
Biological Assay to Determine Gonadotropin Potency: From In Vivo to In Vitro Sustainable Method


Abstract
1. Introduction
2. Results
2.1. Acid Enrichment
2.1.1. Dose–Response Curves
2.1.2. Physicochemical Characteristics
2.1.3. Specific Activity
2.2. Basic Enrichment
2.2.1. Dose–Response Curves
2.2.2. Physicochemical Characteristics
2.2.3. Specific Activity
2.3. Oxidation
2.3.1. Dose–Response Curves
2.3.2. Physicochemical Characteristics
2.3.3. Specific Activity
2.4. Acid pH Stress
2.4.1. Dose–Response Curves
2.4.2. Physicochemical Characteristics
2.4.3. Specific Activity
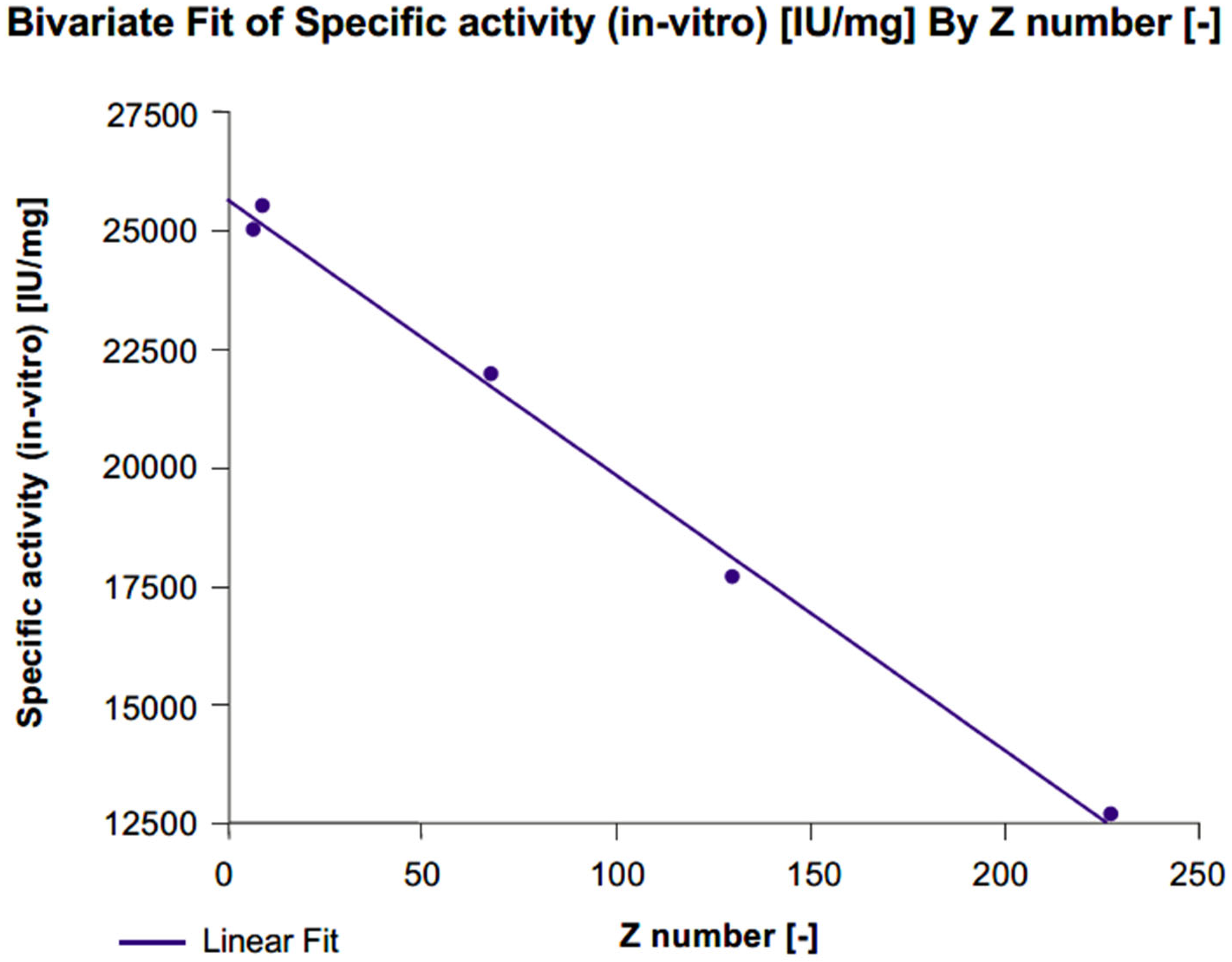
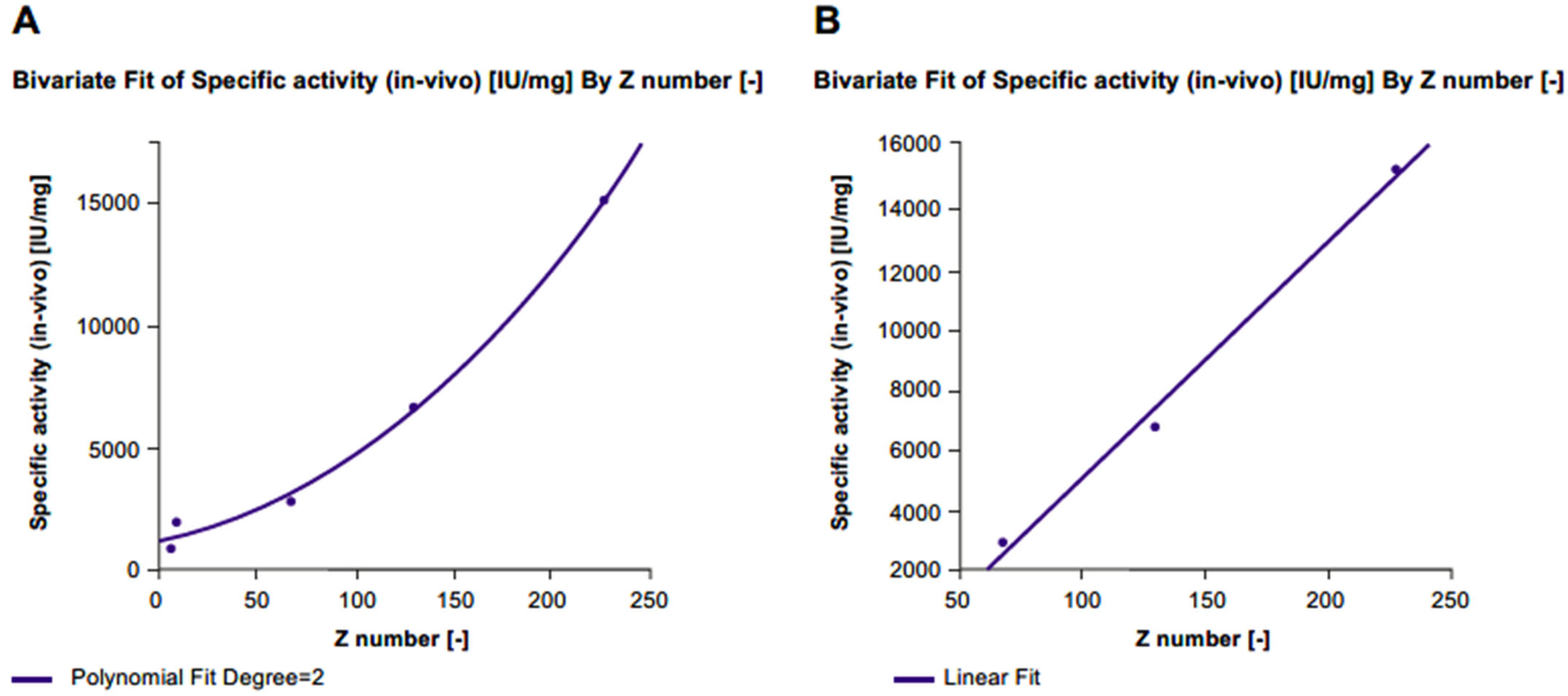
2.5. Total Desialylation
2.5.1. Dose–Response Curves
2.5.2. Physicochemical Characteristics
2.5.3. Specific Activity
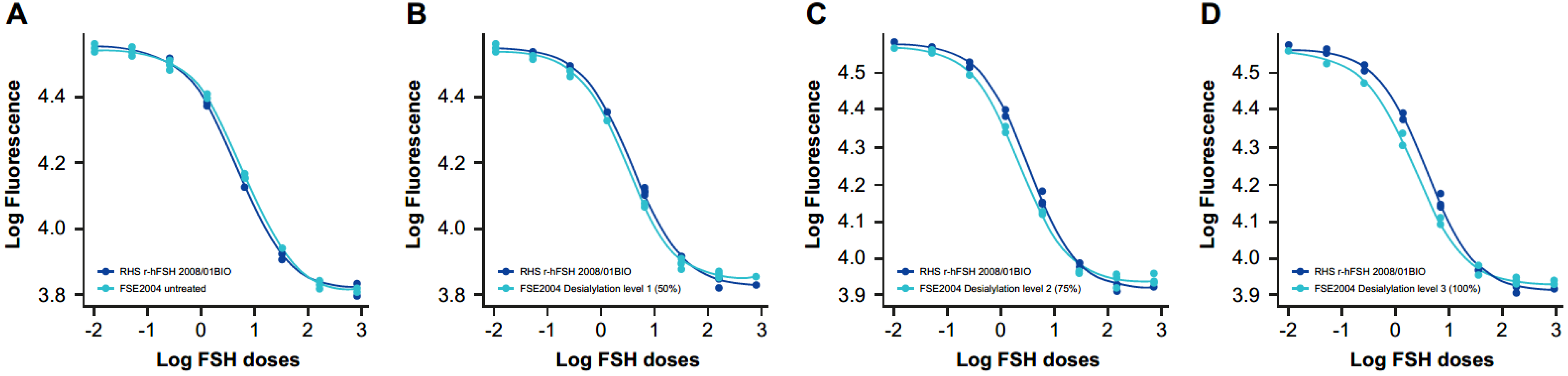
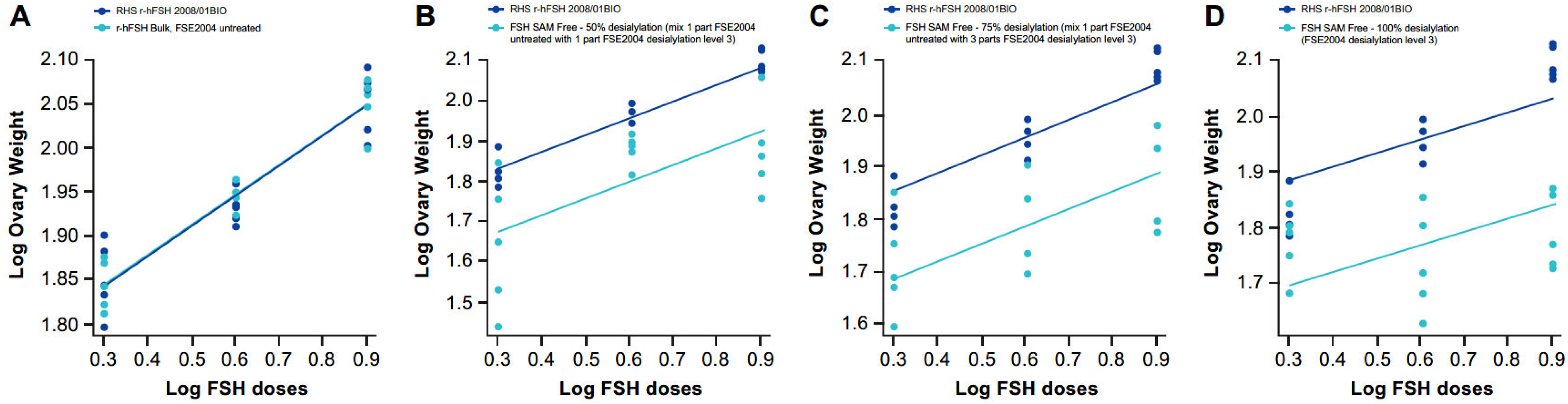
2.6. Different Levels of Sialylation
2.6.1. Dose–Response Curves
2.6.2. Physicochemical Characteristics
2.6.3. Specific Activity
2.7. Desialylation and Degalactosylation
2.7.1. Dose–Response Curves
2.7.2. Physicochemical Characteristics
2.7.3. Specific Activity
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Materials
4.3. Forced Degraded and Molecular Variants
4.4. Determination of Protein Content
4.5. Determination of FSH Potency
4.5.1. In Vitro Bioassay to Assess r-hFSH Biopotency
4.5.2. In Vivo Bioassay to Assess r-hFSH Biopotency
4.6. Calibration of Reference House Standard for the In Vitro Bioassay
4.7. Glycopeptide Mapping
4.8. Peptide Mapping
4.9. Glycan Mapping
4.10. Data Analysis
4.10.1. Potency of Treated and Untreated Samples Obtained by In Vivo and In Vitro Bioassays
4.10.2. In Vitro Bioassay
4.10.3. In Vivo Bioassay
4.10.4. Comparison between In Vivo and In Vitro Specific Activity of Treated and Untreated Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| Four-parameter logistic equation | Also referred to as a dose–response curve. The equation contains four parameters or variables related to the graphical properties of the curve: the bottom and top plateaus of the curve, the half maximal effective concentration (EC50) and the slope factor. |
| Antennarity | The number of branching glycans (bi-, tri- or tetra-antennarity) influences acidity and can impact the binding of the follicle-stimulating hormone receptor (FSHR). Indeed, bulky and extended glycans may result in a delayed receptor response while relatively smaller and more compact follicle-stimulating hormone (FSH) glycoforms (e.g., the bi-antennary at αAsn-52) have more rapid FSHR binding [11]. The higher the number of antennae, the greater the probability of having complete glycan moieties (i.e., with galactose and sialic acid attached). In fact, sialic acid can be added only if a galactose molecule is present in the carbohydrate chain of the terminal branch. Therefore, the number of antennae is indirectly positively correlated with FSH glycoform acidity through sialylation. Alternatively, the sequential addition of N-acetylgalactosamine (GalNAc) and sulfate produces sulfated oligosaccharides [27]. |
