
Multiplexing Biosensor for the Detection of Extracellular Vesicles as Biomarkers of Tissue Damage and Recovery after Ischemic Stroke

 ,
,  ,
,  ,
,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Stroke Patients Profiling
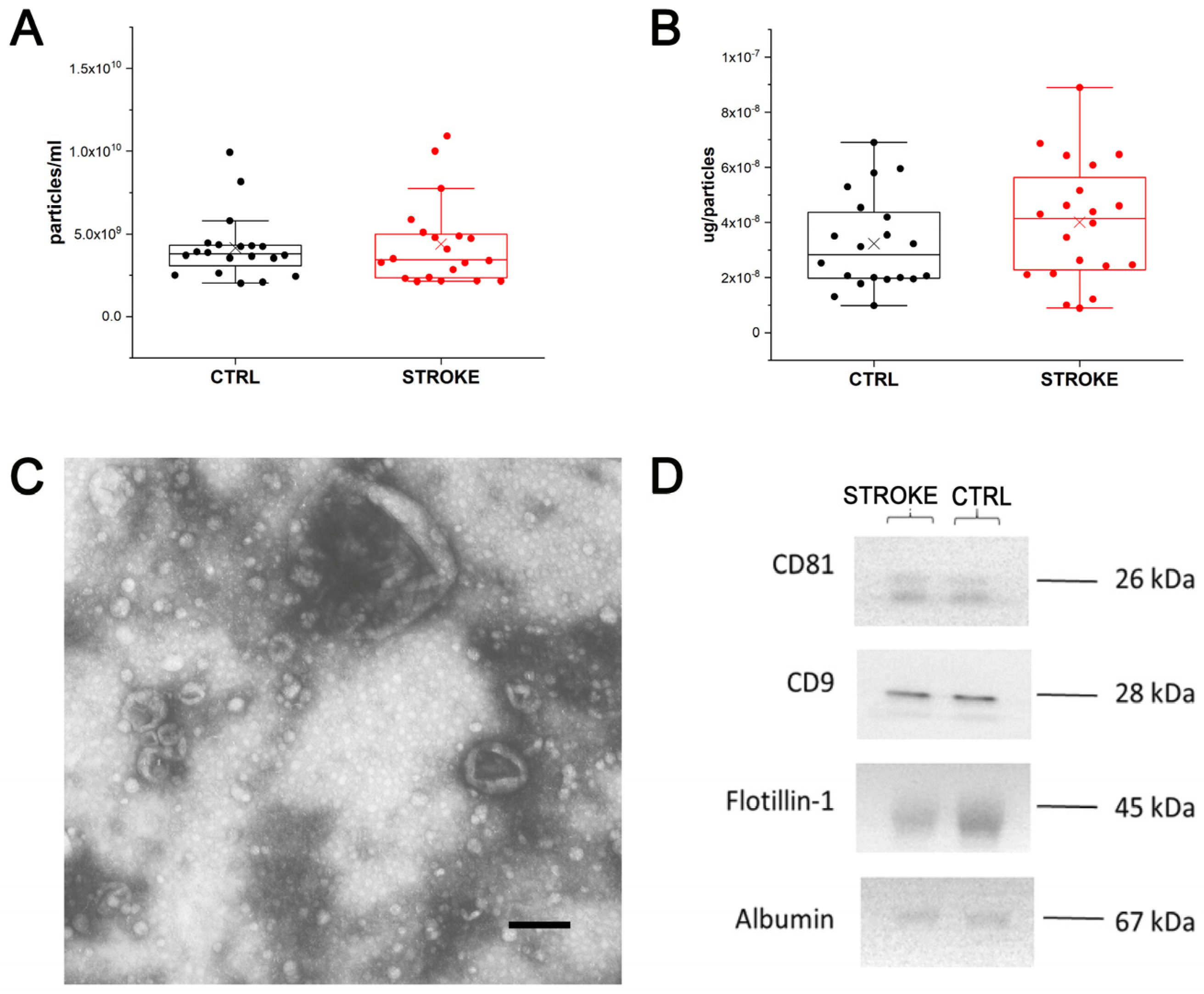
2.2. EV Physico-Chemical Characterization
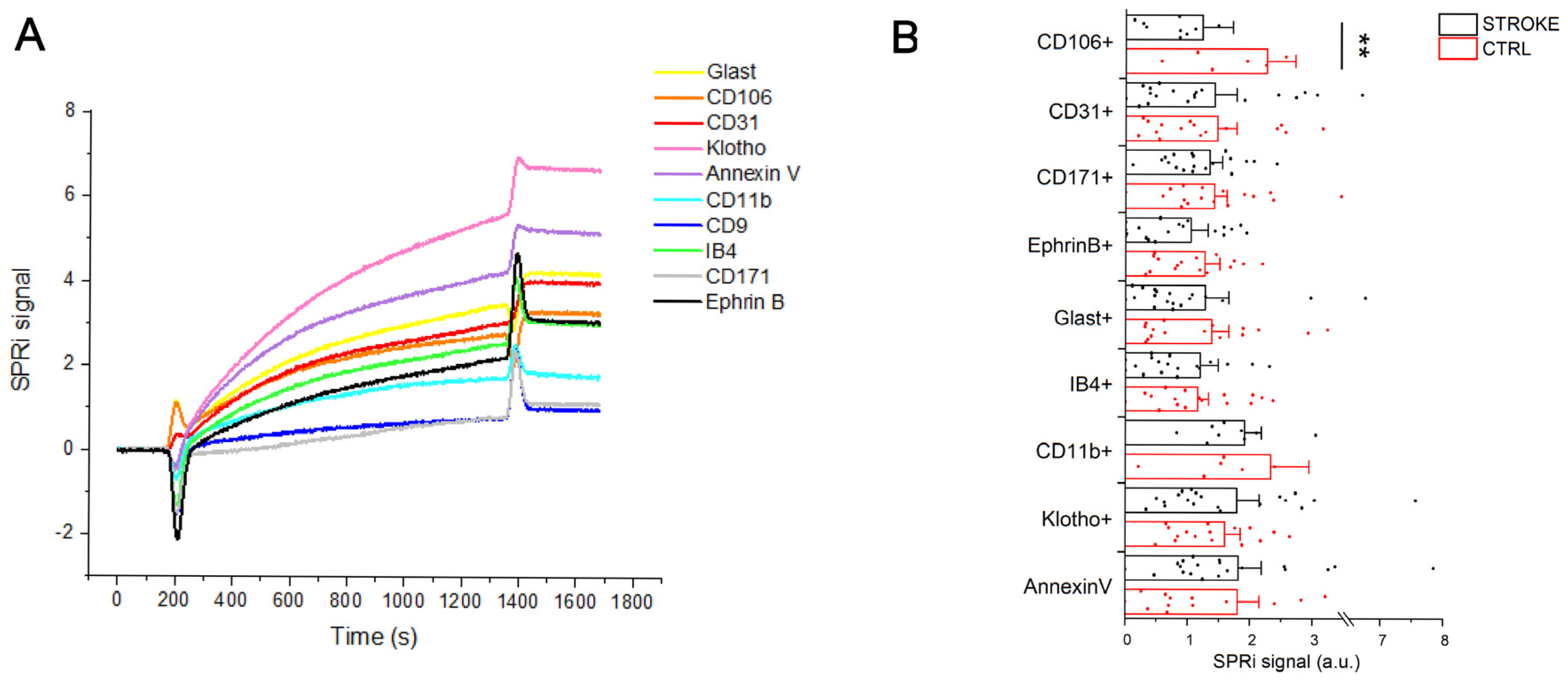
2.3. SPRi Analysis
2.4. Correlation Study
3. Discussion
4. Materials and Methods
4.1. Patients Recruitment and Samples Collection
4.2. Enzyme-Linked Immunosorbent Assay on Serum Samples
4.3. Isolation and Characterization of Extracellular Vesicles
4.4. SPRi Analysis
4.4.1. Biosensor Preparation and EV Detection
4.4.2. Gold Nanoparticle Preparation and Secondary EV Labelling
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bull, F.; Goenka, S.; Lambert, V.; Pratt, M. Physical Activity for the Prevention of Cardiometabolic Disease. In Disease Control Priorities, Third Edition (Volume 5): Cardiovascular, Respiratory, and Related Disorders; World Bank Publications: Washington, DC, USA, 2017; pp. 79–99. [Google Scholar] [CrossRef]
- Taricco, M.; Cecchi, F.; Cassio, A.; Lavezzi, S.; Scarponi, F.; Montis, A.; Gatta, G.; Bernucci, C.; Franceschini, M.; Paolucci, S.; et al. PMIC 2020 Protocollo Di Valutazione Riabilitativa Di Minima Della Persona Con Ictus Cerebrale Versione 2020. Soc. Ital. Med. Fis. 2020, 34, 11–37. [Google Scholar]
- Gualerzi, A.; Picciolini, S.; Rodà, F.; Bedoni, M. Extracellular Vesicles in Regeneration and Rehabilitation Recovery after Stroke. Biology 2021, 10, 843. [Google Scholar] [CrossRef] [PubMed]
- Bembenek, J.P.; Kurczych, K.; Kłysz, B.; Cudna, A.; Antczak, J.; Członkowska, A. Prediction of Recovery and Outcome Using Motor Evoked Potentials and Brain Derived Neurotrophic Factor in Subacute Stroke. J. Stroke Cerebrovasc. Dis. 2020, 29, 105202. [Google Scholar] [CrossRef]
- Gandolfi, M.; Smania, N.; Vella, A.; Picelli, A.; Chirumbolo, S. Assessed and Emerging Biomarkers in Stroke and Training-Mediated Stroke Recovery: State of the Art. Neural Plast. 2017, 2017, 1389475. [Google Scholar] [CrossRef] [PubMed]
- Carandina, A.; Favero, C.; Sacco, R.M.; Hoxha, M.; Torgano, G.; Montano, N.; Bollati, V.; Tobaldini, E. The Role of Extracellular Vesicles in Ischemic Stroke Severity. Biology 2022, 11, 1489. [Google Scholar] [CrossRef]
