
Large-Scale Biogeographical Shifts of Abundance of Antibiotic Resistance Genes and Marine Bacterial Communities as Their Carriers along a Trophic Gradient

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Frequency of Antibiotic Resistance along the Trophic Gradient
2.2. Quantitative Analysis of ARGs and intl1
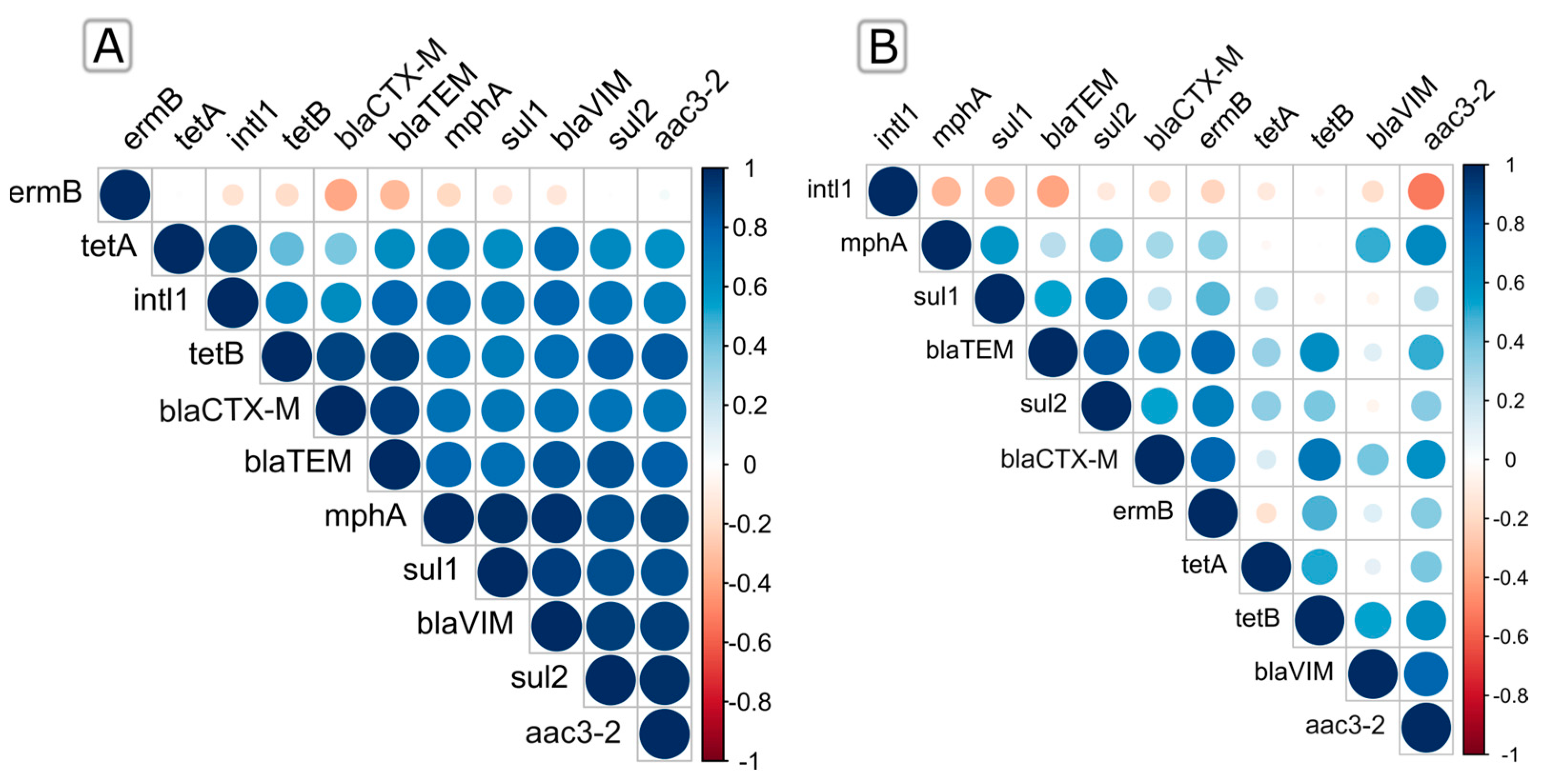
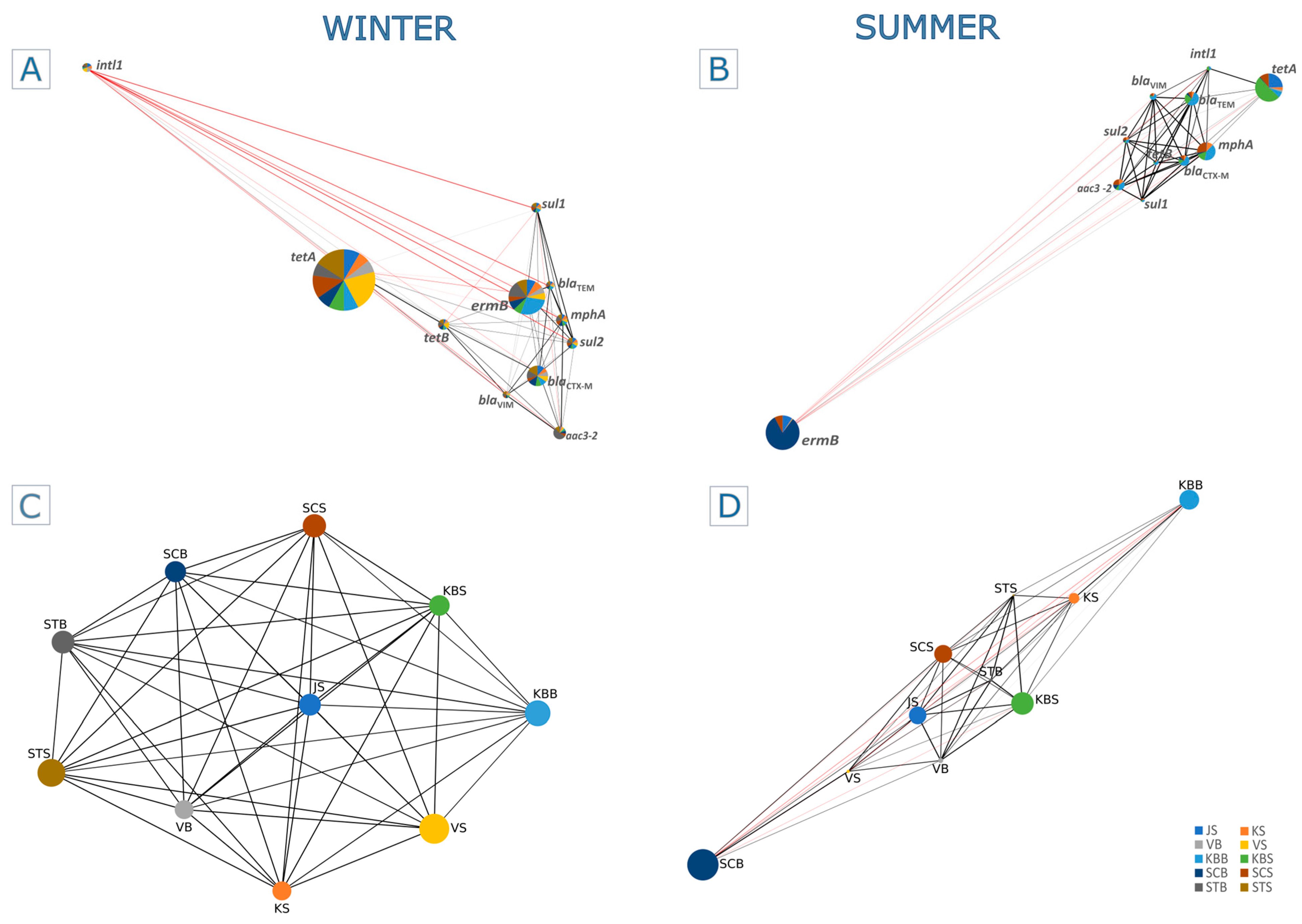
2.3. Seasonal and Spatial Co-Occurrence of Quantified ARGs and intl1
2.4. Relationship between Environmental Parameters and Gene Abundances
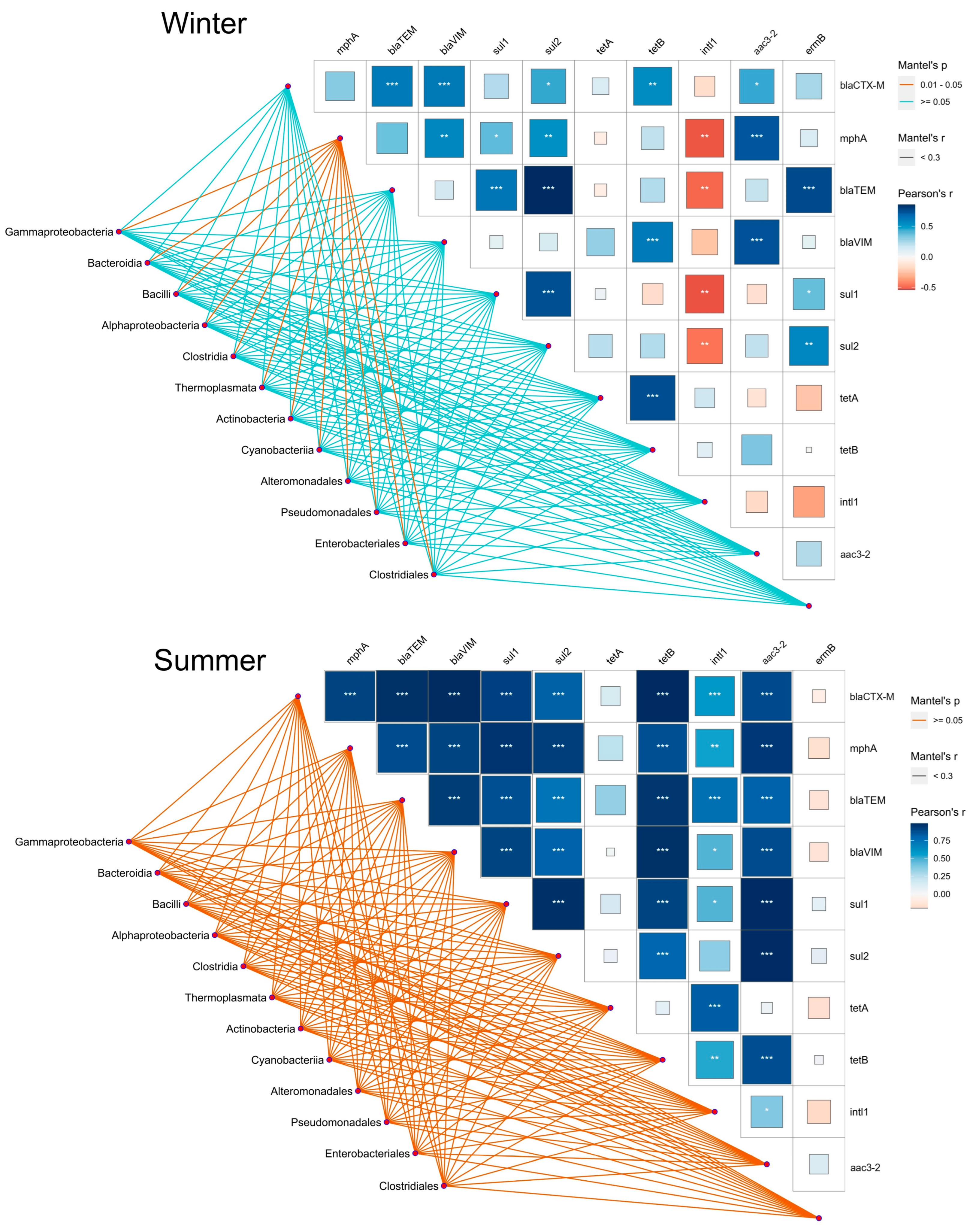
2.5. Association of Quantified ARGs with Bacterial Communities along the Trophic Gradient
3. Discussion