| Biological activity | The specific ability or capacity of a product to achieve a defined biological effect. A valid biological assay to measure the biological activity should be provided by the manufacturer [7]. Biological activity depends on the assay used to measure it. |
| Biopotency/potency | The quantitative measure of biological activity based on the attributes of the product (e.g., the nature and quantity of product-related substances, product-related impurities and process-related impurities) that is linked to the relevant biological properties (e.g., biological activity). A valid biological assay (e.g., in vitro assay or in vivo animal assay or biochemical assays) to measure the biological activity (biopotency) should be provided by the manufacturer of the product. When a valid bioassay is not suitable to measure the biological activity, potency may refer to the quantity of protein (see definition of Quantity in this Glossary) [7]. |
| Glycans (also called “oligosaccharides”) | Carbohydrate-based polymers that are essential for the structure, energy storage, receptor binding affinity and clearance of glycoproteins |
| Glycoforms | Variants (isoforms) of a glycoprotein that have different glycans attached, and/or are glycosylated at different amino acid residues [11]. |
| Glycoproteins | Proteins that contain glycans covalently attached to amino acid side-chains. This post-translational process occurs in the cell’s endoplasmic reticulum (ER) and Golgi apparatus, and is known as “glycosylation” [37]. |
| Glycosylation | The addition of glycans (N- and O-glycans) to proteins. It is a highly regulated mechanism of secondary protein processing within cells, and affects the three-dimensional configuration of proteins, as well as their function and stability. Glycosylation is critical for the action of glycoproteins as well as for their pharmacokinetics and pharmacodynamics (PK/PD) [11]. |
| Inflection point | A point on a curve at which the sign of the curvature changes. |
| In-process control (test) | In-process tests are tests which may be performed during the manufacture of either the drug substance or drug product, rather than as part of the formal battery of tests which are conducted prior to release [7,38]. |
