
Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of c-MET/MAPK Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Compounds 1 and 1a Inhibited the Proliferation of Human Erythroid Leukemia Cells
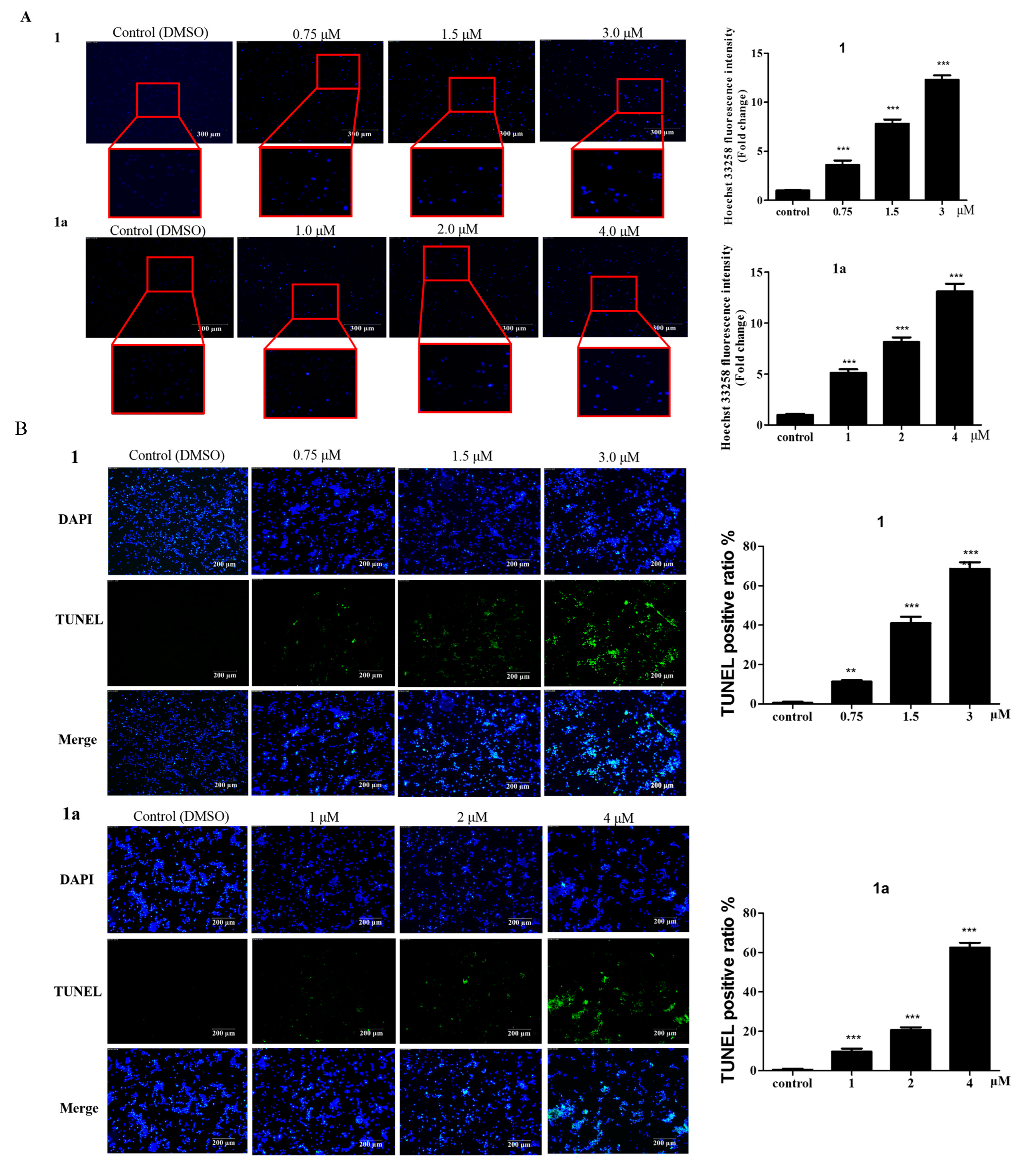
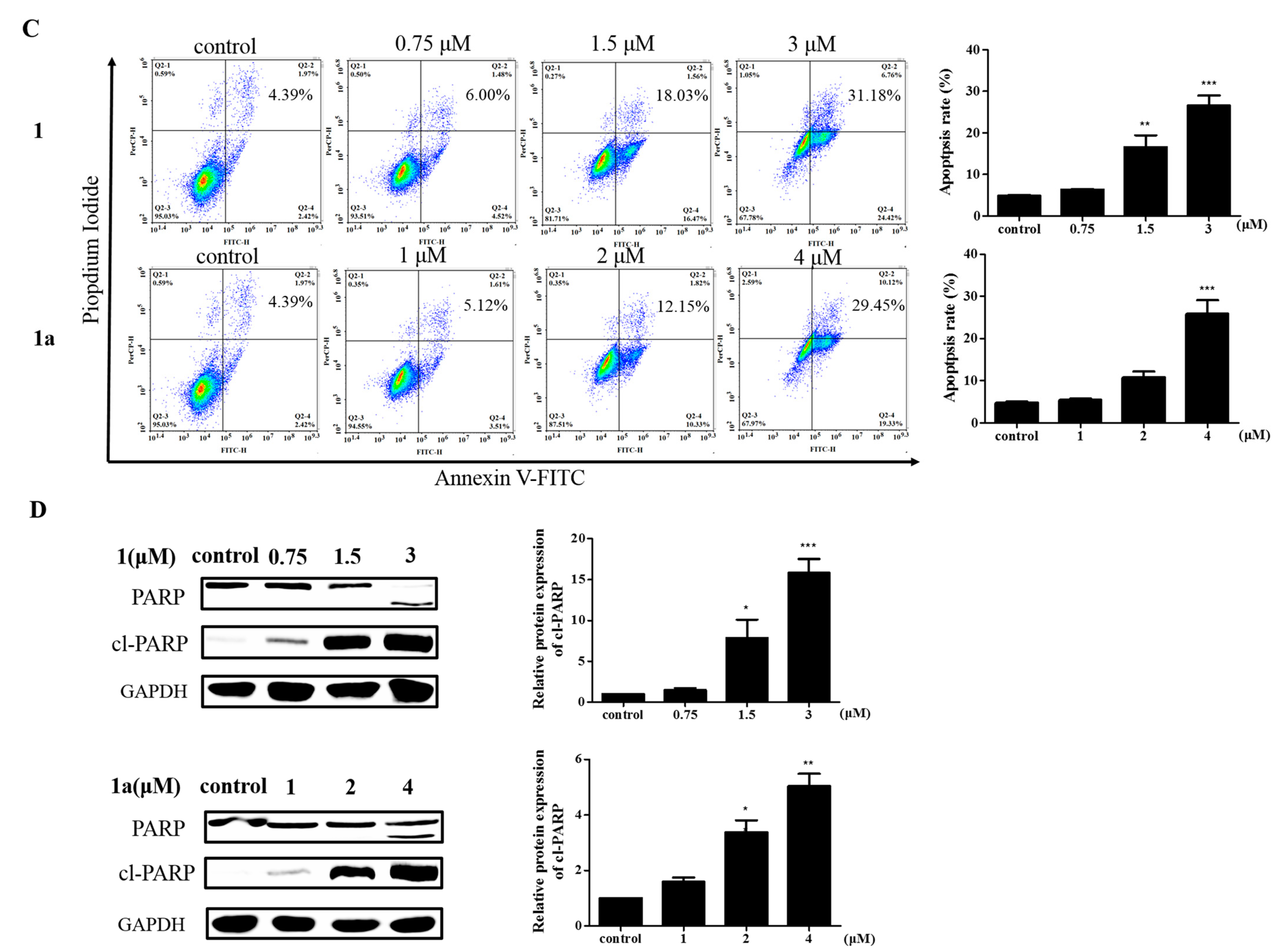
2.2. The Effect of 1 and 1a on Cell Apoptosis
2.3. Compounds 1 and 1a Exerted Pharmacological Antitumor Effects by Blocking MAPK Signaling
2.4. Compounds 1 and 1a Antagonize the MAPK Signaling Pathway to Induce Apoptosis by Inhibiting c-MET Protein Expression
2.5. Compounds 1 and 1a Suppress the MAPK, PI3K/AKT, and STAT3 Signaling Pathways by Decreasing the mRNA and Protein Levels of c-MET
3. Materials and Methods
3.1. Cell Lines and Culture
3.2. Cell Viability Assays
3.3. Apoptosis Analysis
3.4. Hoechst 33258 Staining
3.5. TUNEL Staining
3.6. Western Blotting
3.7. RNA-Sequencing (RNA-Seq) Experiment and Analysis
3.8. Overexpression of c-MET
3.9. Real-Time q-PCR
3.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lou, G.; Wang, J.; Hu, J.; Gan, Q.; Peng, C.; Xiong, H.; Huang, Q. Sanguinarine: A Double-Edged Sword of Anticancer and Carcinogenesis and Its Future Application Prospect. Anti-Cancer Agents Med. Chem. 2021, 21, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Guan, G.; Wang, H. The Anticancer Effect of Sanguinarine: A Review. Curr. Pharm. Des. 2018, 24, 2760–2764. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, N.; Kou, S.; Gao, L.; Peng, B.; Dai, Y.; Zheng, J. Sanguinarine synergistically potentiates aminoglycoside-mediated bacterial killing. Microb. Biotechnol. 2022, 15, 2055–2070. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.B.; Kanungo, N.; Chainy, G.B.N.; Devaraji, V.; Das, S.K.; Dandapat, J. A Computational Insight on the Inhibitory Potential of 8-Hydroxydihydr of Sanguinarine (8-HDS), a Pyridone Containing Analog of Sanguinarine, Against SARS CoV2. Chem Biodivers. 2022, 19, e202200266. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Fan, T.; Li, W.; Huang, H. The Anti-inflammatory Effects of Sanguinarine and Its Modulation of in Flammatory Mediators from Peritoneal Macrophages. Eur. J. Pharmacol. 2012, 689, 262–269. [Google Scholar] [CrossRef]