3.1. Seasonality Is a Driving Factor for Antibiotic Resistance and Distribution of ARGs
3.2. Correlation between ARGs, intl1, and Physico-Chemical Parameters Seasonally
3.3. Association of Quantified ARGs with Bacterial Communities along the Trophic Gradient
4. Materials and Methods
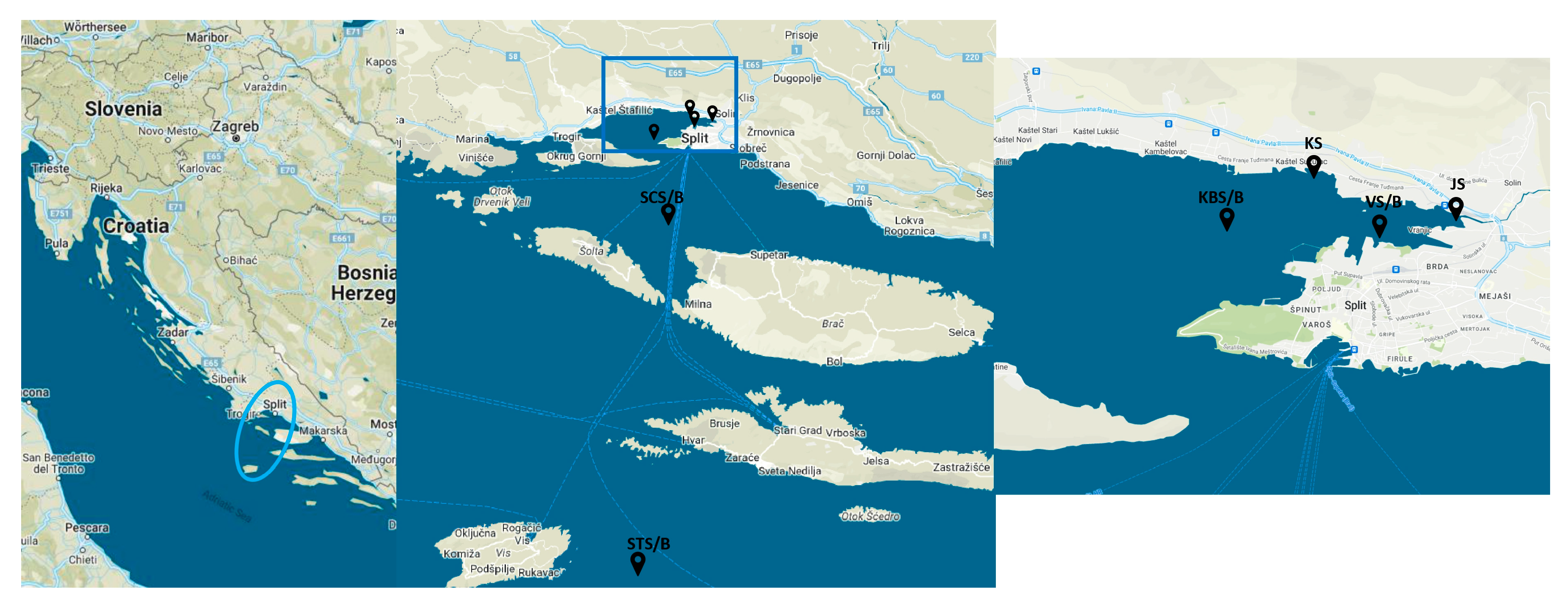
4.1. Study Area and Determination of Environmental and Biological Parameters
4.2. Antibiotic-Resistant Heterotrophic Bacterial Counts
4.3. DNA Extraction and Quantification of Target Genes
4.4. Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.L.; Chueiri, A.; O’Connor, L.; Duane, S.; Maguire, M.; Miliotis, G.; Cormican, M.; Hooban, B.; Leonard, A.; Gaze, W.H.; et al. Assessing the Impact of Recreational Water Use on Carriage of Antimicrobial Resistant Organisms. Sci. Total Environ. 