
Overexpression and Role of HHLA2, a Novel Immune Checkpoint, in Colorectal Cancer

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Results
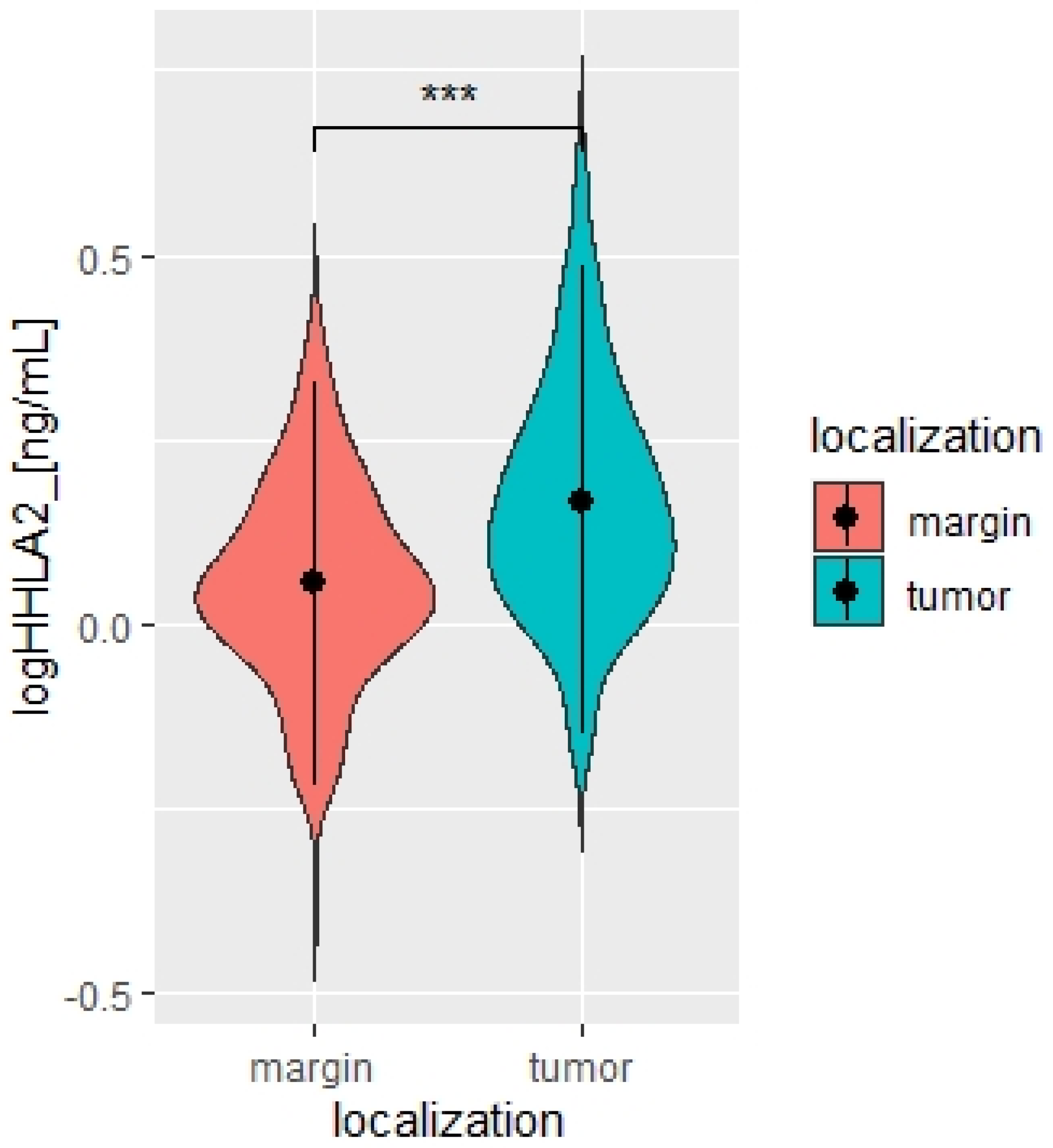
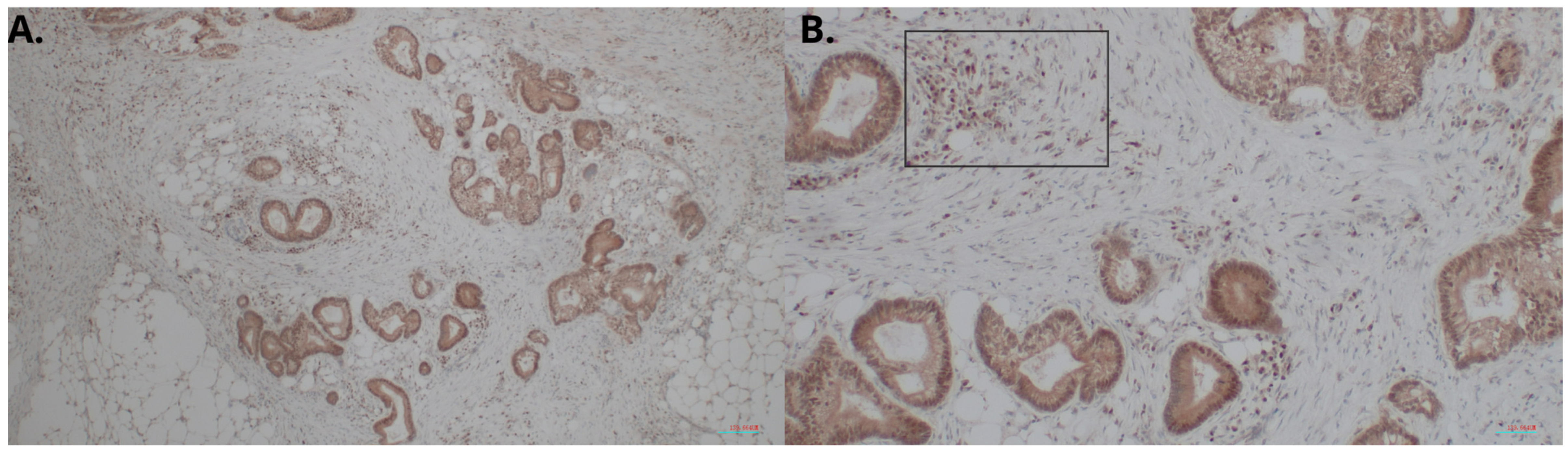
