
Modern Microscopic Approaches to Astrocytes


Abstract
1. Introduction
2. Structural Studies of Astrocytes by Modern Microscopy
2.1. Discovery of Astrocytes
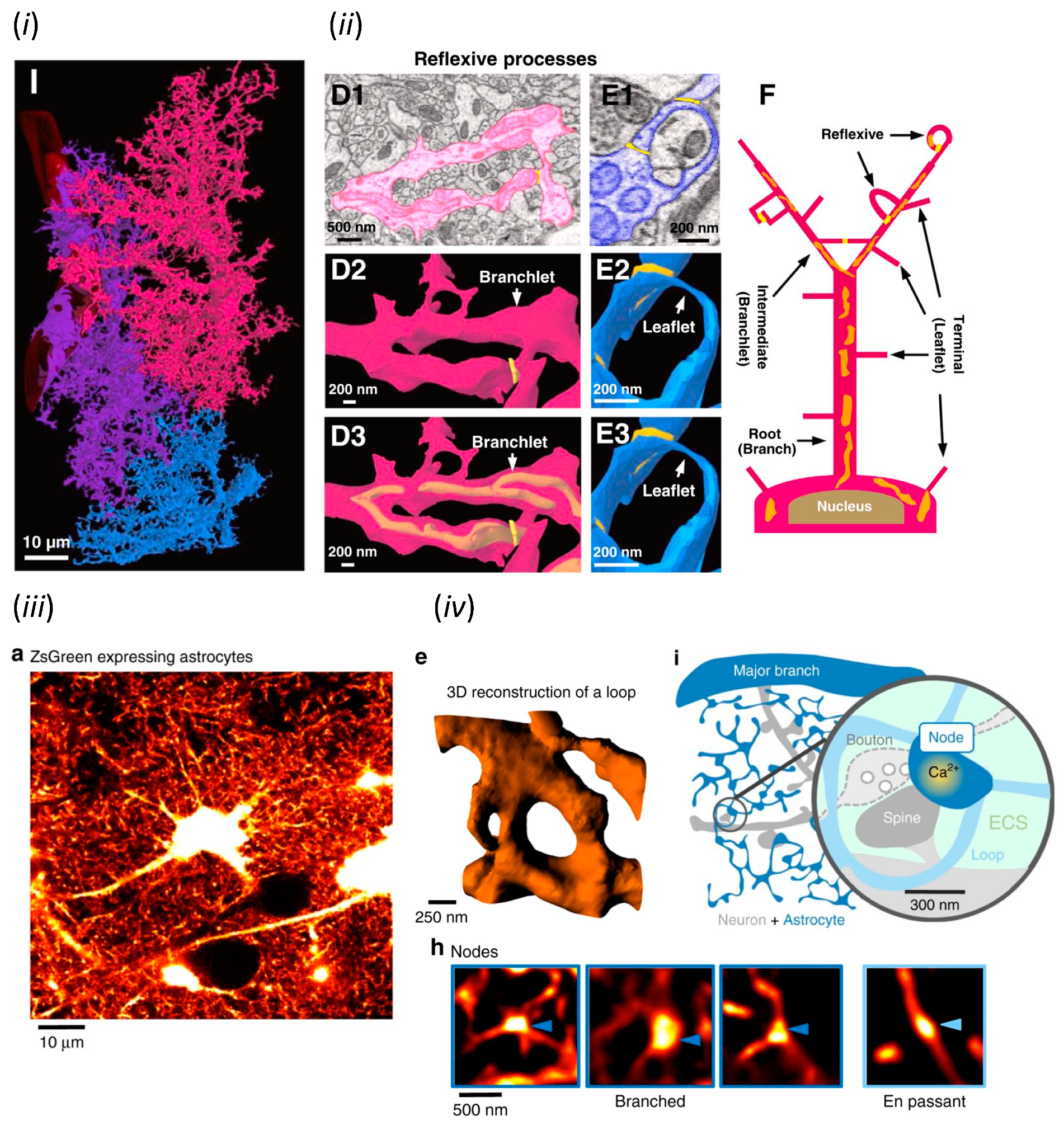
2.2. Electron Microscopic Analysis of Astrocytes
2.3. Analysis of Astrocytic Nanoscale Structures by Light Microscopy
2.4. Analysis of Astrocytic Domains by Multicolor Imaging
3. Physiological Studies of Astrocytes by Modern Microscopy
3.1. Ca2+ Imaging of Astrocytes
3.2. In Vivo Ca2+ Imaging of Astrocytes
3.3. Optical Manipulations of Astrocytes
4. Imaging of Biomolecules and Water in Astrocytes by Modern Microscopy
5. Conclusions
Funding
Conflicts of Interest
References
- Chvátal, A.; Verkhratsky, A. An Early History of Neuroglial Research: Personalities. Neuroglia 2018, 1, 245–257. [Google Scholar] [CrossRef]
- Ramón y Cajal, S. Histology of the Nervous System of Man and Vertebrates; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Lanjakornsiripan, D.; Pior, B.-J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-specific morphological and molecular differences in neocortical astrocytes and their dependence on neuronal layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef]
- Zhou, B.; Zuo, Y.-X.; Jiang, R.-T. Astrocyte morphology: Diversity, plasticity, and role in neurological diseases. CNS Neurosci. Ther. 2019, 25, 665–673. [Google Scholar] [CrossRef]
- Luse, S.A. Electron microscopic observations of the central nervous system. J. Biophys. Biochem. Cytol. 1956, 2, 531–542. [Google Scholar] [CrossRef]
- Schultz, R.L.; Maynard, E.A.; Pease, D.C. Electron microscopy of neurons and neuroglia of cerebral cortex and corpus callosum. Am. J. Anat. 1957, 100, 369–407. [Google Scholar] [CrossRef]
- Aten, S.; Kiyoshi, C.M.; Arzola, E.P.; Patterson, J.A.; Taylor, A.T.; Du, Y.; Guiher, A.M.; Philip, M.; Camacho, E.G.; Mediratta, D.; et al. Ultrastructural view of astrocyte arborization, astrocyte-astrocyte and astrocyte-synapse contacts, intracellular vesicle-like structures, and mitochondrial network. Prog. Neurobiol. 2022, 213, 102264. [Google Scholar] [CrossRef]
- Goggin, P.; Ho EM, L.; Gnaegi, H.; Searle, S.; Oreffo RO, C.; Schneider, P. Development of protocols for the first serial block-face scanning electron microscopy (SBF SEM) studies of bone tissue. Bone 2020, 131, 115107. [Google Scholar] [CrossRef]
- Arizono, M.; Inavalli, V.V.G.K.; Panatier, A.; Pfeiffer, T.; Angibaud, J.; Levet, F.; Ter Veer, M.J.T.; Stobart, J.; Bellocchio, L.; Mikoshiba, K.; et al. Structural basis of astrocytic Ca2+ signals at tripartite synapses. Nat. Commun. 2020, 11, 1906. [Google Scholar] [CrossRef]