2023, 888, 164201. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, Y.; Wu, J.; Wang, J.; Zhang, C.; Lin, Y. Occurrence and Spatial Distribution of Antibiotic Resistance Genes in the Bohai Sea and Yellow Sea Areas, China. Environ. Pollut. 2019, 252, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Xiao, H.; Li, Z.; Wang, J. Tetracyclines, Sulfonamides and Quinolones and Their Corresponding Resistance Genes in Coastal Areas of Beibu Gulf, China. Sci. Total Environ. 2020, 714, 136899. [Google Scholar] [CrossRef] [PubMed]
- Maravić, A.; Skočibušić, M.; Fredotović, Ž.; Šamanić, I.; Cvjetan, S.; Knezović, M.; Puizina, J. Urban Riverine Environment Is a Source of Multidrug-Resistant and ESBL-Producing Clinically Important Acinetobacter spp. Environ. Sci. Pollut. Res. 2016, 23, 3525–3535. [Google Scholar] [CrossRef] [PubMed]
- Kvesić, M.; Kalinić, H.; Dželalija, M.; Šamanić, I.; Andričević, R.; Maravić, A. Microbiome and Antibiotic Resistance Profiling in Submarine Effluent-Receiving Coastal Waters in Croatia. Environ. Pollut. 2022, 292, 118282. [Google Scholar] [CrossRef] [PubMed]
- Guidelines on Recreational Water Quality: Volume 1 Coastal and Fresh Waters. Available online: https://www.who.int/publications-detail-redirect/9789240031302 (accessed on 29 November 2023).