- Dewdney, B.; Trollope, A.; Moxon, J.; Thomas Manapurathe, D.; Biros, E.; Golledge, J. Circulating MicroRNAs as Biomarkers for Acute Ischemic Stroke: A Systematic Review. J. Stroke Cerebrovasc. Dis. 2018, 27, 522–530. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, P.; Zhang, F.; Chen, T. Association of MicroRNAs With Risk of Stroke: A Meta-Analysis. Front. Neurol. 2022, 13, 865265. [Google Scholar] [CrossRef]
- Fullerton, J.L.; Cosgrove, C.C.; Rooney, R.A.; Work, L.M. Extracellular Vesicles and Their MicroRNA Cargo in Ischaemic Stroke. J. Physiol. 2022. [Google Scholar] [CrossRef]
- Erdbrügger, U.; Lannigan, J. Analytical Challenges of Extracellular Vesicle Detection: A Comparison of Different Techniques: Analytical Challenges of Extracellular Vesicle Detection. Cytometry 2016, 89, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Gualerzi, A.; Picciolini, S.; Carlomagno, C.; Rodà, F.; Bedoni, M. Biophotonics for Diagnostic Detection of Extracellular Vesicles. Adv. Drug Deliv. Rev. 2021, 174, 229–249. [Google Scholar] [CrossRef]
- Hartjes, T.; Mytnyk, S.; Jenster, G.; van Steijn, V.; van Royen, M. Extracellular Vesicle Quantification and Characterization: Common Methods and Emerging Approaches. Bioengineering 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, V.; Lucchetti, D.; Gatto, I.; Maiorana, A.; Marcantoni, M.; Maulucci, G.; Papi, M.; Pola, R.; De Spirito, M.; Sgambato, A. Dynamic Light Scattering for the Characterization and Counting of Extracellular Vesicles: A Powerful Noninvasive Tool. J. Nanopart. Res. 2014, 16, 2583. [Google Scholar] [CrossRef]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef]
- Serrano-Pertierra, E.; Oliveira-Rodríguez, M.; Matos, M.; Gutiérrez, G.; Moyano, A.; Salvador, M.; Rivas, M.; Blanco-López, M.C. Extracellular Vesicles: Current Analytical Techniques for Detection and Quantification. Biomolecules 2020, 10, 824. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; LeClaire, M.; Gimzewski, J.K. Ascent of Atomic Force Microscopy as a Nanoanalytical Tool for Exosomes and Other Extracellular Vesicles. Nanotechnology 2018, 29, 132001. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kao, Y.; Zhou, Q.; Wuethrich, A.; Stark, M.S.; Schaider, H.; Soyer, H.P.; Lin, L.L.; Trau, M. An Integrated Microfluidic-SERS Platform Enables Sensitive Phenotyping of Serum Extracellular Vesicles in Early Stage Melanomas. Adv. Funct. Mater. 2022, 32, 2010296. [Google Scholar] [CrossRef]
- Picciolini, S.; Gualerzi, A.; Vanna, R.; Sguassero, A.; Gramatica, F.; Bedoni, M.; Masserini, M.; Morasso, C. Detection and Characterization of Different Brain-Derived Subpopulations of Plasma Exosomes by Surface Plasmon Resonance Imaging. Anal. Chem. 2018, 90, 8873–8880. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Lasek-Bal, A.; Jędrzejowska-Szypułka, H.; Różycka, J.; Bal, W.; Holecki, M.; Duława, J.; Lewin-Kowalik, J. Low Concentration of BDNF in the Acute Phase of Ischemic Stroke as a Factor in Poor Prognosis in Terms of Functional Status of Patients. Med. Sci. Monit. 2015, 21, 3900–3905. [Google Scholar] [CrossRef]
- Shah, S.; Vanclay, F.; Cooper, B. Improving the Sensitivity of the Barthel Index for Stroke Rehabilitation. J. Clin. Epidemiol. 1989, 42, 703–709. [Google Scholar] [CrossRef]
- Sahu, A.; Mamiya, H.; Shinde, S.N.; Cheikhi, A.; Winter, L.L.; Vo, N.V.; Stolz, D.; Roginskaya, V.; Tang, W.Y.; Croix, C.S.; et al. Age-Related Declines in α-Klotho Drive Progenitor Cell Mitochondrial Dysfunction and Impaired Muscle Regeneration. Nat. Commun. 2018, 9, 4859. [Google Scholar] [CrossRef]
- Lee, J.B.; Woo, H.G.; Chang, Y.; Jin, Y.M.; Jo, I.; Kim, J.; Song, T.J. Plasma Klotho Concentrations Predict Functional Outcome at Three Months after Acute Ischemic Stroke Patients. Ann. Med. 2019, 51, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Picciolini, S.; Gualerzi, A.; Carlomagno, C.; Cabinio, M.; Sorrentino, S.; Baglio, F.; Bedoni, M. An SPRi-Based Biosensor Pilot Study: Analysis of Multiple Circulating Extracellular Vesicles and Hippocampal Volume in Alzheimer’s Disease. J. Pharm. Biomed. Anal. 2021, 192, 113649. [Google Scholar] [CrossRef] [PubMed]
