
Efficacy of Immunization against a Novel Synthetic 13-Amino Acid Betaglycan-Binding Peptide Sequence of Inhibin α Subunit on Promoting Fertility in Female Rats
 , ,
, , 
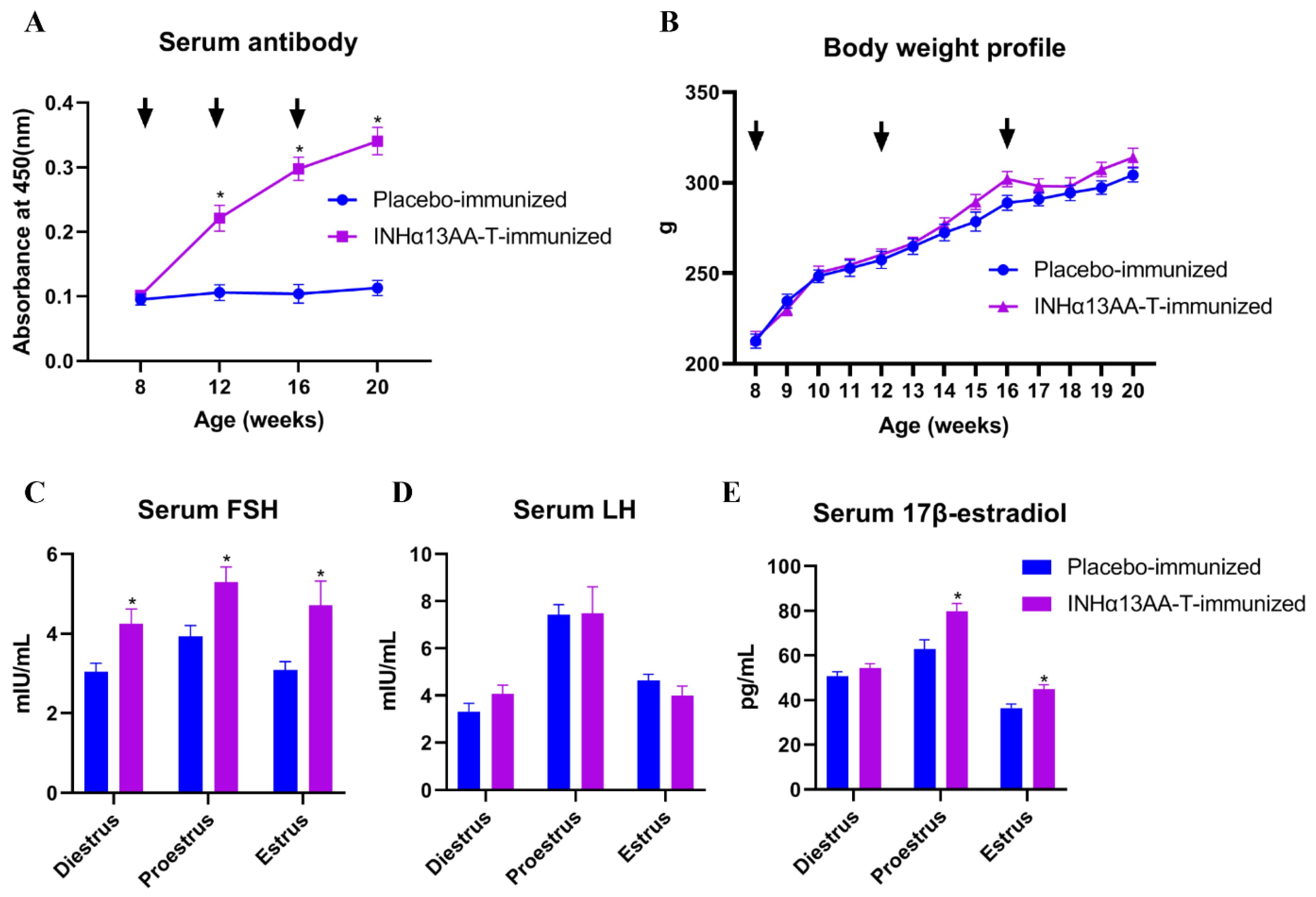{kind=link}
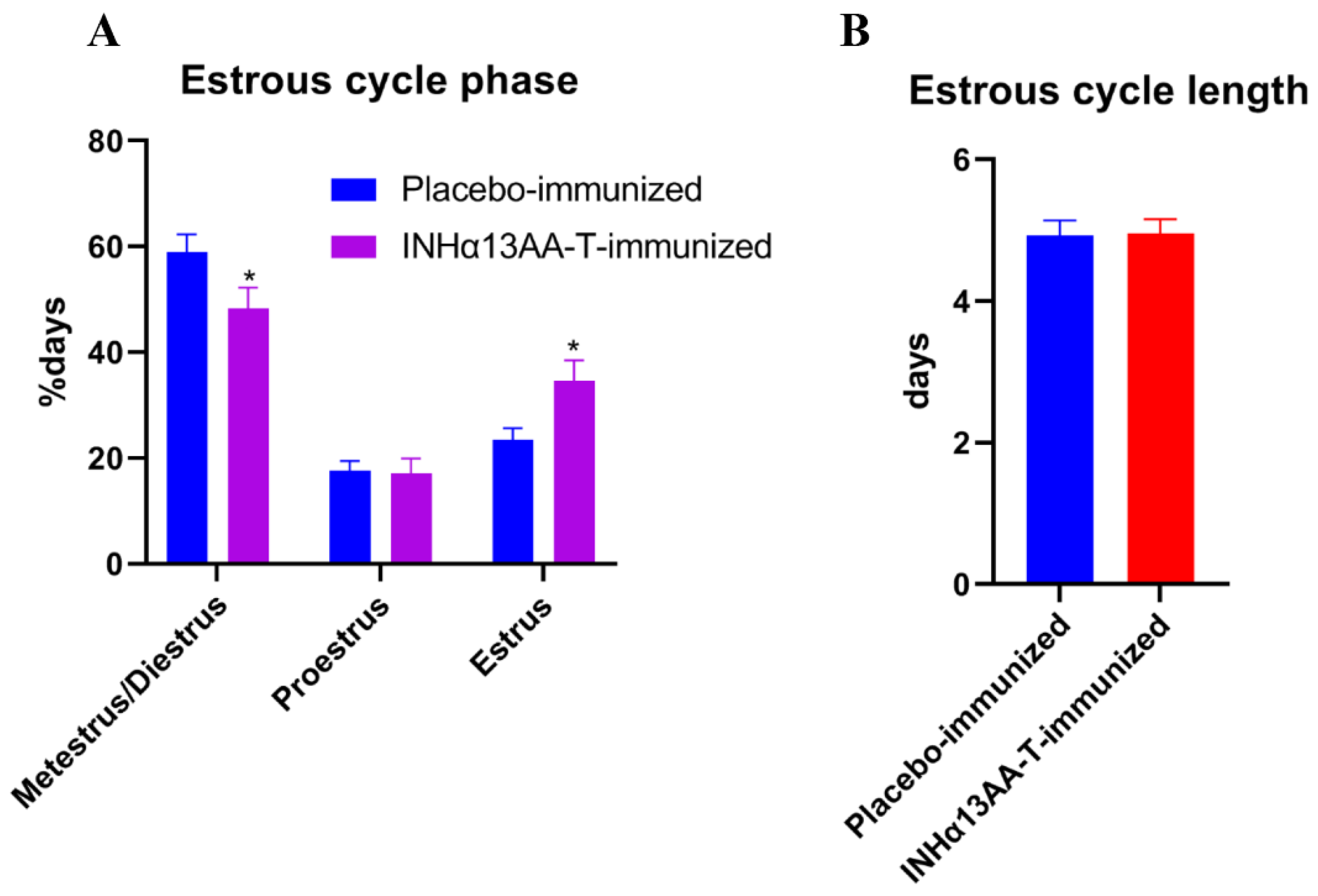{kind=link}
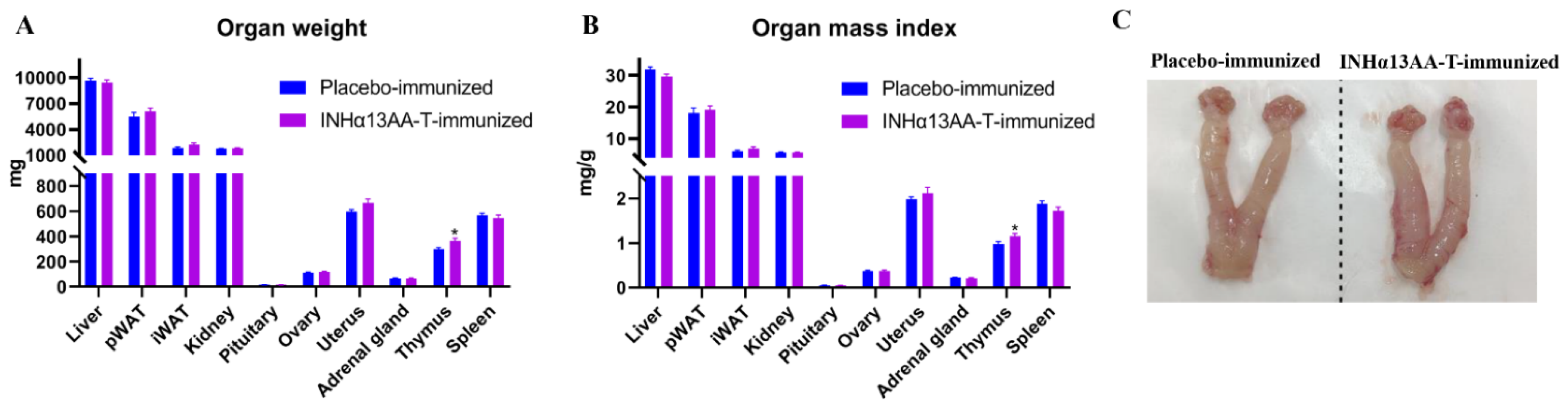{kind=link}
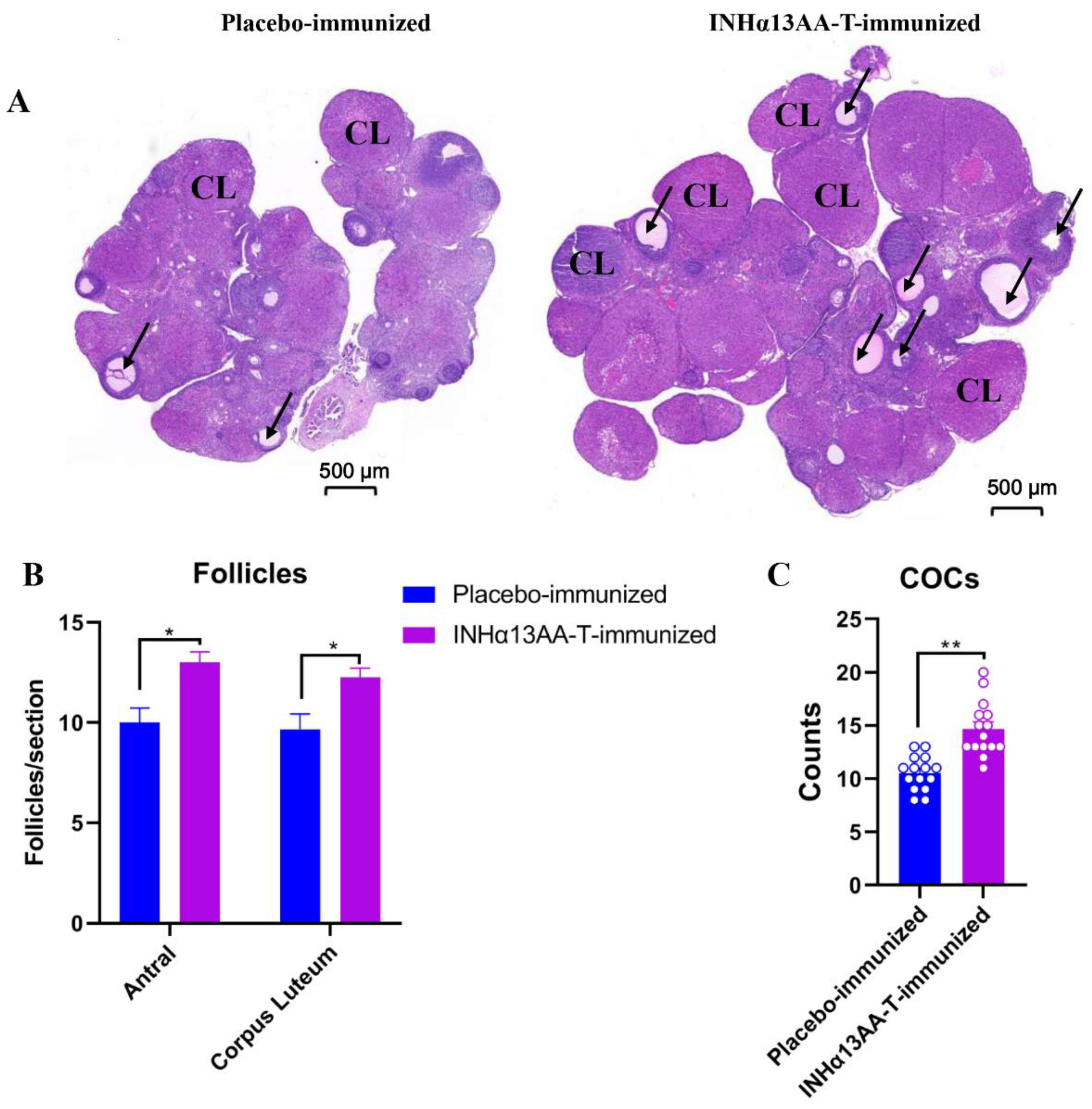{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. INHα13AA-T Immunization Triggered Good Immunological and Biological Responses in Rats
2.2. INHα13AA-T Immunization Prolonged Estrous Cycle Phases
2.3. INHα13AA-T Immunization Increased the Thymus Weight and Index but Without Effects on Other Organs in Female Rats
2.4. INHα13AA-T Immunization Promoted Ovarian Folliculogenesis and Natural Ovulation in Female Rats
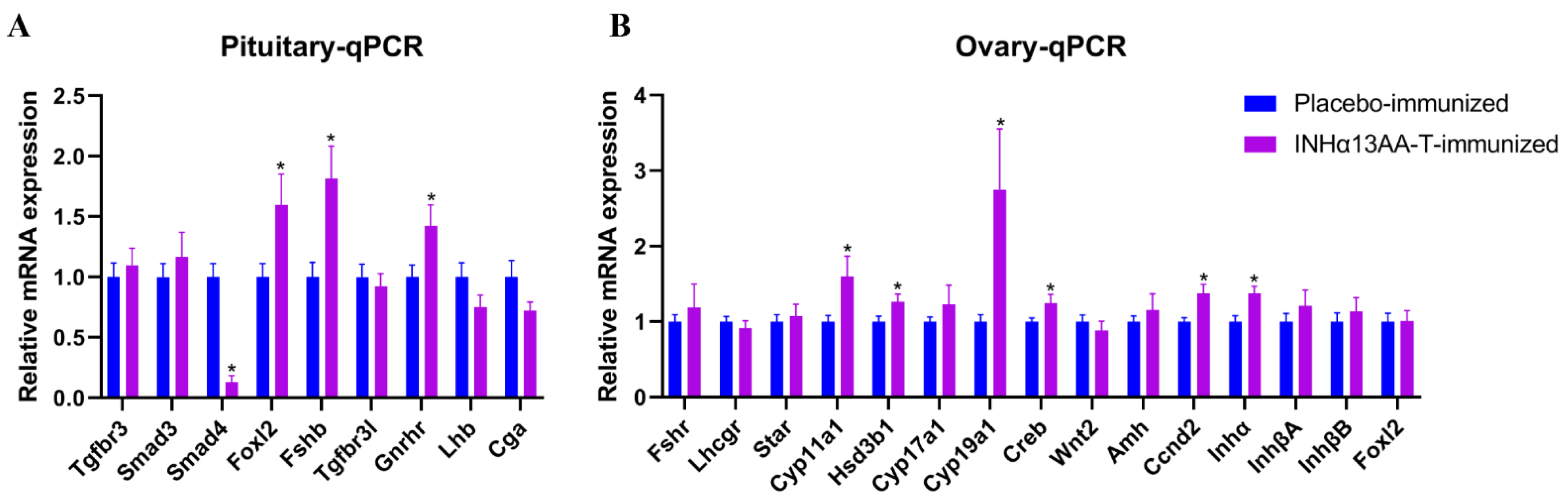
2.5. INHα13AA-T Immunization Promoted Pituitary Fshb and Ovarian Steroidogenesis- and Folliculogenesis-Associated Gene Expression
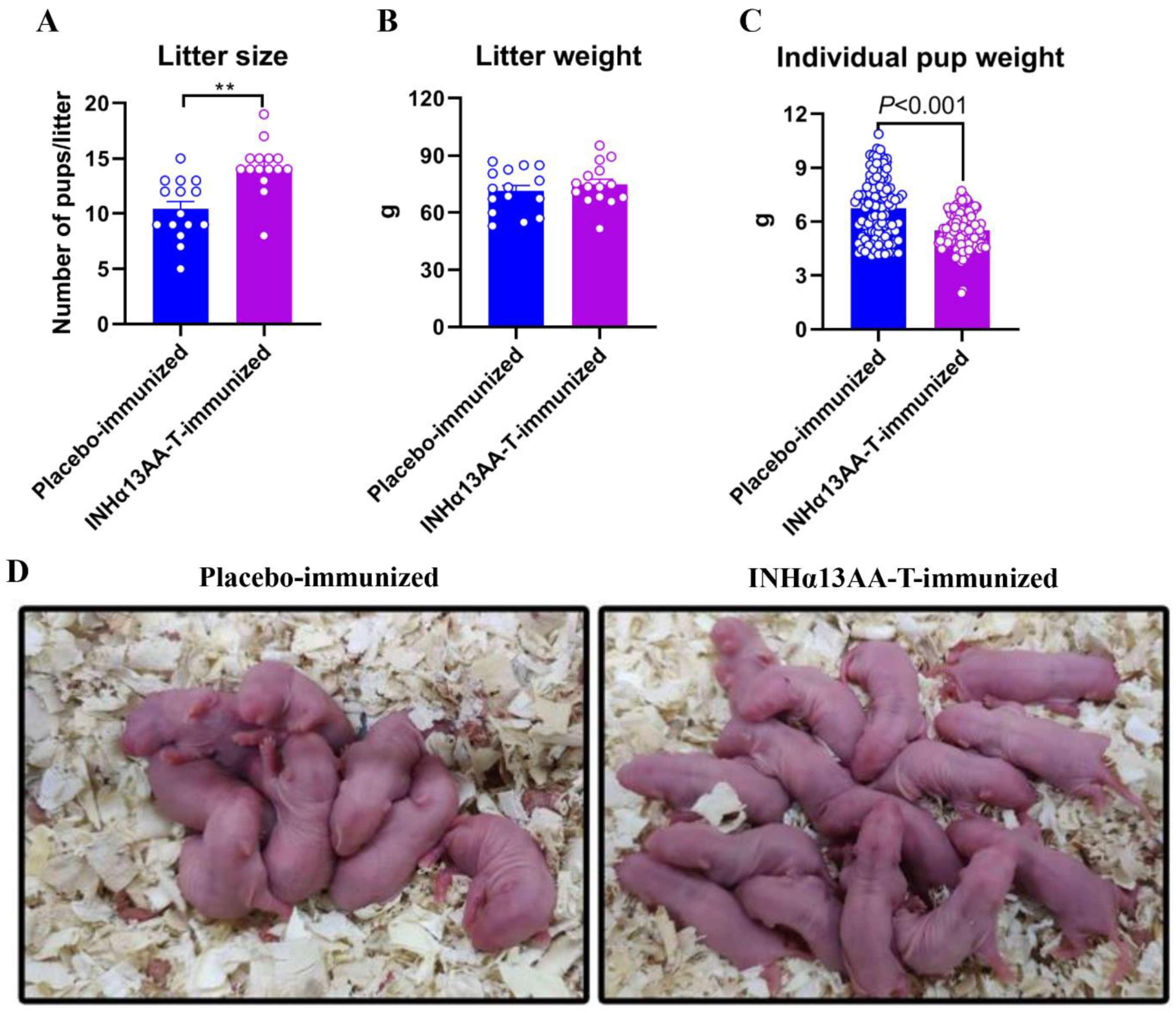
2.6. INHα13AA-T Immunization Improved the Fertility of Female Rats
3. Discussion
4. Materials and Methods
4.1. Homology Conservation Analysis of Betaglycan-Binding Epitope on Inhibin α Subunit
4.2. Antigen Molecular Design, Synthesis and Inhibin Vaccine Formulation
4.3. Animals and Experimental Design
4.4. INHα13AA-T Immunization Improved the Fertility of Female Rats
4.4.1. Experiment 1: Effects of INHα13AA-T Immunization on the Reproductive Physiology and Natural Ovulation of Female Rats
4.4.2. Experiment 2: Effects of INHα13AA-T Immunization on Fertility of Female Rats
4.5. Antibody Titer Assays
4.6. Serum Hormone Assays
4.7. Ovary Histology and Follicle Counts
4.8. Relative Real-Time PCR (RT-PCR)