- Khan, A.Q.; Rashid, K.; AlAmodi, A.A.; Agha, M.V.; Akhtar, S.; Hakeem, I.; Raza, S.S.; Uddin, S. Reactive oxygen species (ROS) in cancer pathogenesis and therapy: An update on the role of ROS in anticancer action of benzophenanthridine alkaloids. Biomed. Pharmacother. 2021, 143, 112142. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Wang, J.; Li, L.; Zhang, Y.; Jin, M.; Chen, X.; Sun, C.; Wang, R.; Liu, K. Cardiotoxicity of sanguinarine via regulating apoptosis and MAPK pathways in zebrafish and HL1 cardiomyocytes. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 252, 109228. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef]
- Tang, Y.; Xu, X.; Li, J.; Deng, L.; Mu, S. Synthesis and Antileukemia Activity Evaluation of Benzophenanthridine Alkaloid Derivatives. Molecules 2022, 27, 3934. [Google Scholar] [CrossRef]
- Laines-Hidalgo, J.I.; Muñoz-Sánchez, J.A.; Loza-Müller, L.; Vázquez-Flota, F. An Update of the Sanguinarine and Benzophenanthridine Alkaloids’ Biosynthesis and Their Applications. Molecules 2022, 27, 1378. [Google Scholar] [CrossRef]
- Yu, C.; Li, P.; Wang, Y.-X.; Zhang, K.-G.; Zheng, Z.-C.; Liang, L.-S. Sanguinarine Attenuates Neuropathic Pain by Inhibiting P38 MAPK Activated Neuroinflammation in Rat Model. Drug Des. Dev. Ther. 2020, 14, 4725–4733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Leng, T.; Zhang, Q.; Nie, X.; Yang, L. Sanguinarine Inhibits Epithelial Ovarian Cancer Development via Regulating Long Non-coding RNA CASC2-EIF4A3 Axis and/or Inhibiting NF-κB Signaling or PI3K/AKT/mTOR Pathway. Biomed. Pharmacother. 2018, 102, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Achkar, I.W.; Siveen, K.S.; Khan, A.Q.; Ahmed, E.I.; Sahir, F.; Jerobin, J.; Raza, A.; Merhi, M.; Elsabah, H.M.; et al. Sanguinarine Induces Apoptosis Pathway in Multiple Myeloma Cell Lines via Inhibition of the JAK2/STAT3 Signaling. Front Oncol. 