
The Role of Nitric Oxide in the Micro- and Macrovascular Response to a 7-Day High-Salt Diet in Healthy Individuals
 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Arterial Blood Pressure and Systemic Hemodynamic
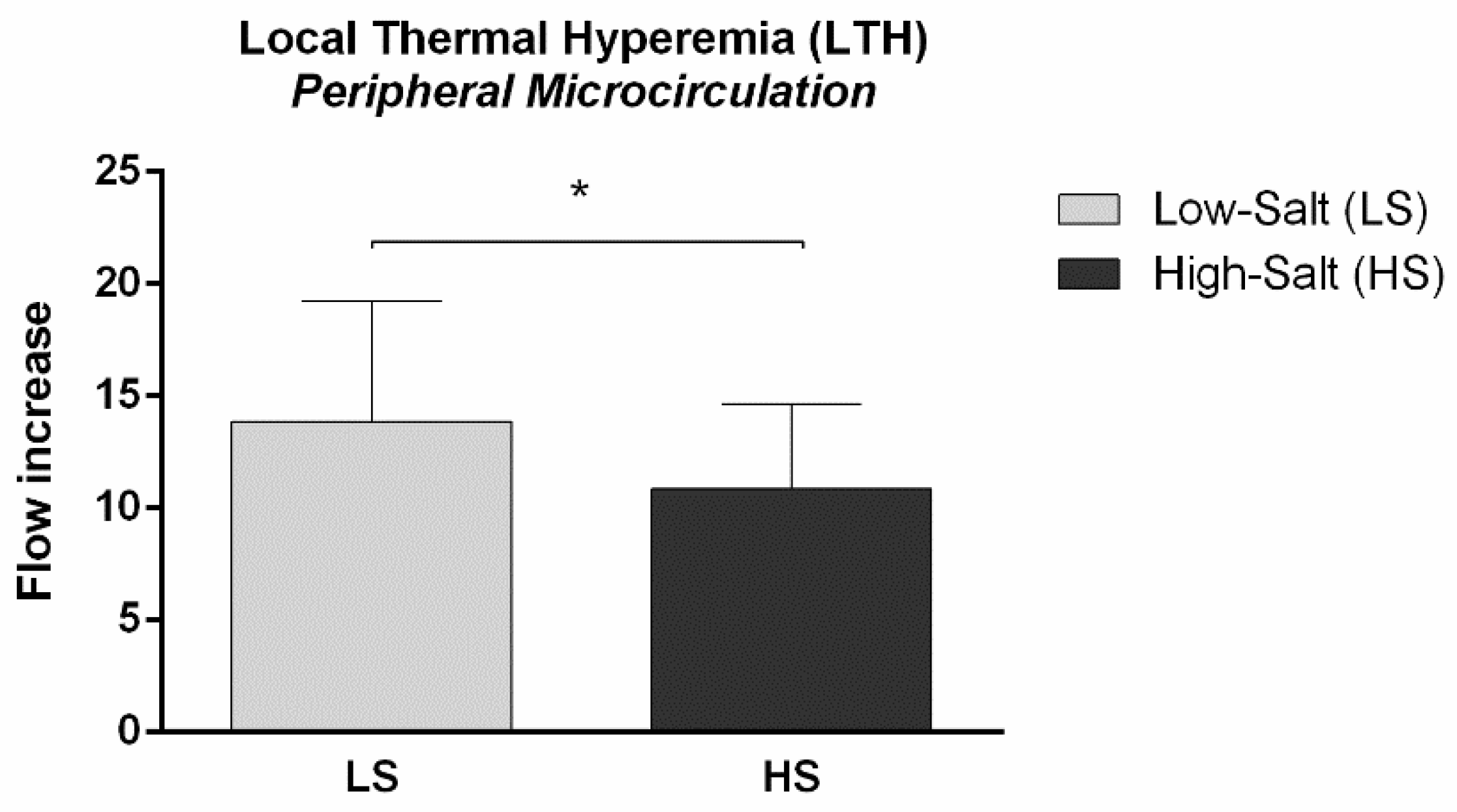
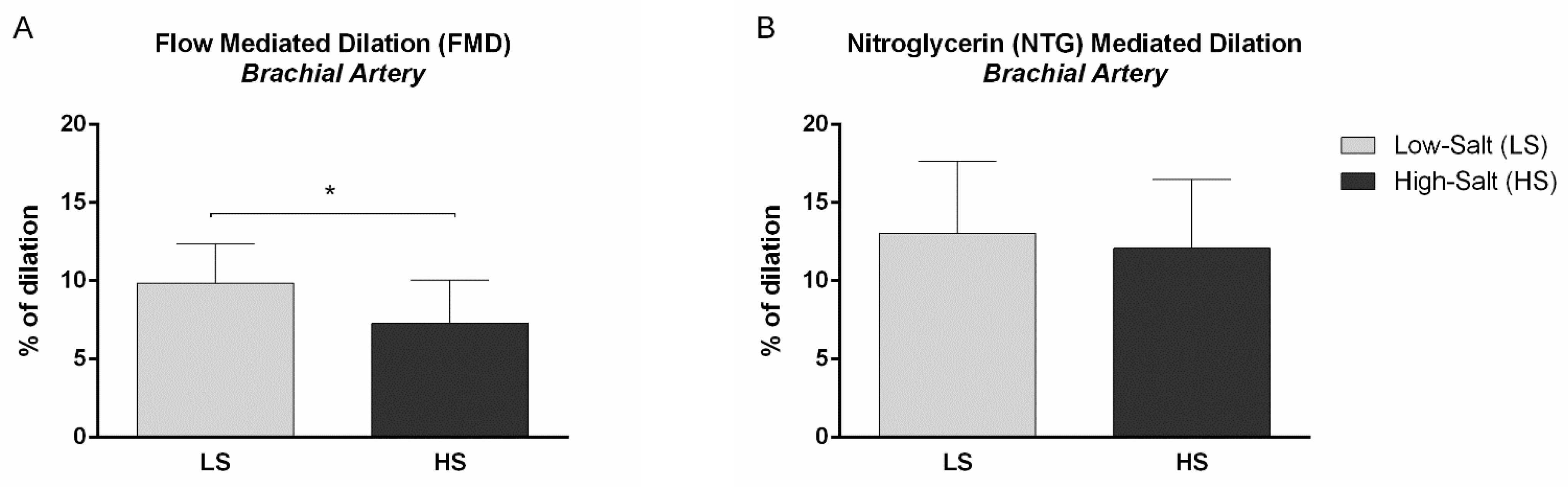
2.2. Micro- and Macrovascular NO-Dependent Endothelial Vasodilation
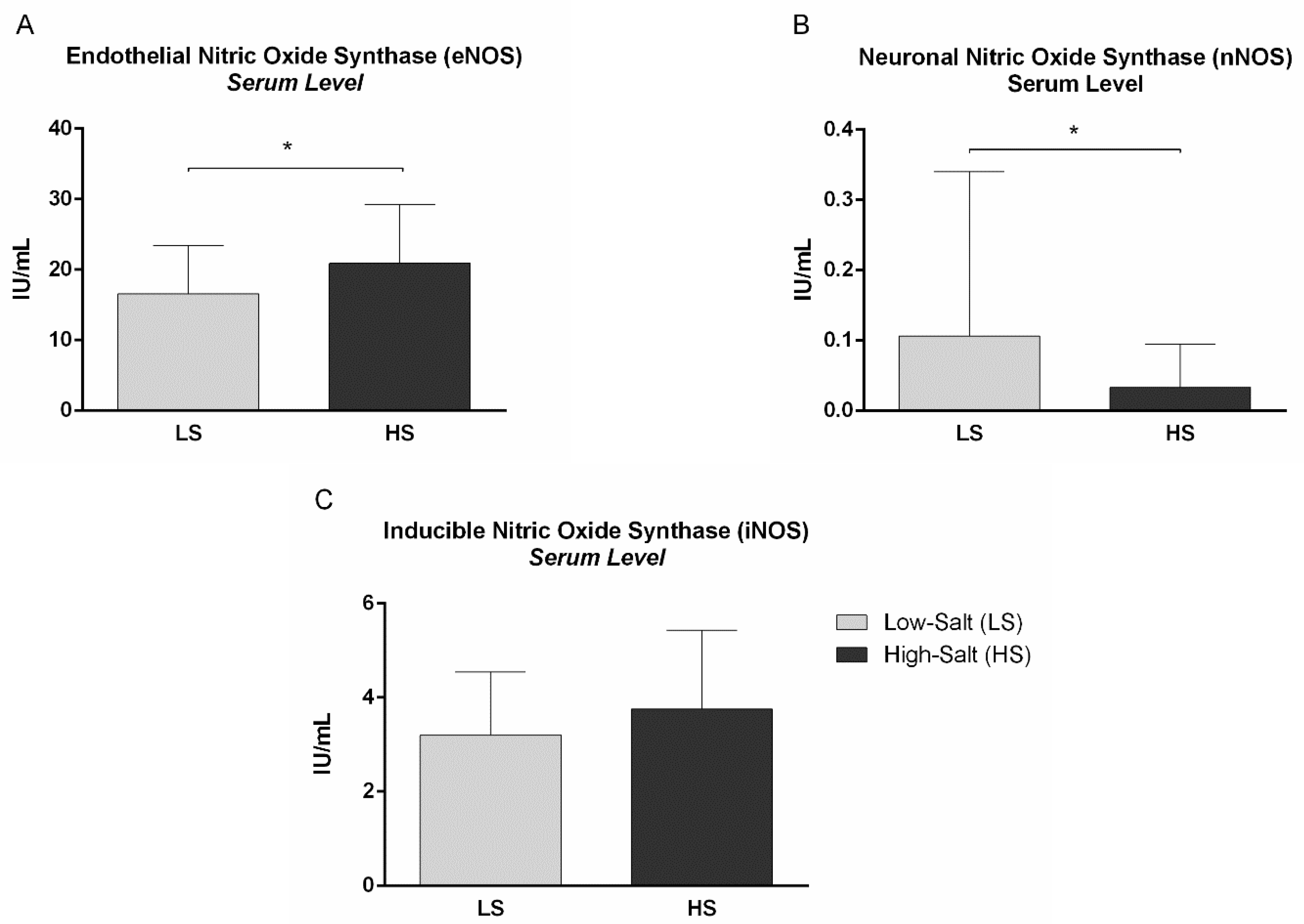
2.3. Nitric Oxide, Nitric Oxide Synthases and Vascular Endothelial Growth Factor C Serum Levels
2.4. Correlations
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Study Design
4.3. Weight Status, Body Composition and Body Fluid Status
4.4. Venous Blood and 24-h Urine Sampling and Analysis
4.5. Systemic Hemodynamic and Arterial Blood Pressure
4.6. Peripheral Microvascular Response to Local Thermal Hyperemia (LTH)
4.7. Flow-Mediated Dilation (FMD) of Brachial Artery
4.8. Serum Nitric Oxide (NO) and Three Isoforms of Nitric Oxide Synthase (NOS) Assay
4.9. Serum Vascular Endothelial Growth Factor C (VEGF-C) Assay
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef]
- Drenjančević-Perić, I.; Jelaković, B.; Lombard, J.H.; Kunert, M.P.; Kibel, A.; Gros, M. High-Salt Diet and Hypertension: Focus on the Renin-Angiotensin System. Kidney Blood Press. Res. 2011, 34, 1–11. [Google Scholar] [CrossRef]
- Jelakovic, B.; Vrdoljak, A.; Pecin, I.; Buzjak, V.; Karanovic, S.; Ivkovic, V.; Dapic, K.; Domislovic, V.; Reiner, Z. Less salt—More health. Croatian Action on Salt and Health (CRASH). J. Hypertens Res. 2016, 2, 61–68. [Google Scholar]
- Jelakovic, B. Salt Intake in Croatia—EHUH 2 Report. In Proceedings of the 41st Symposium Hypertension Highlights in 2020, Rijeka, Croatia, 19 November 2020. [Google Scholar]
- Bragulat, E.; de la Sierra, A. Salt intake, endothelial dysfunction, and salt-sensitive hypertension. J. Clin. Hypertens. 2002, 4, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, M.H. Salt sensitivity is associated with an increased mortality in both normal and hypertensive humans. J. Clin. Hypertens. 2002, 4, 274–276. [Google Scholar] [CrossRef]
- Greaney, J.L.; DuPont, J.J.; Lennon-Edwards, S.L.; Sanders, P.W.; Edwards, D.G.; Farquhar, W.B. Dietary sodium loading impairs microvascular function independent of blood pressure in humans: Role of oxidative stress. J. Physiol. 2012, 590, 5519–5528. [Google Scholar] [CrossRef]