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vale, W.; Wiater, E.; Gray, P.; Harrison, C.; Bilezikjian, L.; Choe, S. Activins and inhibins and their signaling. Ann. N. Y. Acad. Sci. 2004, 1038, 142–147. [Google Scholar] [CrossRef]
- Woodruff, T.K.; Besecke, L.M.; Groome, N.; Draper, L.B.; Schwartz, N.B.; Weiss, J. Inhibin A and inhibin B are inversely correlated to follicle-stimulating hormone, yet are discordant during the follicular phase of the rat estrous cycle, and inhibin A is expressed in a sexually dimorphic manner. Endocrinology 1996, 137, 5463–5467. [Google Scholar] [CrossRef]
- Li, Y.; Fortin, J.; Ongaro, L.; Zhou, X.; Boehm, U.; Schneyer, A.; Bernard, D.J.; Lin, H.Y. Betaglycan (TGFBR3) Functions as an Inhibin A, but Not Inhibin B, Coreceptor in Pituitary Gonadotrope Cells in Mice. Endocrinology 2018, 159, 4077–4091. [Google Scholar] [CrossRef]
- Liu, Q.; Han, L.; Rehman, Z.U.; Dan, X.; Liu, X.; Bhattarai, D.; Yang, L. The efficacy of an inhibin DNA vaccine delivered by attenuated Salmonella choleraesuis on follicular development and ovulation responses in crossbred buffaloes. Anim. Reprod. Sci. 2016, 172, 76–82. [Google Scholar] [CrossRef]
- Han, L.; Mao, D.G.; Zhang, D.K.; Liang, A.X.; Fang, M.; Moaeen-ud-Din, M.; Yang, L.G. Development and evaluation of a novel DNA vaccine expressing inhibin alpha (1-32) fragment for improving the fertility in rats and sheep. Anim. Reprod. Sci. 2008, 109, 251–265. [Google Scholar] [CrossRef]
- Shi, F.; Ozawa, M.; Komura, H.; Watanabe, G.; Tsonis, C.G.; Suzuki, A.K.; Taya, K. Induction of superovulation by inhibin vaccine in cyclic guinea-pigs. J. Reprod. Fertil. 2000, 118, 1–7. [Google Scholar] [CrossRef]