- Tallon, C.; Picciolini, S.; Yoo, S.; Thomas, A.G.; Pal, A.; Alt, J.; Carlomagno, C.; Gualerzi, A.; Rais, R.; Haughey, N.J.; et al. Inhibition of Neutral Sphingomyelinase 2 Reduces Extracellular Vesicle Release from Neurons, Oligodendrocytes, and Activated Microglial Cells Following Acute Brain Injury. Biochem. Pharmacol. 2021, 194, 114796. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Diagnostic and Therapeutic Potential of TSPO Studies Regarding Neurodegenerative Diseases, Psychiatric Disorders, Alcohol Use Disorders, Traumatic Brain Injury, and Stroke: An Update. Cells 2020, 9, 870. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Buller, B.; Chopp, M. Exosomes—Beyond Stem Cells for Restorative Therapy in Stroke and Neurological Injury. Nat. Rev. Neurol. 2019, 15, 193–203. [Google Scholar] [CrossRef]
- Yousif, G.; Qadri, S.; Haik, M.; Haik, Y.; Parray, A.S.; Shuaib, A. Circulating Exosomes of Neuronal Origin as Potential Early Biomarkers for Development of Stroke. Mol. Diagn. Ther. 2021, 25, 163–180. [Google Scholar] [CrossRef]
- Bitencourt, A.C.S.; Timóteo, R.P.; Bazan, R.; Silva, M.V.; Filho, L.G.d.S.; Ratkevicius, C.M.A.; de Assunção, T.S.F.; de Oliveira, A.P.S.; Luvizutto, G.J. Association of Proinflammatory Cytokine Levels with Stroke Severity, Infarct Size, and Muscle Strength in the Acute Phase of Stroke. J. Stroke Cerebrovasc. Dis. 2022, 31, 106187. [Google Scholar] [CrossRef]
- Wytrykowska, A.; Prosba-Mackiewicz, M.; Nyka, W.M. IL-1β, TNF-α, and IL-6 Levels in Gingival Fluid and Serum of Patients with Ischemic Stroke. J. Oral Sci. 2016, 58, 509–513. [Google Scholar] [CrossRef]
- Intiso, D.; Zarrelli, M.M.; Lagioia, G.; Rienzo, F.D.; Ambrosio, C.C.D.; Simone, P.; Tonali, P.; Cioffi, R.P. Tumor Necrosis Factor Alpha Serum Levels and Inflammatory Response in Acute Ischemic Stroke Patients. Neurol. Sci. 2004, 24, 390–396. [Google Scholar] [CrossRef]
- Wu, B.N.; Wu, J.; Hao, D.L.; Mao, L.L.; Zhang, J.; Huang, T.T. High Serum SICAM-1 Is Correlated with Cerebral Microbleeds and Hemorrhagic Transformation in Ischemic Stroke Patients. Br. J. Neurosurg. 2018, 32, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Delgado, P.; Cuadrado, E.; Rosell, A.; Alvarez-Sabin, J.; Ortega-Aznar, A.; Hernandez-Guillamon, M.; Penalba, A.; Molina, C.A.; Montaner, J. Fas System Activation in Perihematomal Areas after Spontaneous Intracerebral Hemorrhage. Stroke 2008, 39, 1730–1734. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Akbar, N.; Davis, S.; Fischer, R.; Dickens, A.M.; Neuhaus, A.A.; Burgess, A.I.; Rothwell, P.M.; Buchan, A.M. Inflammatory Stroke Extracellular Vesicles Induce Macrophage Activation. Stroke 2017, 48, 2292–2296. [Google Scholar] [CrossRef]