- Cavka, A.; Cosic, A.; Jukic, I.; Jelakovic, B.; Lombard, J.H.; Phillips, S.A.; Seric, V.; Mihaljevic, I.; Drenjancevic, I. The role of cyclo-oxygenase-1 in high-salt diet-induced microvascular dysfunction in humans. J. Physiol. 2015, 593, 5313–5324. [Google Scholar] [CrossRef]
- Cavka, A.; Jukic, I.; Ali, M.; Goslawski, M.; Bian, J.T.; Wang, E.; Drenjancevic, I.; Phillips, S.A. Short-term high salt intake reduces brachial artery and microvascular function in the absence of changes in blood pressure. J. Hypertens. 2016, 34, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Barić, L.; Drenjančević, I.; Matić, A.; Stupin, M.; Kolar, L.; Mihaljević, Z.; Lenasi, H.; Šerić, V.; Stupin, A. Seven-Day Salt Loading Impairs Microvascular Endothelium-Dependent Vasodilation without Changes in Blood Pressure, Body Composition and Fluid Status in Healthy Young Humans. Kidney Blood Press. Res. 2019, 44, 835–847. [Google Scholar] [CrossRef]
- Gimbrone, M.A.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Stein, C.M.; Nelson, R.; Brown, M.; Wood, M.; Wood, A.J.J. Dietary Sodium Intake Modulates Vasodilation Mediated by Nitroprusside but Not by Methacholine in the Human Forearm. Hypertension 1995, 25, 1220–1223. [Google Scholar] [CrossRef]
- Dishy, V.; Sofowora, G.G.; Imamura, H.; Nishimi, Y.; Xie, H.-G.; Wood, A.J.; Stein, C.M. Nitric oxide production decreases after salt loading but is not related to blood pressure changes or nitric oxide-mediated vascular responses. J. Hypertens. 2003, 21, 153–157. [Google Scholar] [CrossRef]
- Tzemos, N.; Lim, P.O.; Wong, S.; Struthers, A.D.; MacDonald, T.M. Adverse Cardiovascular Effects of Acute Salt Loading in Young Normotensive Individuals. Hypertension 2008, 51, 1525–1530. [Google Scholar] [CrossRef]
- Eisenach, J.H.; Gullixson, L.R.; Kost, S.L.; Joyner, M.J.; Turner, S.T.; Nicholson, W.T. Sex differences in salt sensitivity to nitric oxide dependent vasodilation in healthy young adults. J. Appl. Physiol. 2012, 112, 1049–1053. [Google Scholar] [CrossRef]
- Dickinson, K.M.; Clifton, P.M.; Keogh, J.B. Endothelial function is impaired after a high-salt meal in healthy subjects. Am. J. Clin. Nutr. 2011, 93, 500–505. [Google Scholar] [CrossRef]