2.1. Overexpression of HHLA2 in CRC Tissues
2.2. The Expression of HHLA2 in Relation to MSI/MSS Status and Clinicopathological Parameters
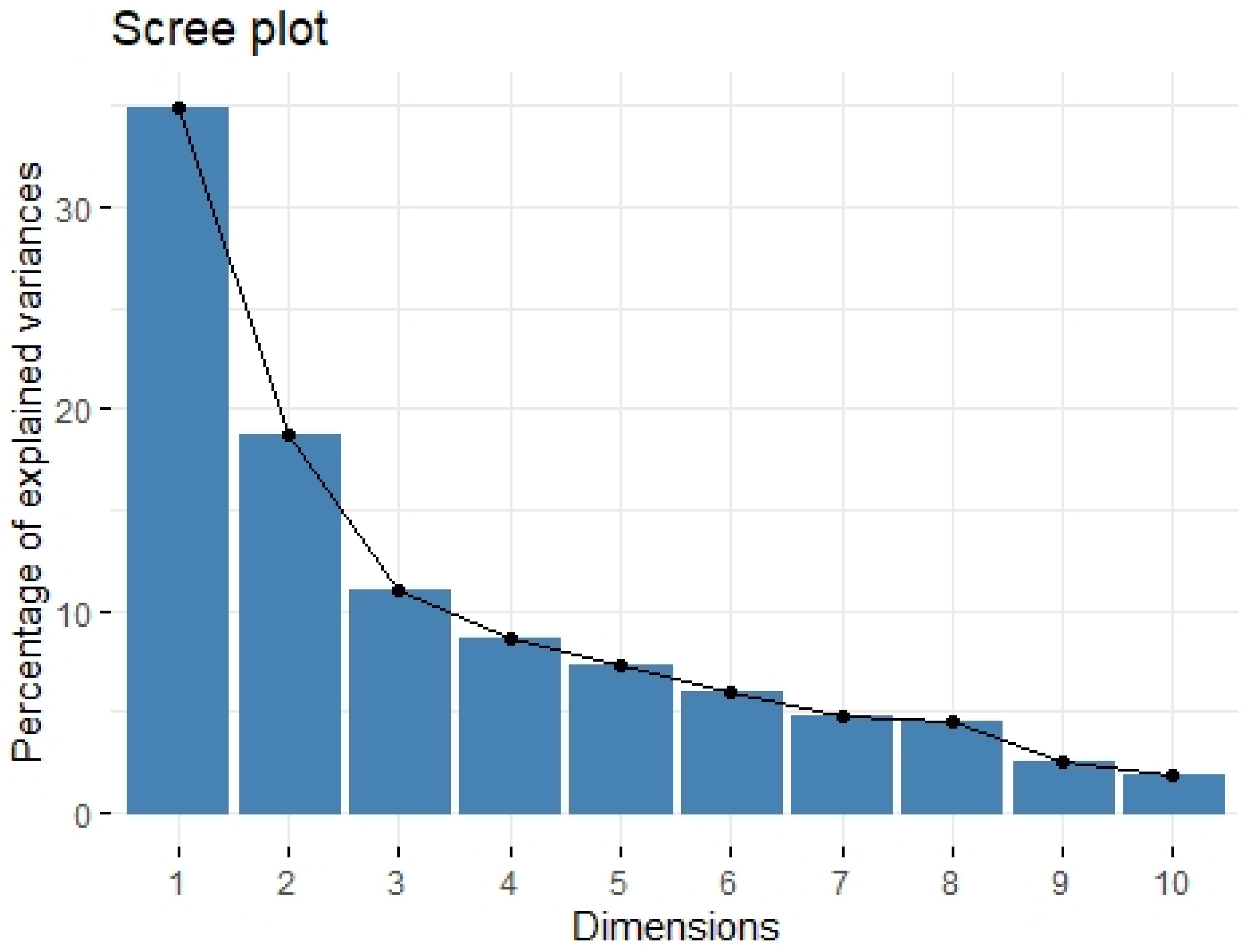
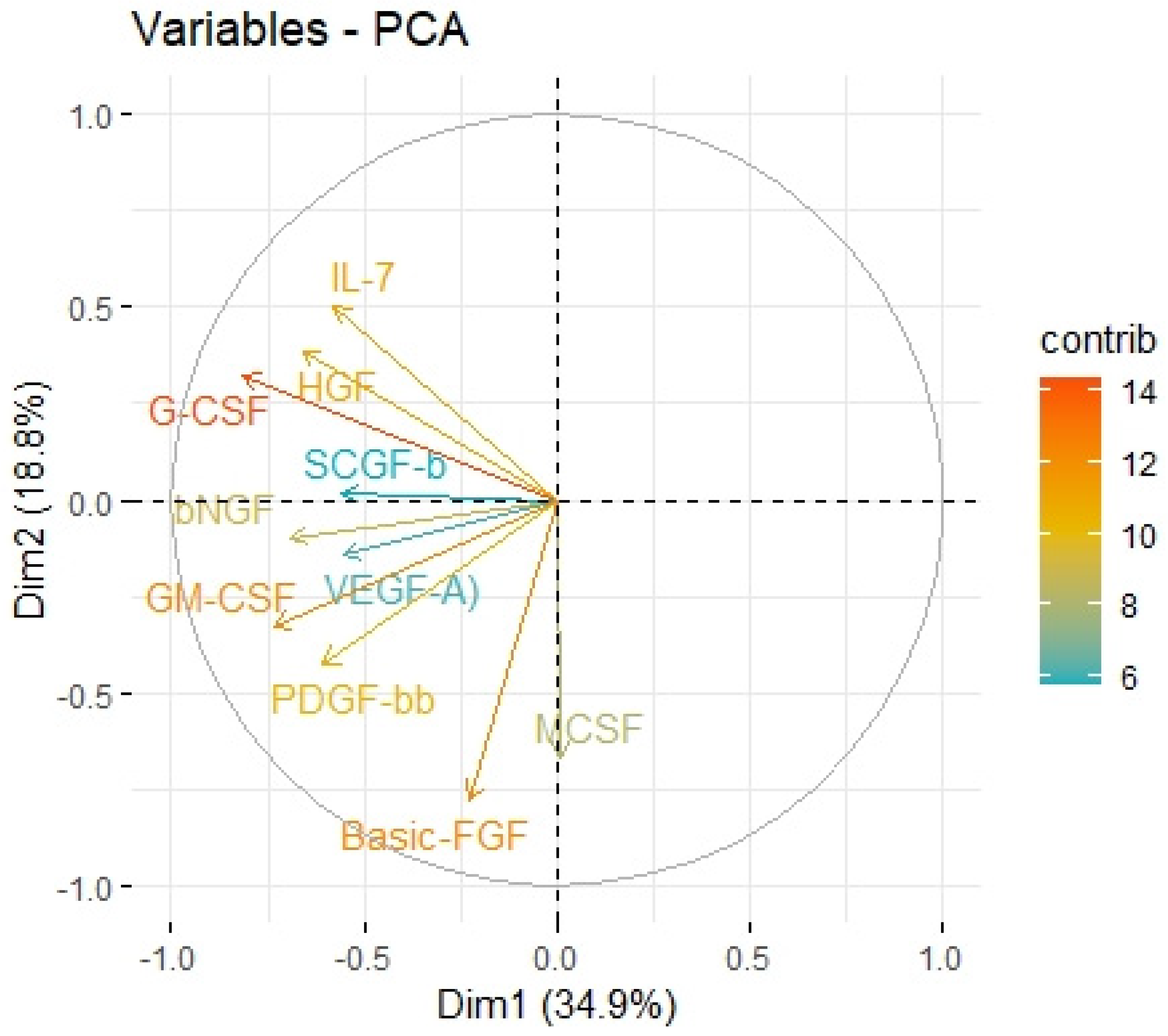