- Santos, G.L.; Alcântara, C.C.; Silva-Couto, M.A.; García-Salazar, L.F.; Russo, T.L. Decreased Brain-Derived Neurotrophic Factor Serum Concentrations in Chronic Post-Stroke Subjects. J. Stroke Cerebrovasc. Dis. 2016, 25, 2968–2974. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.J.; Li, H.; Shi, M.Q.; Mao, X.N.; Liu, D.L.; Chang, Y.R.; Gan, Y.M.; Kuang, X.; Du, J.R. Protective Effect of Klotho against Ischemic Brain Injury Is Associated with Inhibition of RIG-I/NF-ΚB Signaling. Front. Pharmacol. 2018, 8, 950. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Clemens, Z.J.; Shinde, S.N.; Sivakumar, S.; Pius, A.; Bhatia, A.; Picciolini, S.; Carlomagno, C.; Gualerzi, A.; Bedoni, M.; et al. Regulation of Aged Skeletal Muscle Regeneration by Circulating Extracellular Vesicles. Nat. Aging 2021, 1, 1148–1161. [Google Scholar] [CrossRef]
- Bonaventura, A.; Liberale, L.; Vecchié, A.; Casula, M.; Carbone, F.; Dallegri, F.; Montecucco, F. Update on Inflammatory Biomarkers and Treatments in Ischemic Stroke. Int. J. Mol. Sci. 2016, 17, 1967. [Google Scholar] [CrossRef]
- Roy Choudhury, G.; Ryou, M.-G.; Poteet, E.; Wen, Y.; He, R.; Sun, F.; Yuan, F.; Jin, K.; Yang, S.-H. Involvement of P38 MAPK in Reactive Astrogliosis Induced by Ischemic Stroke. Brain Res. 2014, 1551, 45–58. [Google Scholar] [CrossRef]
- Brenna, S.; Altmeppen, H.C.; Mohammadi, B.; Rissiek, B.; Schlink, F.; Ludewig, P.; Krisp, C.; Schlüter, H.; Failla, A.V.; Schneider, C.; et al. Characterization of Brain-derived Extracellular Vesicles Reveals Changes in Cellular Origin after Stroke and Enrichment of the Prion Protein with a Potential Role in Cellular Uptake. J. Extracell. Vesicles 2020, 9, 1809065. [Google Scholar] [CrossRef]
- Barthels, D.; Das, H. Current Advances in Ischemic Stroke Research and Therapies. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165260. [Google Scholar] [CrossRef]
