
Serum Proteomic Profiles Reflect the Stages of Myxomatous Mitral Valve Disease in Dogs

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Patients’ Demographics and Clinical Findings
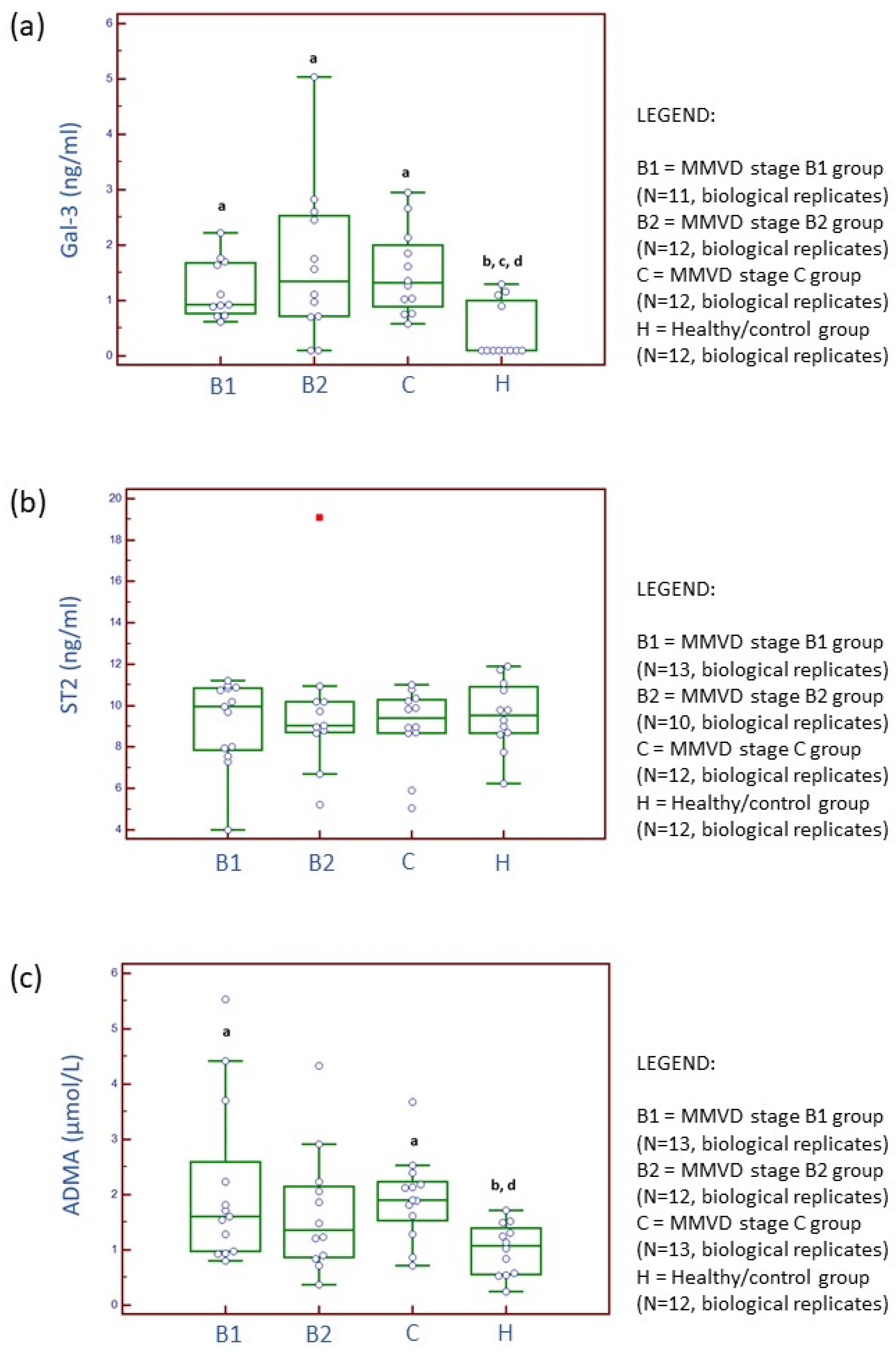
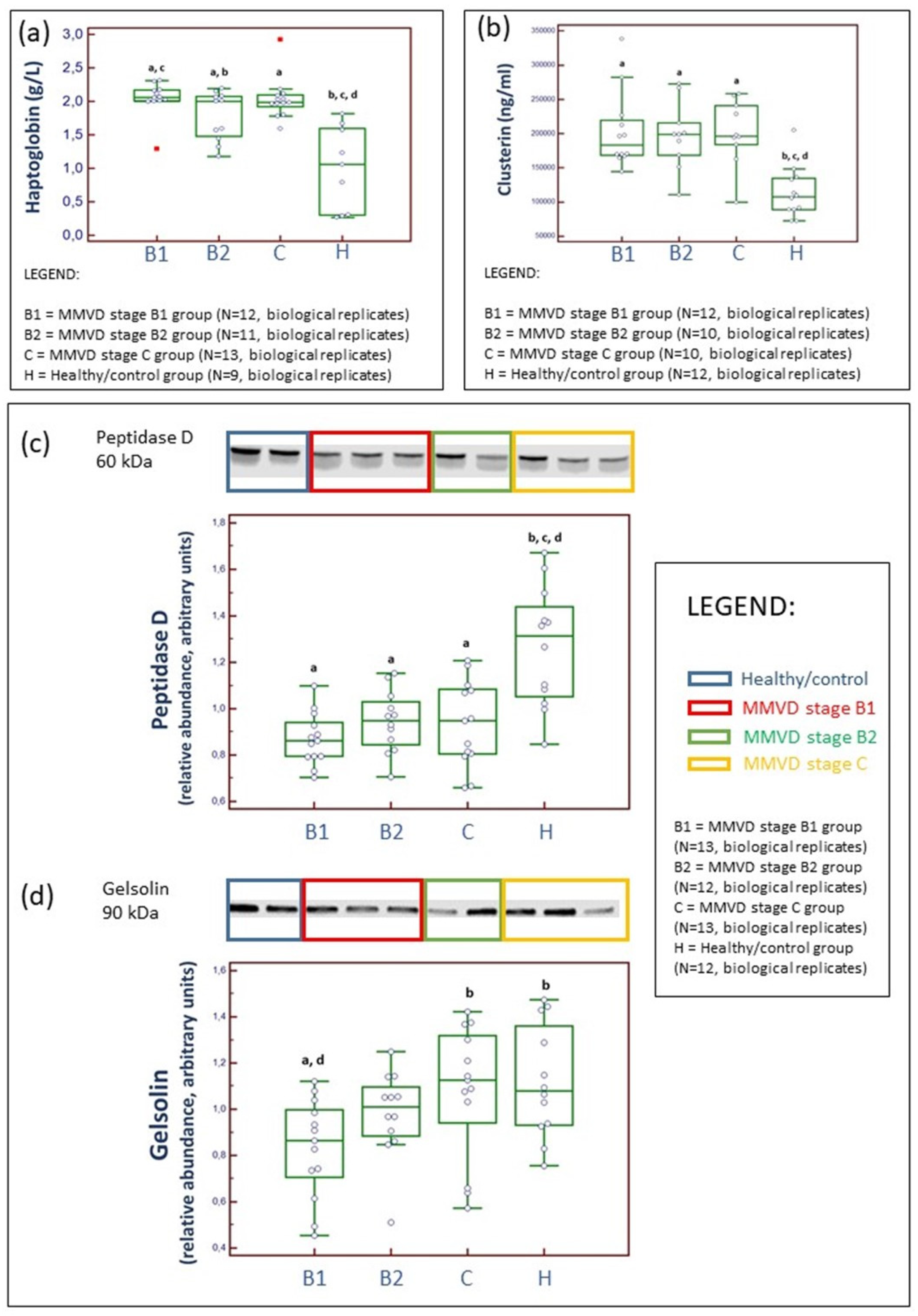
2.2. Serum and Candidate Cardiac Biomarker Analyses
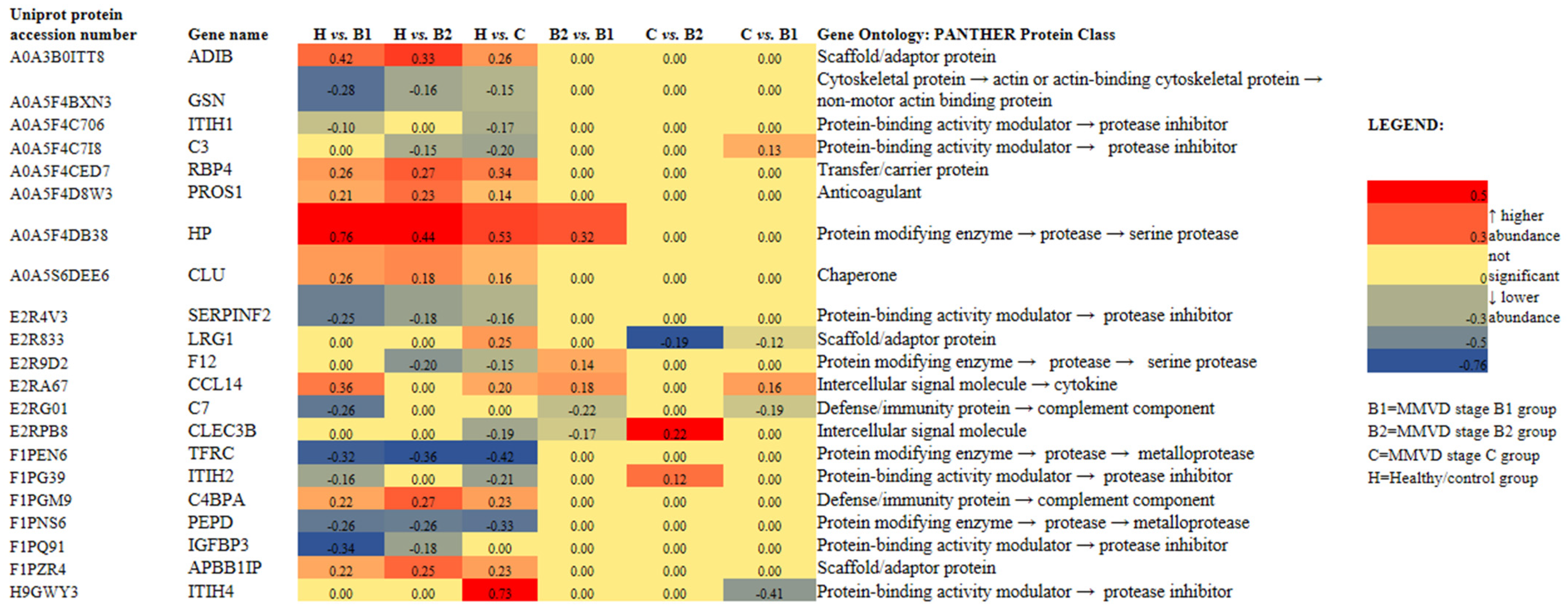
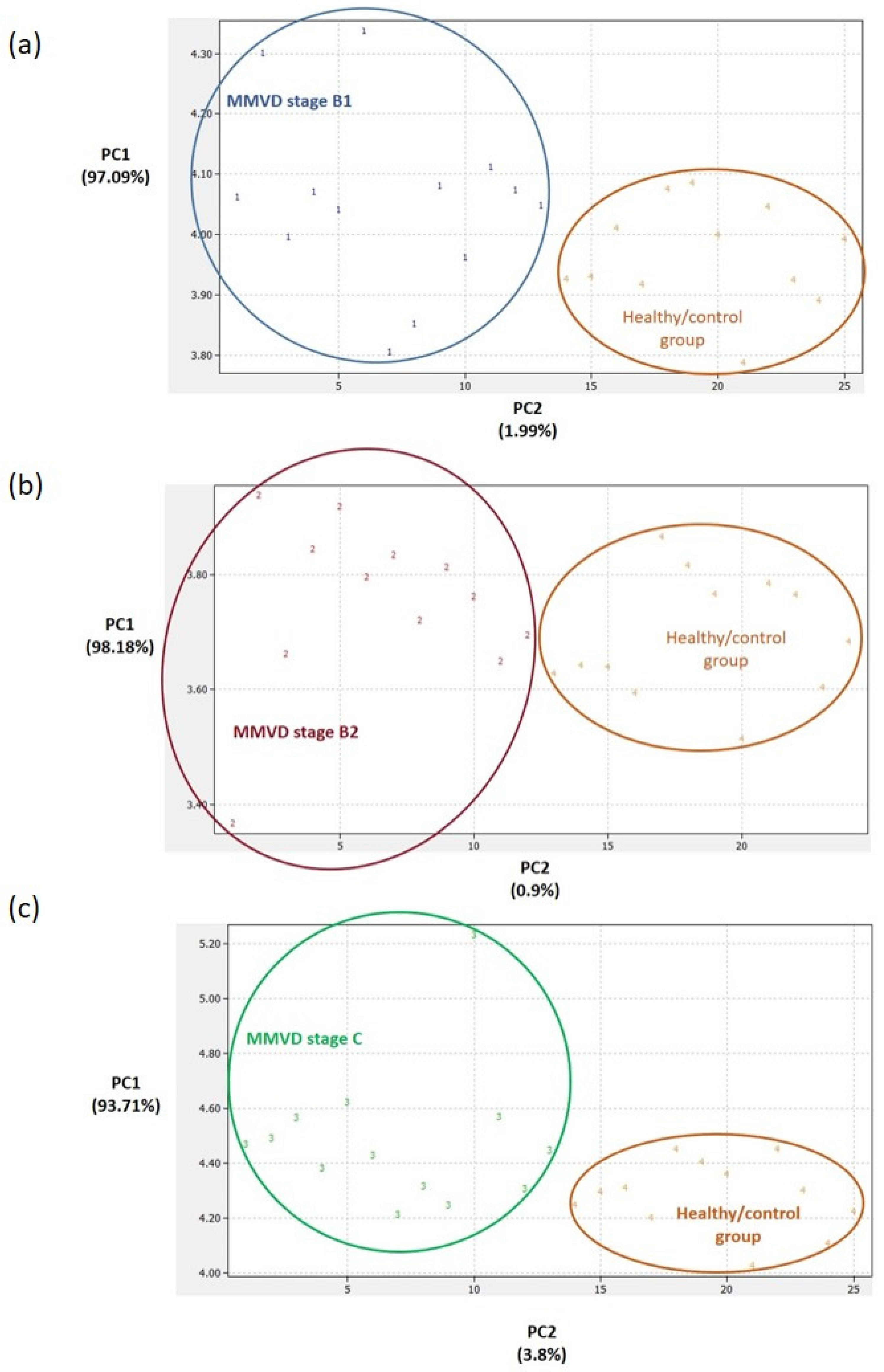
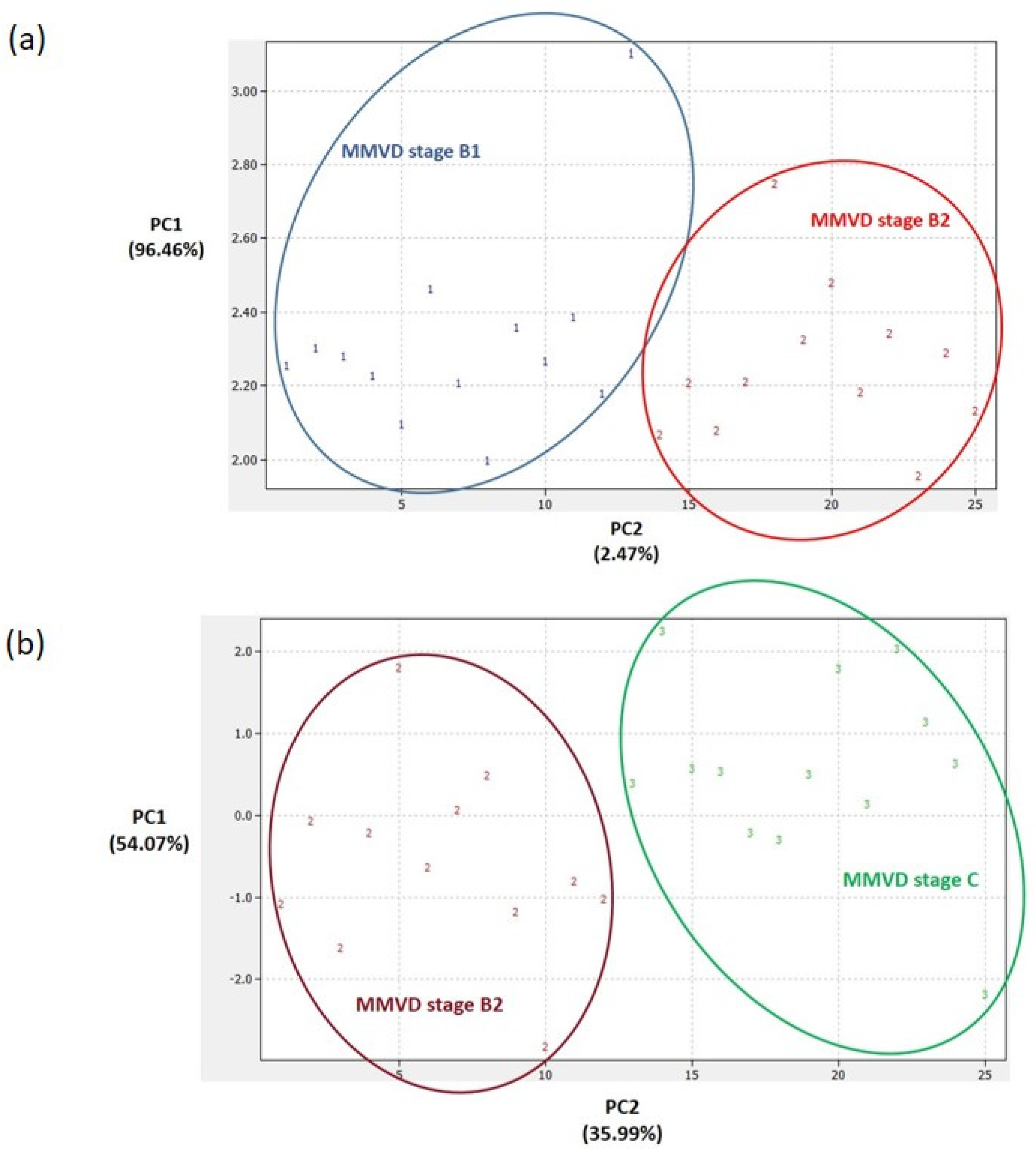
2.3. Proteomics Results
2.4. Analytical Validation of Proteomics Results
3. Discussion
3.1. Serum Parameters and Candidate Cardiac Biomarker Analysis
3.2. Proteomic Results
3.2.1. C-C Motif Chemokine (CCL14) Protein-Cytokine
3.2.2. Haptoglobin (HP), an Acute Phase Protein
3.2.3. Peptidase D (PEPD), a Member of the Matrixin Protein Family
3.2.4. Clusterine (CLU), a Chaperone Protein
3.2.5. Gelsolin (GSN), a Non-Motor Actin-Binding Protein
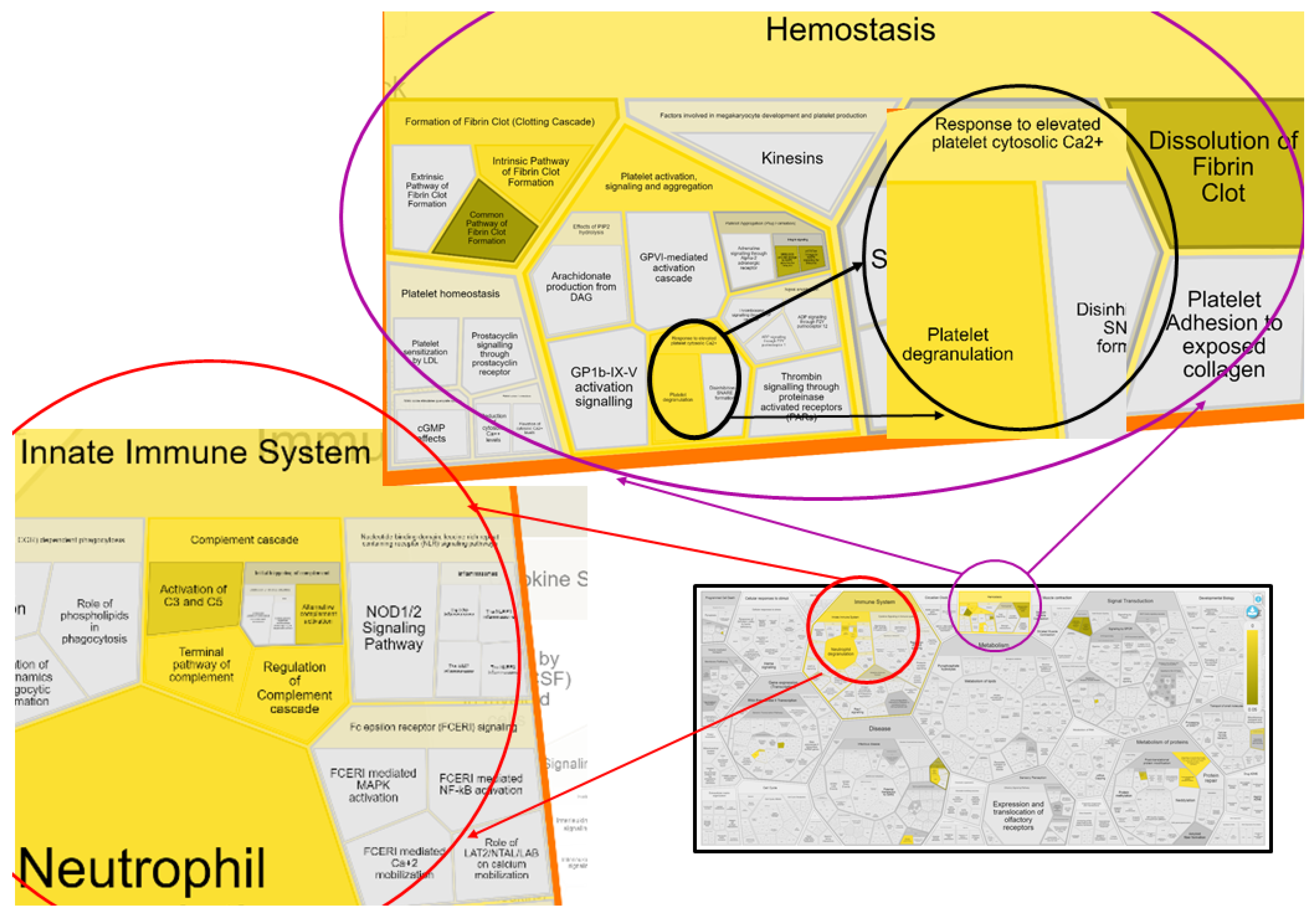
3.2.6. Innate Immune System: Platelets and Complement Cascade
3.2.7. Insulin-like Growth Factor Binding Protein 3 (IGFBP3): Cell and Tissue Hemostasis
3.2.8. Anemia of Chronic Disease
3.2.9. Study Limitations
4. Materials and Methods
4.1. Study Approval, Health Assessment, Cardiac Imaging, and Serum Collection
4.2. Serum Biochemistry and ELISA-Based Candidate Cardiac Biomarker Analyses
4.3. TMT-Based Proteomics and Data Analysis
4.3.1. Sample Preparation and Nano-LC–MS TMT-Based Proteomics
4.3.2. MS Data Processing, Statistics, and Bioinformatics Analysis
4.4. Analytical Validation of TMT Proteomics Results
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oyama, M.A.; Elliott, C.; Loughran, K.A.; Kossar, A.P.; Castillero, E.; Levy, R.J.; Ferrari, G. Comparative pathology of human and canine myxomatous mitral valve degeneration: 5HT and TGF-β mechanisms. Cardiovasc. Pathol. 2020, 46, 107196. [Google Scholar] [CrossRef] [PubMed]
- Keene, B.W.; Atkins, C.E.; Bonagura, J.D.; Fox, P.R.; Häggström, J.; Fuentes, V.L.; Oyama, M.A.; Rush, J.E.; Stepien, R.; Uechi, M. ACVIM consensus guidelines for the diagnosis and treatment of myxomatous mitral valve disease in dogs. J. Vet. Intern. Med. 2019, 33, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Atkins, C.; Bonagura, J.; Ettinger, S.; Fox, P.; Gordon, S.; Haggstrom, J.; Hamlin, R.; Keene, B.; Luis-Fuentes, V.; Stepien, R. Guidelines for the Diagnosis and Treatment of Canine Chronic Valvular Heart Disease. J. Vet. Intern. Med. 2009, 23, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Larouche-Lebel, É.; Loughran, K.A.; Huh, T.P.; Suchodolski, J.S.; Oyama, M.A. Metabolomics Analysis Reveals Deranged Energy Metabolism and Amino Acid Metabolic Reprogramming in Dogs with Myxomatous Mitral Valve Disease. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2021, 10, 18923. [Google Scholar] [CrossRef]
- Pedersen, H.D.; Häggström, J. Mitral valve prolapse in the dog: A model of mitral valve prolapse in man. Cardiovasc. Res. 2000, 47, 234–243. [Google Scholar] [CrossRef]
- Lacerda, C.M.R.; Disatian, S.; Orton, E.C. Differential protein expression between normal, early-stage, and late-stage myxomatous mitral valves from dogs. Proteom. Clin. Appl. 2009, 3, 1422–1429. [Google Scholar] [CrossRef]
- de Oliveira Martins, C.; Santos, K.S.; Ferreira, F.M.; Teixeira, P.C.; Pomerantzeff, P.M.A.; Brandão, C.M.A.; Sampaio, R.O.; Spina, G.S.; Kalil, J.; Guilherme, L.; et al. Distinct mitral valve proteomic profiles in rheumatic heart disease and myxomatous degeneration. Clin. Med. Insights Cardiol. 2014, 8, 79–86. [Google Scholar] [CrossRef]