| International standard | The World Health Organization (WHO) international standards and reference materials provide a common set of standards that are used to ensure the quality of biological medicines worldwide. Currently, the reference standard to measure FSH biopotency is follitropin alfa [39]. |
| International Unit (IU) | For each substance, there is international agreement on the biological effect that is expected for 1 IU based on the potency of a reference standard (see also Biopotency/Potency) [40]. |
| International IU of FSH | The activity contained in a stated amount of the International Standard (IS) of follitropin-alfa [15]. The equivalence in IU of the IS is stated by the WHO for any new certified IS [41]. The first WHO IS defined 1 IU as the amount of FSH exerting an activity corresponding to 0.11388 mg of pure Human Urinary FSH [16,42]. Each new standard is calibrated and validated according to the WHO Expert Committee of International Standard. The replacement IU is defined by the contents of the ampoule of the new standard. Every effort is made to maintain the continuity of the IU, but the replacement IU is not formally traceable to the first IS [41]. |
| Parallel Line Analysis (PLA) software | A statistical test for parallelism. Commonly used to compare dose–response curves to estimate the potency of a test substance relative to the potency of a standard. |
| Pharmacodynamics | Study of pharmacological actions on living systems, including the reactions with and binding to cell constituents, and the biochemical and physiological consequences of these actions [43]. |
| Pharmacokinetics | The activity of drugs in the body over a period of time, including the processes by which drugs are absorbed, distributed in the body, localized in the tissues, metabolized and excreted [40]. |
| Post-translational modifications | The enzyme-mediated covalent addition of functional groups to proteins during or after synthesis that increase the functional diversity of the protein [44,45]. Can include the oxidation of methionine residues. The methionine residues in FSH are not directly located in regions that are critical for binding to the FSHR: methionine 29 is involved in α-β subunit heterodimerization; methionine 47 is located close to the FSHR binding site, but it is not directly involved in the ligand–receptor interaction; methionine 71 is located close to the heterodimerization site but is not directly involved in heterodimerization; and methionine 109 is located in the non-structural C-terminal region. However, the oxidation of these residues may lead to conformational changes, with the potential for indirect effects on biological activity, PK or protein aggregation, with the alteration of the immunogenicity of therapeutic proteins [23]. |