- Chen, M.-K.; Guilarte, T.R. Translocator Protein 18 KDa (TSPO): Molecular Sensor of Brain Injury and Repair. Pharmacol. Ther. 2008, 118, 1–17. [Google Scholar] [CrossRef]
- Betlazar, C.; Harrison-Brown, M.; Middleton, R.J.; Banati, R.; Liu, G.-J. Cellular Sources and Regional Variations in the Expression of the Neuroinflammatory Marker Translocator Protein (TSPO) in the Normal Brain. Int. J. Mol. Sci. 2018, 19, 2707. [Google Scholar] [CrossRef]
- Kozuka, K.; Kohriyama, T.; Ikeda, J.; Nakamura, S.; Nomura, E.; Kajikawa, H. Endothelial Markers and Adhesion Molecules in Acute Ischemic Stroke-Sequential Change and Differences in Stroke Subtype. Atherosclerosis 2002, 161, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Pianta, S.; Lee, J.Y.; Tuazon, J.P.; Castelli, V.; Mantohac, L.M.; Tajiri, N.; Borlongan, C.V. A Short Bout of Exercise Prior to Stroke Improves Functional Outcomes by Enhancing Angiogenesis. Neuromol. Med. 2019, 21, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Poststroke Angiogenesis: Blood, Bloom, or Brood? Stroke A J. Cereb. Circ. 2015, 46, e105–e106. [Google Scholar] [CrossRef] [PubMed]
- Sguassero, A.; Artiga, Á.; Morasso, C.; Jimenez, R.R.; Rapún, R.M.; Mancuso, R.; Agostini, S.; Hernis, A.; Abols, A.; Linē, A.; et al. A Simple and Universal Enzyme-Free Approach for the Detection of Multiple MicroRNAs Using a Single Nanostructured Enhancer of Surface Plasmon Resonance Imaging. Anal. Bioanal. Chem. 2019, 411, 1873–1885. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Hastie, T. Regularization and Variable Selection via the Elastic Net. J. R. Statistical. Soc. B 2005, 67, 301–320. [Google Scholar] [CrossRef]
- Fan, J.; Li, R. Statistical Challenges with High Dimensionality: Feature Selection in Knowledge Discovery. In Proceedings of the International Congress of Mathematicians, Madrid, Spain, 22–30 August 2006. [Google Scholar]
- Yamada, M.; Koh, T.; Iwata, T.; Shawe-Taylor, J.; Kaski, S. Localized Lasso for High-Dimensional Regression. In Proceedings of the 20th International Conference on Artificial Intelligence and Statistics, Fort Lauderdale, FL, USA, 20–22 April 2017; pp. 325–333. Available online: https://proceedings.mlr.press/v54/yamada17a.html (accessed on 20 March 2023).
