
Sex Differences in Tryptophan Metabolism: A Systematic Review Focused on Neuropsychiatric Disorders





Abstract
1. Introduction
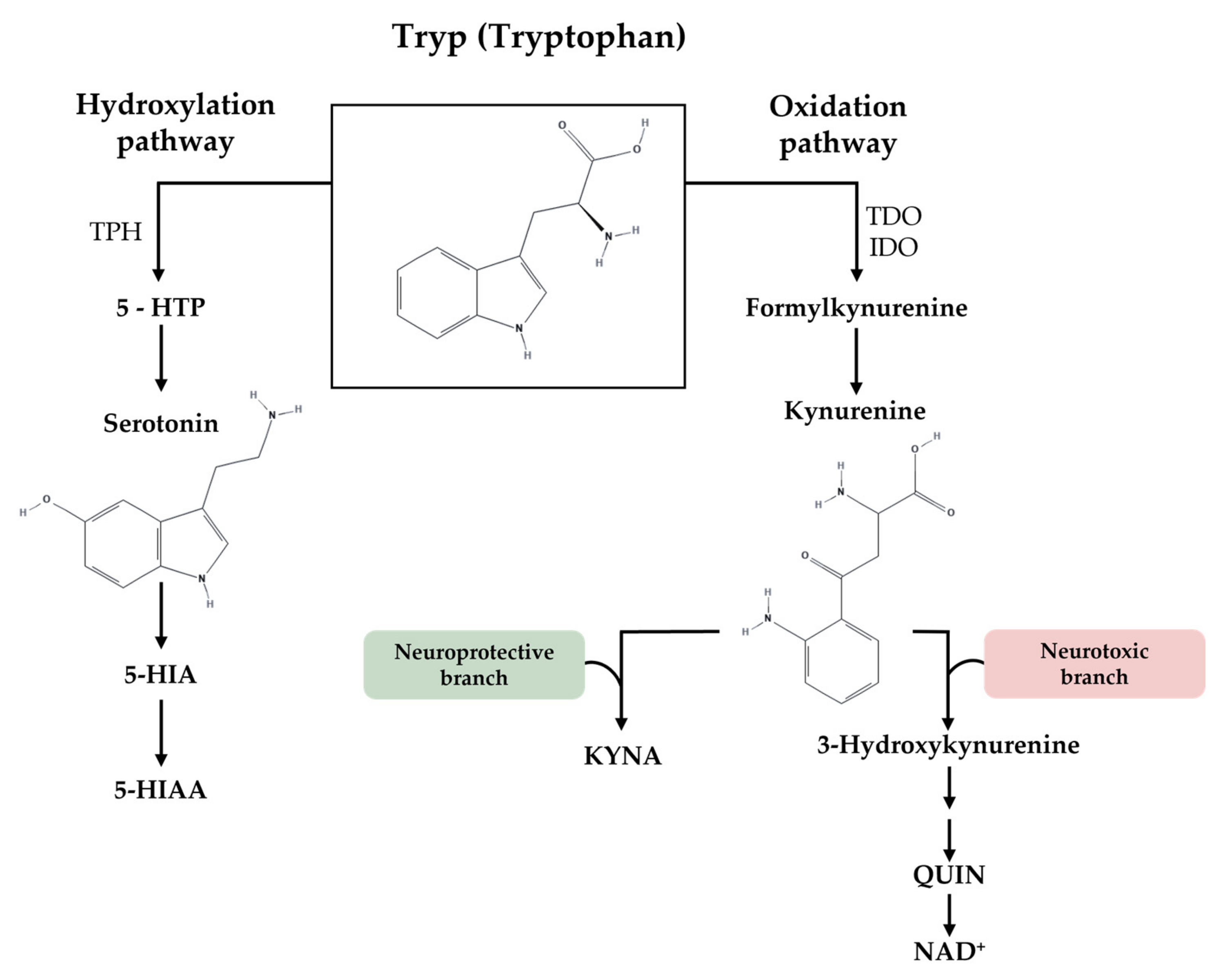
1.1. Sex Differences in Tryptophan Metabolism
1.1.1. Hydroxylation Pathway: Tryptophan and 5-HT Synthesis
1.1.2. Oxidation Pathway: Tryptophan and Kynurenine Metabolites
1.2. Impact of Tryptophan Nutritional Status between Sexes
1.2.1. Tryptophan Supplementation
1.2.2. Tryptophan Depletion
1.3. Effect of Sex Hormones on Tryptophan Metabolism

{kind=link}
{kind=link}
{kind=link}
| Sex Hormone | Study | Effect on Tryptophan Metabolism |
|---|---|---|
| Oestrogen | Preclinical | ↓ extracellular 5-HT in the hypothalamus [74,75] ↓ hippocampal brain levels of Tryp (ATD condition) [29] |
| Clinical | ↓ cortical trapping of α-[11C]methyl-L-tryptophan $ [76] ↓ circulating levels of plasma Tryp [77] ↓ circulating levels of plasma kyn and picolinic acid [78] | |
| Progesterone | In vitro | ↑ KYNA levels (cultures of human macrophages) [79] ↓ QUIN levels (cultures of human macrophages) [79] |
| Preclinical | ↑ brain levels of KYNA after administration of L-kyn [80] ↓ L-kynurenine-induced cortical spreading depression [80] | |
| Clinical | ↑ circulating levels of plasma kyn and urinary kyn excretion [81] ↑ circulating levels of plasma KYNA [17] ↓ inflammation-induced activation of IDO [82] ↓ Tryp catabolism to kyn and neurotoxic metabolites [78,82] | |
| Testosterone | Preclinical | Lack of testosterone attenuated hippocampal KYNA levels (sleep deprivation condition) [30] |
| Clinical | ↑ circulating levels of plasma Tryp [77] |
1.3.1. Oestrogen
1.3.2. Progesterone
1.3.3. Testosterone
2. Results and Discussion
2.1. Sex Differences in Tryptophan Metabolism: Neuropsychiatric Conditions
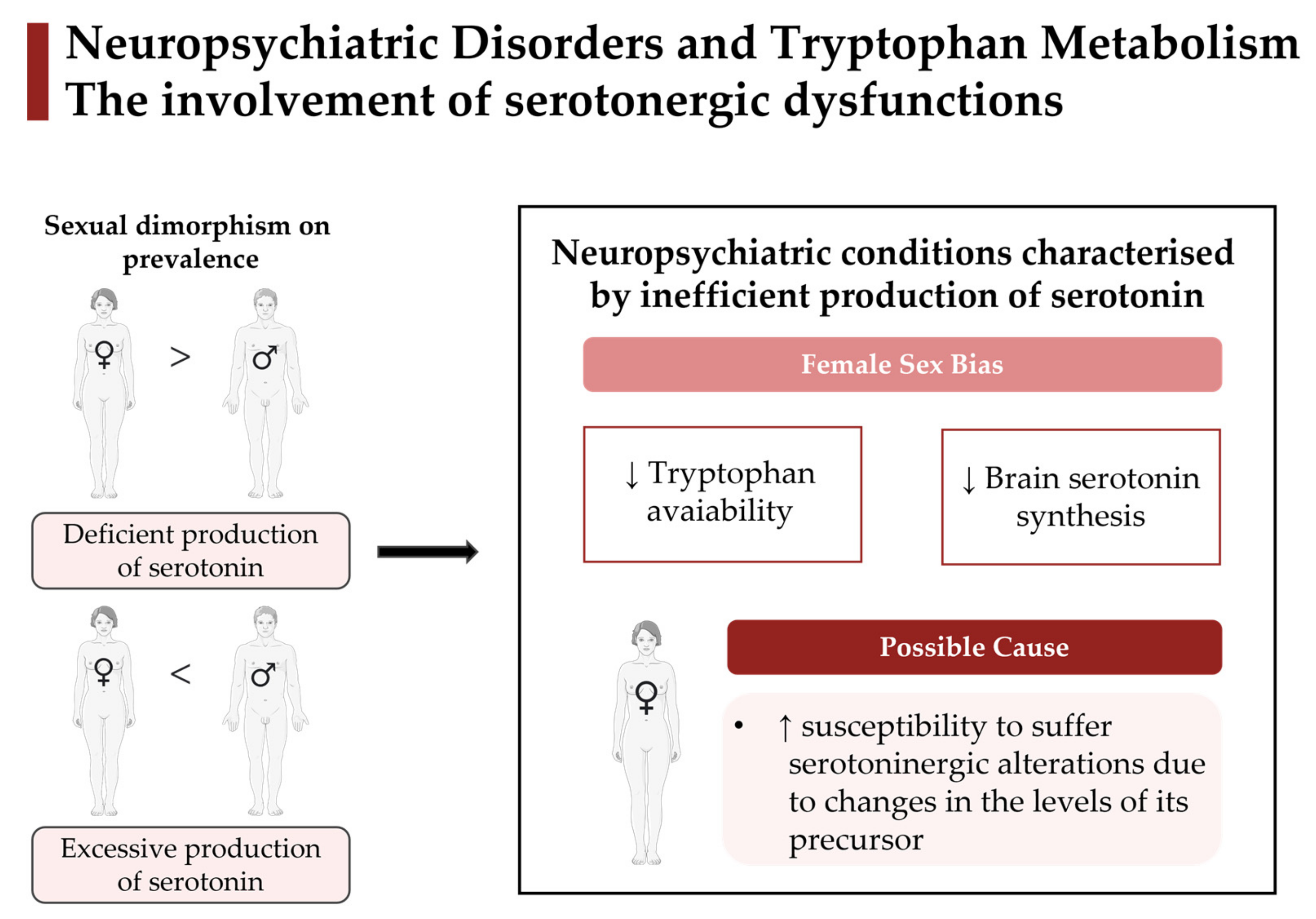
2.1.1. Sexual Dimorphisms on Tryptophan Metabolism: Female Sex Bias for Neuropsychiatric Prevalence
2.1.2. Sexual Dimorphisms on Tryptophan Metabolism: Non-Female Sex Bias for Neuropsychiatric Prevalence
2.1.3. Current Understanding of How Sexual Dimorphisms on Tryptophan Metabolism May Be Involved in the Onset and Progression of Neuropsychiatric Diseases
2.1.4. Limitations and Future Perspectives
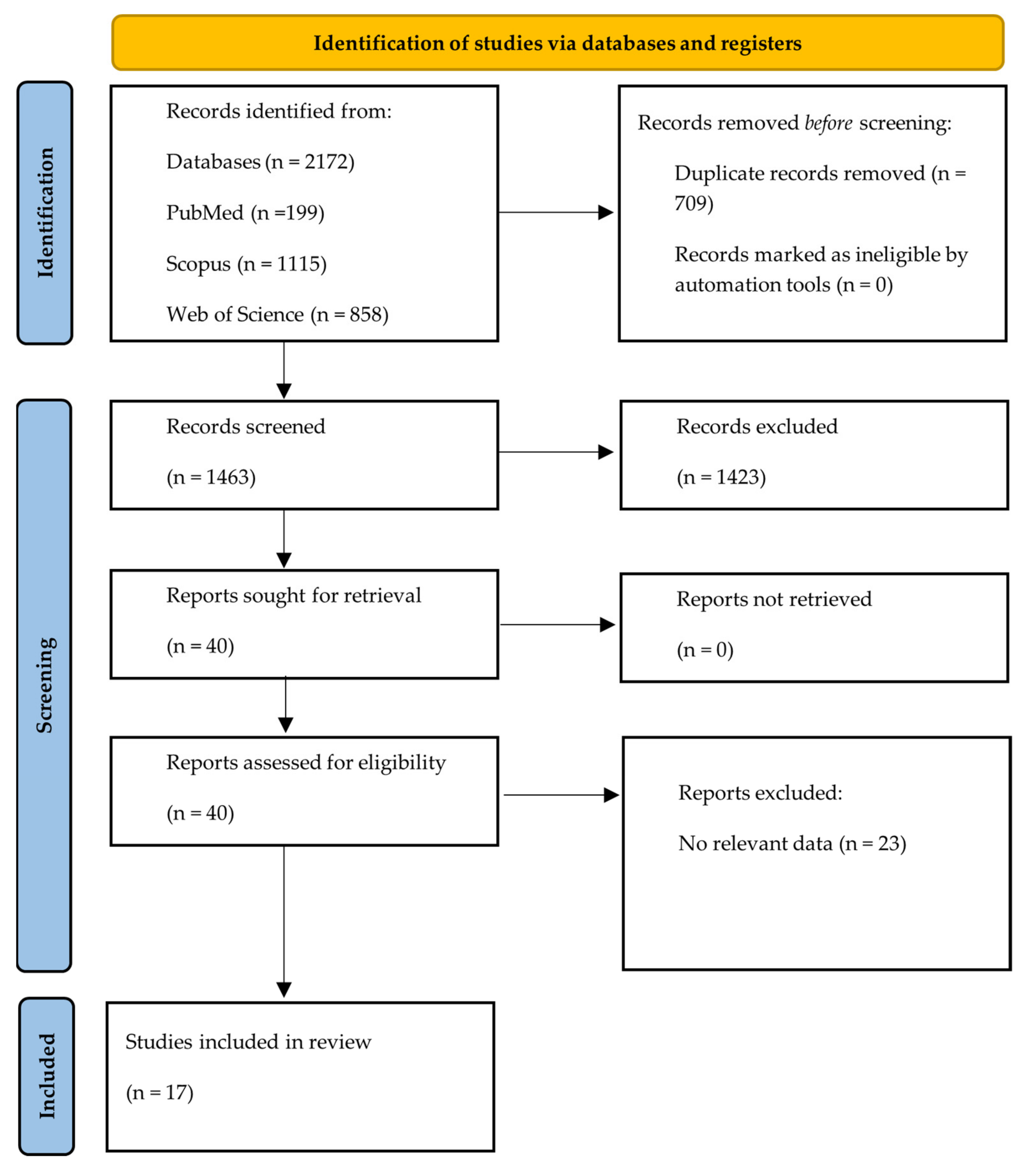
3. Methods
3.1. Study Eligibility
3.2. Search Strategy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruddick, J.P.; Evans, A.K.; Nutt, D.J.; Lightman, S.L.; Rook, G.A.W.; Lowry, C.A. Tryptophan metabolism in the central nervous system: Medical implications. Expert Rev. Mol. Med. 2006, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Van der Leek, A.P.; Yanishevsky, Y.; Kozyrskyj, A.L. The Kynurenine Pathway as a Novel Link between Allergy and the Gut Microbiome. Front. Immunol. 2017, 8, 1374. [Google Scholar] [CrossRef] [PubMed]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Giaroni, C.; Baj, A. Tryptophan Metabolites along the Microbiota-Gut-Brain Axis: An Interkingdom Communication System Influencing the Gut in Health and Disease. Int. J. Tryptophan Res. 2020, 13, 1178646920928984. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, C.E.; Martin, J.A.; Manriquez, F.V.; Dinan, T.G.; Cryan, J.F.; Clarke, G. Focus on the essentials: Tryptophan metabolism and the microbiome-gut-brain axis. Curr. Opin. Pharmacol. 2019, 48, 137–145. [Google Scholar] [CrossRef]
- Zahar, S.; Schneider, N.; Makwana, A.; Chapman, S.; Corthesy, J.; Amico, M.; Hudry, J. Dietary tryptophan-rich protein hydrolysate can acutely impact physiological and psychological measures of mood and stress in healthy adults. Nutr. Neurosci. 2022, 26, 303–312. [Google Scholar] [CrossRef]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in health and disease. Adv. Clin. Chem. 2020, 95, 165–218. [Google Scholar]
- Gostner, J.M.; Geisler, S.; Stonig, M.; Mair, L.; Sperner-Unterweger, B.; Fuchs, D. Tryptophan Metabolism and Related Pathways in Psychoneuroimmunology: The Impact of Nutrition and Lifestyle. Neuropsychobiology 2020, 79, 89–99. [Google Scholar] [CrossRef]
- Ciranna, L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Campbell, B.M.; Charych, E.; Lee, A.W.; Möller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front. Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef]
- Mithaiwala, M.N.; Santana-Coelho, D.; Porter, G.A.; O’Connor, J.C. Neuroinflammation and the Kynurenine Pathway in CNS Disease: Molecular Mechanisms and Therapeutic Implications. Cells 2021, 10, 1548. [Google Scholar] [CrossRef]
- Lemos, H.; Huang, L.; Prendergast, G.C.; Mellor, A.L. Immune control by amino acid catabolism during tumorigenesis and therapy. Nat. Rev. Cancer 2019, 19, 162–175. [Google Scholar] [CrossRef]
- Yeung, A.W.S.; Terentis, A.C.; King, N.J.C.; Thomas, S.R. Role of indoleamine 2,3-dioxygenase in health and disease. Clin. Sci. 2015, 129, 601–672. [Google Scholar] [CrossRef]
- Zinger, A.; Barcia, C.; Herrero, M.T.; Guillemin, G.J. The involvement of neuroinflammation and Kynurenine pathway in Parkinson’s disease. Park. Dis. 2011, 2011, 716859. [Google Scholar] [CrossRef]
- Savino, R.; Carotenuto, M.; Polito, A.N.; Di Noia, S.; Albenzio, M.; Scarinci, A.; Ambrosi, A.; Sessa, F.; Tartaglia, N.; Messina, G. Analyzing the potential biological determinants of Autism Spectrum Disorders: From Neuroinflammation to Kynurenines Pathway. Brain Sci. 2020, 10, 631. [Google Scholar] [CrossRef]