| Quality control | The process used to ensure that all preliminary testing is carried out and that the quality of the final product has been deemed to meet all standards of identity, strength, quality and purity [7,38]. |
| Quantity | The physicochemical measurement of protein content. If physicochemical tests alone are used to quantitate the biological activity (based on the correlation between the expected clinical response and the activity in the biological assay, as established in pharmacodynamic or clinical studies), potency should be expressed in mass [7]. |
| Reference House Standard (RHS) | Biotech material, appropriately characterized, produced from validated production processes and used as reference standard. It is characterized and calibrated versus a suitable IS using an appropriate characterization panel and bioassay. The RHS is intended for use in specified analytical tests, in which its properties are compared with properties of samples under examination. |
| Relative potency | Measured by comparing the concentration–response curves of a manufactured test batch with that of a RHS. |
| Sialylation | The covalent addition of sialic acid to the terminal end of oligosaccharides attached to the protein core. Terminal sialylation, namely the presence or absence of a sialic acid (a2,3 and/or a2,6 N-Acetyl neuraminic acid (Neu5Ac)) that caps glycan branches, is the major factor governing the in vivo clearance rate. More acidic glycoforms have longer elimination rates due to reduced renal clearance, while less acidic glycoforms have a higher affinity for the FSHR [2]. |
| Specific activity (units of biological activity per mg of product) | The quantitative measure of biological activity evaluated as biological activity/mass and can also serve to measure the purity and consistency of a product. |
References
- Casarini, L.; Crepieux, P. Molecular Mechanisms of Action of FSH. Front. Endocrinol. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.A.; Ulloa-Aguirre, A. New Human Follitropin Preparations: How Glycan Structural Differences May Affect Biochemical and Biological Function and Clinical Effect. Front. Endocrinol. 2021, 12, 636038. [Google Scholar] [CrossRef] [PubMed]
- Bergandi, L.; Canosa, S.; Carosso, A.R.; Paschero, C.; Gennarelli, G.; Silvagno, F.; Benedetto, C.; Revelli, A. Human Recombinant FSH and Its Biosimilars: Clinical Efficacy, Safety, and Cost-Effectiveness in Controlled Ovarian Stimulation for In Vitro Fertilization. Pharmaceuticals 2020, 13, 136. [Google Scholar] [CrossRef] [PubMed]
- Lunenfeld, B.; Bilger, W.; Longobardi, S.; Alam, V.; D’Hooghe, T.; Sunkara, S.K. The Development of Gonadotropins for Clinical Use in the Treatment of Infertility. Front. Endocrinol. 2019, 10, 429. [Google Scholar] [CrossRef]