- Brûlé, E.; Wang, Y.; Li, Y.; Lin, Y.F.; Zhou, X.; Ongaro, L.; Alonso, C.A.I.; Buddle, E.R.S.; Schneyer, A.L.; Byeon, C.H.; et al. TGFBR3L is an inhibin B co-receptor that regulates female fertility. Sci. Adv. 2021, 7, eabl4391. [Google Scholar] [CrossRef]
- Lewis, K.A.; Gray, P.C.; Blount, A.L.; MacConell, L.A.; Wiater, E.; Bilezikjian, L.M.; Vale, W. Betaglycan binds inhibin and can mediate functional antagonism of activin signalling. Nature 2000, 404, 411–414. [Google Scholar] [CrossRef]
- Makanji, Y.; Walton, K.L.; Wilce, M.C.; Chan, K.L.; Robertson, D.M.; Harrison, C.A. Suppression of inhibin A biological activity by alterations in the binding site for betaglycan. J. Biol. Chem. 2008, 283, 16743–16751. [Google Scholar] [CrossRef]
- Yan, L.; Li, H.; Shi, Z. Immunization against inhibin improves in vivo and in vitro embryo production. Anim. Reprod. Sci. 2015, 163, 1–9. [Google Scholar] [CrossRef]
- Wrathall, J.H.; McLeod, B.J.; Glencross, R.G.; Beard, A.J.; Knight, P.G. Inhibin immunoneutralization by antibodies raised against synthetic peptide sequences of inhibin alpha subunit: Effects on gonadotrophin concentrations and ovulation rate in sheep. J. Endocrinol. 1990, 124, 167–176. [Google Scholar] [CrossRef]
- King, B.F.; Britt, J.H.; Esbenshade, K.L.; Flowers, W.L.; Sesti, L.A.; Martin, T.L.; Ireland, J.J. Ovulatory and endocrine responses after active immunization of gilts against a synthetic fragment of bovine inhibin. J. Anim. Sci. 1993, 71, 975–982. [Google Scholar] [CrossRef][Green Version]
- Meyer, R.L.; Carlson, K.M.; Rivier, J.; Wheaton, J.E. Antiserum to an inhibin alpha-chain peptide neutralizes inhibin bioactivity and increases ovulation rate in sheep. J. Anim. Sci. 1991, 69, 747–754. [Google Scholar] [CrossRef][Green Version]