- Heyes, M.P.; Saito, K.; Crowley, J.S.; Davis, L.E.; Demitrack, M.A.; Der, M.; Dilling, L.A.; Elia, J.; Kruesi, M.J.P.; Lackner, A.; et al. Quinolinic acid and kynurenine pathway metabolism in inflammatory and non-inflammatory neurological disease. Brain 1992, 115, 1249–1273. [Google Scholar] [CrossRef]
- Meier, T.B.; Drevets, W.C.; Teague, T.K.; Wurfel, B.E.; Mueller, S.C.; Bodurka, J.; Dantzer, R.; Savitz, J. Kynurenic Acid is Reduced in Females and Oral Contraceptive Users: Implications for Depression. Brain Behav. Immun. 2018, 67, 59. [Google Scholar] [CrossRef]
- Stone, T.W.; Mackay, G.M.; Forest, C.M.; Clark, J.; Darlington, L.G. Tryptophan metabolites and brain disorders. Clin. Chem. Lab. Med. 2003, 41, 852–859. [Google Scholar] [CrossRef]
- Jans, L.A.W.; Riedel, W.J.; Markus, C.R.; Blokland, A. Serotonergic vulnerability and depression: Assumptions, experimental evidence and implications. Mol. Psychiatry 2007, 12, 522. [Google Scholar] [CrossRef]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology 2017, 112, 237–247. [Google Scholar] [CrossRef]
- Holden, C. Sex and the suffering brain. Science 2005, 308, 1574–1577. [Google Scholar] [CrossRef] [PubMed]
- Pinares-Garcia, P.; Stratikopoulos, M.; Zagato, A.; Loke, H.; Lee, J. Sex: A Significant Risk Factor for Neurodevelopmental and Neurodegenerative Disorders. Brain Sci. 2018, 8, 154. [Google Scholar] [CrossRef] [PubMed]
- Songtachalert, T.; Roomruangwong, C.; Carvalho, A.F.; Bourin, M.; Maes, M. Anxiety Disorders: Sex Differences in Serotonin and Tryptophan Metabolism. Curr. Top. Med. Chem. 2018, 18, 1704–1715. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Ferreira, H.; Martins, J.; Gonçalves, J.; Castelo-Branco, M. Male sex bias in early and late onset neurodevelopmental disorders: Shared aspects and differences in Autism Spectrum Disorder, Attention Deficit/hyperactivity Disorder, and Schizophrenia. Neurosci. Biobehav. Rev. 2022, 135, 104577. [Google Scholar] [CrossRef]
- Saldanha, D.; Kumar, N.; Ryali, V.S.S.R.; Srivastava, K.; Pawar, A.A. Serum Serotonin Abnormality in Depression. Med. J. Armed Forces India 2009, 65, 108. [Google Scholar] [CrossRef]
- Vahid-Ansari, F.; Albert, P.R. Rewiring of the Serotonin System in Major Depression. Front. Psychiatry 2021, 12, 2275. [Google Scholar] [CrossRef]
- Setiawati, Y.; Wahyuhadi, J.; Mukono, H.J.; Warsiki, E.; Yuniar, S. Is there an Effect of Serotonin on Attention Deficit Hyperactivity Disorder. Artic. Indian J. Public Health Res. Dev. 2020, 11, 1745. [Google Scholar] [CrossRef]
- Clayton, J.A. Sex influences in neurological disorders: Case studies and perspectives. Dialogues Clin. Neurosci. 2016, 18, 357. [Google Scholar] [CrossRef]
- Jans, L.A.W.; Lieben, C.K.J.; Blokland, A. Influence of sex and estrous cycle on the effects of acute tryptophan depletion induced by a gelatin-based mixture in adult Wistar rats. Neuroscience 2007, 147, 304–317. [Google Scholar] [CrossRef]
- Baratta, A.M.; Buck, S.A.; Buchla, A.D.; Fabian, C.B.; Chen, S.; Mong, J.A.; Pocivavsek, A. Sex Differences in Hippocampal Memory and Kynurenic Acid Formation Following Acute Sleep Deprivation in Rats. Sci. Rep. 2018, 8, 6963. [Google Scholar] [CrossRef]
- Alberghina, D.; Giannetto, C.; Visser, E.K.; Ellis, A.D. Effect of diet on plasma tryptophan and serotonin in trained mares and geldings. Vet. Rec. 2010, 166, 133–136. [Google Scholar] [CrossRef]
- Albay, R.; Chen, A.; Anderson, G.M.; Tatevosyan, M.; Janušonis, S. Relationships among body mass, brain size, gut length, and blood tryptophan and serotonin in young wild-type mice. BMC Physiol. 2009, 9, 4. [Google Scholar] [CrossRef]
- Yao, J.; Lu, H.; Wang, Z.; Wang, T.; Fang, F.; Wang, J.; Yu, J.; Gao, R. A sensitive method for the determination of the gender difference of neuroactive metabolites in tryptophan and dopamine pathways in mouse serum and brain by UHPLC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1093–1094, 91–99. [Google Scholar] [CrossRef]
- Guzmán, D.C.; Ruíz, N.L.; García, E.H.; Mejía, G.B.; Téllez, P.P.; Jimenez, G.E.; De la Rosa Apreza, M.; Olguín, H.J. Effect of sibutramine on Na+, K+ ATPase activity and tryptophan levels on male and female rat brain. Horm. Metab. Res. 2009, 41, 363–367. [Google Scholar] [CrossRef]
- Carlsson, M.; Carlsson, A. A regional study of sex differences in rat brain serotonin. Prog. Neuro Psychopharmacol. Biol. Psychiatry 1988, 12, 53–61. [Google Scholar] [CrossRef]
- Domínguez, R.; Cruz-Morales, S.E.; Carvalho, M.C.; Xavier, M.; Brandao, M.L. Sex differences in serotonergic activity in dorsal and median raphe nucleus. Physiol. Behav. 2003, 80, 203–210. [Google Scholar] [CrossRef]
- Haleem, D.J.; Kennett, G.A.; Curzon, G. Hippocampal 5-hydroxytryptamine synthesis is greater in female rats than in males and more decreased by the 5-HT1A agonist 8-OH-DPAT. J. Neural Transm. Gen. Sect. 1990, 79, 93–101. [Google Scholar] [CrossRef]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef]
- Badawy, A.A.B.; Dougherty, D.M. Assessment of the Human Kynurenine Pathway: Comparisons and Clinical Implications of Ethnic and Gender Differences in Plasma Tryptophan, Kynurenine Metabolites, and Enzyme Expressions at Baseline and After Acute Tryptophan Loading and Depletion. Int. J. Tryptophan Res. 2016, 9, 31–49. [Google Scholar] [CrossRef]
- Blennow, K.; Wallin, A.; Gottfries, C.G.; Karlsson, I.; Månsson, J.E.; Skoog, I.; Wikkelsö, C.; Svennerholm, L. Cerebrospinal fluid monoamine metabolites in 114 healthy individuals 18–88 years of age. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 1993, 3, 55–61. [Google Scholar] [CrossRef]