- Vanhoutte, P.M.; Shimokawa, H.; Feletou, M.; Tang, E.H.C. Endothelial dysfunction and vascular disease—A 30th anniversary update. Acta Physiol. 2017, 219, 22–96. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Anderson, T.J. Fundamentals of Endothelial Function for the Clinical Cardiologist. Circulation 2002, 105, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Boegehold, M.A.; Drenjancevic, I.; Lombard, J.H. Salt, Angiotensin II, Superoxide, and Endothelial Function. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 215–254. [Google Scholar]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Zhu, J.; Drenjancevic-Peric, I.; McEwen, S.; Friesema, J.; Schulta, D.; Yu, M.; Roman, R.J.; Lombard, J.H. Role of superoxide and angiotensin II suppression in salt-induced changes in endothelial Ca2+ signaling and NO production in rat aorta. Am. J. Physiol. Circ. Physiol. 2006, 291, H929–H938. [Google Scholar] [CrossRef] [PubMed]
- Barić, L.; Drenjančević, I.; Mihalj, M.; Matić, A.; Stupin, M.; Kolar, L.; Mihaljević, Z.; Mrakovčić-Šutić, I.; Šerić, V.; Stupin, A. Enhanced Antioxidative Defense by Vitamins C and E Consumption Prevents 7-Day High-Salt Diet-Induced Microvascular Endothelial Function Impairment in Young Healthy Individuals. J. Clin. Med. 2020, 9, 843. [Google Scholar] [CrossRef]
- Titze, J. Sodium balance is not just a renal affair. Curr. Opin. Nephrol. Hypertens. 2014, 23, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Titze, J.; Shakibaei, M.; Schafflhuber, M.; Schulze-Tanzil, G.; Porst, M.; Schwind, K.H.; Dietsch, P.; Hilgers, K.F. Glycosaminoglycan polymerization may enable osmotically inactive Na + storage in the skin. Am. J. Physiol. Circ. Physiol. 2004, 287, H203–H208. [Google Scholar] [CrossRef] [PubMed]
- Machnik, A.; Neuhofer, W.; Jantsch, J.; Dahlmann, A.; Tammela, T.; Machura, K.; Park, J.-K.; Beck, F.-X.; Müller, D.N.; Derer, W.; et al. Macrophages regulate salt-dependent volume and blood pressure by a vascular endothelial growth factor-C–dependent buffering mechanism. Nat. Med. 2009, 15, 545–552. [Google Scholar] [CrossRef]
- Ying, W.-Z.; Sanders, P.W. Dietary salt enhances glomerular endothelial nitric oxide synthase through TGF-β1. Am. J. Physiol. Physiol. 1998, 275, F18–F24. [Google Scholar] [CrossRef]