- Sasaki, K.; Medan, M.S.; Watanabe, G.; Sharawy, S.; Taya, K. Immunization of goats against inhibin increased follicular development and ovulation rate. J. Reprod. Dev. 2006, 52, 543–550. [Google Scholar] [CrossRef]
- Medan, M.S.; Watanabe, G.; Sasaki, K.; Nagura, Y.; Sakaime, H.; Fujita, M.; Sharawy, S.; Taya, K. Ovarian and hormonal response of female goats to active immunization against inhibin. J. Endocrinol. 2003, 177, 287–294. [Google Scholar] [CrossRef][Green Version]
- Schanbacher, B.D.; Schemm, S.R.; Rhind, S.M. Gonadotrophin concentrations and ovulation rates in Suffolk ewes actively or passively immunized against inhibin alpha. J. Reprod. Fertil. 1991, 93, 133–139. [Google Scholar] [CrossRef][Green Version]
- O’Shea, T.; Andrews, C.M.; Bindon, B.M.; Hillard, M.A.; Miyamoto, K.; Sinosich, M.J. Immunization of merino ewes with a synthetic inhibin peptide or with preparations obtained from bovine and porcine follicular fluids by immunoaffinity chromatography result in different effects on ovulation rate and on plasma gonadotrophin concentrations. Reprod. Fertil. 1991, 3, 659–670. [Google Scholar] [CrossRef]
- Tannetta, D.S.; Feist, S.A.; Bleach, E.C.; Groome, N.P.; Evans, L.W.; Knight, P.G. Effects of active immunization of sheep against an amino terminal peptide of the inhibin alpha C subunit on intrafollicular levels of activin A, inhibin A and follistatin. J. Endocrinol. 1998, 157, 157–168. [Google Scholar] [CrossRef][Green Version]
- Medan, M.S.; Takedom, T.; Aoyagi, Y.; Konishi, M.; Yazawa, S.; Watanabe, G.; Taya, K. The effect of active immunization against inhibin on gonadotropin secretions and follicular dynamics during the estrous cycle in cows. J. Reprod. 2006, 52, 107–113. [Google Scholar] [CrossRef][Green Version]
