
- Learoyd, B.M.; Taylor, M.G. Alterations with Age in the Viscoelastic Properties of Human Arterial Walls. Circ. Res. 1966, 18, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Elliott, W.; Guo, D.; Veldtman, G.; Tan, W. Effect of Viscoelasticity on Arterial-Like Pulsatile Flow Dynamics and Energy. J. Biomech. Eng. 2020, 142, 0410011. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Han, S.M.; Park, S.H.; Ryu, S.J.; Kim, K.H.; Chang, T.H.; Kim, S.H. Effect of Sufentanil and Fentanyl on Heart Rate and Blood Pressure Changes during Induction of Anesthesia. Korean J. Anesthesiol. 1998, 35, 662–668. [Google Scholar] [CrossRef]
- Agabiti-Rosei, C.; Favero, G.; De Ciuceis, C.; Rossini, C.; Porteri, E.; Rodella, L.F.; Franceschetti, L.; Maria Sarkar, A.; Agabiti-Rosei, E.; Rizzoni, D.; et al. Effect of Long-Term Treatment with Melatonin on Vascular Markers of Oxidative Stress/Inflammation and on the Anticontractile Activity of Perivascular Fat in Aging Mice. Hypertens. Res. 2017, 40, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Lonati, C.; Giugno, L.; Castrezzati, S.; Rodella, L.F.; Rezzani, R. Obesity-Related Dysfunction of the Aorta and Prevention by Melatonin Treatment in Ob/Ob Mice. Acta Histochem. 2013, 115, 783–788. [Google Scholar] [CrossRef]
- Repová-Bednárová, K.; Aziriová, S.; Hrenák, J.; Krajčírovičová, K.; Adamcová, M.; Paulis, L.; Šimko, F. Effect of Captopril and Melatonin on Fibrotic Rebuilding of the Aorta in 24 Hour Light-Induced Hypertension. Physiol. Res. 2013, 62, S135–S141. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Do Amaral, F.G. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef]
- Reiter, R.J.; Richardson, B.A.; Johnson, L.Y.; Ferguson, B.N.; Dinh, D.T. Pineal Melatonin Rhythm: Reduction in Aging Syrian Hamsters. Science 1980, 210, 1372–1373. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.L.; Lewy, A.J.; Erb, D.L.; Vollmer, W.M.; Singer, C.M. Human Melatonin Production Decreases with Age. J. Pineal Res. 1986, 3, 379–388. [Google Scholar] [CrossRef]
- Hardeland, R. Neurobiology, Pathophysiology, and Treatment of Melatonin Deficiency and Dysfunction. Sci. World J. 2012, 2012, 640389. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.M.; Bar-Or, A.; Grossi, D.; Kashur, S.; Johannson, E.; Yie, S.M. Urinary 6-sulphatoxymelatonin, an Index of Pineal Function in the Rat. J. Pineal Res. 1991, 10, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.W. and D.S. The Effect of Melatonin on Pinealectomyinduced Hypertension in the Rat. Br. J. Pharmacol. 1976, 56, 360–361. [Google Scholar]
- Lewy, A.J.; Tetsuo, M.; Markey, S.P.; Goodwin, F.K.; Kopin, I.J. Pinealectomy Abolishes Plasma Melatonin in the Rat. J. Clin. Endocrinol. Metab. 1980, 50, 204–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.C.; Shieh, J.P.; Chung, H.H.; Hung, C.H.; Lin, H.J.; Cheng, J.T. Activation of Peripheral Opioid μ-Receptors in Blood Vessel May Lower Blood Pressure in Spontaneously Hypertensive Rats. Pharmacology 2011, 87, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Wu, C.; Zhang, M.; Yang, Z.; Liu, Z.; Fu, F.; Li, J.; Feng, N.; Gu, X.; Zhang, S.; et al. κ-Opioid Receptor Stimulation Alleviates Rat Vascular Smooth Muscle Cell Calcification via PFKFB3-Lactate Signaling. Aging 2021, 13, 14355–14371. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Sato, K.; Shirai, R.; Seki, T.; Shibata, K.; Yamashita, T.; Koide, A.; Tezuka, H.; Mori, Y.; Hirano, T.; et al. β-Endorphin Mediates the Development and Instability of Atherosclerotic Plaques. Int. J. Endocrinol. 2020, 2020, 4139093. [Google Scholar] [CrossRef] [PubMed]