- Henneberger, C.; Bard, L.; Panatier, A.; Reynolds, J.P.; Kopach, O.; Medvedev, N.I.; Minge, D.; Herde, M.K.; Anders, S.; Kraev, I.; et al. LTP Induction Boosts Glutamate Spillover by Driving Withdrawal of Perisynaptic Astroglia. Neuron 2020, 108, 919–936.e11. [Google Scholar] [CrossRef]
- Vicidomini, G.; Bianchini, P.; Diaspro, A. STED super-resolved microscopy. Nat. Methods 2018, 15, 173–182. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Livet, J.; Weissman, T.A.; Kang, H.; Draft, R.W.; Lu, J.; Bennis, R.A.; Sanes, J.R.; Lichtman, J.W. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature 2007, 450, 56–62. [Google Scholar] [CrossRef]
- Abdeladim, L.; Matho, K.S.; Clavreul, S.; Mahou, P.; Sintes, J.-M.; Solinas, X.; Arganda-Carreras, I.; Turney, S.G.; Lichtman, J.W.; Chessel, A.; et al. Multicolor multiscale brain imaging with chromatic multiphoton serial microscopy. Nat. Commun. 2019, 10, 1662. [Google Scholar] [CrossRef]
- Clavreul, S.; Abdeladim, L.; Hernández-Garzón, E.; Niculescu, D.; Durand, J.; Ieng, S.-H.; Barry, R.; Bonvento, G.; Beaurepaire, E.; Livet, J.; et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nat. Commun. 2019, 10, 4884. [Google Scholar] [CrossRef]
- Grienberger, C.; Konnerth, A. Imaging calcium in neurons. Neuron 2012, 73, 862–885. [Google Scholar] [CrossRef]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef]
- Sugino, H.; Ogura, A.; Kudo, Y.; Amano, T. Intracellular Ca2+ elevation induced by a neurotransmitter in a glial cell clone. Brain Res. 1984, 322, 127–130. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Kettenmann, H. Calcium signalling in glial cells. Trends Neurosci. 1996, 19, 346–352. [Google Scholar] [CrossRef]
- Agronskaia, A.V.; Tertoolen, L.; Gerritsen, H.C. Fast fluorescence lifetime imaging of calcium in living cells. J. Biomed. Opt. 2004, 9, 1230–1237. [Google Scholar] [CrossRef]
- Wilms, C.D.; Schmidt, H.; Eilers, J. Quantitative two-photon Ca2+ imaging via fluorescence lifetime analysis. Cell Calcium. 2006, 40, 73–79. [Google Scholar] [CrossRef]
- King, C.M.; Bohmbach, K.; Minge, D.; Delekate, A.; Zheng, K.; Reynolds, J.; Rakers, C.; Zeug, A.; Petzold, G.C.; Rusakov, D.A.; et al. Local Resting Ca2+ Controls the Scale of Astroglial Ca2+ Signals. Cell Rep. 2020, 30, 3466–3477.e4. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa-Ankerhold, H.C.; Ankerhold, R.; Drummen, G.P.C. Advanced fluorescence microscopy techniques—FRAP, FLIP, FLAP, FRET and FLIM. Molecules 2012, 17, 4047–4132. [Google Scholar] [CrossRef]
- Köhler, S.; Winkler, U.; Junge, T.; Lippmann, K.; Eilers, J.; Hirrlinger, J. Gray and white matter astrocytes differ in basal metabolism but respond similarly to neuronal activity. Glia 2023, 71, 229–244. [Google Scholar] [CrossRef]