- Wrathall, J.H.; McLeod, B.J.; Glencross, R.G.; Knight, P.G. Effects of active immunization against a synthetic peptide sequence of the inhibin alpha-subunit on plasma gonadotrophin concentrations, ovulation rate and lambing rate in ewes. J. Reprod. Fertil. 1992, 95, 175–182. [Google Scholar] [CrossRef][Green Version]
- Glencross, R.G.; Bleach, E.C.; McLeod, B.J.; Beard, A.J.; Knight, P.G. Effect of active immunization of heifers against inhibin on plasma FSH concentrations, ovarian follicular development and ovulation rate. J. Endocrinol. 1992, 134, 11–18. [Google Scholar] [CrossRef]
- Glencross, R.G.; Bleach, E.C.; Wood, S.C.; Knight, P.G. Active immunization of heifers against inhibin: Effects on plasma concentrations of gonadotrophins, steroids and ovarian follicular dynamics during prostaglandin-synchronized cycles. J. Reprod. Fertil. 1994, 100, 599–605. [Google Scholar] [CrossRef]
- Walton, K.L.; Goney, M.P.; Peppas, Z.; Stringer, J.M.; Winship, A.; Hutt, K.; Goodchild, G.; Maskey, S.; Chan, K.L.; Brûlé, E.; et al. Inhibin Inactivation in Female Mice Leads to Elevated FSH Levels, Ovarian Overstimulation, and Pregnancy Loss. Endocrinology 2022, 163, bqac025. [Google Scholar] [CrossRef]
- Tran, S.; Zhou, X.; Lafleur, C.; Calderon, M.J.; Ellsworth, B.S.; Kimmins, S.; Boehm, U.; Treier, M.; Boerboom, D.; Bernard, D.J. Impaired fertility and FSH synthesis in gonadotrope-specific Foxl2 knockout mice. Mol. Endocrinol. 2013, 27, 407–421. [Google Scholar] [CrossRef]
- Corpuz, P.S.; Lindaman, L.L.; Mellon, P.L.; Djurdjica, C.J.M.E. FoxL2 Is required for activin induction of the mouse and human follicle-stimulating hormone beta-subunit genes. Mol. Endocrinol. 2010, 24, 1037–1051. [Google Scholar] [CrossRef]