- Locatelli, C.; Piras, C.; Riscazzi, G.; Alloggio, I.; Spalla, I.; Soggiu, A.; Greco, V.; Bonizzi, L.; Roncada, P. Serum proteomic profiles in CKCS with Mitral valve disease. BMC Vet. Res. 2017, 13, 331. [Google Scholar] [CrossRef]
- Kuleš, J.; Bilić, P.; Horvatić, A.; Kovačević, A.; Guillemin, N.; Ljubić, B.B.; Galan, A.; Jović, I.; Torti, M. Serum proteome profiling in canine chronic valve disease using a TMT-based quantitative proteomics approach. J. Proteom. 2020, 223, 103825. [Google Scholar] [CrossRef]
- Levent, P.; Kocaturk, M.; Akgun, E.; Saril, A.; Cevik, O.; Baykal, A.T.; Tanaka, R.; Ceron, J.J.; Yilmaz, Z. Platelet proteome changes in dogs with congestive heart failure. BMC Vet. Res. 2020, 16, 466. [Google Scholar] [CrossRef]
- Saril, A.; Kocaturk, M.; Shimada, K.; Uemura, A.; Akgün, E.; Levent, P.; Baykal, A.T.; Prieto, A.M.; Agudelo, C.F.; Tanaka, R.; et al. Serum Proteomic Changes in Dogs with Different Stages of Chronic Heart Failure. Animals 2022, 12, 490. [Google Scholar] [CrossRef] [PubMed]
- Wilshaw, J.; Boswood, A.; Chang, Y.M.; Sands, C.J.; Camuzeaux, S.; Lewis, M.R.; Xia, D.; Connolly, D.J. Evidence of altered fatty acid metabolism in dogs with naturally occurring valvular heart disease and congestive heart failure. Metabolomics 2022, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Freeman, L.M.; Rush, J.E.; Huggins, G.S.; Kennedy, A.D.; Labuda, J.A.; Laflamme, D.P.; Hannah, S.S. Veterinary Medicine and Multi-Omics Research for Future Nutrition Targets: Metabolomics and Transcriptomics of the Common Degenerative Mitral Valve Disease in Dogs. OMICS 2015, 19, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Mallat, Z.; Weyand, C. Immune and inflammatory mechanisms mediate cardiovascular diseases from head to toe. Cardiovasc. Res. 2021, 117, 2503. [Google Scholar] [CrossRef] [PubMed]
- Deroyer, C.; Magne, J.; Moonen, M.; Le Goff, C.; Dupont, L.; Hulin, A.; Radermecker, M.; Colige, A.; Cavalier, E.; Kolh, P.; et al. New biomarkers for primary mitral regurgitation. Clin. Proteom. 2015, 12, 25. [Google Scholar] [CrossRef]
- Hara, A.; Niwa, M.; Kanayama, T.; Noguchi, K.; Niwa, A.; Matsuo, M.; Kuroda, T.; Hatano, Y.; Okada, H.; Tomita, H. Galectin-3: A Potential Prognostic and Diagnostic Marker for Heart Disease and Detection of Early Stage Pathology. Biomolecules 2020, 10, 1277. [Google Scholar] [CrossRef]
- Klein, S.; Nolte, I.; Granados-Soler, J.L.; Lietz, P.; Sehn, M.; Raue, J.F.; Rohn, K.; Packeiser, E.M.; Bach, J.P. Evaluation of new and old biomarkers in dogs with degenerative mitral valve disease. BMC Vet. Res. 2022, 18, 256. [Google Scholar] [CrossRef]
- De Boer, R.A.; Voors, A.A.; Muntendam, P.; Van Gilst, W.H.; Van Veldhuisen, D.J. Galectin-3: A novel mediator of heart failure development and progression. Eur. J. Heart Fail. 2009, 11, 811–817. [Google Scholar] [CrossRef]
- Ho, J.E.; Liu, C.; Lyass, A.; Courchesne, P.; Pencina, M.J.; Vasan, R.S.; Larson, M.G.; Levy, D. Galectin-3, a Marker of Cardiac Fibrosis, Predicts Incident Heart Failure in the Community. J. Am. Coll. Cardiol. 2012, 60, 1249–1256. [Google Scholar] [CrossRef]
- Lok, D.J.A.; Van Der Meer, P.; De La Porte, P.W.B.A.; Lipsic, E.; Van Wijngaarden, J.; Hillege, H.L.; Van Veldhuisen, D.J. Prognostic value of galectin-3, a novel marker of fibrosis, in patients with chronic heart failure: Data from the DEAL-HF study. Clin. Res. Cardiol. 2010, 99, 323–328. [Google Scholar] [CrossRef]