- Ben-Rafael, Z.; Levy, T.; Schoemaker, J. Pharmacokinetics of Follicle-Stimulating Hormone: Clinical Significance. Fertil. Steril. 1998, 69, 40S–49S. [Google Scholar] [CrossRef]
- Bousfield, G.R.; May, J.V.; Davis, J.S.; Dias, J.A.; Kumar, T.R. In Vivo and In Vitro Impact of Carbohydrate Variation on Human Follicle-Stimulating Hormone Function. Front. Endocrinol. 2018, 9, 216. [Google Scholar] [CrossRef]
- EMA; ICH. Topic Q 6 B Specifications: Test Procedures and Acceptance Criteria for Biotechnological/Biological Products. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-q-6-b-test-procedures-acceptance-criteria-biotechnological/biological-products-step-5_en.pdf (accessed on 4 April 2023).
- Barrios-De-Tomasi, J.; Timossi, C.; Merchant, H.; Quintanar, A.; Avalos, J.M.; Andersen, C.Y.; Ulloa-Aguirre, A. Assessment of the in vitro and in vivo biological activities of the human follicle-stimulating isohormones. Mol. Cell. Endocrinol. 2002, 186, 189–198. [Google Scholar] [CrossRef]
- EMA. Guideline on Potency Testing of Cell Based Immunotherapy Medicinal Products for the Treatment of Cancer. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-potency-testing-cell-based-immunotherapy-medicinal-products-treatment-cancer_en.pdf (accessed on 4 April 2023).
- Manzi, L.; Sepe, N.; Migliaccio, W.; Lanzoni, L.; Iozzino, L.; D’Angelo, F.; Colarusso, L.; Montenegro, S.; Palmese, A.; D’Hooghe, T.; et al. Comparative Assessment of the Structural Features of Originator Recombinant Human Follitropin Alfa Versus Recombinant Human Follitropin Alfa Biosimilar Preparations Approved in Non-European Regions. Int. J. Mol. Sci. 2022, 23, 6762. [Google Scholar] [CrossRef]
- Bousfield, G.R.; Harvey, D.J. Follicle-Stimulating Hormone Glycobiology. Endocrinology 2019, 160, 1515–1535. [Google Scholar] [CrossRef]
- Meher, B.R.; Dixit, A.; Bousfield, G.R.; Lushington, G.H. Glycosylation Effects on FSH-FSHR Interaction Dynamics: A Case Study of Different FSH Glycoforms by Molecular Dynamics Simulations. PLoS ONE 2015, 10, e0137897. [Google Scholar] [CrossRef]
- Leao Rde, B.; Esteves, S.C. Gonadotropin therapy in assisted reproduction: An evolutionary perspective from biologics to biotech. Clinics 2014, 69, 279–293. [Google Scholar] [CrossRef]
- Lugan, I.; Giartosio, C.; Bassett, R.; Howles, C.; Munafo, A. The impact of more acidic or more basic Follicle Stimulating Hormone (FSH) isoforms on ovarian follicle development in down-regulated healthy women. Hum. Reprod. 2007, 22, I137. [Google Scholar]
- European Directorate for the Quality of Medicines & HealthCare European Pharmacopoeia. Available online: https://www.edqm.eu/en/european-pharmacopoeia-ph-eur-10th-edition-(June) (accessed on 4 April 2023).