- Engels, M.; Kalia, M.; Rahmati, S.; Petersilie, L.; Kovermann, P.; van Putten, M.J.A.M.; Rose, C.R.; Meijer HG, E.; Gensch, T.; Fahlke, C. Glial Chloride Homeostasis Under Transient Ischemic Stress. Front. Cell. Neurosci. 2021, 15, 735300. [Google Scholar] [CrossRef]
- Requardt, R.P.; Hirrlinger, P.G.; Wilhelm, F.; Winkler, U.; Besser, S.; Hirrlinger, J. Ca2+ signals of astrocytes are modulated by the NAD+/NADH redox state. J. Neurochem. 2012, 120, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Higuchi, C.; Moto, T.; Kozuka, N.; Susuki, J.; Itofusa, R.; Yamashita, J.; Kudo, Y. Dual Regulation of Calcium Oscillation in Astrocytes by Growth Factors and Pro-Inflammatory Cytokines via the Mitogen-Activated Protein Kinase Cascade. J. Neurosci. 2003, 23, 10944–10952. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Maekawa, S.; Morita, M. Astrocyte calcium waves propagate proximally by gap junction and distally by extracellular diffusion of ATP released from volume-regulated anion channels. Sci. Rep. 2017, 7, 13115. [Google Scholar] [CrossRef]
- Porter, J.T.; McCarthy, K.D. Hippocampal astrocytes in situ respond to glutamate released from synaptic terminals. J. Neurosci. 1996, 16, 5073–5081. [Google Scholar] [CrossRef]
- Wang, X.; Lou, N.; Xu, Q.; Tian, G.F.; Peng, W.G.; Han, X.; Kang, J.; Takano, T.; Nedergaard, M. Astrocytic Ca2+ signaling evoked by sensory stimulation in vivo. Nat. Neurosci. 2006, 9, 816–823. [Google Scholar] [CrossRef]
- Bindocci, E.; Savtchouk, I.; Liaudet, N.; Becker, D.; Carriero, G.; Volterra, A. Three-dimensional Ca2+ imaging advances understanding of astrocyte biology. Science 2017, 356, eaai8185. [Google Scholar] [CrossRef]
- Streich, L.; Boffi, J.C.; Wang, L.; Alhalaseh, K.; Barbieri, M.; Rehm, R.; Deivasigamani, S.; Gross, C.T.; Agarwal, A.; Prevedel, R. High-resolution structural and functional deep brain imaging using adaptive optics three-photon microscopy. Nat. Methods 2021, 18, 1253–1258. [Google Scholar] [CrossRef]
- Gómez-Gaviro, M.V.; Sanderson, D.; Ripoll, J.; Desco, M. Biomedical Applications of Tissue Clearing and Three-Dimensional Imaging in Health and Disease. iScience 2020, 23, 101432. [Google Scholar] [CrossRef]
- Wang, K.; Sun, W.; Richie, C.T.; Harvey, B.K.; Betzig, E.; Ji, N. Direct wavefront sensing for high-resolution in vivo imaging in scattering tissue. Nat. Commun. 2015, 6, 7276. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Sun, S.; Lin, H.-Z.; Liu, W.-T.; Chen, P.-X. Imaging through scattering layers exceeding memory effect range with spatial-correlation-achieved point-spread-function. Opt. Lett. 2018, 43, 1670–1673. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.; Moro, D.H.; Mouffle, C.; Didienne, S.; Hepp, R.; Pfrieger, F.W.; Mangin, J.-M.; Legendre, P.; Martin, C.; Luquet, S.; et al. Mapping astrocyte activity domains by light sheet imaging and spatio-temporal correlation screening. Neuroimage 2002, 220, 117069. [Google Scholar] [CrossRef]