- Sakarin, S.; Rungsipipat, A.; Surachetpong, S.D. Galectin-3 in cardiac muscle and circulation of dogs with degenerative mitral valve disease. J. Vet. Cardiol. 2016, 18, 34–46. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.A.; Lok, D.J.A.; Jaarsma, T.; Van Der Meer, P.; Voors, A.A.; Hillege, H.L.; Van Veldhuisen, D.J. Predictive value of plasma galectin-3 levels in heart failure with reduced and preserved ejection fraction. Ann. Med. 2011, 43, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Andrés, N.; Rossignol, P.; Iraqi, W.; Fay, R.; Nuée, J.; Ghio, S.; Cleland, J.G.F.; Zannad, F.; Lacolley, P. Association of galectin-3 and fibrosis markers with long-term cardiovascular outcomes in patients with heart failure, left ventricular dysfunction, and dyssynchrony: Insights from the CARE-HF (Cardiac Resynchronization in Heart Failure) trial. Eur. J. Heart Fail. 2012, 14, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.W.; Kang, M.H.; Ro WBin Song, D.W.; Park, H.M. Circulating Galectin-3 Evaluation in Dogs with Cardiac and Non-cardiac Diseases. Front. Vet. Sci. 2021, 8, 741210. [Google Scholar] [CrossRef]
- Sabbah, H.N.; Singh-Gupta, V.; Ramesh, C.G. Abstract 12472: Inhibition of Galectin-3 Reverses Reactive Interstitial Fibrosis in Left Ventricular Myocardium of Dogs with Chronic Heart Failure. Circulation 2018, 138, A12472. [Google Scholar]
- Patric, B.; Camille, A.; Mauro, I.; Angelika, H.-L.; Tobias, B.; Christian, M.; Alan, M.; Frank, R. Soluble ST2—A new biomarker in heart failure. Cardiovasc. Med. 2019, 22, w02008. [Google Scholar]
- Garcia-Pena, A.; Ibarrola, J.; Navarro, A.; Sadaba, A.; Tiraplegui, C.; Garaikoetxea, M.; Arrieta, V.; Matilla, L.; Fernández-Celis, A. Activation of the Interleukin-33/ST2 Pathway Exerts Deleterious Effects in Myxomatous Mitral Valve Disease. Int. J. Mol. Sci. 2021, 22, 2310. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, J.S.; Seo, K.W.; Song, K.H. Evaluation of ST2 and NT-proBNP as Cardiac Biomarkers in Dogs with Chronic Mitral Valve Disease. J. Vet. Clin. 2018, 35, 35–38. [Google Scholar] [CrossRef]
- Pedersen, L.G.; Tarnow, I.; Olsen, L.H.; Teerlink, T.; Pedersen, H.D. Body size, but neither age nor asymptomatic mitral regurgitation, influences plasma concentrations of dimethylarginines in dogs. Res. Vet. Sci. 2006, 80, 336–342. [Google Scholar] [CrossRef]
- Valente, C.; Guglielmini, C.; Baron Toaldo, M.; Romito, G.; Artusi, C.; Brugnolo, L.; Contiero, B.; Poser, H. Plasmatic Dimethylarginines in Dogs with Myxomatous Mitral Valve Disease. Front. Vet. Sci. 2021, 8, 738898. [Google Scholar] [CrossRef]
- Ci’Han, H.; Tural, M. Assessment of asymmetric dimethyl arginine, cardiac troponin I, thyroxine, cholesterol, and triglyceride levels in obese dogs and dogs with normal body condition. Turk. J. Vet. Anim. Sci. 2019, 43, 271–275. [Google Scholar] [CrossRef]
- Gori, E.; Pierini, A.; Lippi, I.; Meucci, V.; Perondi, F.; Marchetti, V. Evaluation of asymmetric dimethylarginine as an inflammatory and prognostic marker in dogs with acute pancreatitis. J. Vet. Intern. Med. 2020, 34, 1144. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.C.; Langford-Smith, A.W.W.; Birchenough, H.L.; Jowitt, T.A.; Kielty, C.M.; Enghild, J.J.; Baldock, C.; Milner, C.M. Inter-α-inhibitor heavy chain-1 has an integrin-like 3D structure mediating immune regulatory activities and matrix stabilization during ovulation. J Biol Chem. 2020, 295, 5278–5291. [Google Scholar] [CrossRef] [PubMed]