- Dželalija, M.; Kvesić-Ivanković, M.; Jozić, S.; Ordulj, M.; Kalinić, H.; Pavlinović, A.; Šamanić, I.; Maravić, A. Marine Resistome of a Temperate Zone: Distribution, Diversity, and Driving Factors across the Trophic Gradient. Water Res. 2023, 246, 120688. [Google Scholar] [CrossRef]
- Ali, L.; Wang, Y.-Q.; Zhang, J.; Ajmal, M.; Xiao, Z.; Wu, J.; Chen, J.-L.; Yu, D. Nutrient-Induced Antibiotic Resistance in Enterococcus Faecalis in the Eutrophic Environment. J. Glob. Antimicrob. Resist. 2016, 7, 78–83. [Google Scholar] [CrossRef]
- Huang, H.; Zheng, X.; Chen, Y.; Liu, H.; Wan, R.; Su, Y. Alkaline Fermentation of Waste Sludge Causes a Significant Reduction of Antibiotic Resistance Genes in Anaerobic Reactors. Sci. Total Environ. 2017, 580, 380–387. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Hu, H.-W.; Yan, H.; Wang, J.-T.; Lam, S.K.; Chen, Q.-L.; Chen, D.; He, J.-Z. Salinity as a Predominant Factor Modulating the Distribution Patterns of Antibiotic Resistance Genes in Ocean and River Beach Soils. Sci. Total Environ. 2019, 668, 193–203. [Google Scholar] [CrossRef]
- Yang, J.; Wang, H.; Roberts, D.J.; Du, H.-N.; Yu, X.-F.; Zhu, N.-Z.; Meng, X.-Z. Persistence of Antibiotic Resistance Genes from River Water to Tap Water in the Yangtze River Delta. Sci. Total Environ. 2020, 742, 140592. [Google Scholar] [CrossRef] [PubMed]
- Dželalija, M.; Kvesić, M.; Novak, A.; Fredotović, Ž.; Kalinić, H.; Šamanić, I.; Ordulj, M.; Jozić, S.; Goić Barišić, I.; Tonkić, M.; et al. Microbiome Profiling and Characterization of Virulent and Vancomycin-Resistant Enterococcus Faecium from Treated and Untreated Wastewater, Beach Water and Clinical Sources. Sci. Total Environ. 2023, 858, 159720. [Google Scholar] [CrossRef]
- Hernroth, B.; Lothigius, Å.; Bölin, I. Factors Influencing Survival of Enterotoxigenic Escherichia Coli, Salmonella Enterica (Serovar Typhimurium) and Vibrio Parahaemolyticus in Marine Environments. FEMS Microbiol. Ecol. 2009, 71, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Reda, R.M.; Ibrahim, R.E.; Ahmed, E.-N.G.; El-Bouhy, Z.M. Effect of Oxytetracycline and Florfenicol as Growth Promoters on the Health Status of Cultured Oreochromis Niloticus. Egypt. J. Aquat. Res. 2013, 39, 241–248. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Xiong, P.; Zhu, Q.; Liao, C.; Jiang, G. Occurrence, Fate, and Risk Assessment of Typical Tetracycline Antibiotics in the Aquatic Environment: A Review. Sci. Total Environ. 2021, 753, 141975. [Google Scholar] [CrossRef] [PubMed]
- Cuadrat, R.R.C.; Sorokina, M.; Andrade, B.G.; Goris, T.; Dávila, A.M.R. Global Ocean Resistome Revealed: Exploring Antibiotic Resistance Gene Abundance and Distribution in TARA Oceans Samples. GigaScience 2020, 9, giaa046. [Google Scholar] [CrossRef] [PubMed]
- Milaković, M.; Vestergaard, G.; González-Plaza, J.J.; Petrić, I.; Šimatović, A.; Senta, I.; Kublik, S.; Schloter, M.; Smalla, K.; Udiković-Kolić, N. Pollution from Azithromycin-Manufacturing Promotes Macrolide-Resistance Gene Propagation and Induces Spatial and Seasonal Bacterial Community Shifts in Receiving River Sediments. Environ. Int. 2019, 123, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Su, H.; Cao, Y.; Wang, L.; Hu, X.; Xu, W.; Xu, Y.; Li, Z.; Wen, G. Antibiotic Resistance Genes and Bacterial Community Dynamics in the Seawater Environment of Dapeng Cove, South China. Sci. Total Environ. 2020, 723, 138027. [Google Scholar] [CrossRef]