- Parra, L.; Pérez-Vizcaíno, F.; Alsasua, A.; Martín, M.I.; Tamargo, J. Μ- and Δ-Opioid Receptor-Mediated Contractile Effects on Rat Aortic Vascular Smooth Muscle. Eur. J. Pharmacol. 1995, 277, 99–105. [Google Scholar] [CrossRef]
- Saeed, R.W. Expression of Functional Delta Opioid Receptors in Vascular Smooth Muscle. Int. J. Mol. Med. 2000, 6, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B.; Hartman, A.; Bilfinger, T.V.; Magazine, H.I.; Liu, Y.; Casares, F.; Goligorsky, M.S. Presence of the Μ3 Opiate Receptor in Endothelial Cells: Coupling to Nitric Oxide Production and Vasodilation. J. Biol. Chem. 1995, 270, 30290–30293. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, F.; Iwanov, V.; Moulds, R.F.W. Effects of Fentanyl on the Rat Aorta Are Mediated by Alpha-Adrenoceptors Rather than by the Endothelium. Br. J. Anaesth. 1993, 71, 877–880. [Google Scholar] [CrossRef] [PubMed]
- Park, K.E.; Sohn, J.T.; Jeong, Y.S.; Sung, H.J.; Shin, I.W.; Lee, H.K.; Chung, Y.K. Inhibitory Effect of Fentanyl on Phenylephrine-Induced Contraction of the Rat Aorta. Yonsei Med. J. 2009, 50, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, F.; Iwanov, V.; Moulds, R.F.W. Sufentanil and Alfentanil Cause Vasorelaxation By Mechanisms Independent of the Endothelium. Clin. Exp. Pharmacol. Physiol. 1993, 20, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Migheli, R.; Lostia, G.; Galleri, G.; Rocchitta, G.; Serra, P.A.; Campesi, I.; Bassareo, V.; Acquas, E.; Peana, A.T. New Perspective for an Old Drug: Can Naloxone Be Considered an Antioxidant Agent? Biochem. Biophys. Rep. 2023, 34, 101441. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive Oxygen Species: Key Regulators in Vascular Health and Diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, K.J.; Corey, R.A.; Alhosan, N.; Cavallo, D.; Groom, S.; Santiago, M.; Bailey, C.; Charlton, S.J.; Sessions, R.B.; Henderson, G.; et al. Interaction With the Lipid Membrane Influences Fentanyl Pharmacology. Adv. Drug Alcohol Res. 2022, 2, 10280. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.A.; Reiter, R.J. Rapid Pinealectomy in Hamsters and other Small Rodents. Anat. Rec. 1965, 153, 19–21. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Nenchovska, Z.; Atanasova, D.; Atanasova, M.; Kortenska, L.; Stefanova, M.; Alova, L.; Lazarov, N. Consequences of Long-Term Treatment with Agomelatine on Depressive-like Behavior and Neurobiological Abnormalities in Pinealectomized Rats. Behav. Brain Res. 2016, 302, 11–28. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Stoyanova, T.; Nenchovska, Z.; Ivanova, N.; Atanasova, D.; Atanasova, M.; Georgieva, K. Effect of Endurance Training on Diurnal Rhythms of Superoxide Dismutase Activity, Glutathione and Lipid Peroxidation in Plasma of Pinealectomized Rats. Neurosci. Lett. 2020, 716, 134637. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Hrischev, P.; Ivanova, P.; Boyadjiev, N.; Georgieva, K. Metabolic Footprint in Young, Middle-Aged and Elderly Rats with Melatonin Deficit. Physiol. Behav. 2022, 250, 113786. [Google Scholar] [CrossRef] [PubMed]