- Agarwal, A.; Wu, P.H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient Opening of the Mitochondrial Permeability Transition Pore Induces Microdomain Calcium Transients in Astrocyte Processes. Neuron 2017, 93, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.-H.; Noh, K.; Lee, B.H.; Barcelon, E.; Jun, S.B.; Park, H.Y.; Lee, S.J. Hippocampal astrocytes modulate anxiety-like behavior. Nat. Commun. 2022, 13, 6536. [Google Scholar] [CrossRef]
- Yu, X.; Taylor, A.M.W.; Nagai, J.; Golshani, P.; Evans, C.J.; Coppola, G.; Khakh, B.S. Reducing Astrocyte Calcium Signaling In Vivo Alters Striatal Microcircuits and Causes Repetitive Behavior. Neuron 2018, 99, 1170–1187.e9. [Google Scholar] [CrossRef]
- Pittolo, S.; Yokoyama, S.; Willoughby, D.D.; Taylor, C.R.; Reitman, M.E.; Tse, V.; Wu, Z.; Etchenique, R.; Li, Y.; Poskanzer, K.E. Dopamine activates astrocytes in prefrontal cortex via α1-adrenergic receptors. Cell Rep. 2022, 40, 111426. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.-F.; Lin, J.-Y.; Yeh, P.-T.; Hsu, K.-J.; Tsai, Y.-H.; Chen, S.-K.; Chu, S.-W. Dual GRIN lens two-photon endoscopy for high-speed volumetric and deep brain imaging. Biomed. Opt. Express. 2021, 12, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.M.; Resendez, S.L.; Chen, K.-S.; Favero, M.; Liang-Guallpa, J.; Nassi, J.J.; Neufeld, S.Q.; Visscher, K.; Ghosh, K.K. Miniature microscopes for manipulating and recording in vivo brain activity. Microscopy 2021, 70, 399–414. [Google Scholar] [CrossRef]
- Klioutchnikov, A.; Wallace, D.J.; Frosz, M.H.; Zeltner, R.; Sawinski, J.; Pawlak, V.; Voit, K.-M.; Russell, P.S.J.; Kerr, J.N.D. Three-photon head-mounted microscope for imaging deep cortical layers in freely moving rats. Nat. Methods 2020, 17, 509–513. [Google Scholar] [CrossRef]
- Rynes, M.L.; Surinach, D.A.; Linn, S.; Laroque, M.; Rajendran, V.; Dominguez, J.; Hadjistamoulou, O.; Navabi, Z.S.; Ghanbari, L.; Johnson, G.W.; et al. Miniaturized head-mounted microscope for whole-cortex mesoscale imaging in freely behaving mice. Nat. Methods 2021, 18, 417–425. [Google Scholar] [CrossRef]
- Monai, H.; Ohkura, M.; Tanaka, M.; Oe, Y.; Konno, A.; Hirai, H.; Mikoshiba, K.; Itohara, S.; Nakai, J.; Iwai, Y.; et al. Calcium imaging reveals glial involvement in transcranial direct current stimulation-induced plasticity in mouse brain. Nat. Commun. 2016, 7, 11100. [Google Scholar] [CrossRef]
- Ma, Y.; Shaik, M.A.; Kim, S.H.; Kozberg, M.G.; Thibodeaux, D.N.; Zhao, H.T.; Yu, H.; Hillman, E.M.C. Wide-field optical mapping of neural activity and brain haemodynamics: Considerations and novel approaches. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150360. [Google Scholar] [CrossRef] [PubMed]