- Ni, Z.; Oveisi, F.; Vaziri, N.D. Nitric Oxide Synthase Isotype Expression in Salt-Sensitive and Salt-Resistant Dahl Rats. Hypertension 1999, 34, 552–557. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wang, X.Q.; Oveisi, F.; Rad, B. Induction of Oxidative Stress by Glutathione Depletion Causes Severe Hypertension in Normal Rats. Hypertension 2000, 36, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Drenjancevic-Peric, I.; Frisbee, J.C.; Lombard, J.H. Skeletal Muscle Arteriolar Reactivity in SS.BN13 Consomic Rats and Dahl Salt-Sensitive Rats. Hypertension 2003, 41, 1012–1015. [Google Scholar] [CrossRef]
- Durand, M.J.; Moreno, C.; Greene, A.S.; Lombard, J.H. Impaired relaxation of cerebral arteries in the absence of elevated salt intake in normotensive congenic rats carrying the Dahl salt-sensitive renin gene. Am. J. Physiol. Circ. Physiol. 2010, 299, H1865–H1874. [Google Scholar] [CrossRef] [PubMed]
- Roustit, M.; Cracowski, J.-L. Non-invasive Assessment of Skin Microvascular Function in Humans: An Insight into Methods. Microcirculation 2012, 19, 47–64. [Google Scholar] [CrossRef]
- DuPont, J.J.; Greaney, J.L.; Wenner, M.M.; Lennon-Edwards, S.L.; Sanders, P.W.; Farquhar, W.B.; Edwards, D.G. High dietary sodium intake impairs endothelium-dependent dilation in healthy salt-resistant humans. J. Hypertens. 2013, 31, 530–536. [Google Scholar] [CrossRef]
- Lennon-Edwards, S.; Ramick, M.G.; Matthews, E.L.; Brian, M.S.; Farquhar, W.B.; Edwards, D.G. Salt loading has a more deleterious effect on flow-mediated dilation in salt-resistant men than women. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 990–995. [Google Scholar] [CrossRef]
- Matthews, E.L.; Brian, M.S.; Ramick, M.G.; Lennon-Edwards, S.; Edwards, D.G.; Farquhar, W.B. High dietary sodium reduces brachial artery flow-mediated dilation in humans with salt-sensitive and salt-resistant blood pressure. J. Appl. Physiol. 2015, 118, 1510–1515. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Ally, A.; Powell, I.; Ally, M.M.; Chaitoff, K.; Nauli, S.M. Role of neuronal nitric oxide synthase on cardiovascular functions in physiological and pathophysiological states. Nitric Oxide 2020, 102, 52–73. [Google Scholar] [CrossRef] [PubMed]