- Brewerton, T.D.; Putnam, K.T.; Lewine, R.R.J.; Risch, S.C. Seasonality of cerebrospinal fluid monoamine metabolite concentrations and their associations with meteorological variables in humans. J. Psychiatr. Res. 2018, 99, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B.; Morgan, C.J.; Llewelyn, M.B.; Albuquerque, S.R.J.; Farmer, A. Heterogeneity of serum tryptophan concentration and availability to the brain in patients with the chronic fatigue syndrome. J. Psychopharmacol. 2005, 19, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Raheja, U.K.; Fuchs, D.; Giegling, I.; Brenner, L.A.; Rovner, S.F.; Mohyuddin, I.; Weghuber, D.; Mangge, H.; Rujescu, D.; Postolache, T.T. In psychiatrically healthy individuals, overweight women but not men have lower tryptophan levels. Pteridines 2015, 26, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Vidal, R.; García-Marchena, N.; O’Shea, E.; Requena-Ocaña, N.; Flores-López, M.; Araos, P.; Serrano, A.; Suárez, J.; Rubio, G.; Rodríguez de Fonseca, F.; et al. Plasma tryptophan and kynurenine pathway metabolites in abstinent patients with alcohol use disorder and high prevalence of psychiatric comorbidity. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 102, 109958. [Google Scholar] [CrossRef]
- Hestad, K.A.; Engedal, K.; Whist, J.E.; Farup, P.G. The Relationships among Tryptophan, Kynurenine, Indoleamine 2,3-Dioxygenase, Depression, and Neuropsychological Performance. Front. Psychol. 2017, 8, 1561. [Google Scholar] [CrossRef]
- Horn, J.; Mayer, D.E.; Chen, S.; Mayer, E.A. Role of diet and its effects on the gut microbiome in the pathophysiology of mental disorders. Transl. Psychiatry 2022, 12, 164. [Google Scholar] [CrossRef]
- Grzymisławska, M.; Puch, E.A.; Zawada, A.; Grzymisławski, M. Do nutritional behaviors depend on biological sex and cultural gender? Adv. Clin. Exp. Med. 2020, 29, 165–172. [Google Scholar] [CrossRef]
- Gibson, E.L. Tryptophan supplementation and serotonin function: Genetic variations in behavioural effects. Proc. Nutr. Soc. 2018, 77, 174–188. [Google Scholar] [CrossRef]
- Nayak, B.; Singh, R.; Buttar, H. Role of Tryptophan in Health and Disease: Systematic Review of the Anti-Oxidant, Anti-Inflammation, and Nutritional Aspects of Tryptophan and Its Metabolites. 2019. Available online: https://www.researchgate.net/publication/336104734_Role_of_Tryptophan_in_Health_and_Disease_Systematic_Review_of_the_Anti-Oxidant_Anti-Inflammation_and_Nutritional_Aspects_of_Tryptophan_and_Its_Metabolites (accessed on 10 July 2022).
- Beretich, G.R. Reversal of autistic symptoms by removal of low-relative tryptophan foods: Case report. Med. Hypotheses 2009, 73, 856–857. [Google Scholar] [CrossRef]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chirumbolo, S.; Chartrand, M.S.; Bjørklund, G. How important is tryptophan in human health? Crit. Rev. Food Sci. Nutr. 2019, 59, 72–88. [Google Scholar] [CrossRef]
- Badawy, A.A.B. Modulation of Tryptophan and Serotonin Metabolism as a Biochemical Basis of the Behavioral Effects of Use and Withdrawal of Androgenic-Anabolic Steroids and Other Image- and Performance-Enhancing Agents. Int. J. Tryptophan Res. 2018, 11, 1178646917753422. [Google Scholar] [CrossRef]
- Walsh, A.E.S.; Oldman, A.D.; Franklin, M.; Fairburn, C.G.; Cowen, P.J. Dieting decreases plasma tryptophan and increases the prolactin response to d-fenfluramine in women but not men. J. Affect. Disord. 1995, 33, 89–97. [Google Scholar] [CrossRef]
- Suarez, E.C.; Krishnan, K.R.R. The relation of free plasma tryptophan to anger, hostility, and aggression in a nonpatient sample of adult men and women. Ann. Behav. Med. A Publ. Soc. Behav. Med. 2006, 31, 254–260. [Google Scholar] [CrossRef]
- Blankfield, A. A Brief Historic Overview of Clinical Disorders Associated with Tryptophan: The Relevance to Chronic Fatigue Syndrome (CFS) and Fibromyalgia (FM). Int. J. Tryptophan Res. 2012, 5, 27. [Google Scholar] [CrossRef]
- McDougle, C. Effects of Tryptophan Depletion in Drug-Free Adults With Autistic Disorder. Arch. Gen. Psychiatry 1996, 53, 993. [Google Scholar] [CrossRef]
- Correia, A.S.; Vale, N. Tryptophan Metabolism in Depression: A Narrative Review with a Focus on Serotonin and Kynurenine Pathways. Int. J. Mol. Sci. 2022, 23, 8493. [Google Scholar] [CrossRef]
- Weaver, D.; Gupta, M.; Meek, A.; Wang, Y.; Wu, F. Alzheimer’s Disease as a Disorder of Tryptophan Metabolism (2745). Neurology 2020, 94. [Google Scholar]
- Nishizawa, S.; Benkelfat, C.; Young, S.N.; Leyton, M.; Mzengeza, S.; De Montigny, C.; Blier, P.; Diksic, M. Differences between males and females in rates of serotonin synthesis in human brain. Proc. Natl. Acad. Sci. USA 1997, 94, 5308–5313. [Google Scholar] [CrossRef]
- Sambeth, A.; Blokland, A.; Harmer, C.J.; Kilkens, T.O.C.; Nathan, P.J.; Porter, R.J.; Schmitt, J.A.J.; Scholtissen, B.; Sobczak, S.; Young, A.H.; et al. Sex differences in the effect of acute tryptophan depletion on declarative episodic memory: A pooled analysis of nine studies. Neurosci. Biobehav. Rev. 2007, 31, 516–529. [Google Scholar] [CrossRef]
- Ellenbogen, M.A.; Young, S.N.; Dean, P.; Palmour, R.M.; Benkelfat, C. Mood response to acute tryptophan depletion in healthy volunteers: Sex differences and temporal stability. Neuropsychopharmacology 1996, 15, 465–474. [Google Scholar] [CrossRef]