- Couto, J.; Musall, S.; Sun, X.R.; Khanal, A.; Gluf, S.; Saxena, S.; Kinsella, I.; Abe, T.; Cunningham, J.P.; Paninski, L.; et al. Chronic, cortex-wide imaging of specific cell populations during behavior. Nat. Protoc. 2021, 16, 3241–3263. [Google Scholar] [CrossRef] [PubMed]
- Valley, M.T.; Moore, M.G.; Zhuang, J.; Mesa, N.; Castelli, D.; Sullivan, D.; Reimers, M.; Waters, J. Separation of hemodynamic signals from GCaMP fluorescence measured with wide-field imaging. J. Neurophysiol. 2020, 123, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Komiyama, T. Characterizing Cortex-Wide Dynamics with Wide-Field Calcium Imaging. J. Neurosci. 2021, 41, 4160–4168. [Google Scholar] [CrossRef]
- Liu, Q.S.; Xu, Q.; Arcuino, G.; Kang, J.; Nedergaard, M. Astrocyte-mediated activation of neuronal kainate receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 3172–3177. [Google Scholar] [CrossRef]
- Gordon, G.R.J.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.R.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–749. [Google Scholar] [CrossRef]
- Perea, G.; Yang, A.; Boyden, E.S.; Sur, M. Optogenetic astrocyte activation modulates response selectivity of visual cortex neurons in vivo. Nat. Commun. 2014, 5, 3262. [Google Scholar] [CrossRef] [PubMed]
- Masamoto, K.; Unekawa, M.; Watanabe, T.; Toriumi, H.; Takuwa, H.; Kawaguchi, H.; Kanno, I.; Matsui, K.; Tanaka, K.F.; Tomita, Y.; et al. Unveiling astrocytic control of cerebral blood flow with optogenetics. Sci. Rep. 2015, 5, 11455. [Google Scholar] [CrossRef] [PubMed]
- Pettit, D.L.; Wang, S.S.; Gee, K.R.; Augustine, G.J. Chemical two-photon uncaging: A novel approach to mapping glutamate receptors. Neuron 1997, 19, 465–471. [Google Scholar] [CrossRef]
- Liang, C.W.; Mohammadi, M.; Santos, M.D.; Tang, C.-M. Patterned photostimulation with digital micromirror devices to investigate dendritic integration across branch points. J. Vis. Exp. 2011, 49, e2003. [Google Scholar] [CrossRef]
- Pégard, N.C.; Mardinly, A.R.; Oldenburg, I.A.; Sridharan, S.; Waller, L.; Adesnik, H. Three-dimensional scanless holographic optogenetics with temporal focusing (3D-SHOT). Nat. Commun. 2017, 8, 1228. [Google Scholar] [CrossRef]
- Xue, Y.; Waller, L.; Adesnik, H.; Pégard, N. Three-dimensional multi-site random access photostimulation (3D-MAP). Elife 2022, 11, e73266. [Google Scholar] [CrossRef]
- Carmi, I.; De Battista, M.; Maddalena, L.; Carroll, E.C.; Kienzler, M.A.; Berlin, S. Holographic two-photon activation for synthetic optogenetics. Nat. Protoc. 2019, 14, 864–900. [Google Scholar] [CrossRef]