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, X.; Deng, J.; Zheng, H.; Liu, W.; Chen, S.; Tian, J.; Wang, F. p53-dependent upregulation of miR-16-2 by sanguinarine induces cell cycle arrest and apoptosis in hepatocellular carcinoma. Cancer Lett. 2019, 459, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ye, W.; Wang, Y.-D.; Chen, W.-D. HGF/c-Met: A Key Promoter in Liver Regeneration. Front. Pharmacol. 2022, 13, 808855. [Google Scholar] [CrossRef]
- Stefani, C.; Miricescu, D.; Stanescu, I.; Nica, R.I.; Greabu, M.; Totan, A.R.; Jinga, M. (Growth Factors, PI3K/AKT/mTOR and MAPK Signaling Pathways in Colorectal Cancer Pathogenesis: Where Are we Now? Int. J. Mol. Sci. 2021, 22, 10260. [Google Scholar] [CrossRef]
- Blangé, D.; Stroes, C.I.; Derks, S.; Bijlsma, M.F.; van Laarhoven, H.W. Resistance Mechanisms to HER2-targeted Therapy in Aastroesophageal Adenocarcinoma: A Systematic Review. Cancer Treat. Rev. 2022, 108, 102418. [Google Scholar] [CrossRef]
- Uchikawa, E.; Chen, Z.; Xiao, G.-Y.; Zhang, X.; Bai, X.-C. Structural basis of the activation of c-MET receptor. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Damghani, T.; Elyasi, M.; Pirhadi, S.; Haghighijoo, Z.; Ghazi, S. Type II c-Met inhibitors: Molecular insight into crucial interactions for effective inhibition. Mol. Divers. 2021, 26, 1411–1423. [Google Scholar] [CrossRef]
- Wang, H.; Rao, B.; Lou, J.; Li, J.; Liu, Z.; Li, A.; Cui, G.; Ren, Z.; Yu, Z. The Function of the HGF/c-Met Axis in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2020, 8, 55. [Google Scholar] [CrossRef]
- Ci, T.; Zhang, W.; Qiao, Y.; Li, H.; Zang, J.; Li, H.; Feng, N.; Gu, Z. Delivery strategies in treatments of leukemia. Chem. Soc. Rev. 2022, 51, 2121–2144. [Google Scholar] [CrossRef] [PubMed]
- Roloff, G.W.; Odenike, O.; Bajel, A.; Wei, A.H.; Foley, N.; Uy, G.L. Contemporary Approach to Acute Myeloid Leukemia Therapy in 2022. Am. Soc. Clin. Oncol. Educ. Book 2022, 568–583. [Google Scholar] [CrossRef]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S.; Liu, C.; Li, Q.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2020, 263, 128346. [Google Scholar] [CrossRef]