- Knezović, A.; Kolobarić, N.; Drenjančević, I.; Mihaljević, Z.; Šušnjara, P.; Jukić, I.; Stupin, M.; Kibel, A.; Marczi, S.; Mihalj, M.; et al. Role of Oxidative Stress in Vascular Low-Grade Inflammation Initiation Due to Acute Salt Loading in Young Healthy Individuals. Antioxidants 2022, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wallerath, T.; Münzel, T.; Förstermann, U. Regulation of endothelial-type NO synthase expression in pathophysiology and in response to drugs. Nitric Oxide 2002, 7, 149–164. [Google Scholar] [CrossRef]
- Drummond, G.R.; Cai, H.; Davis, M.E.; Ramasamy, S.; Harrison, D.G. Transcriptional and Posttranscriptional Regulation of Endothelial Nitric Oxide Synthase Expression by Hydrogen Peroxide. Circ. Res. 2000, 86, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Mori, T.; Huang, T.; Lombard, J.H. Effect of high-salt diet on NO release and superoxide production in rat aorta. Am. J. Physiol. Circ. Physiol. 2004, 286, H575–H583. [Google Scholar] [CrossRef] [PubMed]
- Nurkiewicz, T.R.; Boegehold, M.A. High salt intake reduces endothelium-dependent dilation of mouse arterioles via superoxide anion generated from nitric oxide synthase. Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R1550–R1556. [Google Scholar] [CrossRef]
- Cosic, A.; Jukic, I.; Stupin, A.; Mihalj, M.; Mihaljevic, Z.; Novak, S.; Vukovic, R.; Drenjancevic, I. Attenuated flow-induced dilatation of middle cerebral arteries is related to increased vascular oxidative stress in rats on a short-term high salt diet. J. Physiol. 2016, 594, 4917–4931. [Google Scholar] [CrossRef]
- Heer, M.; Baisch, F.; Kropp, J.; Gerzer, R.; Drummer, C. High dietary sodium chloride consumption may not induce body fluid retention in humans. Am. J. Physiol. Physiol. 2000, 278, F585–F595. [Google Scholar] [CrossRef]
- Qi, N.; Rapp, J.P.; Brand, P.H.; Metting, P.J.; Britton, S.L. Body fluid expansion is not essential for salt-induced hypertension in SS/Jr rats. Am. J. Physiol. Integr. Comp. Physiol. 1999, 277, R1392–R1400. [Google Scholar] [CrossRef] [PubMed]
- Titze, J.; Krause, H.; Hecht, H.; Dietsch, P.; Rittweger, J.; Lang, R.; Kirsch, K.A.; Hilgers, K.F. Reduced osmotically inactive Na storage capacity and hypertension in the Dahl model. Am. J. Physiol. Physiol. 2002, 283, F134–F141. [Google Scholar] [CrossRef]