- Steelman, S.L.; Pohley, F.M. Assay of the follicle stimulating hormone based on the augmentation with human chorionic gonadotropin. Endocrinology 1953, 53, 604–616. [Google Scholar] [CrossRef]
- Mastrangeli, R.; Satwekar, A.; Cutillo, F.; Ciampolillo, C.; Palinsky, W.; Longobardi, S. In-vivo biological activity and glycosylation analysis of a biosimilar recombinant human follicle-stimulating hormone product (Bemfola) compared with its reference medicinal product (GONAL-f). PLoS ONE 2017, 12, e0184139. [Google Scholar] [CrossRef]
- USP. United States Pharmacopeia Convention (USP 28-NF); United States Pharmacopeia: Rockville, MD, USA, 2005; pp. 2274–2286. [Google Scholar]
- Driebergen, R.; Baer, G. Quantification of follicle stimulating hormone (follitropin alfa): Is in vivo bioassay still relevant in the recombinant age? Curr. Med. Res. Opin. 2003, 19, 41–46. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959; p. 238. [Google Scholar]
- Wang, L.Y.; Liang, C.G.; Yang, H.M.; Li, J.; Li, Z.J.; Zhang, H.; Lv, P.; Li, Y.; Wang, J.Z. Development of a robust reporter gene-based assay for the bioactivity determination of recombinant human follicle stimulating hormone (rhFSH) pharmaceutical products. J. Pharm. Biomed. Anal. 2020, 177, 112855. [Google Scholar] [CrossRef]
- Thomas, R.M.; Nechamen, C.A.; Mazurkiewicz, J.E.; Ulloa-Aguirre, A.; Dias, J.A. The adapter protein APPL1 links FSH receptor to inositol 1,4,5-trisphosphate production and is implicated in intracellular Ca(2+) mobilization. Endocrinology 2011, 152, 1691–1701. [Google Scholar] [CrossRef]
- Hermeling, S.; Crommelin, D.J.; Schellekens, H.; Jiskoot, W. Structure-immunogenicity relationships of therapeutic proteins. Pharm. Res. 2004, 21, 897–903. [Google Scholar] [CrossRef]
- Timossi, C.M.; Barrios-de-Tomasi, J.; Gonzalez-Suarez, R.; Arranz, M.C.; Padmanabhan, V.; Conn, P.M.; Ulloa-Aguirre, A. Differential effects of the charge variants of human follicle-stimulating hormone. J. Endocrinol. 2000, 165, 193–205. [Google Scholar] [CrossRef]
- Simoni, M.; Gromoll, J.; Nieschlag, E. The follicle-stimulating hormone receptor: Biochemistry, molecular biology, physiology, and pathophysiology. Endocr. Rev. 1997, 18, 739–773. [Google Scholar]
- D’Antonio, M.; Borrelli, F.; Datola, A.; Bucci, R.; Mascia, M.; Polletta, P.; Piscitelli, D.; Papoian, R. Biological characterization of recombinant human follicle stimulating hormone isoforms. Hum. Reprod. 1999, 14, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Andreone, L.; Ambao, V.; Urrutia, M.; Calandra, R.S.; Rulli, S.B. Hormonal Regulation of Follicle-Stimulating Hormone Glycosylation in Males. Front. Endocrinol. 2019, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.P.; Gaines Das, R.E.; Balen, A.H. Definition and measurement of follicle stimulating hormone. Endocr. Rev. 2000, 21, 5–22. [Google Scholar] [CrossRef] [PubMed]
- de Leeuw, R.; Mulders, J.; Voortman, G.; Rombout, F.; Damm, J.; Kloosterboer, L. Structure-function relationship of recombinant follicle stimulating hormone (Puregon). Mol. Hum. Reprod. 1996, 2, 361–369. [Google Scholar] [CrossRef]
- EMA. ICH Guideline Q10 on Pharmaceutical Quality System; EMA: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Hugues, J.N.; Barlow, D.H.; Rosenwaks, Z.; Cédrin-Durnerin, I.; Robson, S.; Pidoux, L.; Loumaye, E. Improvement in consistency of response to ovarian stimulation with recombinant human follicle stimulating hormone resulting from a new method for calibrating the therapeutic preparation. Reprod. Biomed. Online 2003, 6, 185–190. [Google Scholar] [CrossRef]
- USP. United States Pharmacopeia—Design and Development of Biological Assays (USP 35-NF 30), 1032. Available online: https://www.drugfuture.com/Pharmacopoeia/usp35/PDF/5160-5174%20%5B1032%5D%20Development%20and%20Design%20of%20Bioassays.pdf (accessed on 4 April 2023).