- Okada, T.; Kato, D.; Nomura, Y.; Obata, N.; Quan, X.; Morinaga, A.; Yano, H.; Guo, Z.; Aoyama, Y.; Tachibana, Y.; et al. Pain induces stable, active microcircuits in the somatosensory cortex that provide a therapeutic target. Sci Adv. 2021, 337, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Ikeshima-Kataoka, H.; Kreft, M.; Vardjan, N.; Zorec, R.; Noda, M. Metabolic Plasticity of Astrocytes and Aging of the Brain. Int. J. Mol. Sci. 2019, 20, 941. [Google Scholar] [CrossRef]
- Zimmer, E.R.; Parent, M.J.; Souza, D.G.; Leuzy, A.; Lecrux, C.; Kim, H.-I.; Gauthier, S.; Pellerin, L.; Hamel, E.; Rosa-Neto, P. [18F]FDG PET signal is driven by astroglial glutamate transport. Nat. Neurosci. 2017, 20, 393–395. [Google Scholar] [CrossRef]
- Yao, J.; Xia, J.; Maslov, K.I.; Nasiriavanaki, M.; Tsytsarev, V.; Demchenko, A.V.; Wang, L.V. Noninvasive photoacoustic computed tomography of mouse brain metabolism in vivo. Neuroimage 2013, 64, 257–266. [Google Scholar] [CrossRef]
- Chuquet, J.; Quilichini, P.; Nimchinsky, E.A.; Buzsáki, G. Predominant enhancement of glucose uptake in astrocytes versus neurons during activation of the somatosensory cortex. J. Neurosci. 2010, 30, 15298–15303. [Google Scholar] [CrossRef]
- Lundgaard, I.; Li, B.; Xie, L.; Kang, H.; Sanggaard, S.; Haswell, J.D.; Sun, W.; Goldman, S.; Blekot, S.; Nielsen, M.; et al. Direct neuronal glucose uptake heralds activity-dependent increases in cerebral metabolism. Nat. Commun. 2015, 6, 6807. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.; Sugawara, I.; Harada, K.; Ito, M.; Takizawa, M.; Ishida, K.; Ueda, H.; Kitaguchi, T.; Tsuboi, T. Development of red genetically encoded biosensor for visualization of intracellular glucose dynamics. Cell Chem Biol. 2022, 29, 98–108.e4. [Google Scholar] [CrossRef]
- Bekdash, R.; Quejada, J.R.; Ueno, S.; Kawano, F.; Morikawa, K.; Klein, A.D.; Matsumoto, K.; Lee, T.C.; Nakanishi, K.; Chalan, A.; et al. GEM-IL: A highly responsive fluorescent lactate indicator. Cell. Rep. Methods 2021, 1, 100092. [Google Scholar] [CrossRef] [PubMed]
- Durkee, C.A.; Araque, A. Diversity and Specificity of Astrocyte-neuron Communication. Neuroscience 2019, 396, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Vandebroek, A.; Yasui, M. Regulation of AQP4 in the Central Nervous System. Int. J. Mol. Sci. 2020, 21, 1603. [Google Scholar] [CrossRef]
- Kojima, S.; Nakamura, T.; Nidaira, T.; Nakamura, K.; Ooashi, N.; Ito, E.; Watase, K.; Tanaka, K.; Wada, K.; Kudo, Y.; et al. Optical detection of synaptically induced glutamate transport in hippocampal slices. J. Neurosci. 1999, 19, 2580–2588. [Google Scholar] [CrossRef]
- Pál, I.; Kardos, J.; Dobolyi, Á.; Héja, L. Appearance of fast astrocytic component in voltage-sensitive dye imaging of neural activity. Mol. Brain 2015, 8, 35. [Google Scholar] [CrossRef]