- Aupperle, H.; Disatian, S. Pathology, protein expression and signaling in myxomatous mitral valve degeneration: Comparison of dogs and humans. J. Vet. Cardiol. 2012, 14, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Piantedosi, D.; Musco, N.; Palatucci, A.T.; Carriero, F.; Rubino, V.; Pizzo, F.; Nasir, S.; Molinaro, G.; Ruggiero, G.; Terrazzano, G.; et al. Pro-Inflammatory and Immunological Profile of Dogs with Myxomatous Mitral Valve Disease. Vet. Sci. 2022, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Yutzey, K.E. Therapeutic CCR2 Blockade Prevents Inflammation and Alleviates Myxomatous Valve Disease in Marfan Syndrome. JACC Basic Transl. Sci. 2022, 7, 1143–1157. [Google Scholar] [CrossRef]
- Sridhar, S.; Pham, D.H.; Gee, T.W.; Hua, J.; Butcher, J.T. Monocytes and macrophages in heart valves: Uninvited guests or critical performers? Curr. Opin. Biomed. Eng. 2018, 5, 82–89. [Google Scholar] [CrossRef]
- Theilgaard-Mönch, K.; Jacobsen, L.C.; Nielsen, M.J.; Rasmussen, T.; Udby, L.; Gharib, M.; Arkwright, P.D.; Gombart, A.F.; Calafat, J.; Moestrup, S.K.; et al. Haptoglobin is synthesized during granulocyte differentiation, stored in specific granules, and released by neutrophils in response to activation. Blood 2006, 108, 353–361. [Google Scholar] [CrossRef]
- Druhan, L.J.; Lance, A.; Li, S.; Price, A.E.; Emerson, J.T.; Baxter, S.A.; Gerber, J.M.; Avalos, B.R. Leucine Rich α-2 Glycoprotein: A Novel Neutrophil Granule Protein and Modulator of Myelopoiesis. PLoS ONE 2017, 12, e0170261. [Google Scholar] [CrossRef]
- Eni-Aganga, I.; Lanaghan, Z.M.; Balasubramaniam, M.; Dash, C.; Pandhare, J. PROLIDASE: A Review from Discovery to its Role in Health and Disease. Front. Mol. Biosci. 2021, 8, 849. [Google Scholar] [CrossRef]
- Turkieh, A.; Fertin, M.; Bouvet, M.; Mulder, P.; Drobecq, H.; Lemesle, G.; Lamblin, N.; De Groote, P.; Porouchani, S.; Chwastyniak, M.; et al. Expression and Implication of Clusterin in Left Ventricular Remodeling After Myocardial Infarction. Circ. Heart Fail. 2018, 11, e004838. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Gonos, E.S. Clusterin/Apolipoprotein J in human aging and cancer. Int. J. Biochem. Cell Biol. 2002, 34, 1430–1448. [Google Scholar] [CrossRef] [PubMed]
- Yuste-Checa, P.; Bracher, A.; Hartl, F.U. The chaperone Clusterin in neurodegeneration−friend or foe? BioEssays 2022, 44, 2100287. [Google Scholar] [CrossRef]
- Rabkin, E.; Aikawa, M.; Stone, J.R.; Fukumoto, Y.; Libby, P.; Schoen, F.J. Activated interstitial myofibroblasts express catabolic enzymes and mediate matrix remodeling in myxomatous heart valves. Circulation 2001, 104, 2525–2532. [Google Scholar] [CrossRef] [PubMed]
- Darke, P.G. Valvular incompetence in cavalier King Charles spaniels. Vet. Rec. 1987, 120, 365–366. [Google Scholar] [CrossRef]
- Dye, B.K.; Butler, C.; Lincoln, J. Smooth Muscle α-Actin Expression in Mitral Valve Interstitial Cells is Important for Mediating Extracellular Matrix Remodeling. J. Cardiovasc. Dev. Dis. 2020, 7, 32. [Google Scholar] [CrossRef]