- USP. United States Pharmacopeia—Biological Assay Validation (USP 35-NF 30); United States Pharmacopeia: Rockville, MD, USA, 2023; p. 1033. Available online: https://doi.usp.org/USPNF/USPNF_M912_01_01.html (accessed on 4 April 2023).
- Agrawal, G.; Dighe, R.R. Critical involvement of the hinge region of the follicle-stimulating hormone receptor in the activation of the receptor. J. Biol. Chem. 2009, 284, 2636–2647. [Google Scholar] [CrossRef]
- Casarini, L.; Lazzaretti, C.; Paradiso, E.; Limoncella, S.; Riccetti, L.; Sperduti, S.; Melli, B.; Marcozzi, S.; Anzivino, C.; Sayers, N.S.; et al. Membrane Estrogen Receptor (GPER) and Follicle-Stimulating Hormone Receptor (FSHR) Heteromeric Complexes Promote Human Ovarian Follicle Survival. iScience 2020, 23, 101812. [Google Scholar] [CrossRef]
- ICH. Q5D Derivation and Characterisation of Cell Substrates Used for Production of Biotechnological/Biological Products. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-q-5-d-derivation-characterisation-cell-substrates-used-production-biotechnological/biological-products-step-5_en.pdf (accessed on 4 April 2023).
- Eichler, J. Protein glycosylation. Curr. Biol. 2019, 29, R229–R231. [Google Scholar] [CrossRef]
- EMA. Good Manufacturing Practice Guidelines. Volume 4. Available online: https://health.ec.europa.eu/medicinal-products/eudralex/eudralex-volume-4_en (accessed on 7 March 2023).
- MHRA. WHO International Standard Follicle Stimulating Hormone (FSH), Human, Recombinant, For Immunoassay. Available online: https://www.nibsc.org/documents/ifu/92-510.pdf (accessed on 4 April 2023).
- NCI/NIH. Definition of Mechanism of Action-NCI Dictionary of Cancer Terms. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/pharmacokinetics (accessed on 4 April 2023).
- MHRA. WHO: International Standards. Available online: https://www.nibsc.org/standardisation/international_standards.aspx (accessed on 4 April 2023).
- WHO. WHO Expert Committee on Biological Standardisation, 26th Report; WHO Technical Report Series No. 565; WHO: Geneva, Switzerland, 1975; p. 17. [Google Scholar]
- IUPAC. Compendium of Chemical Terminology. In The “Gold Book”, 2nd ed.; McNaught, A.D., Wilkinson, A., Eds.; Blackwell Science: Oxford, UK; Malden, MA, USA, 1997; p. vii, 450p. [Google Scholar]
- Becares, N.; Gage, M.C.; Pineda-Torra, I. Posttranslational Modifications of Lipid-Activated Nuclear Receptors: Focus on Metabolism. Endocrinology 2017, 158, 213–225. [Google Scholar] [CrossRef]
- Chen, L.; Kashina, A. Post-translational Modifications of the Protein Termini. Front. Cell Dev. Biol. 2021, 9, 719590. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Sample | In Vitro Activity (IU/mg) * | Δ% In Vitro | In Vivo Activity (IU/mg) † | Δ% In Vivo | Z-Number |
|---|---|---|---|---|---|---|
| Acid enriched | FSE2003 untreated | 11,858 | 14,316 | 226 | ||
| FSE2003 acid enriched | 7153 | −40% | 20,785 | +45% | 294 | |
| FSE2004 untreated | 12,688 | 15,139 | 221 | |||
| FSE2004 acid enriched | 6442 | −49% | 23,580 | +56% | 289 | |
| FSE2005 untreated | 11,804 | 14,038 | 221 | |||
| FSE2005 acid enriched | 8011 | −32% | 27,414 | +95% | 289 | |
| Basic enriched | FSE2003 untreated | 11,858 | 14,316 | 226 | ||
| FSE2003 basic enriched | 14,219 | +20% | 7467 | −48% | 167 | |
| FSE2004 untreated | 12,688 | 15,139 | 221 | |||
| FSE2004 basic enriched | 12,230 | −4% | 7912 | −48% | 169 | |
| FSE2005 untreated | 11,804 | 14,038 | 221 | |||
| FSE2005 basic enriched | 13,744 | +16% | 4434 | −68% | 165 | |
| Oxidized | FSE2003 untreated | 11,858 | 14,316 | ·· | ||
| FSE2003 oxidized | 2963 | −75% | 12,717 | −11% | ·· | |
| FSE2004 untreated | 12,688 | 15,139 | ·· | |||
| FSE2004 oxidized | 2222 | −82% | 12,764 | −16% | ·· | |