- Schmeck, K.; Sadigorsky, S.; Englert, E.; Demisch, L.; Dierks, T.; Barta, S.; Poustka, F. Mood Changes following Acute Tryptophan Depletion in Healthy Adults. Psychopathology 2002, 35, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Rogers, R.D.; Tunbridge, E.; Cowen, P.J.; Goodwin, G.M. Tryptophan depletion decreases the recognition of fear in female volunteers. Psychopharmacology 2003, 167, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, J.; McEwen, B.S. Sex in the brain: Hormones and sex differences. Dialogues Clin. Neurosci. 2016, 18, 373. [Google Scholar] [CrossRef] [PubMed]
- Tramunt, B.; Smati, S.; Grandgeorge, N.; Lenfant, F.; Arnal, J.F.; Montagner, A.; Gourdy, P. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia 2020, 63, 453–461. [Google Scholar] [CrossRef]
- Simpson, J.; Kelly, J.P. An investigation of whether there are sex differences in certain behavioural and neurochemical parameters in the rat. Behav. Brain Res. 2012, 229, 289–300. [Google Scholar] [CrossRef]
- Jayawickrama, G.S.; Nematollahi, A.; Sun, G.; Gorrell, M.D.; Church, W.B. Inhibition of human kynurenine aminotransferase isozymes by estrogen and its derivatives. Sci. Rep. 2017, 7, 17559. [Google Scholar] [CrossRef]
- Raghunath, R.S.; Venables, Z.C.; Millington, G.W.M. The menstrual cycle and the skin. Clin. Exp. Dermatol. 2015, 40, 111–115. [Google Scholar] [CrossRef]
- Fink, G.; Sumner, B.E.H.; McQueen, J.K.; Wilson, H.; Rosie, R. Sex steroid control of mood, mental state and memory. Clin. Exp. Pharmacol. Physiol. 1998, 25, 764–775. [Google Scholar] [CrossRef]
- Roeder, H.J.; Leira, E.C. Effects of the Menstrual Cycle on Neurological Disorders. Curr. Neurol. Neurosci. Rep. 2021, 21, 34. [Google Scholar] [CrossRef]
- Frokjaer, V.G.; Pinborg, A.; Holst, K.K.; Overgaard, A.; Henningsson, S.; Heede, M.; Larsen, E.C.; Jensen, P.S.; Agn, M.; Nielsen, A.P.; et al. Role of Serotonin Transporter Changes in Depressive Responses to Sex-Steroid Hormone Manipulation: A Positron Emission Tomography Study. Biol. Psychiatry 2015, 78, 534–543. [Google Scholar] [CrossRef]
- Munk-Olsen, T.; Laursen, T.M.; Pedersen, C.B.; Mors, O.; Mortensen, P.B. New parents and mental disorders: A population-based register study. JAMA 2006, 296, 2582–2589. [Google Scholar] [CrossRef]
- Scharfman, H.E.; MacLusky, N.J. The influence of gonadal hormones on neuronal excitability, seizures, and epilepsy in the female. Epilepsia 2006, 47, 1423–1440. [Google Scholar] [CrossRef]
- Gundlah, C.; Simon, L.D.; Auerbach, S.B. Differences in hypothalamic serotonin between estrous phases and gender: An in vivo microdialysis study. Brain Res. 1998, 785, 91–96. [Google Scholar] [CrossRef]
- Maswood, S.; Truitt, W.; Hotema, M.; Caldarola-Pastuszka, M.; Uphouse, L. Estrous cycle modulation of extracellular serotonin in mediobasal hypothalamus: Role of the serotonin transporter and terminal autoreceptors. Brain Res. 1999, 831, 146–154. [Google Scholar] [CrossRef]
- Sakai, Y.; Nishikawa, M.; Leyton, M.; Benkelfat, C.; Young, S.N.; Diksic, M. Cortical trapping of alpha-[(11)C]methyl-l-tryptophan, an index of serotonin synthesis, is lower in females than males. NeuroImage 2006, 33, 815–824. [Google Scholar] [CrossRef]
- Giltay, E.J.; Bunck, M.C.; Gooren, L.J.; Zitman, F.G.; Diamant, M.; Teerlink, T. Effects of sex steroids on the neurotransmitter-specific aromatic amino acids phenylalanine, tyrosine, and tryptophan in transsexual subjects. Neuroendocrinology 2008, 88, 103–110. [Google Scholar] [CrossRef]
- Sha, Q.; Achtyes, E.; Nagalla, M.; Keaton, S.; Smart, L.A.; Leach, R.; Brundin, L. Associations between estrogen and progesterone, the kynurenine pathway, and inflammation in the post-partum. J. Affect. Disord. 2021, 281, 9–12. [Google Scholar] [CrossRef]
- De Bie, J.; Lim, C.K.; Guillemin, G.J. Progesterone Alters Kynurenine Pathway Activation in IFN-γ-Activated Macrophages—Relevance for Neuroinflammatory Diseases. Int. J. Tryptophan Res. 2016, 9, 89–93. [Google Scholar] [CrossRef]
- Chauvel, V.; Vamos, E.; Pardutz, A.; Vecsei, L.; Schoenen, J.; Multon, S. Effect of systemic kynurenine on cortical spreading depression and its modulation by sex hormones in rat. Exp. Neurol. 2012, 236, 207–214. [Google Scholar] [CrossRef]
- Hrboticky, N.; Leiter, L.A.; Anderson, G.H. Menstrual cycle effects on the metabolism of tryptophan loads. Am. J. Clin. Nutr. 1989, 50, 46–52. [Google Scholar] [CrossRef]
- Nagy, B.; Szekeres-Barthó, J.; Kovács, G.L.; Sulyok, E.; Farkas, B.; Várnagy, Á.; Vértes, V.; Kovács, K.; Bódis, J. Key to Life: Physiological Role and Clinical Implications of Progesterone. Int. J. Mol. Sci. 2021, 22, 11039. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Akhter, S.; Afridi, M.I. Gender based response to fluoxetine hydrochloride medication in endogenous depression. J. Coll. Physicians Surg. Pak. 2004, 14, 161–165. [Google Scholar] [PubMed]
- Pivac, N.; Mück-Šeler, D.; Mustapić, M.; Nenadić-Šviglin, K.; Kozarić-Kovačić, D. Platelet serotonin concentration in alcoholic subjects. Life Sci. 2004, 76, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Booij, L.; Van Der Does, A.J.W.; Haffmans, P.M.J.; Spinhoven, P.; McNally, R.J. Acute tryptophan depletion as a model of depressive relapse: Behavioural specificity and ethical considerations. Br. J. Psychiatry J. Ment. Sci. 2005, 187, 148–154. [Google Scholar] [CrossRef]
- Moreno, F.A.; McGahuey, C.A.; Freeman, M.P.; Delgado, P.L. Sex differences in depressive response during monoamine depletions in remitted depressive subjects. J. Clin. Psychiatry 2006, 67, 1618–1623. [Google Scholar] [CrossRef]
- Weaver, S.A.; Janal, M.N.; Aktan, N.; Ottenweller, J.E.; Natelson, B.H. Sex differences in plasma prolactin response to tryptophan in chronic fatigue syndrome patients with and without comorbid fibromyalgia. J. Women’s Health 2010, 19, 951–958. [Google Scholar] [CrossRef]