- Zahra, Q.; Gul, J.; Shah, A.R.; Yasir, M.; Karim, A.M. Antibiotic Resistance Genes Prevalence Prediction and Interpretation in Beaches Affected by Urban Wastewater Discharge. One Health 2023, 17, 100642. [Google Scholar] [CrossRef]
- Czekalski, N.; Sigdel, R.; Birtel, J.; Matthews, B.; Bürgmann, H. Does Human Activity Impact the Natural Antibiotic Resistance Background? Abundance of Antibiotic Resistance Genes in 21 Swiss Lakes. Environ. Int. 2015, 81, 45–55. [Google Scholar] [CrossRef]
- McKinney, C.W.; Loftin, K.A.; Meyer, M.T.; Davis, J.G.; Pruden, A. Tet and Sul Antibiotic Resistance Genes in Livestock Lagoons of Various Operation Type, Configuration, and Antibiotic Occurrence. Environ. Sci. Technol. 2010, 44, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, W.; Xu, C.; Wei, B.; Wang, J. Antibiotic Resistance Genes in Lakes from Middle and Lower Reaches of the Yangtze River, China: Effect of Land Use and Sediment Characteristics. Chemosphere 2017, 178, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Fresia, P.; Antelo, V.; Salazar, C.; Giménez, M.; D’Alessandro, B.; Afshinnekoo, E.; Mason, C.; Gonnet, G.H.; Iraola, G. Urban Metagenomics Uncover Antibiotic Resistance Reservoirs in Coastal Beach and Sewage Waters. Microbiome 2019, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Haldar, A.; Bhattacharyya, M.; Ghosh, A. Anthropogenic Influence Shapes the Distribution of Antibiotic Resistant Bacteria (ARB) in the Sediment of Sundarban Estuary in India. Sci. Total Environ. 2019, 647, 1626–1639. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Yasir, M.; Bibi, F.; Abujamel, T.S.; Hashem, A.M.; Sohrab, S.S.; Al-Ansari, A.; Al-Sofyani, A.A.; Al-Ghamdi, A.K.; Al-sieni, A.; et al. Taxonomic Diversity of Antimicrobial-Resistant Bacteria and Genes in the Red Sea Coast. Sci. Total Environ. 2019, 677, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.-C. Integron Involvement in Environmental Spread of Antibiotic Resistance. Front. Microbiol. 2012, 3, 119. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Variatza, E.; Balcazar, J.L. The Role of Aquatic Ecosystems as Reservoirs of Antibiotic Resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef]
- Chen, B.; Liang, X.; Huang, X.; Zhang, T.; Li, X. Differentiating Anthropogenic Impacts on ARGs in the Pearl River Estuary by Using Suitable Gene Indicators. Water Res. 2013, 47, 2811–2820. [Google Scholar] [CrossRef]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. MMBR 2014, 78, 257–277. [Google Scholar] [CrossRef]
- Ma, L.; Li, A.-D.; Yin, X.-L.; Zhang, T. The Prevalence of Integrons as the Carrier of Antibiotic Resistance Genes in Natural and Man-Made Environments. Environ. Sci. Technol. 2017, 51, 5721–5728. [Google Scholar] [CrossRef]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene Cassettes and Cassette Arrays in Mobile Resistance Integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling Antibiotic Resistance: The Environmental Framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Šantić, D.; Krstulović, N.; Šolić, M.; Kušpilić, G. HNA and LNA Bacteria in Relation to the Activity of Heterotrophic Bacteria. Acta Adriat. 2012, 53, 25–40. [Google Scholar]