- Coleman, J.R.; Moore, E.E.; Freeman, K.; Grubinger, N.D.; Hennig, G.W.; Cohen, M.J.; Samuels, J.M.; Hansen, K. Actin is associated with tissue injury in trauma patients and produces a hypercoagulable profile in vitro. J. Trauma Acute Care Surg. 2020, 89, 87. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, H.; Jiang, Y.; Xue, M.; Guo, C.; Shi, D.; Chen, K. Correlation between Platelet Gelsolin and Platelet Activation Level in Acute Myocardial Infarction Rats and Intervention Effect of Effective Components of Chuanxiong Rhizome and Red Peony Root. Evid. Based Complement. Alternat. Med. 2013, 2013, 985746. [Google Scholar] [CrossRef]
- McDonald, K.; Glezeva, N.; Collier, P.; O’Reilly, J.; O’Connell, E.; Tea, I.; Russell-Hallinan, A.; Tonry, C.; Pennington, S. Tetranectin, a potential novel diagnostic biomarker of heart failure, is expressed within the myocardium and associates with cardiac fibrosis. Sci. Rep. 2020, 10, 7507. [Google Scholar] [CrossRef]
- Tarnow, I.; Falk, T.; Tidholm, A.; Martinussen, T.; Jensen, A.L.; Olsen, L.H.; Pedersen, H.D.; Kristensen, A.T. Hemostatic Biomarkers in Dogs with Chronic Congestive Heart Failure. J. Vet. Intern. Med. 2007, 21, 451–457. [Google Scholar] [CrossRef]
- Prihirunkit, K.; Sastravaha, A.; Lekcharoensuk, C.; Chanloinapha, P. Hemostatic Markers in Congestive Heart Failure Dogs with Mitral Valve Disease. J. Vet. Med. 2014, 2014, 589873. [Google Scholar] [CrossRef] [PubMed]
- Caivano, D.; Marchesi, M.C.; Birettoni, F.; Lepri, E.; Porciello, F. Left Atrial Mural Thrombosis and Hemopericardium in a Dog with Myxomatous Mitral Valve Disease. Vet. Sci. 2021, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Burchell, R.K.; Schoeman, J.P. Advances in the understanding of the pathogenesis, progression and diagnosis of myxomatous mitral valve disease in dogs. J. S. Afr. Vet. Assoc. 2014, 85, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Steinle, J.J. IGFBP-3 inhibits TNF-α production and TNFR-2 signaling to protect against Retinal Endothelial Cell Apoptosis. Microvasc. Res. 2014, 95, 76–81. [Google Scholar] [CrossRef]
- Varma Shrivastav, S.; Bhardwaj, A.; Pathak, K.A.; Shrivastav, A. Insulin-Like Growth Factor Binding Protein-3 (IGFBP-3): Unraveling the Role in Mediating IGF-Independent Effects within the Cell. Front. Cell Dev. Biol. 2020, 8, 286. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.P.; Ebihara, Y.; LaRue, A.C.; Fleming, P.A.; McQuinn, T.C.; Masuya, M.; Minamiguchi, H.; Markwald, R.R.; Ogawa, M.; Drake, C.J. An In Vivo Analysis of Hematopoietic Stem Cell Potential. Circ. Res. 2006, 98, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Chikazawa, S.; Dunning, M.D. A review of anaemia of inflammatory disease in dogs and cats. J. Small Anim. Pract. 2016, 57, 348–353. [Google Scholar] [CrossRef]
- Thomas, W.P.; Gaber, C.E.; Jacobs, G.J.; Kaplan, P.M.; Lombard, C.W.; Vet, M.; Moise, N.S.; Moses, B.L. Recommendations for standards in transthoracic two-dimensional echocardiography in the dog and cat. Echocardiography Committee of the Specialty of Cardiology, American College of Veterinary Internal Medicine. J. Vet. Intern. Med. 1993, 7, 247–252. [Google Scholar] [CrossRef]
- Kvart, C.; Häggström, J.; Pedersen, H.D.; Hansson, K.; Eriksson, A.; Järvinen, A.K.; Tidholm, A.; Bsenko, K.; Ahlgren, E.; Ilves, M.; et al. Efficacy of Enalapril for Prevention of Congestive Heart Failure in Dogs with Myxomatous Valve Disease and Asymptomatic Mitral Regurgitation. J. Vet. Intern. Med. 2002, 16, 80–88. [Google Scholar] [CrossRef]
- Atkins, C.E.; Keene, B.W.; Brown, W.A.; Coats, J.R.; Crawford, M.A.; DeFrancesco, T.C.; Edwards, N.J.; Fox, P.R.; Lehmkuhl, L.B.; Luethy, M.W.; et al. Results of the veterinary enalapril trial to prove reduction in onset of heart failure in dogs chronically treated with enalapril alone for compensated, naturally occurring mitral valve insufficiency. J. Am. Vet. Med. Assoc 2007, 231, 1061–1069. [Google Scholar] [CrossRef]