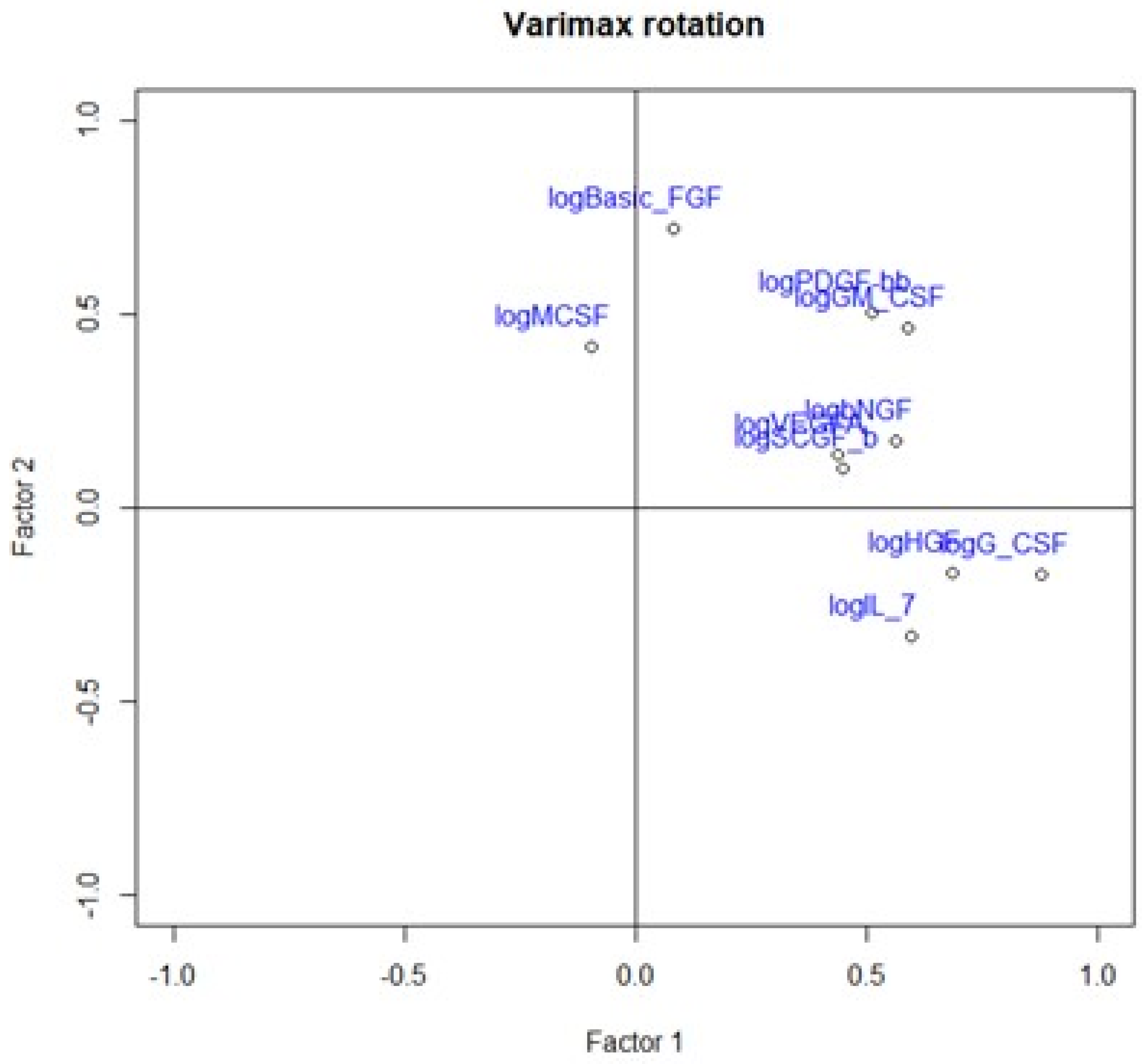
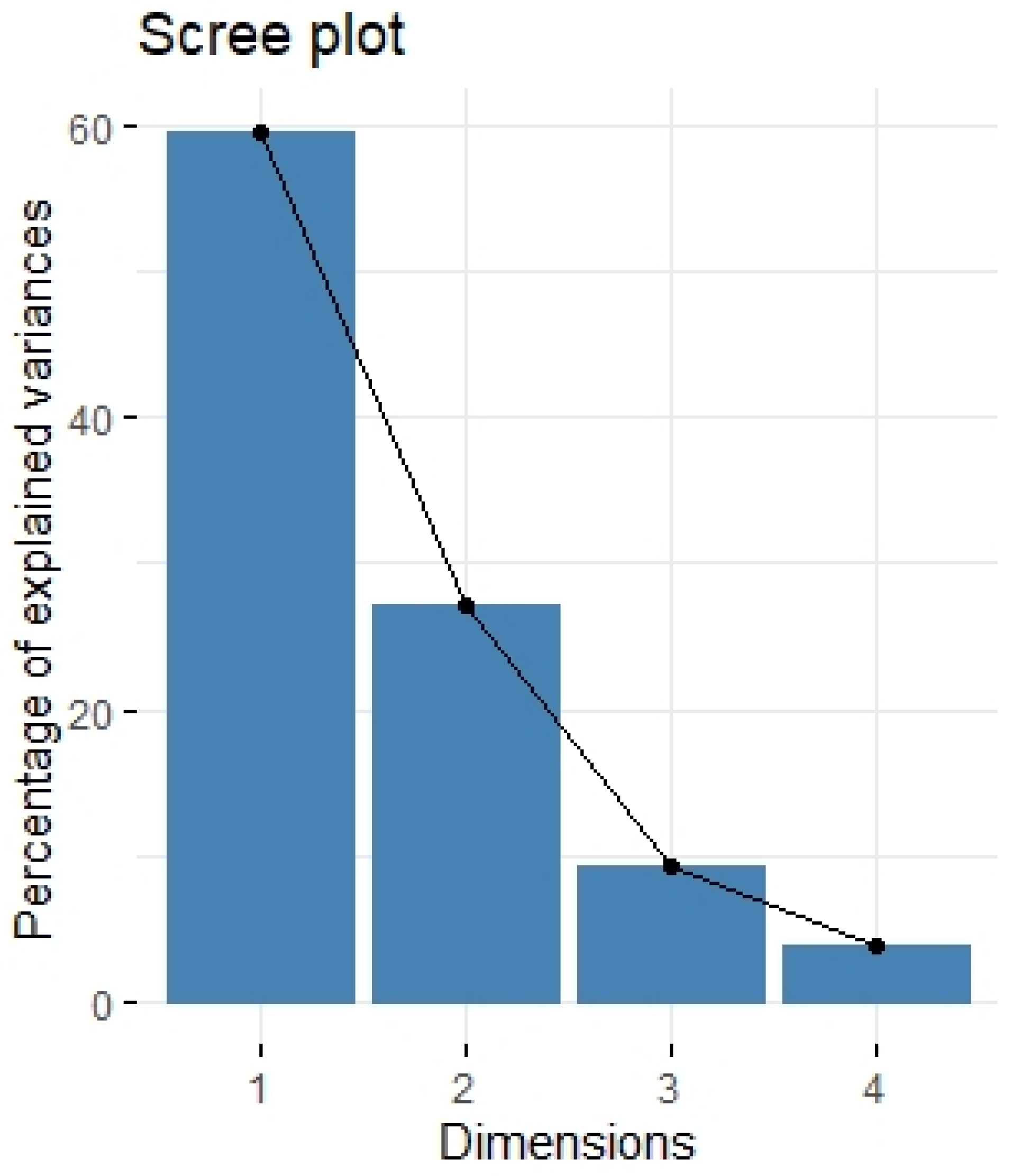
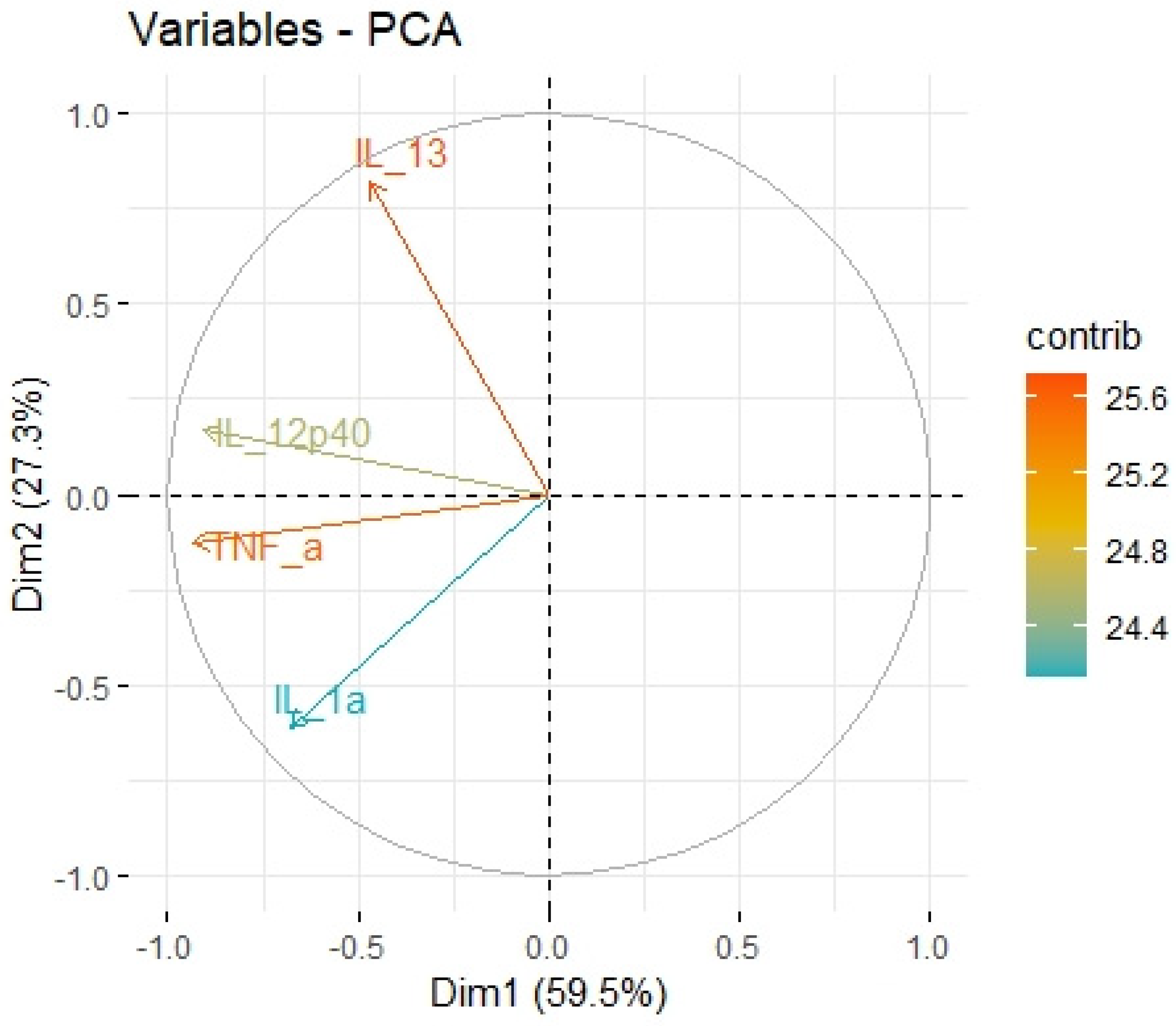
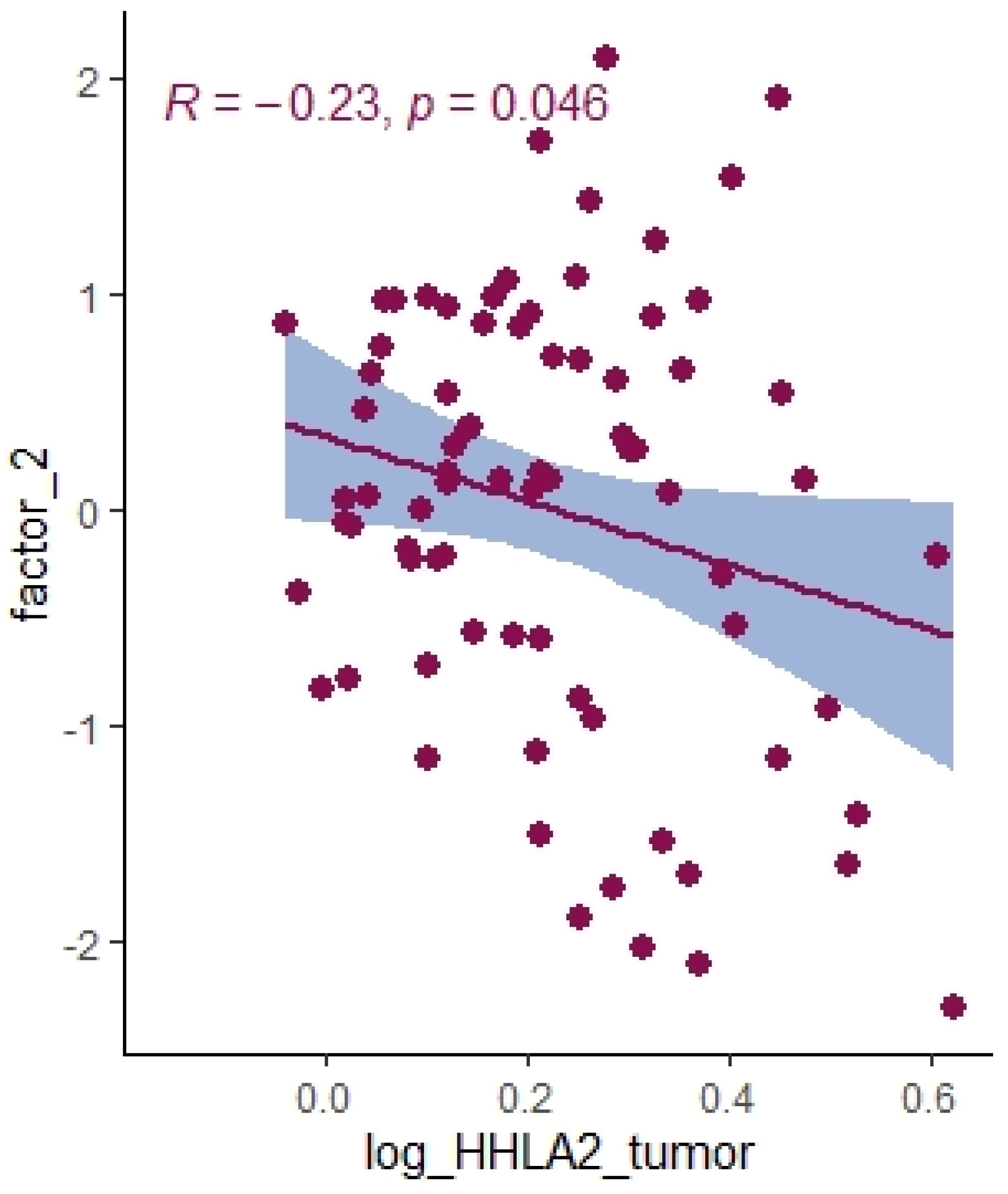
2.3. Principal Component Analysis
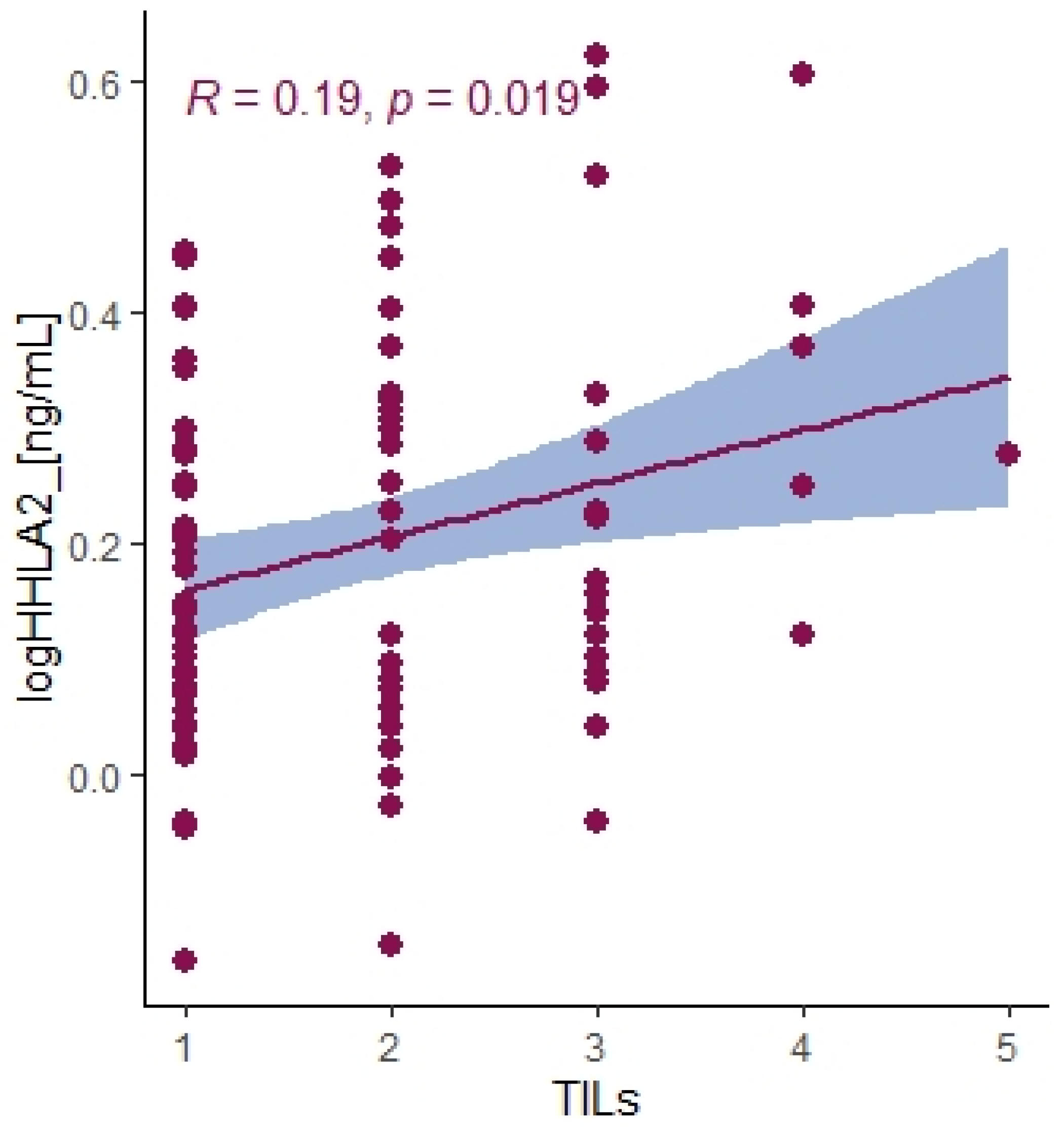