- Frey, B.N.; Skelin, I.; Sakai, Y.; Nishikawa, M.; Diksic, M. Gender differences in alpha-[(11)C]MTrp brain trapping, an index of serotonin synthesis, in medication-free individuals with major depressive disorder: A positron emission tomography study. Psychiatry Res. 2010, 183, 157–166. [Google Scholar] [CrossRef]
- Kötting, W.F.; Bubenzer, S.; Helmbold, K.; Eisert, A.; Gaber, T.J.; Zepf, F.D. Effects of tryptophan depletion on reactive aggression and aggressive decision-making in young people with ADHD. Acta Psychiatr. Scand. 2013, 128, 114–123. [Google Scholar] [CrossRef]
- Young, S.N.; Regoli, M.; Leyton, M.; Pihl, R.O.; Benkelfat, C. The effect of acute tryptophan depletion on mood and impulsivity in polydrug ecstasy users. Psychopharmacology 2014, 231, 707–716. [Google Scholar] [CrossRef]
- Booij, L.; Soucy, J.P.; Young, S.N.; Regoli, M.; Gravel, P.; Diksic, M.; Leyton, M.; Pihl, R.O.; Benkelfat, C. Brain serotonin synthesis in MDMA (ecstasy) polydrug users: An alpha-[(11) C]methyl-l-tryptophan study. J. Neurochem. 2014, 131, 634–644. [Google Scholar] [CrossRef]
- Platzer, M.; Dalkner, N.; Fellendorf, F.T.; Birner, A.; Bengesser, S.A.; Queissner, R.; Kainzbauer, N.; Pilz, R.; Herzog-Eberhard, S.; Hamm, C.; et al. Tryptophan breakdown and cognition in bipolar disorder. Psychoneuroendocrinology 2017, 81, 144–150. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, W.; Liu, W.; Wang, C.; Zhan, Y.; Li, H.; Chen, L.; Ning, Y. Cross-sectional relationship between kynurenine pathway metabolites and cognitive function in major depressive disorder. Psychoneuroendocrinology 2019, 101, 72–79. [Google Scholar] [CrossRef]
- Reininghaus, E.Z.; Dalkner, N.; Riedrich, K.; Fuchs, D.; Gostner, J.M.; Reininghaus, B. Sex Specific Changes in Tryptophan Breakdown Over a 6 Week Treatment Period. Front. Psychiatry 2019, 10, 74. [Google Scholar] [CrossRef]
- Xu, J.; Green, R.; Kim, M.; Lord, J.; Ebshiana, A.; Westwood, S.; Baird, A.L.; Nevado-Holgado, A.J.; Shi, L.; Hye, A.; et al. Sex-Specific Metabolic Pathways Were Associated with Alzheimer’s Disease (AD) Endophenotypes in the European Medical Information Framework for AD Multimodal Biomarker Discovery Cohort. Biomedicines 2021, 9, 1610. [Google Scholar] [CrossRef]
- Cheng, Z.; Peng, Y.; Wen, J.; Chen, W.; Pan, W.; Xu, X.; Lu, X.; Cai, Q.; Ge, F.; Fan, Y.; et al. Sex-specific metabolic signatures in methamphetamine addicts. Addict. Biol. 2023, 28, e13255. [Google Scholar] [CrossRef]
- Berney, A.; Leyton, M.; Gravel, P.; Sibon, I.; Sookman, D.; Rosa Neto, P.; Diksic, M.; Nakai, A.; Pinard, G.; Todorov, C.; et al. Brain regional α-[11C]methyl-L-tryptophan trapping in medication-free patients with obsessive-compulsive disorder. Arch. Gen. Psychiatry 2011, 68, 732–741. [Google Scholar] [CrossRef]
- Chugani, D.C. Serotonin in autism and pediatric epilepsies. Ment. Retard. Dev. Disabil. Res. Rev. 2004, 10, 112–116. [Google Scholar] [CrossRef]
- Sağlam, E.; Bilgiç, A.; Abuşoğlu, S.; Ünlü, A.; Sivrikaya, A. The role of tryptophan metabolic pathway in children with attention deficit hyperactivity disorder with and without comorbid oppositional defiant disorder and conduct disorder. Psychiatry Res. 2021, 298, 113770. [Google Scholar] [CrossRef]
- Nakatomi, Y.; Mizuno, K.; Ishii, A.; Wada, Y.; Tanaka, M.; Tazawa, S.; Onoe, K.; Fukuda, S.; Kawabe, J.; Takahashi, K.; et al. Neuroinflammation in Patients with Chronic Fatigue Syndrome/Myalgic Encephalomyelitis: An 11 C-( R )-PK11195 PET Study. J. Nucl. Med. 2014, 55, 945–950. [Google Scholar] [CrossRef]
- Kohno, M.; Link, J.; Dennis, L.E.; McCready, H.; Huckans, M.; Hoffman, W.F.; Loftis, J.M. Neuroinflammation in addiction: A review of neuroimaging studies and potential immunotherapies. Pharmacol. Biochem. Behav. 2019, 179, 34. [Google Scholar] [CrossRef]
- Widner, B.; Leblhuber, F.; Walli, J.; Tilz, G.P.; Demel, U.; Fuchs, D. Tryptophan degradation and immune activation in Alzheimer’s disease. J. Neural Transm. 2000, 107, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Whiley, L.; Chappell, K.E.; D’Hondt, E.; Lewis, M.R.; Jiménez, B.; Snowden, S.G.; Soininen, H.; Kłoszewska, I.; Mecocci, P.; Tsolaki, M.; et al. Metabolic phenotyping reveals a reduction in the bioavailability of serotonin and kynurenine pathway metabolites in both the urine and serum of individuals living with Alzheimer’s disease. Alzheimer’s Res. Ther. 2021, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M. Potential Role of Neuroactive Tryptophan Metabolites in Central Fatigue: Establishment of the Fatigue Circuit. Int. J. Tryptophan Res. 2020, 13, 1178646920936279. [Google Scholar] [CrossRef] [PubMed]
- Ogyu, K.; Kubo, K.; Noda, Y.; Iwata, Y.; Tsugawa, S.; Omura, Y.; Wada, M.; Tarumi, R.; Plitman, E.; Moriguchi, S.; et al. Kynurenine pathway in depression: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2018, 90, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.M.; Flaviano, G. The kynurenine pathway and neurodegenerative disease. Semin. Cell Dev. Biol. 2015, 40, 134–141. [Google Scholar]
- Evangelisti, M.; De Rossi, P.; Rabasco, J.; Donfrancesco, R.; Lionetto, L.; Capi, M.; Sani, G.; Simmaco, M.; Nicoletti, F.; Villa, M.P. Changes in serum levels of kynurenine metabolites in paediatric patients affected by ADHD. Eur. Child Adolesc. Psychiatry 2017, 26, 1433–1441. [Google Scholar] [CrossRef]
- Badawy, A.A.-B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef]