- Wess, G.; Kresken, J.G.; Wendt, R.; Gaugele, J.; Killich, M.; Keller, L.; Simak, J.; Holler, P.; Bauer, A. Efficacy of adding ramipril (VAsotop) to the combination of furosemide (Lasix) and pimobendan (VEtmedin) in dogs with mitral valve degeneration: The VALVE trial. J. Vet. Intern. Med. 2020, 34, 2232–2241. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Schäffer, A.A.; Aravind, L.; Madden, T.L.; Shavirin, S.; Spouge, J.L.; Wolf, Y.I.; Koonin, E.V.; Altschul, S.F. Improving the accuracy of PSI-BLAST protein database searches with composition-based statistics and other refinements. Nucleic Acids Res. 2001, 29, 2994–3005. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F. The Reactome pathway Knowledgebase. Nucleic Acids Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef]
- Eckersall, P.D.; Duthie, S.; Safi, S.; Moffatt, D.; Horadagoda, N.U.; Doyle, S.; Parton, R.; Bennett, D.; Fitzpatrick, J.L. An automated biochemical assay for haptoglobin: Prevention of interference from albumin. Comp. Haematol. Int. 1999, 9, 117–124. [Google Scholar] [CrossRef]
- Brady, N.; O’Reilly, E.L.; McComb, C.; Macrae, A.I.; Eckersall, P.D. An immunoturbidimetric assay for bovine haptoglobin. Comp. Clin. Path. 2019, 28, 21–27. [Google Scholar] [CrossRef]
- Kjelgaard-Hansen, M.; Jacobsen, S. Assay Validation and Diagnostic Applications of Major Acute-phase Protein Testing in Companion Animals. Clin. Lab. Med. 2011, 31, 51–70. [Google Scholar] [CrossRef]
- Goggs, R.; Myers, M.; De Rosa, S.; Zager, E.; Fletcher, D.J. Chloride:Sodium Ratio May Accurately Predict Corrected Chloride Disorders and the Presence of Unmeasured Anions in Dogs and Cats. Front. Vet. Sci. 2017, 4, 122. [Google Scholar] [CrossRef] [PubMed]
- Ortega, O.; Rodriguez, I.; Hinostroza, J.; Laso, N.; Callejas, R.; Gallar, P.; Mon, C.; Herrero, J.C.; Ortiz, M.; Oliet, A.; et al. Serum Alkaline Phosphatase Levels and Left Ventricular Diastolic Dysfunction in Patients with Advanced Chronic Kidney Disease. Nephron Extra 2011, 1, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Morningstar, J.E.; Gensemer, C.; Moore, R.; Fulmer, D.; Beck, T.C.; Wang, C.; Moore, K.; Guo, L.; Sieg, F.; Nagata, Y.; et al. Mitral Valve Prolapse Induces Regionalized Myocardial Fibrosis. J. Am. Heart Assoc. 2021, 10, e022332. [Google Scholar] [CrossRef] [PubMed]
- Urfer, S.R.; Kaeberlein, T.L.; Mailheau, S.; Bergman, P.J.; Creevy, K.E.; Promislow, D.E.L.; Kaeberlein, M. Asymptomatic heart valve dysfunction in healthy middle-aged companion dogs and its implications for cardiac aging. GeroScience 2017, 39, 43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, M.-J.; Yang, H.-M.; Ma, C.-Y.; Jia, P.-Y.; Jia, D.-L.; Hou, A.-J. Association between increased serum alkaline phosphatase and the coronary slow flow phenomenon. BMC Cardiovasc Disord. 2018, 18, 138. [Google Scholar] [CrossRef]
- Zimmerman, K.L.; Panciera, D.L.; Hoeschele, I.; Monroe, W.E.; Todd, S.M.; Werre, S.R.; Leroith, T.; Fecteau, K.; Lake, B.B. Adrenocortical Challenge Response and Genomic Analyses in Scottish Terriers With Increased Alkaline Phosphate Activity. Front. Vet. Sci. 2018, 5, 231. [Google Scholar] [CrossRef]