- Feng, H.; Hu, L.; Zhu, H.; Tao, L.; Wu, L.; Zhao, Q.; Gao, Y.; Gong, Q.; Mao, F.; Li, X.; et al. Repurposing antimycotic ciclopirox olamine as a promising anti-ischemic stroke agent. Acta Pharm. Sin. B 2019, 10, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Zhang, Y.J.; Lei, J.; Li, S.Q.; He, L.J.; Tang, D.Y.; Xu, C.; Zhang, L.T.; Wen, J.; Lin, H.K.; et al. Discovery of Fused Benzimidazole-imidazole Autophagic Flux Inhibitors for Treatment of Triple-negative Treast Cancer. Eur. J. Med. Chem. 2022, 240, 114565. [Google Scholar] [CrossRef] [PubMed]
- Zaky, M.Y.; Liu, X.; Wang, T.; Wang, S.; Liu, F.; Wang, D.; Wu, Y.; Zhang, Y.; Guo, D.; Sun, Q.; et al. Dynasore potentiates c-Met inhibitors against hepatocellular carcinoma through destabilizing c-Met. Arch. Biochem. Biophys. 2020, 680, 108239. [Google Scholar] [CrossRef]
- Rose, M.; Burgess, J.T.; O’byrne, K.; Richard, D.J.; Bolderson, E. PARP Inhibitors: Clinical Relevance, Mechanisms of Action and Tumor Resistance. Front. Cell Dev. Biol. 2020, 8, 564601. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Yang, M.; Bai, Y.; Ren, H.; Zou, Y.; Yao, Z.; Zhang, B.; Li, Y. RNA-Seq Based Transcriptome Analysis of Endothelial Differentiation of Bone Marrow Mesenchymal Stem Cells. Eur. J. Vasc. Endovasc. Surg. 2019, 59, 834–842. [Google Scholar] [CrossRef]
- Lu, S.; Zhu, N.; Guo, W.; Wang, X.; Li, K.; Yan, J.; Jiang, C.; Han, S.; Xiang, H.; Xiaohan, W.; et al. RNA-Seq revealed a circular RNA-microRNA-mRNA Regulatory Network in Hantaan Virus Infection. Front. Cell Infect. Microbiol. 2022, 10, 97. [Google Scholar] [CrossRef]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Raj, S.; Kesari, K.K.; Kumar, A.; Rathi, B.; Sharma, A.; Gupta, P.K.; Jha, S.K.; Jha, N.K.; Slama, P.; Roychoudhury, S.; et al. Molecular mechanism(s) of regulation(s) of c-MET/HGF signaling in head and neck cancer. Mol. Cancer 2022, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Besser, D.; Bardelli, A.; Didichenko, S.; Thelen, M.; Comoglio, P.M.; Ponzetto, C.; Nagamine, Y. Regulation of the urokinase-type plasminogen activator gene by the oncogene Tpr-Met involves GRB2. Oncogene 1997, 14, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Inagaki, Y.; Song, P.; Qu, X.; Kokudo, N.; Tang, W. Targeting c-Met as a promising strategy for the treatment of hepatocellular carcinoma. Pharmacol. Res. 2012, 65, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Pons, E.; Uphoff, C.; Drexler, H. Expression of hepatocyte growth factor and its receptor c-met in human leukemia-lymphoma cell lines. Leuk. Res. 1998, 22, 797–804. [Google Scholar] [CrossRef]
- Paiva, R.S.; Gomes, I.; Casimiro, S.; Fernandes, I.; Costa, L. c-Met expression in renal cell carcinoma with bone metastases. J. Bone Oncol. 2020, 25, 100315. [Google Scholar] [CrossRef]
- Hu, Y.; Gong, C.; Li, Z.; Liu, J.; Chen, Y.; Huang, Y.; Luo, Q.; Wang, S.; Hou, Y.; Yang, S.; et al. Demethylase ALKBH5 suppresses invasion of gastric cancer via PKMYT1 m6A modification. Mol. Cancer 2022, 21, 1–15. [Google Scholar] [CrossRef]
- Sheng, W.; Guo, W.; Lu, F.; Liu, H.; Xia, R.; Dong, F. Upregulation of Linc00284 Promotes Lung Cancer Progression by Regulating the miR-205-3p/c-Met Axis. Front. Genet. 2021, 12, 694571. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Deng, L.; Tang, Y.; Li, J.; Zhong, T.; Hao, X.; Fan, Y.; Mu, S. Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of c-MET/MAPK Signaling. Int. J. Mol. Sci. 2023, 24, 8113. https://doi.org/10.3390/ijms24098113
Xu X, Deng L, Tang Y, Li J, Zhong T, Hao X, Fan Y, Mu S. Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of c-MET/MAPK Signaling. International Journal of Molecular Sciences. 2023; 24(9):8113. https://doi.org/10.3390/ijms24098113
Chicago/Turabian StyleXu, Xinglian, Lulu Deng, Yaling Tang, Jiang Li, Ting Zhong, Xiaojiang Hao, Yanhua Fan, and Shuzhen Mu. 2023. "Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of c-MET/MAPK Signaling" International Journal of Molecular Sciences 24, no. 9: 8113. https://doi.org/10.3390/ijms24098113
APA StyleXu, X., Deng, L., Tang, Y., Li, J., Zhong, T., Hao, X., Fan, Y., & Mu, S. (2023). Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of c-MET/MAPK Signaling. International Journal of Molecular Sciences, 24(9), 8113. https://doi.org/10.3390/ijms24098113