- Frens, G. Controlled Nucleation for the Regulation of the Particle Size in Monodisperse Gold Suspensions. Nat. Phys. Sci. 1973, 241, 20–22. [Google Scholar] [CrossRef]
- Ojea-Jiménez, I.; Bastús, N.G.; Puntes, V. Influence of the Sequence of the Reagents Addition in the Citrate-Mediated Synthesis of Gold Nanoparticles. J. Phys. Chem. C 2011, 115, 15752–15757. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Stroke (n = 19) | Control (n = 20) |
|---|---|---|
| Age, years | 74 (18) | 66 (8) |
| Sex, female | 8 (42.1%) | 10 (50.0%) |
| Modified Barthel Index admission, points | 30 (26) | -- |
| Modified Barthel Index at discharge, points | 56 (54) | -- |
| Time post onset, days | 21 (14) | -- |
| Relapsing patients, number | 8 (42.1%) | -- |
| Patients with diabetes, number | 10 (62.6%) | -- |
| Independent Variables | MBI at Admission | MBI at Discharge | |||
|---|---|---|---|---|---|
| R2 | p | R2 | p | ||
| SPRi | CD11b+ | 0.694 | 0.056 | 0.881 | 0.004 |
| VEGFR2 on CD11b+ | −0.680 | 0.044 | −0.644 | 0.061 | |
| VEGFR2 on IB4+ | −0.244 | 0.330 | −0.563 | 0.015 | |
| TSPO on CD106+ | 0.671 | 0.048 | 0.402 | 0.283 | |
| TSPO on CD31+ | 0.487 | 0.035 | 0.694 | 0.001 | |
| TSPO on CD9+ | 0.458 | 0.049 | 0.258 | 0.287 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picciolini, S.; Mangolini, V.; Rodà, F.; Montesano, A.; Arnaboldi, F.; Liuzzi, P.; Mannini, A.; Bedoni, M.; Gualerzi, A. Multiplexing Biosensor for the Detection of Extracellular Vesicles as Biomarkers of Tissue Damage and Recovery after Ischemic Stroke. Int. J. Mol. Sci. 2023, 24, 7937. https://doi.org/10.3390/ijms24097937
Picciolini S, Mangolini V, Rodà F, Montesano A, Arnaboldi F, Liuzzi P, Mannini A, Bedoni M, Gualerzi A. Multiplexing Biosensor for the Detection of Extracellular Vesicles as Biomarkers of Tissue Damage and Recovery after Ischemic Stroke. International Journal of Molecular Sciences. 2023; 24(9):7937. https://doi.org/10.3390/ijms24097937
Chicago/Turabian StylePicciolini, Silvia, Valentina Mangolini, Francesca Rodà, Angelo Montesano, Francesca Arnaboldi, Piergiuseppe Liuzzi, Andrea Mannini, Marzia Bedoni, and Alice Gualerzi. 2023. "Multiplexing Biosensor for the Detection of Extracellular Vesicles as Biomarkers of Tissue Damage and Recovery after Ischemic Stroke" International Journal of Molecular Sciences 24, no. 9: 7937. https://doi.org/10.3390/ijms24097937
APA StylePicciolini, S., Mangolini, V., Rodà, F., Montesano, A., Arnaboldi, F., Liuzzi, P., Mannini, A., Bedoni, M., & Gualerzi, A. (2023). Multiplexing Biosensor for the Detection of Extracellular Vesicles as Biomarkers of Tissue Damage and Recovery after Ischemic Stroke. International Journal of Molecular Sciences, 24(9), 7937. https://doi.org/10.3390/ijms24097937