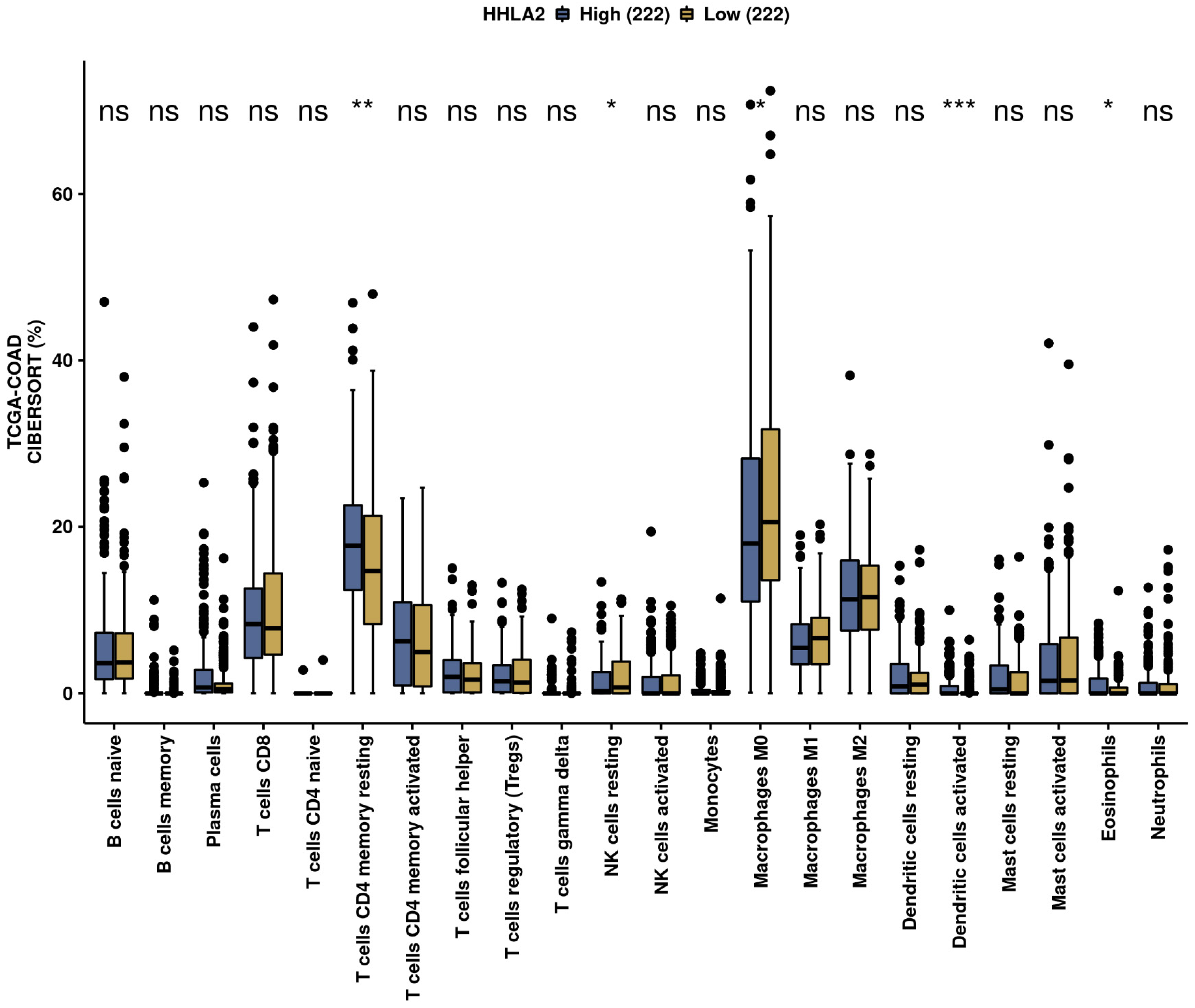
2.4. Immune Infiltration Landscape of HHLA2 Based on the Cancer Genome Atlas-Colon Adenocarcinoma (TCGA-COAD) Data
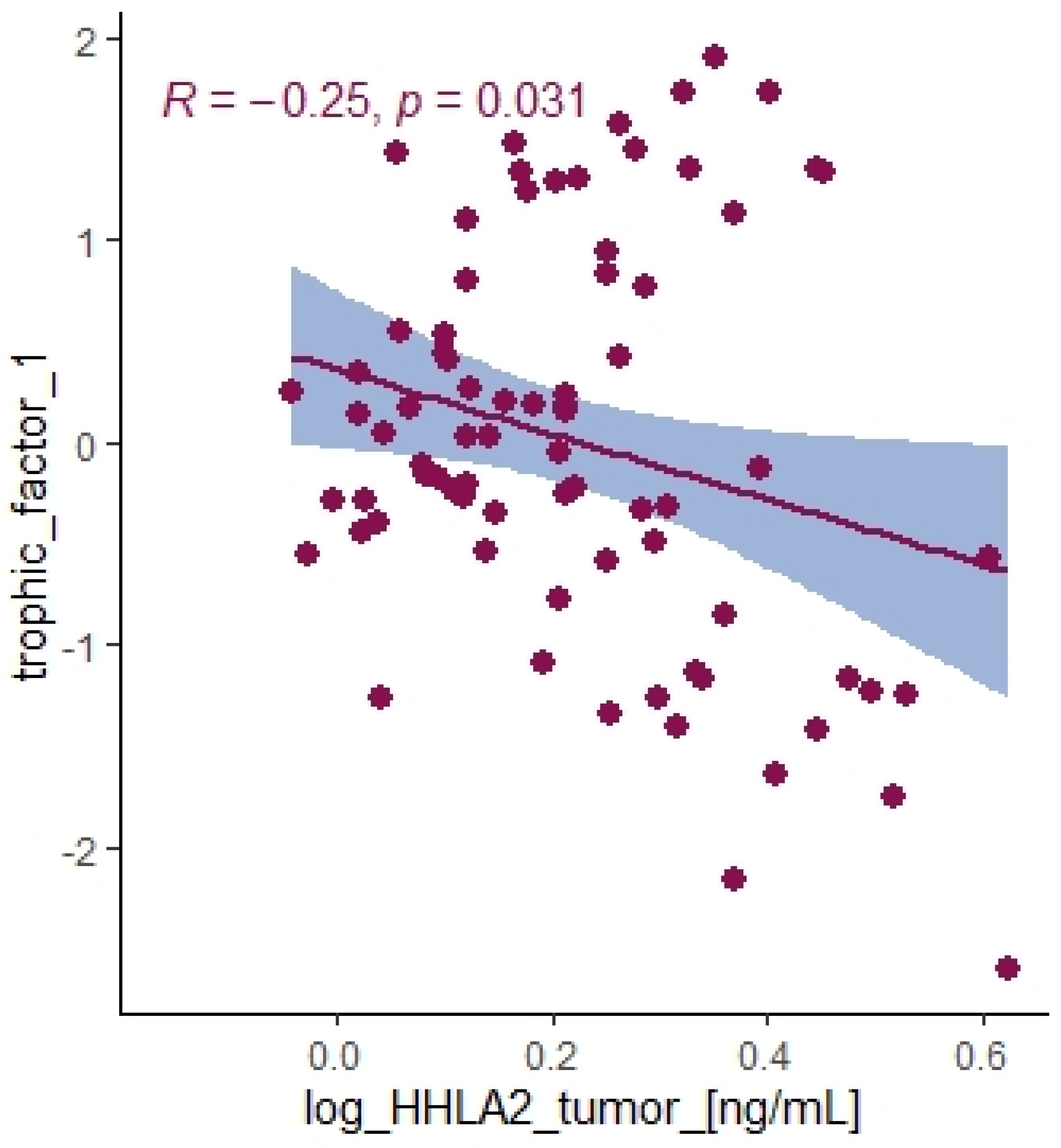
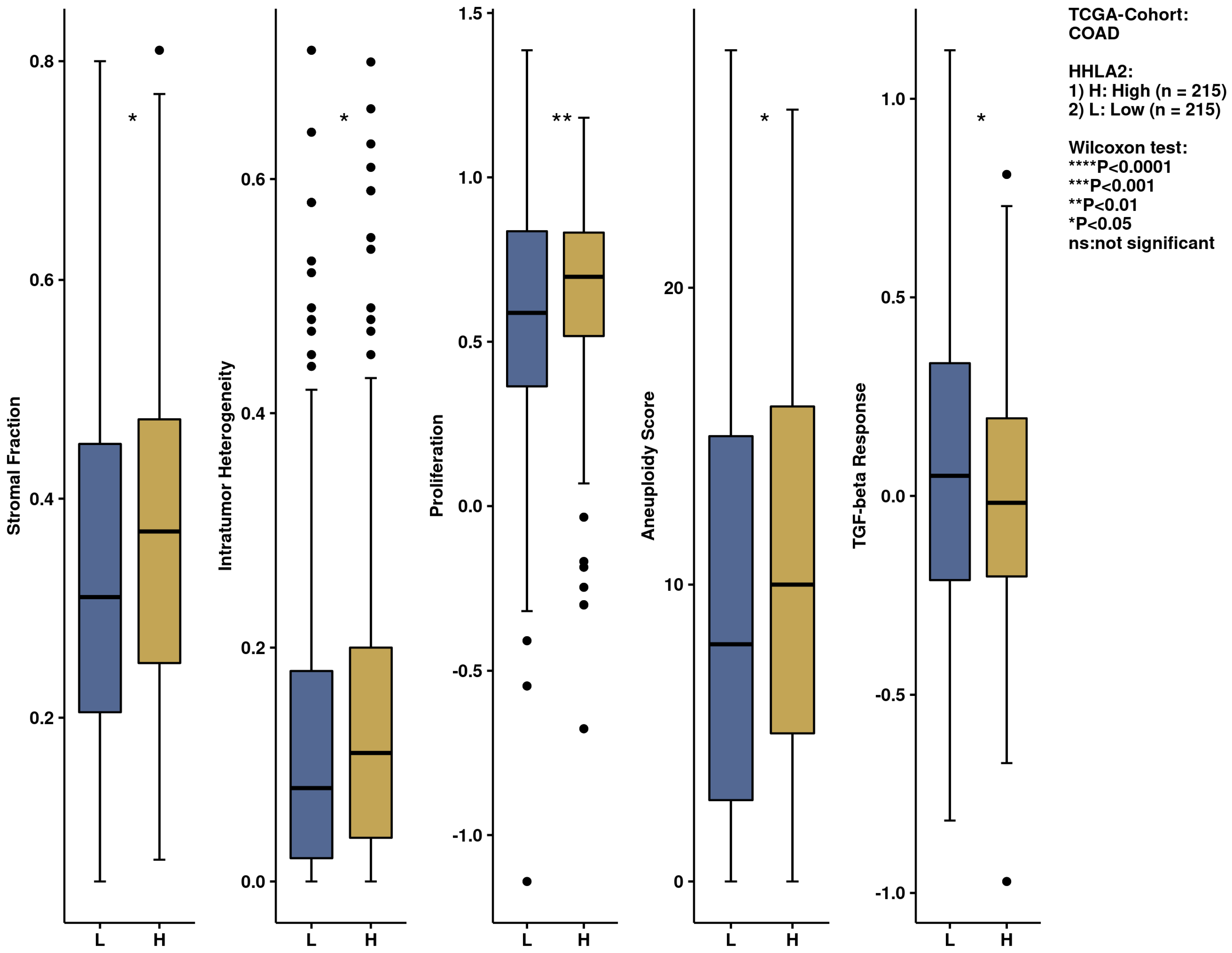