- Dahlmann, A.; Dörfelt, K.; Eicher, F.; Linz, P.; Kopp, C.; Mössinger, I.; Horn, S.; Büschges-Seraphin, B.; Wabel, P.; Hammon, M.; et al. Magnetic resonance–determined sodium removal from tissue stores in hemodialysis patients. Kidney Int. 2015, 87, 434–441. [Google Scholar] [CrossRef]
- Slagman, M.C.J.; Kwakernaak, A.J.; Yazdani, S.; Laverman, G.D.; van den Born, J.; Titze, J.; Navis, G. Vascular endothelial growth factor C levels are modulated by dietary salt intake in proteinuric chronic kidney disease patients and in healthy subjects. Nephrol. Dial. Transplant. 2012, 27, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Soria, J.-C.; DeBraud, F.; Bahleda, R.; Adamo, B.; Andre, F.; Dientsmann, R.; Delmonte, A.; Cereda, R.; Isaacson, J.; Litten, J.; et al. Phase I/IIa study evaluating the safety, efficacy, pharmacokinetics, and pharmacodynamics of lucitanib in advanced solid tumors. Ann. Oncol. 2014, 25, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Lankhorst, S.; Severs, D.; Markó, L.; Rakova, N.; Titze, J.; Müller, D.N.; Danser, A.H.J.; van den Meiracker, A.H. Salt Sensitivity of Angiogenesis Inhibition–Induced Blood Pressure Rise. Hypertension 2017, 69, 919–926. [Google Scholar] [CrossRef]
- Bryan, N.S. Nitrite in nitric oxide biology: Cause or consequence? Free. Radic. Biol. Med. 2006, 41, 691–701. [Google Scholar] [CrossRef]
- Stupin, A.; Drenjančević, I.; Šušnjara, P.; Debeljak, Ž.; Kolobarić, N.; Jukić, I.; Mihaljević, Z.; Martinović, G.; Selthofer-Relatić, K. Is There Association between Altered Adrenergic System Activity and Microvascular Endothelial Dysfunction Induced by a 7-Day High Salt Intake in Young Healthy Individuals. Nutrients 2021, 13, 1731. [Google Scholar] [CrossRef]
- Brunt, V.E.; Minson, C.T. Cutaneous thermal hyperemia: More than skin deep. J. Appl. Physiol. 2011, 111, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Flammer, A.J.; Anderson, T.; Celermajer, D.S.; Creager, M.A.; Deanfield, J.; Ganz, P.; Hamburg, N.M.; Lüscher, T.F.; Shechter, M.; Taddei, S.; et al. The Assessment of Endothelial Function. Circulation 2012, 126, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, D.H.J.; Black, M.A.; Pyke, K.E.; Padilla, J.; Atkinson, G.; Harris, R.A.; Parker, B.; Widlansky, M.E.; Tschakovsky, M.E.; Green, D.J. Assessment of flow-mediated dilation in humans: A methodological and physiological guideline. Am. J. Physiol. Circ. Physiol. 2011, 300, H2–H12. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | LS | HS |