- Herndon, M.K.; Nilson, J.H. Maximal expression of Foxl2 in pituitary gonadotropes requires ovarian hormones. PLoS ONE 2015, 10, e0126527. [Google Scholar] [CrossRef]
- Raju, G.A.; Chavan, R.; Deenadayal, M.; Gunasheela, D.; Gutgutia, R.; Haripriya, G.; Govindarajan, M.; Patel, N.H.; Patki, A.S. Luteinizing hormone and follicle stimulating hormone synergy: A review of role in controlled ovarian hyper-stimulation. J. Hum. Reprod. Sci. 2013, 6, 227–234. [Google Scholar] [CrossRef]
- Samardzija Nenadov, D.; Pogrmic-Majkic, K.; Fa, S.; Stanic, B.; Tubic, A.; Andric, N. Environmental mixture with estrogenic activity increases Hsd3b1 expression through estrogen receptors in immature rat granulosa cells. J. Appl. Toxicol. 2018, 38, 879–887. [Google Scholar] [CrossRef]
- Cayton Vaught, K.C.; Hazimeh, D.; Carter, A.S.; Devine, K.; Maher, J.Y.; Maguire, M.; McGee, E.A.; Driggers, P.H.; Segars, J.H. AKAP13 Enhances CREB1 Activation by FSH in Granulosa Cells. Reprod. Sci. 2023; ahead of print. [Google Scholar]
- Sicinski, P.; Donaher, J.L.; Geng, Y.; Parker, S.B.; Gardner, H.; Park, M.Y.; Robker, R.L.; Richards, J.S.; McGinnis, L.K.; Biggers, J.D.; et al. Cyclin D2 is an FSH-responsive gene involved in gonadal cell proliferation and oncogenesis. Nature 1996, 384, 470–474. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Q.; Wen, Z.; Yuan, K.; Su, Z.; Wang, Y.; Zhong, Y.; Ge, R.S. Androgen and Luteinizing Hormone Stimulate the Function of Rat Immature Leydig Cells Through Different Transcription Signals. Front. Endocrinol. 2021, 12, 599149. [Google Scholar] [CrossRef]
- Johnson, A.L.; Bridgham, J.T. Regulation of steroidogenic acute regulatory protein and luteinizing hormone receptor messenger ribonucleic acid in hen granulosa cells. Endocrinology 2001, 142, 3116–3124. [Google Scholar] [CrossRef]