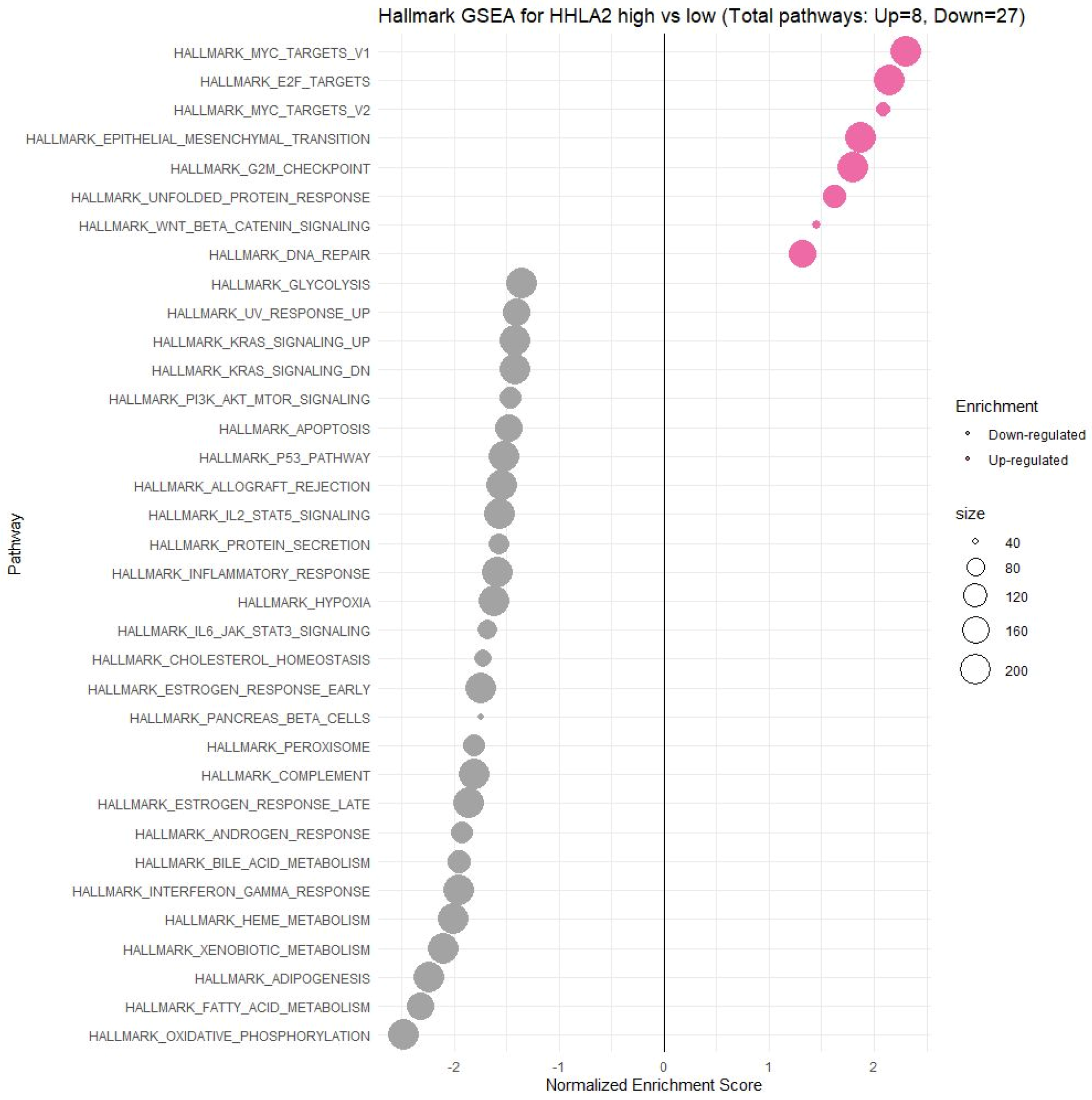
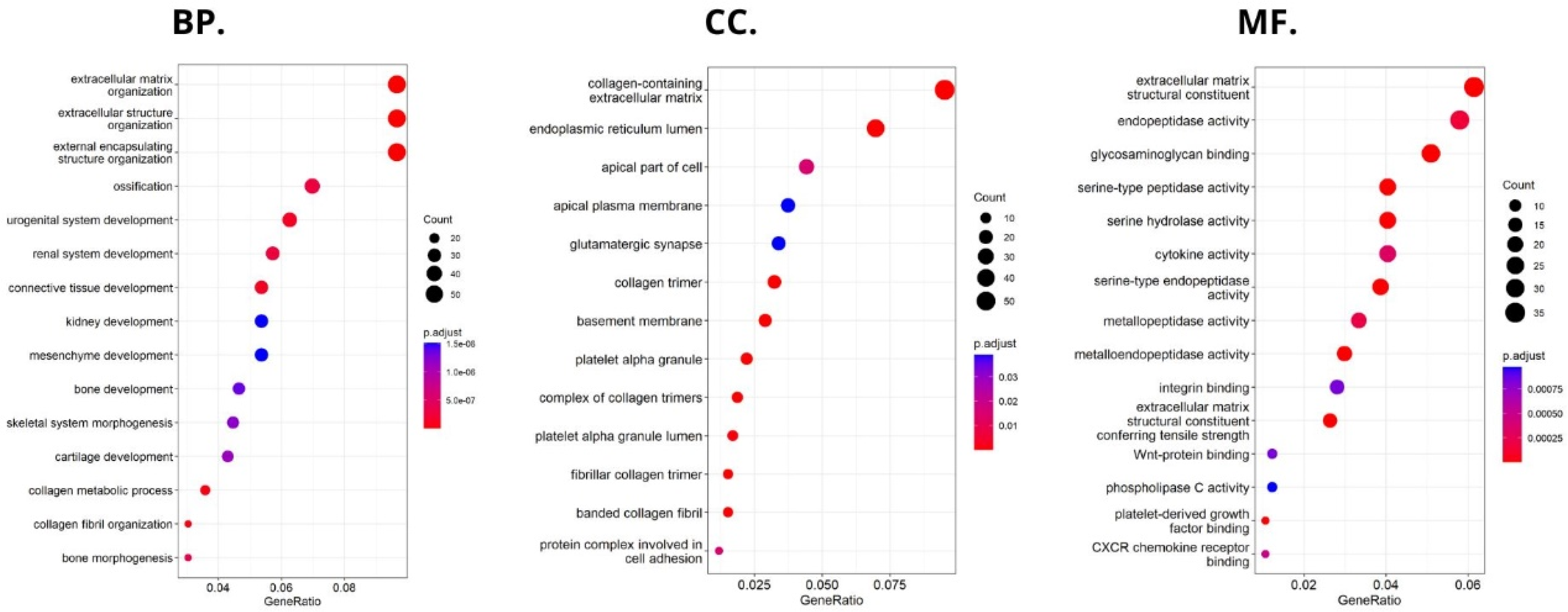
2.5. Comprehensive Analysis of Signaling Pathways for HHLA2 Expression
3. Discussion
4. Materials and Methods
4.1. Study Sample
4.2. Evaluation of HHLA2 Level by ELISA
4.3. Evaluation of the HHLA2 Expression by IHC
4.4. Assessment of the MSI Status
- -
- Mouse Monoclonal Antibody MSH2 (G219-1129), Cell Marque; incubation time, 30′; dilution, 1:400; room temperature.