- Dehhaghi, M.; Panahi, H.K.S.; Kavyani, B.; Heng, B.; Tan, V.; Braidy, N.; Guillemin, G.J. The Role of Kynurenine Pathway and NAD+ Metabolism in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Aging Dis. 2022, 13, 698. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Scharpè, S.; Bosmans, E.; Suy, E.; De Meester, I.; Calabrese, J.; Cosyns, P. Relationships between lower plasma L-tryptophan levels and immune-inflammatory variables in depression. Psychiatry Res. 1993, 49, 151–165. [Google Scholar] [CrossRef]
- Maes, M.; Wauters, A.; Verkerk, R.; Demedts, P.; Neels, H.; Van Gastel, A.; Cosyns, P.; Scharpé, S.; Desnyder, R. Lower serum L-tryptophan availability in depression as a marker of a more generalized disorder in protein metabolism. Neuropsychopharmacology 1996, 15, 243–251. [Google Scholar] [CrossRef]
- Simonato, M.; Dall’acqua, S.; Zilli, C.; Sut, S.; Tenconi, R.; Gallo, N.; Sfriso, P.; Sartori, L.; Cavallin, F.; Fiocco, U.; et al. Tryptophan Metabolites, Cytokines, and Fatty Acid Binding Protein 2 in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Biomedicines 2021, 9, 1724. [Google Scholar] [CrossRef]
- Groven, N.; Reitan, S.K.; Fors, E.A.; Guzey, I.C. Kynurenine metabolites and ratios differ between Chronic Fatigue Syndrome, Fibromyalgia, and healthy controls. Psychoneuroendocrinology 2021, 131, 105287. [Google Scholar] [CrossRef]
- Chugani, D.C.; Muzik, O. α[C-11]methyl-l-tryptophan PET maps brain serotonin synthesis and Kynurenine pathway metabolism. J. Cereb. Blood Flow Metab. 2000, 20, 2–9. [Google Scholar] [CrossRef]
- Chatterjee, P.; Zetterberg, H.; Goozee, K.; Lim, C.K.; Jacobs, K.R.; Ashton, N.J.; Hye, A.; Pedrini, S.; Sohrabi, H.R.; Shah, T.; et al. Plasma neurofilament light chain and amyloid-β are associated with the kynurenine pathway metabolites in preclinical Alzheimer’s disease. J. Neuroinflammation 2019, 16, 1–12. [Google Scholar] [CrossRef]
- Jeon, S.W.; Kim, Y.K. The role of neuroinflammation and neurovascular dysfunction in major depressive disorder. J. Inflamm. Res. 2018, 11, 179. [Google Scholar] [CrossRef]
- Broadbear, J.H.; Kabel, D.; Tracy, L.; Mak, P. Oxytocinergic regulation of endogenous as well as drug-induced mood. Pharmacol. Biochem. Behav. 2014, 119, 61–71. [Google Scholar] [CrossRef]
- Guinle, M.I.B.; Sinha, R. The Role of Stress, Trauma, and Negative Affect in Alcohol Misuse and Alcohol Use Disorder in Women. Alcohol Res. Curr. Rev. 2020, 40, 1–17. [Google Scholar] [CrossRef]
- Arnold, L.M. Gender differences in bipolar disorder. Psychiatr. Clin. N. Am. 2003, 26, 595–620. [Google Scholar] [CrossRef]
- Hebbrecht, K.; Skorobogatov, K.; Giltay, E.J.; Coppens, V.; De Picker, L.; Morrens, M. Tryptophan Catabolites in Bipolar Disorder: A Meta-Analysis. Front. Immunol. 2021, 12, 667179. [Google Scholar] [CrossRef]
- Zhang, P.; Huang, H.; Gao, X.; Jiang, J.; Xi, C.; Wu, L.; Fu, Y.; Lai, J.; Hu, S. Involvement of Kynurenine Metabolism in Bipolar Disorder: An Updated Review. Front. Psychiatry 2021, 12, 1270. [Google Scholar] [CrossRef] [PubMed]
- Plitman, E.; Iwata, Y.; Caravaggio, F.; Nakajima, S.; Chung, J.K.; Gerretsen, P.; Kim, J.; Takeuchi, H.; Chakravarty, M.M.; Remington, G.; et al. Kynurenic Acid in Schizophrenia: A Systematic Review and Meta-analysis. Schizophr. Bull. 2017, 43, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Neupane, S.P.; Lien, L.; Martinez, P.; Hestad, K.; Bramness, J.G. The relationship of alcohol-use disorders and depressive symptoms to tryptophan metabolism: Cross-sectional data from a Nepalese alcohol treatment sample. Alcohol. Clin. Exp. Res. 2015, 39, 514. [Google Scholar] [CrossRef] [PubMed]
- Sari, Y.; Johnson, V.R.; Weedman, J.M. Role of the Serotonergic System in Alcohol Dependence: From Animal Models to Clinics. Prog. Mol. Biol. Transl. Sci. 2011, 98, 401. [Google Scholar]
- Murakami, Y.; Imamura, Y.; Saito, K.; Sakai, D.; Motyama, J. Altered kynurenine pathway metabolites in a mouse model of human attention-deficit hyperactivity/autism spectrum disorders: A potential new biological diagnostic marker. Sci. Rep. 2019, 9, 13182. [Google Scholar] [CrossRef]
- Shuffrey, L.C.; Guter, S.J.; Delaney, S.; Jacob, S.; Anderson, G.M.; Sutcliffe, J.S.; Cook, E.H.; Veenstra-VanderWeele, J. Is there sexual dimorphism of hyperserotonemia in Autism Spectrum Disorder? Autism Res. 2017, 10, 1417. [Google Scholar] [CrossRef]
- Chakraborti, B.; Verma, D.; Guhathakurta, S.; Jaiswal, P.; Singh, A.S.; Sinha, S.; Ghosh, S.; Mukhopadhyay, K.; Mohanakumar, K.P.; Rajamma, U. Gender-Specific Effect of 5-HT and 5-HIAA on Threshold Level of Behavioral Symptoms and Sex-Bias in Prevalence of Autism Spectrum Disorder. Front. Neurosci. 2020, 13, 1375. [Google Scholar] [CrossRef]
- Jacobsen, J.P.; Rudder, M.L.; Roberts, W.; Royer, E.L.; Robinson, T.J.; Oh, A.; Spasojevic, I.; Sachs, B.D.; Caron, M.G. SSRI Augmentation by 5-Hydroxytryptophan Slow Release: Mouse Pharmacodynamic Proof of Concept. Neuropsychopharmacology 2016, 41, 2324–2334. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 88, 105906. [Google Scholar]



| Path | Study | Sexual Dimorphisms (♀ vs. ♂) | |
|---|---|---|---|
| Peripherical levels | Tryp | Preclinical | ↑ plasma Tryp levels [29,30,31] |
| Clinical | ↑ plasma-free Tryp levels [5,39] (=) plasma Tryp levels [42,43] ↓ plasma Tryp levels [44,45] | ||
| 5-HT | Preclinical | ↑ plasma 5-HT/Tryp ratio [32,33] | |
| Clinical | - | ||
| Kyn | Preclinical | ↓ plasma Kyn levels [30] (=) plasma KYNA levels [30] (=) plasma Kyn/Tryp ratio [33] | |