- Donovan, C.E.; Hazzard, T.; Schmidt, A.; LeMieux, J.; Hathaway, F.; Kutzler, M.A. Effects of a commercial canine gonadotropin releasing hormone vaccine on estrus suppression and estrous behavior in mares. Anim. Reprod. Sci. 2013, 142, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Yao, H.; Li, J.; Zhuo, Y.; Yu, G.; Bu, G.; Cao, X.; Du, X.; Liang, Q.; Zeng, X.; et al. Effects of active immunization against a 13-amino acid receptor-binding epitope of FSHbeta on fertility regulation in female mice. Reprod. Biol. 2022, 22, 100669. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, A.F.; Akhigbe, R.E. Staging of the estrous cycle and induction of estrus in experimental rodents: An update. Fertil. Res. Pract. 2020, 6, 5. [Google Scholar] [CrossRef]
- Thapa, P.; Farber, D.L. The Role of the Thymus in the Immune Response. Thorac. Surg. Clin. 2019, 29, 123–131. [Google Scholar] [CrossRef]
- Han, X.; Guan, Z.; Xu, M.; Zhang, Y.; Yao, H.; Meng, F.; Zhuo, Y.; Yu, G.; Cao, X.; Du, X.; et al. A novel follicle-stimulating hormone vaccine for controlling fat accumulation. Theriogenology 2020, 148, 103–111. [Google Scholar] [CrossRef]
- Meloen, R.H.; Turkstra, J.A.; Lankhof, H.; Puijk, W.C.; Schaaper, W.M.; Dijkstra, G.; Wensing, C.J.; Oonk, R.B. Efficient immunocastration of male piglets by immunoneutralization of GnRH using a new GnRH-like peptide. Vaccine 1994, 12, 741–746. [Google Scholar] [CrossRef]