- -
- Mouse Monoclonal Antibody MSH6 (44), Cell Marque; incubation time, 45′; dilution, 1:100; room temperature.
- -
- Mouse Monoclonal Antibody PMS2 (MRQ-28), Cell Marque; incubation time, 40′; dilution, 1:50; room temperature.
- -
- Mouse Monoclonal Antibody MLH1 (G168-728), Cell Marque; incubation time, 40′; dilution, 1:100; room temperature.
4.5. Assessment of the Tumor-Infiltrating Cells and Budding
4.6. Assessment of the TILs and Budding
4.7. Bio-Plex Pro Human Cytokines Screening
4.8. Exploration of Biological Characteristics of HHLA2
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nojadeh, J.N.; Behrouz Sharif, S.; Sakhinia, E. Microsatellite instability in colorectal cancer. EXCLI J. 2018, 17, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Na, Y.; Joung, S.Y.; Lee, S.I.; Oh, S.C.; Min, B.W. The significance of microsatellite instability in colorectal cancer after controlling for clinicopathological factors. Medicine 2018, 97, e0019. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.F.; Bretes, L.; Furtado, I. Review of PD-1/PD-L1 Inhibitors in Metastatic dMMR/MSI-H Colorectal Cancer. Front. Oncol. 2019, 9, 396. [Google Scholar] [CrossRef]
- Núñez Abad, M.; Calabuig-Fariñas, S.; Lobo de Mena, M.; Torres-Martínez, S.; García González, C.; García García, J.Á.; Iranzo González-Cruz, V.; Camps Herrero, C. Programmed Death-Ligand 1 (PD-L1) as Immunotherapy Biomarker in Breast Cancer. Cancers 2022, 14, 307. [Google Scholar] [CrossRef]
- Wang, C.; Fakih, M. Targeting MSS colorectal cancer with immunotherapy: Are we turning the corner? Expert Opin. Biol. Ther. 2021, 21, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Huyghe, N.; Benidovskaya, E.; Stevens, P.; van den Eynde, M. Biomarkers of Response and Resistance to Immunotherapy in Microsatellite Stable Colorectal Cancer: Toward a New Personalized Medicine. Cancers 2022, 14, 2241. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, R.W.; Barbie, D.A.; Flaherty, K.T. Mechanisms of resistance to immune checkpoint inhibitors. Br. J. Cancer 2018, 118, 9–16. [Google Scholar] [CrossRef]
- Schoenfeld, A.J.; Hellmann, M.D. Acquired Resistance to Immune Checkpoint Inhibitors. Cancer Cell 2020, 37, 443–455. [Google Scholar] [CrossRef]
- Cheng, H.; Janakiram, M.; Borczuk, A.; Lin, J.; Qiu, W.; Liu, H.; Chinai, J.M.; Halmos, B.; Perez-Soler, R.; Zang, X. HHLA2, a New Immune Checkpoint Member of the B7 Family, Is Widely Expressed in Human Lung Cancer and Associated with EGFR Mutational Status. Clin. Cancer Res. 2017, 23, 825–832. [Google Scholar] [CrossRef]
- Farrag, M.S.; Ibrahim, E.M.; El-Hadidy, T.A.; Akl, M.F.; Elsergany, A.R.; Abdelwahab, H.W. Human Endogenous Retrovirus-H Long Terminal Repeat- Associating Protein 2 (HHLA2) is a Novel Immune Checkpoint Protein in Lung Cancer which Predicts Survival. Asian Pac. J. Cancer Prev. 2021, 22, 1883–1889. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, J.; Zhang, C.; Li, F.; Li, L.; Wang, D.; Chand, D.; Guan, F.; Zang, X.; Zhang, Y. Over-Expression and Prognostic Significance of HHLA2, a New Immune Checkpoint Molecule, in Human Clear Cell Renal Cell Carcinoma. Front. Cell Dev. Biol. 2020, 8, 280. [Google Scholar] [CrossRef]
- Zhou, Q.-H.; Li, K.-W.; Chen, X.; He, H.-X.; Peng, S.-M.; Peng, S.-R.; Wang, Q.; Li, Z.-A.; Tao, Y.-R.; Cai, W.-L.; et al. HHLA2 and PD-L1 co-expression predicts poor prognosis in patients with clear cell renal cell carcinoma. J. Immunother. Cancer 2020, 8, e000157. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, D.; Ye, X.; Lin, X.; Zhang, M.; Su, Y.; Xie, Q.; Ni, W. HHLA2 Used as a Potential Prognostic and Immunological Biomarker and Correlated with Tumor Microenvironment in Pan-Cancer. Biomed Res. Int. 2022, 2022, 3924400. [Google Scholar] [CrossRef]
- Zhang, W.; Acuna-Villaorduna, A.; Kuan, K.; Gupta, S.; Hu, S.; Ohaegbulam, K.; Albanese, J.; Kaumaya, M.; Levy, R.; Hwang, R.R.; et al. B7-H3 and PD-L1 Expression Are Prognostic Biomarkers in a Multi-racial Cohort of Patients with Colorectal Cancer. Clin. Color. Cancer 2021, 20, 161–169. [Google Scholar] [CrossRef]
- Payandeh, Z.; Khalili, S.; Somi, M.H.; Mard-Soltani, M.; Baghbanzadeh, A.; Hajiasgharzadeh, K.; Samadi, N.; Baradaran, B. PD-1/PD-L1-dependent immune response in colorectal cancer. J. Cell. Physiol. 2020, 235, 5461–5475. [Google Scholar] [CrossRef]
- Ying, H.; Xu, J.; Zhang, X.; Liang, T.; Bai, X. Human endogenous retrovirus-H long terminal repeat-associating 2: The next immune checkpoint for antitumour therapy. eBioMedicine 2022, 79, 103987. [Google Scholar] [CrossRef]
- Wang, C.; Feng, H.; Cheng, X.; Liu, K.; Cai, D.; Zhao, R. Potential Therapeutic Targets of B7 Family in Colorectal Cancer. Front. Immunol. 2020, 11, 681. [Google Scholar] [CrossRef]
- Zhu, G.-X.; Gao, D.; Shao, Z.-Z.; Chen, L.; Ding, W.-J.; Yu, Q.-F. Wnt/β-catenin signaling: Causes and treatment targets of drug resistance in colorectal cancer (Review). Mol. Med. Rep. 2021, 23, 105. [Google Scholar] [CrossRef]
- Mlecnik, B.; Bindea, G.; Angell, H.K.; Maby, P.; Angelova, M.; Tougeron, D.; Church, S.E.; Lafontaine, L.; Fischer, M.; Fredriksen, T.; et al. Integrative Analyses of Colorectal Cancer Show Immunoscore Is a Stronger Predictor of Patient Survival Than Microsatellite Instability. Immunity 2016, 44, 698–711. [Google Scholar] [CrossRef]
- Angell, H.K.; Bruni, D.; Barrett, J.C.; Herbst, R.; Galon, J. The Immunoscore: Colon Cancer and Beyond. Clin. Cancer Res. 2020, 26, 332–339. [Google Scholar] [CrossRef]
- Yang, L.; Chao, L.; Yang, Y.; Baokang, W.; Yizhou, Z.; Qi, L.; Zhiyun, L.; Chongli, Z.; Yu, S.; Shukun, H.; et al. KIR3DL3-HHLA2 and TMIGD2-HHLA2 pathways: The dual role of HHLA2 in immune responses and its potential therapeutic approach for cancer immunotherapy. J. Adv. Res. 2022, in press. [Google Scholar] [CrossRef]
- Bolandi, N.; Derakhshani, A.; Hemmat, N.; Baghbanzadeh, A.; Asadzadeh, Z.; Afrashteh Nour, M.; Brunetti, O.; Bernardini, R.; Silvestris, N.; Baradaran, B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 10719. [Google Scholar] [CrossRef]