| Clinical | ↓ KYNA plasma levels [17] (=) Kyn and KYNA plasma levels [39] | ||
| Brain levels | Tryp | Preclinical | ↑ brain Tryp levels (brainstem, striatum and cortex) [35] ↑ brain Tryp levels (whole brain) [34] |
| Clinical | ↑ plasma Tryp/CAA ratios [5,39] (=) plasma Tryp/CAA ratios [42] | ||
| 5-HT | Preclinical | ↑ 5-HT levels (brainstem and limbic forebrain) and 5-HIAA (whole brain) [35] ↑ 5-HIAA/5-HT ratios (hypothalamus/preoptic area and limbic forebrain) [35] ↑ 5-HIAA/5-HT ratios (brainstem—dorsal raphe nucleus) [36] ↑ 5-HT and 5-HIAA (limbic forebrain—hippocampus) [37] | |
| Clinical | ↑ cerebrospinal fluid 5-HIAA levels [40,41] | ||
| Kyn | Preclinical | (=) KYNA levels (cortex and hippocampus) [30] (=) Kyn/Tryp ratios (whole brain) [33] | |
| Clinical | - |
| Condition | Parameters | Sexual Dimorphisms (♀ vs ♂) |
|---|---|---|
| Tryp supplementation | Biochemical | ↑ plasma-free Tryp [39] (=) plasma total kynurenines [39] |
| Behavioural | (≠) interaction between Tryp and mood (a positive trend in men and a negative trend in women) [5] | |
| Tryp depletion | Biochemical | ↓ trapping of α-[11C]methyl-L-tryptophan $ (whole brain) [59] |
| Behavioural | ↑ memory impairment [60] * ↑ mood-lowering [61] ↑ mood-lowering with pre-existing aggressive traits [62] ↑ impairment of the recognition of fearful facial expressions [63] |
| Reference | Sample Size | Health/Condition | Assessment Methods | Major Findings Between Sexes (♀ vs. ♂) |
|---|---|---|---|---|
| Bano et al. (2004) [83] | N= 16 (Did not specify the sample size of each sex) Randomised controlled clinical trial | Depressed patients treated with Prozac # for four weeks | Fluorometric method on blood samples |
|
| Pivac et al. (2004) [84] | Healthy controls: N = 233 (123 women and 110 men) Alcohol dependency patients: N = 190 (42 women and 148 men) | Drug-free patients with AUD | Fluorimetric method on platelet-rich-blood samples |
|
| Badawy et al. (2005) [42] | Healthy controls: N= 42 (16 females and 26 males) CFS patients: N= 23 (14 females and 9 males) | CFS patients | Fluorimetric method on blood samples |
|
| Booij et al. (2005) [85] | N = 21 (10 women and 11 men) Double-blind cross-over approach | Remitted depression participant’s in ATD condition | Psychiatric symptoms and behavioural measures of depression |
|
| Moreno et al. (2006) [86] | N = 59 (41 women and 18 men) | Remitted depressive patients in ATD condition | Behavioural measures of depressive symptoms |
|
| Weaver et al. (2010) [87] | Healthy controls: N= 16 (10 women * and 6 men) CFS patients: N= 22 (15 women* and 7 men) | CFS participants in Tryp enhancement condition | HPLC on blood samples |
|
| Frey et al. (2010) [88] | N= 25 (13 women * and 12 men) | MDD patients | PET with α-Methyl-L-tryptophan $ |
|
| Kötting et al. (2013) [89] | N = 20 (10 girls and 10 boys) Double-blind within-subject cross-over approach | ADHD participants in control or ATD condition | Behavioural measures of aggression 2.5 h after administration |
|
| Young et al. (2014) [90] | Ecstasy user participants: N = 13 (7 women * and 6 men) Ecstasy non-user participants: N = 17 (9 women* and 8 men) | Participants who had used ecstasy at least 25 times in control or ATD condition | Behavioural measures of mood states and impulsivity |
|
| Booij et al. (2014) [91] | Healthy controls: N= 18 (9 women * and 9 men) Ecstasy polydrug users: N = 17 (8 women * and 9 men) | Ecstasy polydrug users | HPLC on blood samples PET with α-Methyl-L-tryptophan $ |
|
| Platzer et al. (2017) [92] | Healthy participants: N = 93(57 women and 37 men) Bipolar patients: N =68 (26 women and 42 men) | Bipolar patients | UPLC on blood samples |
|
| Hestad et al. (2017) [45] | Healthy participants: N = 31(22 women and 9 men) Depression patients: N =44 (23 women and 21 men) | Depressed patients | HPLC on blood and cerebrospinal fluid |
|
| Zhou et al. (2018) [93] | Healthy participants: N = 72(31 women and 41 men) MDD patients: N =146 (70 women and 76 men) | MDD patients | HPLC-MS/MS on blood samples Behavioural measures of depressive symptoms, cognition, and memory |
|
| Vidal et al. (2020) [44] | Healthy controls: N= 80 (40 women and 40 men) AUD patients: N= 130 (35 women and 95 men) | AUD patients with a high prevalence of psychiatric comorbidity | HPLC on blood samples |
|
| Reininghaus et al. (2019) [94] | N= 426 (242 women and 184 men) | MDD patients over a 6-week rehabilitation treatment | HPLC on blood samples |
|
| Xu et al. (2021) [95] | N = 695 participants (377 women and 318 men) | AD patients | Metabolomics analysis over 665 plasma metabolites on blood samples |
|
| Cheng et al. (2023) [96] | N = 27 (10 women and 17 men) | Methamphetamine addicts | UHPLC–MS/MS on blood samples |
|
| Path | Condition | Sexual Dimorphisms (♀ vs. ♂) | |
|---|---|---|---|
| Peripherical levels | Tryp | MDD | ↓ plasma Tryp levels [45,83] ↓ plasma Tryp levels [94] |
| CFS | (=) plasma Tryp levels [42] (=) plasma Tryp levels [87] * (=) plasma Tryp and Kyn levels (Tryp supplementation condition) [87] * | ||
| Substance/drug abuse | (=) plasma Tryp levels [44,91] (≠) shift on plasma Tryp levels relative to controls (↓ on ♂; (=) on ♀) [96] | ||
| AD | ↓ plasma Tryp levels [95] | ||
| 5-HT | Substance/drug abuse | ↓ platelet 5-HT levels [84] (=) plasma 5-HT levels [44] | |
| Kyn | CFS | ↑ plasma Kyn levels [42] (=) plasma Kyn levels [87] * | |
| Bipolar disorder | ↓ plasma Kyn and KYNA levels [92] | ||
| MDD | (=) plasma Kyn levels [45] | ||