- Aktas, M.; Auguste, D.; Lefebvre, H.P.; Toutain, P.L.; Braun, J.P. Creatine kinase in the dog: A review. Vet. Res. Commun. 1993, 17, 353–369. [Google Scholar] [CrossRef]
- Bakirel, U.; Gunes, S. Value of cardiac markers in dogs with chronic mitral valve disease. Acta Vet. 2009, 59, 223–229. [Google Scholar]
- Adin, D.; Atkins, C.; Londoño, L.; Del Nero, B. Correction of serum chloride concentration in dogs with congestive heart failure. J. Vet. Intern. Med. 2020, 35, 51–57. [Google Scholar] [CrossRef]
- Adin, D.; Kurtz, K.; Atkins, C.; Papich, M.G.; Vaden, S. Role of electrolyte concentrations and renin-angiotensin-aldosterone activation in the staging of canine heart disease. J. Vet. Intern. Med. 2019, 34, 53–64. [Google Scholar] [CrossRef]
- Banner, N.R.; Bonser, R.S.; Clark, A.L.; Clark, S.; Cowburn, P.J.; Gardner, R.S.; Kalra, P.R.; McDonagh, T.; Rogers, C.A.; Swan, L.; et al. UK guidelines for referral and assessment of adults for heart transplantation. Heart 2011, 97, 1520–1527. [Google Scholar] [CrossRef] [PubMed]
- Christopoulou, E.C.; Filippatos, T.D.; Megapanou, E.; Elisaf, M.S.; Liamis, G. Phosphate imbalance in patients with heart failure. Heart Fail. Rev. 2017, 22, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, J.; Kestenbaum, B.; Chonchol, M. Phosphate and Cardiovascular Disease. Adv. Chronic Kidney Dis. 2011, 18, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Voelkl, J.; Egli-Spichtig, D.; Alesutan, I.; Wagner, C.A. Inflammation: A putative link between phosphate metabolism and cardiovascular disease. Clin. Sci. 2021, 135, 201–227. [Google Scholar] [CrossRef] [PubMed]
- Szczepankiewicz, B.; Pasławska, U.; Siwińska, N.; Plens, K.; Pasławski, R. Evaluation of the diagnostic value of the renal resistive index as a marker of the subclinical development of cardiorenal syndrome in MMVD dogs. J. Renin-Angiotensin-Aldosterone Syst. 2021, 22, 1–10. [Google Scholar] [CrossRef]
- Lan, Q.; Zheng, L.; Zhou, X.; Wu, H.; Buys, N.; Liu, Z.; Sun, J.; Fan, H. The Value of Blood Urea Nitrogen in the Prediction of Risks of Cardiovascular Disease in an Older Population. Front. Cardiovasc. Med. 2021, 8, 614117. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Healthy Dogs (Control) | MMVD Stage B1 | MMVD Stage B2 | MMVD Stage C |
|---|---|---|---|---|
| Demographics | ||||
| Number of dogs per group | 12 | 13 | 12 | 13 |
| Number of male/female dogs | 4/8 | 7/6 | 4/8 | 8/5 |
| Breeds (frequency) | Mixed breed (4) Belgian Shepherd (3) Dachshund (2) Labrador Retriever Beauceron Border collie | Maltese dog (4) Mixed breed (2) CKCS Havanese Poodle Shih-tzu Bichon frisé Miniature schnauzer Pekingese | CKCS (6) Maltese dog Poodle Beagle Mixed breed German spitz Miniature schnauzer | CKCS (5) Mixed breed (2) Yorkshire Terrier (2) Maltese dog Poodle Shih-tzu Bichon frisé |
| Age (years) | ||||
| Median (Q1–Q3) | 7 (4–10) b, c, d | 12 (7–14) a | 11 (8–13) a | 11 (9–14) a |
| Average ± StDev. Min Max Mode | 7 ± 3 3 13 3 | 11 ± 3 4 15 12 | 10 ± 3 6 16 12 | 11 ± 3 6 16 11 |
| Weight (kg) | ||||
| Median (Q1–Q3) | 15.9 (8.7–26.2) b, c, d | 7.5 (4.7–9.9) a | 10.1 (5.2–11.6) a | 7.2 (5.1–10.2) a |
| Average ± StDev. | 19.2 ± 11.3 | 7.8 ± 4.1 | 9 ± 3.7 | 7.4 ± 2.9 |
| Echocardiographic measurements | ||||
| LA/Ao | ||||
| Median (Q1–Q3) | 1.3 (1.3–1.4) b, c, d | 1.4 (1.4–1.5) a, c, d | 1.78 (1.6–1.9) a, b, d | 2.2 (2–2.6) a, b, c |
| Average ± StDev. | 1.3 ± 0.1 | 1.5 ± 0.1 | 1.8 ± 0.17 | 2.26 ± 0.37 |
| LVIDDn | ||||