- Zhang, Y.; Phillips, G.J.; Li, Q.; Yeung, E.S. Imaging localized astrocyte ATP release with firefly luciferase beads attached to the cell surface. Anal. Chem. 2008, 80, 9316–9325. [Google Scholar] [CrossRef]
- Yamashiro, K.; Fujii, Y.; Maekawa, S.; Morita, M. Multiple pathways for elevating extracellular adenosine in the rat hippocampal CA1 region characterized by adenosine sensor cells. J. Neurochem. 2016, 140, 24–36. [Google Scholar] [CrossRef]
- Wu, Z.; He, K.; Chen, Y.; Li, H.; Pan, S.; Li, B.; Liu, T.; Xi, F.; Deng, F.; Wang, H.; et al. A sensitive GRAB sensor for detecting extracellular ATP in vitro and in vivo. Neuron 2022, 110, 770–782.e5. [Google Scholar] [CrossRef]
- Bezzi, P.; Gundersen, V.; Galbete, J.L.; Seifert, G.; Steinhäuser, C.; Pilati, E.; Volterra, A. Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat. Neurosci. 2004, 7, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Shuttleworth, C.W. Use of NAD(P)H and flavoprotein autofluorescence transients to probe neuron and astrocyte responses to synaptic activation. Neurochem. Int. 2010, 56, 379–386. [Google Scholar] [CrossRef]
- Gomes da Costa, S.; Richter, A.; Schmidt, U.; Breuninger, S.; Hollricher, O. Confocal Raman microscopy in life sciences. Morphologie 2019, 103, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Palonpon, A.F.; Smith, N.I.; Chiu, L.; Kasai, A.; Hashimoto, H.; Kawata, S.; Fujita, K. Structured line illumination Raman microscopy. Nat. Commun. 2015, 6, 10095. [Google Scholar] [CrossRef]
- Palombo, F.; Tamagnini, F.; Jeynes, J.C.G.; Mattana, S.; Swift, I.; Nallala, J.; Hancock, J.; Brown, J.T.; Randall, A.D.; Stone, N. Detection of Aβ plaque-associated astrogliosis in Alzheimer’s disease brain by spectroscopic imaging and immunohistochemistry. Analyst 2018, 143, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Xu, J.; Brinkhof, B.; Wang, H.; Cui, Z.; Huang, W.E.; Ye, H. A single-cell Raman-based platform to identify developmental stages of human pluripotent stem cell-derived neurons. Proc. Natl. Acad. Sci. USA 2002, 117, 18412–18423. [Google Scholar] [CrossRef]
- Ibata, K.; Takimoto, S.; Morisaku, T.; Miyawaki, A.; Yasui, M. Analysis of Aquaporin-Mediated Diffusional Water Permeability by Coherent Anti-Stokes Raman Scattering Microscopy. Biophys. J. 2011, 101, 2277–2283. [Google Scholar] [CrossRef]
- Yu, Y.-C.; Sohma, Y.; Takimoto, S.; Miyauchi, T.; Yasui, M. Direct visualization and quantitative analysis of water diffusion in complex biological tissues using CARS microscopy. Sci. Rep. 2013, 3, 2745. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Category | Microscope | Achievements and Findings | References |
|---|---|---|---|