- Pierpaoli, W. The Pineal Gland as Ontogenetic Scanner of Reproduction, Immunity, and Aging. The Aging Clock. In Proceedings of the Annals of the New York Academy of Sciences; Wiley: Hoboken, NJ, USA, 1994; Volume 741, pp. 46–49. [Google Scholar]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program. I. Grafting of Old Pineals in Young Mice Accelerates Their Aging. J. Anti. Aging Med. 2001, 4, 31–37. [Google Scholar] [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program: Life Prolongation in Pre-Aging Pinealectomized Mice. In Proceedings of the Annals of the New York Academy of Sciences; Wiley: Hoboken, NJ, USA, 2005; Volume 1057, pp. 133–144. [Google Scholar]
- Costa, R.M.; Filgueira, F.P.; Tostes, R.C.; Carvalho, M.H.C.; Akamine, E.H.; Lobato, N.S. H2O2 Generated from Mitochondrial Electron Transport Chain in Thoracic Perivascular Adipose Tissue Is Crucial for Modulation of Vascular Smooth Muscle Contraction. Vascul. Pharmacol. 2016, 84, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Li, H. The Role of Perivascular Adipose Tissue in Obesity-Induced Vascular Dysfunction. Br. J. Pharmacol. 2017, 174, 3425–3442. [Google Scholar] [CrossRef] [PubMed]
- Molcan, L.; Maier, A.; Zemančíková, A.; Gelles, K.; Török, J.; Zeman, M.; Ellinger, I. Expression of Melatonin Receptor 1 in Rat Mesenteric Artery and Perivascular Adipose Tissue and Vasoactive Action of Melatonin. Cell Mol. Neurobiol. 2021, 41, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Schepelmann, M.; Molcan, L.; Uhrova, H.; Zeman, M.; Ellinger, I. The Presence and Localization of Melatonin Receptors in the Rat Aorta. Cell Mol. Neurobiol. 2011, 31, 1257–1265. [Google Scholar] [CrossRef]
- Antonova, M.; Georgiev, A.; Totev, C. A Device for Implementation of the Forced Oscillation Method on Strip Preparations of Biological Tissues in Vitro in Prolonged Experiments. J. Theor. Appl. Mech. 2024; in press. [Google Scholar]
- Randall, R.B. Frequency Analysis; Bruel & Kjaer: Nærum, Denmark, 1980; ISBN 8787355078. [Google Scholar]
- Antonova, M.; Kasakov, L. Dynamic Viscoelastic Characteristics of Strips of Vital, Devitalized and Artificial Blood Vessels at Resonance. I: Natural Frequency, Dynamic Modulus of Elasticity, Coefficient of Viscosity. C. R. Acad. Bulg. Sci. 1999, 52, 123–126. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgiev, A.; Kaneva, M.; Shikova, L.; Mateeva, P.; Tchekalarova, J.; Antonova, M. In Vitro Effects of Fentanyl on Aortic Viscoelasticity in a Rat Model of Melatonin Deficiency. Int. J. Mol. Sci. 2024, 25, 5669. https://doi.org/10.3390/ijms25115669
Georgiev A, Kaneva M, Shikova L, Mateeva P, Tchekalarova J, Antonova M. In Vitro Effects of Fentanyl on Aortic Viscoelasticity in a Rat Model of Melatonin Deficiency. International Journal of Molecular Sciences. 2024; 25(11):5669. https://doi.org/10.3390/ijms25115669
Chicago/Turabian StyleGeorgiev, Andreyan, Maria Kaneva, Lyudmila Shikova, Polina Mateeva, Jana Tchekalarova, and Mariya Antonova. 2024. "In Vitro Effects of Fentanyl on Aortic Viscoelasticity in a Rat Model of Melatonin Deficiency" International Journal of Molecular Sciences 25, no. 11: 5669. https://doi.org/10.3390/ijms25115669
APA StyleGeorgiev, A., Kaneva, M., Shikova, L., Mateeva, P., Tchekalarova, J., & Antonova, M. (2024). In Vitro Effects of Fentanyl on Aortic Viscoelasticity in a Rat Model of Melatonin Deficiency. International Journal of Molecular Sciences, 25(11), 5669. https://doi.org/10.3390/ijms25115669