| Median (Q1–Q3) | 1.6 (1.5–1.7) c, d | 1.5 (1.5–1.7) c, d | 1.8 (1.6–2.0) a, b, d | 2.3 (2.2–2.5) a, b, c |
| Average ± StDev. | 1.6 ± 5.7 | 1.5 ± 0.3 | 1.8 ± 0.2 | 2.3 ± 0.2 |
| FS% | ||||
| Median (Q1–Q3) | 31.7 (28.8–35.9) b, c, d | 41 (39–52) a | 46 (42–55) a | 47.5 (39.3–50.8) a |
| Average ± StDev. | 32.9 ± 5.7 | 44 ± 8 | 48 ± 6 | 45.4 ± 6.6 |
| MV E/A ratio | ||||
| Median (Q1–Q3) | 1.2 (1.2–1.2) b | 0.9 (0.8–1) a, c, d | 1.1 (0.9–1.4) b | 1.7 (0.9–2.5) b |
| Average ± StDev. | 1.2 ± 0.04 | 0.9 ± 0.2 | 1.2 ± 0.3 | 1.8 ± 1 |
| Pathway Name | Associated Gene Names | p-Value | FDR * |
|---|---|---|---|
| Regulation of complement cascade # | C3, C4BPA, C7, PROS1, CLU | 1.50 × 10−7 | 1.56 × 10−5 |
| Complement cascade # | C3, C4BPA, C7, PROS1, CLU | 2.94 × 10−7 | 1.56 × 10−04 |
| Platelet degranulation # | CLEC3B, PROS1, CLU, ITIH4, SERPINF2 | 4.76 × 10−6 | 1.56 × 10−04 |
| Response to elevated platelet cytosolic Ca2+ # | CLEC3B, PROS1, CLU, ITIH4, SERPINF3 | 6.02 × 10−6 | 1.56 × 10−04 |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | C3, ITIH2, IGFBP3 | 7.44 × 10−5 | 0.002 |
| Terminal pathway of complement | C7, CLU | 9.71 × 10−5 | 0.002 |
| Platelet activation, signaling, and aggregation # | CLEC3B, PROS1, CLU, ITIH4, SERPINF2 | 1.53 × 10−4 | 0.002 |
| Innate immune system # | HP, C4BPA, C7, LRG1, PROS1, C3, GSN, CLU | 3.65 × 10−4 | 0.005 |
| TP53 regulates transcription of death receptors and ligands | IGFBP3 | 4.86 × 10−4 | 0.005 |
| Post-translational protein phosphorylation | C3, ITIH2, IGFBP3 | 9.59 × 10−4 | 0.009 |
| Intrinsic pathway of fibrin clot formation | PROS1, F12 | 0.0010 | 0.009 |
| Hemostasis # | PROS1, ITIH4, F12, SERPINF2, CLEC3B, CLU | 0.0030 | 0.020 |
| Formation of a fibrin clot (clotting cascade) | PROS1, F12 | 0.0030 | 0.022 |
| Retinoid metabolism disease events | RBP4 | 0.0040 | 0.025 |
| Defective SERPING1 causes hereditary angioedema | F12 | 0.0050 | 0.037 |
| Defective factor XII causes hereditary angioedema | F12 | 0.0070 | 0.043 |
| Iron uptake and transport | TFRC | 0.0100 | 0.048 |
| TP53 regulates transcription of cell death genes | IGFBP3 | 0.0100 | 0.048 |
| Neutrophil degranulation | HP, C3, GSN, LRG1 | 0.0100 | 0.049 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rešetar Maslov, D.; Farkaš, V.; Rubić, I.; Kuleš, J.; Beletić, A.; Beer Ljubić, B.; Šmit, I.; Mrljak, V.; Torti, M. Serum Proteomic Profiles Reflect the Stages of Myxomatous Mitral Valve Disease in Dogs. Int. J. Mol. Sci. 2023, 24, 7142. https://doi.org/10.3390/ijms24087142
Rešetar Maslov D, Farkaš V, Rubić I, Kuleš J, Beletić A, Beer Ljubić B, Šmit I, Mrljak V, Torti M. Serum Proteomic Profiles Reflect the Stages of Myxomatous Mitral Valve Disease in Dogs. International Journal of Molecular Sciences. 2023; 24(8):7142. https://doi.org/10.3390/ijms24087142
Chicago/Turabian StyleRešetar Maslov, Dina, Vladimir Farkaš, Ivana Rubić, Josipa Kuleš, Anđelo Beletić, Blanka Beer Ljubić, Iva Šmit, Vladimir Mrljak, and Marin Torti. 2023. "Serum Proteomic Profiles Reflect the Stages of Myxomatous Mitral Valve Disease in Dogs" International Journal of Molecular Sciences 24, no. 8: 7142. https://doi.org/10.3390/ijms24087142
APA StyleRešetar Maslov, D., Farkaš, V., Rubić, I., Kuleš, J., Beletić, A., Beer Ljubić, B., Šmit, I., Mrljak, V., & Torti, M. (2023). Serum Proteomic Profiles Reflect the Stages of Myxomatous Mitral Valve Disease in Dogs. International Journal of Molecular Sciences, 24(8), 7142. https://doi.org/10.3390/ijms24087142