| Substance/drug abuse | ↑ plasma Kyn levels [44] ↓plasma KYNA levels [44] | ||
| AD | ↑ plasma Kyn levels [95] | ||
| Brain levels | Tryp | MDD | ↓ cerebrospinal fluid Tryp levels [45] ↑ depressive response (ATD condition) [85,86] ↑uptake tracer of α-Methyl-L-tryptophan $ (prefrontal cortex and limbic system) [88] * |
| ADHD | ↑ relative risk of presenting with increased aggression after a low level of provocation (ATD condition) [89] | ||
| Substance/drug abuse | ↑uptake tracer of α-Methyl-L-tryptophan$ (frontal regions) [91] * (≠) mood reduction relative to controls (↓ only on ♀) (ATD condition) [90] * |
| Condition | Sex-Bias on Incidence (♀:♂) | Dysfunctions of Tryp Metabolism | Tryp metabolism and Disease Severity | Sexual Dimorphisms and Dysfunctions of Tryp Metabolism | Reference |
|---|---|---|---|---|---|
| CFS | 4:1 | No reported dysfunctions | - | No reported sexual dimorphisms | [44,48,112,113] |
| Depression and related disorders | 2:1 | ↓Circulating Tryp ↓Brain 5-HT and 5-HIAA ↑Circulating Kyn IDO ↑Brain QUIN | Tryp depletion and lowering of brain 5-HT were associated with transient depressive symptoms. Elevated Kyn, IDO and QUIN levels were associated with depressive symptoms. | ↓Circulating Tryp (♀ vs. ♂) ↑Depressive response to Tryp depletion (♀ vs. ♂) | [1,9,10,19,25,26,45,83,85,86,94,110,111,116] |
| AD | 2:1 | ↓Circulating Tryp and 5-HT ↑Circulating Kyn ↑Brain kynurenine, IDO and QUIN | Neurodegeneration biomarkers, such as amyloid-β, senile plaques and neurofilament light chain, were positively correlated with the plasma Kyn/Tryp ratio and IDO levels. | ↓Circulating Tryp (♀ vs. ♂) ↑Circulating Kyn (♀ vs. ♂) | [9,10,58,95,103,115] |
| HD | 1:1 | ↑Brain Kyn and QUIN | Symptom severity was associated with plasma Kyn/Tryp ratio. QUIN induced behavioural and motor deficits in Huntingtin models. | No data | [18,20,92] |
| Bipolar Disorder | 1:1 | ↓Circulating Tryp and KYNA ↑Circulating Kyn | Plasma Kyn/Tryp ratio was associated with depressive severity while trending toward a negative association with mania symptoms. | ↓Circulating Tryp and KYNA (♀ vs. ♂) ↑Circulating Kyn (♂ vs. ♀) | [120,121] |
| OCD | 1:1 | ↑Brain α-[11C]methyl -L-tryptophan$ uptake | Symptom severity correlated positively with uptake from the caudate and temporal lobe. | No data | [97] |
| Epilepsy | 1:1 | ↑Brain α-[11C]methyl -L-tryptophan$ uptake, serotonin immunoreactivity, 5-HIAA and QUIN ↑Circulating 3HK and QUIN | Brain α-[11C]methyl-L-tryptophan $ uptake was correlated with seizure onset. | No data | [10,98] |
| Schizophrenia | 1:1.4 | ↑Brain and cerebrospinal fluid KYNA | Increased KYNA levels were correlated with cognitive impairments. | ↑Circulating KYNA (♂ vs. ♀) | [9,10,122] |
| Substance/ drug abuse disorder | 1:2 | ↑Circulating Kyn | Reduced Tryp availability and deficient 5-HT synthesis as higher serum levels of Kyn had been associated with increased consumption and vulnerability to addiction. | ↑Circulating Kyn (♀ vs. ♂) ↓Circulating KYNA (♀ vs. ♂)Incoherent results on 5-HT | [44,84,96,98,123,124] |
| ADHD | 1:2.3 | ↓ 3HK ↑Circulating Kyn, KYNA and 5-HT | Kyn levels were positively correlated to the pathological conditions in a mouse model of ADHD. | ↑Relative risk for aggressive behaviour during Tryp depletion (♀ vs. ♂) | [10,27,89,99,125] |
| ASD | 1:3 | ↓Circulating KYNA ↑Circulating 5-HT, Kyn and QUIN | Decreased central 5-HT associates with increased aggression. Tryp depletion exacerbates ASD symptoms, including stereotypies and repetitive behaviours. | ↑Circulating 5-HT (♂ vs. ♀) Platelet 5-HT content was inversely correlated to the score for intellectual ability (♂ vs. ♀) | [15,126,127] |
| PD | 1:3.5 | ↓Circulating KYNA ↑Brain IDO and QUIN | Kyn inhibitors significantly reduce the severity of dystonia in animal models. | No data | [10,14] |
| Database | Search Formula |
|---|---|
| Medline (via PubMed) | ((((“tryptophan” [Mesh] AND “Sex Characteristics” [Mesh]) OR (“tryptophan” [title] AND “gender” [All fields]) OR (“tryptophan” [title] AND “sex” [All fields])) NOT (“review” [Publication Type])) NOT (“systematic review” [Publication Type])) NOT (“meta analysis” [Publication Type]) |
| Scopus | TITLE-ABS-KEY ((“tryptophan” AND “Sex Characteristics”) OR (“tryptophan” AND “gender”) OR (“tryptophan” AND “sex”)) AND (LIMIT-TO(DOCTYPE, “ar”)) |
| Web of Science Medline | (TS= (“tryptophan” AND (“Sex Characteristics” OR “gender” OR “sex”))) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pais, M.L.; Martins, J.; Castelo-Branco, M.; Gonçalves, J. Sex Differences in Tryptophan Metabolism: A Systematic Review Focused on Neuropsychiatric Disorders. Int. J. Mol. Sci. 2023, 24, 6010. https://doi.org/10.3390/ijms24066010
Pais ML, Martins J, Castelo-Branco M, Gonçalves J. Sex Differences in Tryptophan Metabolism: A Systematic Review Focused on Neuropsychiatric Disorders. International Journal of Molecular Sciences. 2023; 24(6):6010. https://doi.org/10.3390/ijms24066010
Chicago/Turabian StylePais, Mariana Lapo, João Martins, Miguel Castelo-Branco, and Joana Gonçalves. 2023. "Sex Differences in Tryptophan Metabolism: A Systematic Review Focused on Neuropsychiatric Disorders" International Journal of Molecular Sciences 24, no. 6: 6010. https://doi.org/10.3390/ijms24066010
APA StylePais, M. L., Martins, J., Castelo-Branco, M., & Gonçalves, J. (2023). Sex Differences in Tryptophan Metabolism: A Systematic Review Focused on Neuropsychiatric Disorders. International Journal of Molecular Sciences, 24(6), 6010. https://doi.org/10.3390/ijms24066010