|---|---|---|
| N (W/M) | 46 (24/22) | |
| Age (years) | 21 [18–24] | |
| Weight Status | ||
| BMI (kg/m2) | 23.1 [17.8–29.9] | 23.1 [17.8–29.9] |
| Waist Circumference (cm) | 77.8 ± 9.4 | 77.6 ± 9.3 |
| Body Composition and Fluid Status | ||
| Fat Free Mass (%) | 82.0 ± 5.7 | 82.3 ± 5.4 |
| Fat (%) | 18.0 ± 5.7 | 17.7 ± 5.4 |
| Total Body Water (%) | 61.4 ± 6.3 | 61.4 ± 4.9 |
| Extracellular Water (%) | 42.9 ± 1.4 | 42.8 ± 1.2 |
| Intracellular Water (%) | 57.1 ± 1.4 | 57.2 ± 1.2 |
| ECW/ICW | 0.75 ± 0.04 | 0.75 ± 0.04 |
| Plasma Fluid (L) | 3.8 ± 0.8 | 3.8 ± 0.9 |
| Interstitial Fluid (L) | 13.4 ± 2.8 | 13.4 ± 3.2 |
| Body Density (kg/L) | 1.06 ± 0.01 | 1.06 ± 0.01 |
| Parameter | LS | HS | Reference Range † |
|---|---|---|---|
| N (W/M) | 46 (24/22) | ||
| Serum Biochemical Parameters | |||
| urea (mmol/L) | 5.2 ± 1.4 | 5.5 ± 1.3 | 2.8–8.3 |
| creatinine (µmol/L) | 78.3 ± 12.7 | 76.9 ± 12.3 | 64–104 |
| sodium (mmol/L) | 139.0 ± 12.7 | 138.9 ± 12.3 | 137–146 |
| potassium (mmol/L) | 4.3 ± 0.4 | 4.2 ± 0.2 | 3.9–5.1 |
| calcium (mmol/L) | 2.46 ± 0.08 | 2.43 ± 0.07 * | 2.14–2.53 |
| hsCRP (mg/L) | 0.84 [0.20–4.09] | 0.64 [0.12–1.70] | <5.00 |
| 24-h Urine Biochemical Parameters | |||
| 24 h urine volume (mL) | 1560 ± 714 | 1826 ± 723 * | |
| 24 h creatinine coefficient (µmol/24 h/kg) | 182 ± 45 | 191 ± 50 | 94–253 |
| 24 h urine urea (mmol/dU) | 333 ± 122 | 364 ± 140 | 200–350 |
| 24 h urine protein (mg/dU) | 88 [23–298] | 108 [27–183] | <150 |
| 24 h urine albumin (mg/dU) | 9.83 ± 10.88 | 9.17 ± 8.18 | <30.0 |
| 24 h sodium (mmol/dU) | 118 ± 42 | 267 ± 82 * | 40.0–220.0 |
| 24 h potassium (mmol/dU) | 50.4 [11.5–121.2] | 58.7 [22.7–132.6] | 25.0–125.0 |
| calculated salt intake (g/day) | 6.7 ± 2.6 | 15.6 ± 4.8 * | |
| Parameter | LS | HS |
|---|---|---|
| N (W/M) | 46 (24/22) | |
| Hemodynamic Parameters | ||
| Systolic ABP (mmHg) | 115 ± 10 | 115 ± 12 |
| Diastolic ABP (mmHg) | 74 ± 8 | 72 ± 10 |