| FSE2005 untreated | 11,804 | 14,038 | ·· | |||
| FSE2005 oxidized | 3172 | −73% | 12,771 | −9% | ·· | |
| Acid pH | FSE2003 untreated | 11,858 | 14,316 | ·· | ||
| FSE2003 acid pH | 2444 | −79% | 5383 | −62% | ·· | |
| FSE2004 untreated | 12,688 | 15,139 | ·· | |||
| FSE2004 acid pH | 2168 | −83% | 4573 | −70% | ·· | |
| FSE2005 untreated | 11,804 | 14,038 | ·· | |||
| FSE2005 acid pH | 2165 | −82% | 4576 | −67% | ·· | |
| Total desialylation | FSE2003 untreated | 11,858 | 14,316 | 226 | ||
| FSE2003 desialyl | 23,241 | +96% | 849 | −94% | 6 | |
| FSE2004 untreated | 12,688 | 15,139 | 221 | |||
| FSE2004 desialyl | 25,058 | +97% | 843 | −94% | 7 | |
| FSE2005 untreated | 11,804 | 14,038 | 221 | |||
| FSE2005 desialyl | 25,296 | +114% | 1900 | −86% | 7 | |
| FSE2012 untreated | 12,177 | 15,689 | ||||
| FSE2012 desialyl T1 | 23,193 | +90% | 3932 | −81% | 13 | |
| FSE2012 desialyl T2 | 23,571 | +94% | 2960 | −85% | 12 | |
| FSE2012 desialyl T3 | 25,545 | +110% | 3735 | −82% | 11 | |
| FSE2012 desialyl T4 | 26,207 | +115% | 2915 | −86% | 7 | |
| Different sialylation levels | FSE2004 untreated | 12,688 | 15,139 | 227 | ||
| Desialylation level 1 (50%) | 17,723 | +40% | 6617 | −56% | 130 | |
| Desialylation level 2 (75%) | 21,963 | +73% | 2819 | −81% | 68 | |
| Desialylation level 3 (100%) | 25,530 | +101% | 1925 | −87% | 9 | |
| FSE2004 desialylated | 25,058 | +97% | 843 | −94% | 7 | |
| Desialylated and degalactosylated | FSE2003 untreated | 11,858 | 14,316 | 226 | ||
| FSE2003 Desial/Degal | 17,709 | +49% | 1324 | −91% | 6 | |
| FSE2004 untreated | 12,688 | 15,139 | 227 | |||
| FSE2004 Desial/Degal | 17,317 | +36% | 743 | −95% | 6 | |
| FSE2005 untreated | 11,804 | 14,038 | 224 | |||
| FSE2005 Desial/Degal | 19,379 | +64% | 2788 | -80% | 7 |
| Category | Procedure |
|---|---|
| Acidic variant | Acid fractionation performed by manufacturing site. |
| Basic variant | Basic fractionation performed by CRO. |
| Oxidation | Incubation at room temperature with 0.5% H2O2 for 60 min. |
| Acidic pH | 1:1 dilution with sodium citrate 0.5 M pH 3.0 and incubation with sialidase at 25 °C for 3 days. |
| Glycosylation (desialylated sample) | Incubation at 37 °C for 18 h with Sialidase A. The enzymatic activity was stopped by freezing at −20 ± 5 °C. |
| Glycosylation (desialylated + degalactosylated sample) | Incubation at +37 °C for 18 h with Sialidase A, followed by galactosidase. The enzymatic activity was stopped by freezing at −20 ± 5 °C. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nevelli, F.; Palmese, A.; Gleixner, R.; Peroglio, F.; D’Acunto, C.-W.; Dadone, A.; D’Hooghe, T.; Lispi, M. Biological Assay to Determine Gonadotropin Potency: From In Vivo to In Vitro Sustainable Method. Int. J. Mol. Sci. 2023, 24, 8040. https://doi.org/10.3390/ijms24098040
Nevelli F, Palmese A, Gleixner R, Peroglio F, D’Acunto C-W, Dadone A, D’Hooghe T, Lispi M. Biological Assay to Determine Gonadotropin Potency: From In Vivo to In Vitro Sustainable Method. International Journal of Molecular Sciences. 2023; 24(9):8040. https://doi.org/10.3390/ijms24098040
Chicago/Turabian StyleNevelli, Francesco, Angelo Palmese, Ralf Gleixner, Flavio Peroglio, Cosimo-Walter D’Acunto, Aurora Dadone, Thomas D’Hooghe, and Monica Lispi. 2023. "Biological Assay to Determine Gonadotropin Potency: From In Vivo to In Vitro Sustainable Method" International Journal of Molecular Sciences 24, no. 9: 8040. https://doi.org/10.3390/ijms24098040
APA StyleNevelli, F., Palmese, A., Gleixner, R., Peroglio, F., D’Acunto, C.-W., Dadone, A., D’Hooghe, T., & Lispi, M. (2023). Biological Assay to Determine Gonadotropin Potency: From In Vivo to In Vitro Sustainable Method. International Journal of Molecular Sciences, 24(9), 8040. https://doi.org/10.3390/ijms24098040