| Structure | Serial block-face scanning electron microscopy (SBF-SEM) | Reflexive process | Aten et al, 2022 [7] |
| Stimulated emission depletion (STED) microscopy | Structure of living PAP | Arizono et al, 2020 [9] | |
| Fluorescence recovery after photobleaching (FRAP) | Dynamics of living PAP | Henneberger et al, 2020 [10] | |
| Confocal microscopy and dye injection | Astrocytic domain | Bushong et al, 2002 [12] | |
| Chromatic multiphoton serial (ChroMS) microscopy | Developmental process forming astrocytic domain | Clavreul et al, 2019 [15] | |
| Physiology | Two photon fluorescence lifetime imaging microscopy (2P-FLIM) | Astocyte subpopulations of different resting Ca2+ level | King et al, 2020 [22] |
| Redox state and ATP concentration | Köhler et al, 2023 [24] | ||
| Cl- imaging | Engels et al, 2021 [25] | ||
| Three photon excitation with adaptive optics | Ca2+ imaging in subcortial structure | Streich et al, 2021 [31] | |
| Membrance-anchored GECI | Ca2+ imaging of PAP | Agarwal et al, 2013 [37] | |
| Two-photon endoscopy (GRIN lens) | Deep brain imaging (no astrocyte data) | Chien et al, 2021 [41] | |
| Head-mounted microscopy with three-photon excitation | Deep brain Ca2+ imagng of freely beehaving mouse (no astrocyte data) | Klioutchnikov et al, 2020 [43] | |
| Head-mounted microscopy with a low magnification objective | Wide field Ca2+ imaging of freely beehaving mouse (no astrocyte data) | Rynes et al, 2021 [44] | |
| Wide field Ca2+ imaging | Wide field Ca2+ imaging | Monai et al, 2016 [45] | |
| Furthermore, three-dimensional multi-site random access photostimulation (3D-MAP) | Target cell optogenetic stimulation (no astrocyte data) | Xue et al, 2022 [57] | |
| Two-photon Computer generated holography (CGH) | Target cell optogenetic stimulation (no astrocyte data) | Carmi et al, 2019 [58] | |
| Biomolecule | GFP-based biosensor | In vivo ATP imaging | Wu et al, 2022 [73] |
| Raman microscopy | Myelin imaging | Gomes et al, 2019 [76] | |
| Beta-amyloid imaging | Watanabe et al, 2015 [77] | ||
| Water flow imaging | Yu et al, 2014 [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morita, M. Modern Microscopic Approaches to Astrocytes. Int. J. Mol. Sci. 2023, 24, 5883. https://doi.org/10.3390/ijms24065883
Morita M. Modern Microscopic Approaches to Astrocytes. International Journal of Molecular Sciences. 2023; 24(6):5883. https://doi.org/10.3390/ijms24065883
Chicago/Turabian StyleMorita, Mitsuhiro. 2023. "Modern Microscopic Approaches to Astrocytes" International Journal of Molecular Sciences 24, no. 6: 5883. https://doi.org/10.3390/ijms24065883
APA StyleMorita, M. (2023). Modern Microscopic Approaches to Astrocytes. International Journal of Molecular Sciences, 24(6), 5883. https://doi.org/10.3390/ijms24065883