- Jacquet, S.; Miki, T.; Noble, R.; Peduzzi, P.; Wilhelm, S. Viruses in Aquatic Ecosystems: Important Advancements of the Last 20 Years and Prospects for the Future in the Field of Microbial Oceanography and Limnology. Adv. Oceanogr. Limnol. 2010, 1, 97–141. [Google Scholar] [CrossRef]
- Bourdonnais, E.; Colcanap, D.; Le Bris, C.; Brauge, T.; Midelet, G. Occurrence of Indicator Genes of Antimicrobial Resistance Contamination in the English Channel and North Sea Sectors and Interactions with Environmental Variables. Front. Microbiol. 2022, 13, 1750. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, L.; Lu, Z.; He, C.; Li, Q.; Na, G. Complex Migration of Antibiotic Resistance in Natural Aquatic Environments. Environ. Pollut. 2018, 232, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.A.S.; Xu, J.; Moore, J.E.; Blair, I.S.; McDowell, D.A. Environmental Stress and Antibiotic Resistance in Food-Related Pathogens. Appl. Environ. Microbiol. 2007, 73, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Keen, P.L.; Knapp, C.W.; Hall, K.J.; Graham, D.W. Seasonal Dynamics of Tetracycline Resistance Gene Transport in the Sumas River Agricultural Watershed of British Columbia, Canada. Sci. Total Environ. 2018, 628–629, 490–498. [Google Scholar] [CrossRef]
- Ordulj, M.; Krstulović, N.; Šantić, D.; Jozić, S.; Šolić, Μ. Distribution of Marine Viruses in the Central and South Adriatic Sea. Mediterr. Mar. Sci. 2015, 2015, 65–72. [Google Scholar] [CrossRef][Green Version]
- Zhang, T.; Ji, Z.; Li, J.; Yu, L. Metagenomic Insights into the Antibiotic Resistome in Freshwater and Seawater from an Antarctic Ice-Free Area. Environ. Pollut. 2022, 309, 119738. [Google Scholar] [CrossRef]
- Ohore, O.E.; Wei, Y.; Wang, Y.; Nwankwegu, A.S.; Wang, Z. Tracking the Influence of Antibiotics, Antibiotic Resistomes, and Salinity Gradient in Modulating Microbial Community Assemblage of Surface Water and the Ecological Consequences. Chemosphere 2022, 305, 135428. [Google Scholar] [CrossRef] [PubMed]
- Uyaguari-Díaz, M.I.; Croxen, M.A.; Luo, Z.; Cronin, K.I.; Chan, M.; Baticados, W.N.; Nesbitt, M.J.; Li, S.; Miller, K.M.; Dooley, D.; et al. Human Activity Determines the Presence of Integron-Associated and Antibiotic Resistance Genes in Southwestern British Columbia. Front. Microbiol. 2018, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- González-Plaza, J.J.; Blau, K.; Milaković, M.; Jurina, T.; Smalla, K.; Udiković-Kolić, N. Antibiotic-Manufacturing Sites Are Hot-Spots for the Release and Spread of Antibiotic Resistance Genes and Mobile Genetic Elements in Receiving Aquatic Environments. Environ. Int. 2019, 130, 104735. [Google Scholar] [CrossRef] [PubMed]
- Adelowo, O.O.; Ikhimiukor, O.O.; Knecht, C.; Vollmers, J.; Bhatia, M.; Kaster, A.-K.; Müller, J.A. A Survey of Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Urban Wetlands in Southwestern Nigeria as a Step towards Generating Prevalence Maps of Antimicrobial Resistance. PLoS ONE 2020, 15, e0229451. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarawi, H.A.; Habibi, N.; Uddin, S.; Jha, A.N.; Al-Sarawi, M.A.; Lyons, B.P. Antibiotic Resistance Mediated by Escherichia Coli in Kuwait Marine Environment as Revealed through Genomic Analysis. Antibiotics 2023, 12, 1366. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Yuan, K.; Liang, X.; Chen, X.; Zhao, Z.; Yang, Y.; Zou, S.; Luan, T.; Chen, B. Occurrences and Distribution of Sulfonamide and Tetracycline Resistance Genes in the Yangtze River Estuary and Nearby Coastal Area. Mar. Pollut. Bull. 2015, 100, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Tuyet Hoa, T.T.; Harada, K.; Warisaya, M.; Asayama, M.; Hinenoya, A.; Lee, J.W.; Phu, T.M.; Ueda, S.; Sumimura, Y.; et al. Water Metagenomic Analysis Reveals Low Bacterial Diversity and the Presence of Antimicrobial Residues and Resistance Genes in a River Containing Wastewater from Backyard Aquacultures in the Mekong Delta, Vietnam. Environ. Pollut. 2017, 222, 294–306. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Liu, P.; Sun, Y.; Song, Z.; Hu, X. Characteristics of Bacterial Community Structure and Function Associated with Nutrients and Heavy Metals in Coastal Aquaculture Area. Environ. Pollut. 2021, 275, 116639. [Google Scholar] [CrossRef]
- CLSI Publishes M100—Performance Standards for Antimicrobial Susceptibility Testing, 31st Edition. Available online: https://clsi.org/about/press-releases/clsi-publishes-m100-performance-standards-for-antimicrobial-susceptibility-testing-31st-edition/ (accessed on 29 November 2023).
- Eucast: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 29 November 2023).
- Weber, N.; Liou, D.; Dommer, J.; MacMenamin, P.; Quiñones, M.; Misner, I.; Oler, A.J.; Wan, J.; Kim, L.; Coakley McCarthy, M.; et al. Nephele: A Cloud Platform for Simplified, Standardized and Reproducible Microbiome Data Analysis. Bioinform. Oxf. Engl. 2018, 34, 1411–1413. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dželalija, M.; Fredotović, Ž.; Udiković-Kolić, N.; Kalinić, H.; Jozić, S.; Šamanić, I.; Ordulj, M.; Maravić, A. Large-Scale Biogeographical Shifts of Abundance of Antibiotic Resistance Genes and Marine Bacterial Communities as Their Carriers along a Trophic Gradient. Int. J. Mol. Sci. 2024, 25, 654. https://doi.org/10.3390/ijms25010654
Dželalija M, Fredotović Ž, Udiković-Kolić N, Kalinić H, Jozić S, Šamanić I, Ordulj M, Maravić A. Large-Scale Biogeographical Shifts of Abundance of Antibiotic Resistance Genes and Marine Bacterial Communities as Their Carriers along a Trophic Gradient. International Journal of Molecular Sciences. 2024; 25(1):654. https://doi.org/10.3390/ijms25010654
Chicago/Turabian StyleDželalija, Mia, Željana Fredotović, Nikolina Udiković-Kolić, Hrvoje Kalinić, Slaven Jozić, Ivica Šamanić, Marin Ordulj, and Ana Maravić. 2024. "Large-Scale Biogeographical Shifts of Abundance of Antibiotic Resistance Genes and Marine Bacterial Communities as Their Carriers along a Trophic Gradient" International Journal of Molecular Sciences 25, no. 1: 654. https://doi.org/10.3390/ijms25010654
APA StyleDželalija, M., Fredotović, Ž., Udiković-Kolić, N., Kalinić, H., Jozić, S., Šamanić, I., Ordulj, M., & Maravić, A. (2024). Large-Scale Biogeographical Shifts of Abundance of Antibiotic Resistance Genes and Marine Bacterial Communities as Their Carriers along a Trophic Gradient. International Journal of Molecular Sciences, 25(1), 654. https://doi.org/10.3390/ijms25010654