| Mean ABP (mmHg) | 87 ± 7 | 86 ± 9 |
| Heart Rate (beats/min) | 74 ± 13 | 73 ± 9 |
| Stroke Volume (mL) | 96.7 ± 26.7 | 93.0 ± 17.0 |
| Cardiac Output (L/min) | 6.96 ± 1.30 | 6.72 ± 1.12 |
| Cardiac Index (L/min/m2) | 3.86 ± 0.54 | 3.71 ± 0.48 |
| Systemic Vascular Resistance Index (dyn.s.cm−5 m2) | 1639 ± 297 | 1754 ± 318 |
| Total Arterial Compliance Index (mL/m2/mmHg) | 1.087 ± 0.269 | 1.056 ± 0.193 |
| Parameter | LS | HS |
|---|---|---|
| N (W/M) | 46 (24/22) | |
| Nitric Oxide Serum Level | ||
| NO (μmol/L) | 39.5 ± 8.3 | 39.5 ± 11.0 |
| VEGF-C Serum Level | ||
| VEGF-C (ng/mL) | 2.93 ± 0.90 | 2.78 ± 0.69 |
| 24-h Sodium Excretion (mmol/dU) | NO Serum Concentration (μmol/L) | ||
|---|---|---|---|
| LTH (flow increase) | R | −0.254 | 0.555 |
| p | 0.050 * | 0.017 * | |
| FMD (% of dilation) | R | −0.322 | −0.268 |
| p | 0.015 * | 0.278 | |
| eNOS (IU/mL) | R | 0.071 | −0.128 |
| p | 0.664 | 0.648 | |
| nNOS (IU/mL) | R | −0.483 | −0.166 |
| p | 0.002 * | 0.562 | |
| iNOS (IU/mL) | R | 0.199 | 0.267 |
| p | 0.224 | 0.348 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tolj, I.; Stupin, A.; Drenjančević, I.; Šušnjara, P.; Perić, L.; Stupin, M. The Role of Nitric Oxide in the Micro- and Macrovascular Response to a 7-Day High-Salt Diet in Healthy Individuals. Int. J. Mol. Sci. 2023, 24, 7157. https://doi.org/10.3390/ijms24087157
Tolj I, Stupin A, Drenjančević I, Šušnjara P, Perić L, Stupin M. The Role of Nitric Oxide in the Micro- and Macrovascular Response to a 7-Day High-Salt Diet in Healthy Individuals. International Journal of Molecular Sciences. 2023; 24(8):7157. https://doi.org/10.3390/ijms24087157
Chicago/Turabian StyleTolj, Ivana, Ana Stupin, Ines Drenjančević, Petar Šušnjara, Leon Perić, and Marko Stupin. 2023. "The Role of Nitric Oxide in the Micro- and Macrovascular Response to a 7-Day High-Salt Diet in Healthy Individuals" International Journal of Molecular Sciences 24, no. 8: 7157. https://doi.org/10.3390/ijms24087157
APA StyleTolj, I., Stupin, A., Drenjančević, I., Šušnjara, P., Perić, L., & Stupin, M. (2023). The Role of Nitric Oxide in the Micro- and Macrovascular Response to a 7-Day High-Salt Diet in Healthy Individuals. International Journal of Molecular Sciences, 24(8), 7157. https://doi.org/10.3390/ijms24087157