- Han, X.F.; Cao, X.H.; Tang, J.; Du, X.G.; Zeng, X.Y. Active immunization against GnRH reduces the synthesis of GnRH in male rats. Theriogenology 2013, 80, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Franca, M.M.; Han, X.; Funari, M.F.A.; Lerario, A.M.; Nishi, M.Y.; Fontenele, E.G.P.; Domenice, S.; Jorge, A.A.L.; Garcia-Galiano, D.; Elias, C.F.; et al. Exome Sequencing Reveals the POLR3H Gene as a Novel Cause of Primary Ovarian Insufficiency. J. Clin. Endocrinol. Metab. 2019, 104, 2827–2841. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Xia, X.; Chen, W.; Meng, F.; Cao, X.; Bu, G.; Gan, T.; Du, X.; Liang, Q.; Zeng, X. Efficacy of Immunization against a Novel Synthetic 13-Amino Acid Betaglycan-Binding Peptide Sequence of Inhibin α Subunit on Promoting Fertility in Female Rats. Int. J. Mol. Sci. 2023, 24, 6914. https://doi.org/10.3390/ijms24086914
Han X, Xia X, Chen W, Meng F, Cao X, Bu G, Gan T, Du X, Liang Q, Zeng X. Efficacy of Immunization against a Novel Synthetic 13-Amino Acid Betaglycan-Binding Peptide Sequence of Inhibin α Subunit on Promoting Fertility in Female Rats. International Journal of Molecular Sciences. 2023; 24(8):6914. https://doi.org/10.3390/ijms24086914
Chicago/Turabian StyleHan, Xingfa, Xue Xia, Weihao Chen, Fengyan Meng, Xiaohan Cao, Guixian Bu, Tian Gan, Xiaogang Du, Qiuxia Liang, and Xianyin Zeng. 2023. "Efficacy of Immunization against a Novel Synthetic 13-Amino Acid Betaglycan-Binding Peptide Sequence of Inhibin α Subunit on Promoting Fertility in Female Rats" International Journal of Molecular Sciences 24, no. 8: 6914. https://doi.org/10.3390/ijms24086914
APA StyleHan, X., Xia, X., Chen, W., Meng, F., Cao, X., Bu, G., Gan, T., Du, X., Liang, Q., & Zeng, X. (2023). Efficacy of Immunization against a Novel Synthetic 13-Amino Acid Betaglycan-Binding Peptide Sequence of Inhibin α Subunit on Promoting Fertility in Female Rats. International Journal of Molecular Sciences, 24(8), 6914. https://doi.org/10.3390/ijms24086914