- Wei, Y.; Ren, X.; Galbo, P.M., Jr.; Moerdler, S.; Wang, H.; Sica, R.A.; Etemad-Gilbertson, B.; Shi, L.; Zhu, L.; Tang, X.; et al. KIR3DL3-HHLA2 is a human immunosuppressive pathway and a therapeutic target. Sci. Immunol. 2021, 6, eabf9792. [Google Scholar] [CrossRef] [PubMed]
- Manfioletti, G.; Fedele, M. Epithelial-Mesenchymal Transition (EMT) 2021. Int. J. Mol. Sci. 2022, 23, 5848. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.; Datta, P.K. Regulation of EMT in Colorectal Cancer: A Culprit in Metastasis. Cancers 2017, 9, 171. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, D.; Feng, J.; Zhou, Y.; Wang, Q.; Feng, H.; Zhang, J.; Jiang, J. Overexpression of HHLA2 in human clear cell renal cell carcinoma is significantly associated with poor survival of the patients. Cancer Cell Int. 2019, 19, 101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Lv, C.; Wu, B.; Yu, Y.; Zhong, C.; Lang, Q.; Liang, Z.; Li, Y.; Shi, Y.; et al. HHLA2 promotes tumor progression by long non-coding RNA H19 in human gallbladder cancer. Int. J. Oncol. 2022, 61, 112. [Google Scholar] [CrossRef]
- Rochlitz, C.F.; Herrmann, R.; de Kant, E. Overexpression and amplification of c-myc during progression of human colorectal cancer. Oncology 1996, 53, 448–454. [Google Scholar] [CrossRef]
- Strippoli, A.; Cocomazzi, A.; Basso, M.; Cenci, T.; Ricci, R.; Pierconti, F.; Cassano, A.; Fiorentino, V.; Barone, C.; Bria, E.; et al. c-MYC Expression Is a Possible Keystone in the Colorectal Cancer Resistance to EGFR Inhibitors. Cancers 2020, 12, 638. [Google Scholar] [CrossRef]
- Urbanski, L.; Brugiolo, M.; Park, S.; Angarola, B.L.; Leclair, N.K.; Yurieva, M.; Palmer, P.; Sahu, S.K.; Anczuków, O. MYC regulates a pan-cancer network of co-expressed oncogenic splicing factors. Cell Rep. 2022, 41, 111704. [Google Scholar] [CrossRef]
- Schulze, A.; Oshi, M.; Endo, I.; Takabe, K. MYC Targets Scores Are Associated with Cancer Aggressiveness and Poor Survival in ER-Positive Primary and Metastatic Breast Cancer. Int. J. Mol. Sci. 2020, 21, 8127. [Google Scholar] [CrossRef]
- Oshi, M.; Patel, A.; Le, L.; Tokumaru, Y.; Yan, L.; Matsuyama, R.; Endo, I.; Takabe, K. G2M checkpoint pathway alone is associated with drug response and survival among cell proliferation-related pathways in pancreatic cancer. Am. J. Cancer Res. 2021, 11, 3070–3084. [Google Scholar]
- Oshi, M.; Takahashi, H.; Tokumaru, Y.; Yan, L.; Rashid, O.M.; Nagahashi, M.; Matsuyama, R.; Endo, I.; Takabe, K. The E2F Pathway Score as a Predictive Biomarker of Response to Neoadjuvant Therapy in ER+/HER2- Breast Cancer. Cells 2020, 9, 1643. [Google Scholar] [CrossRef]
- Xie, D.; Pei, Q.; Li, J.; Wan, X.; Ye, T. Emerging Role of E2F Family in Cancer Stem Cells. Front. Oncol. 2021, 11, 723137. [Google Scholar] [CrossRef]
- Oshi, M.; Takahashi, H.; Tokumaru, Y.; Yan, L.; Rashid, O.M.; Matsuyama, R.; Endo, I.; Takabe, K. G2M Cell Cycle Pathway Score as a Prognostic Biomarker of Metastasis in Estrogen Receptor (ER)-Positive Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2921. [Google Scholar] [CrossRef]
- Ramón, Y.; Cajal, S.; Sesé, M.; Capdevila, C.; Aasen, T.; de Mattos-Arruda, L.; Diaz-Cano, S.J.; Hernández-Losa, J.; Castellví, J. Clinical implications of intratumor heterogeneity: Challenges and opportunities. J. Mol. Med. 2020, 98, 161–177. [Google Scholar] [CrossRef]
- Micke, P.; Strell, C.; Mattsson, J.; Martín-Bernabé, A.; Brunnström, H.; Huvila, J.; Sund, M.; Wärnberg, F.; Ponten, F.; Glimelius, B.; et al. The prognostic impact of the tumour stroma fraction: A machine learning-based analysis in 16 human solid tumour types. eBioMedicine 2021, 65, 103269. [Google Scholar] [CrossRef]
- Zhao, R.; Chinai, J.M.; Buhl, S.; Scandiuzzi, L.; Ray, A.; Jeon, H.; Ohaegbulam, K.C.; Ghosh, K.; Zhao, A.; Scharff, M.D.; et al. HHLA2 is a member of the B7 family and inhibits human CD4 and CD8 T-cell function. Proc. Natl. Acad. Sci. USA 2013, 110, 9879–9884. [Google Scholar] [CrossRef]
- Kula, A.; Dawidowicz, M.; Mielcarska, S.; Kiczmer, P.; Chrabańska, M.; Rynkiewicz, M.; Świętochowska, E.; Waniczek, D. Periostin in Angiogenesis and Inflammation in CRC—A Preliminary Observational Study. Medicina 2022, 58, 96. [Google Scholar] [CrossRef]
- Zhu, Z.; Dong, W. Overexpression of HHLA2, a member of the B7 family, is associated with worse survival in human colorectal carcinoma. OncoTargets Ther. 2018, 11, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Fassan, M.; Scarpa, A.; Remo, A.; de Maglio, G.; Troncone, G.; Marchetti, A.; Doglioni, C.; Ingravallo, G.; Perrone, G.; Parente, P.; et al. Current prognostic and predictive biomarkers for gastrointestinal tumors in clinical practice. Pathologica 2020, 112, 248–259. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Idzik, M.; Poloczek, J.; Skrzep-Poloczek, B.; Dróżdż, E.; Chełmecka, E.; Czuba, Z.; Jochem, J.; Stygar, D. The Effects of 21-Day General Rehabilitation after Hip or Knee Surgical Implantation on Plasma Levels of Selected Interleukins, VEGF, TNF-α, PDGF-BB, and Eotaxin-1. Biomolecules 2022, 12, 605. [Google Scholar] [CrossRef]
- Grudzińska, E.; Grzegorczyn, S.; Czuba, Z.P. Chemokines and Growth Factors Produced by Lymphocytes in the Incompetent Great Saphenous Vein. Mediat. Inflamm. 2019, 2019, 7057303. [Google Scholar] [CrossRef] [PubMed]
- Dampier, C.H.; Devall, M.; Jennelle, L.T.; Díez-Obrero, V.; Plummer, S.J.; Moreno, V.; Casey, G. Oncogenic Features in Histologically Normal Mucosa: Novel Insights Into Field Effect From a Mega-Analysis of Colorectal Transcriptomes. Clin. Transl. Gastroenterol. 2020, 11, e00210. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HHLA2 Concentration | Mean | SD | p |
|---|---|---|---|
| Log HHLA2 tumor | 0.17 | 0.16 | <0.0001 |
| Log HHLA2 margin | 0.05 | 0.14 |
| Characteristics | HHLA2 Tumor Expression | |
|---|---|---|
| all | positive | negative |
| 77 (100%) | 75 (97.40%) | 2 (2.60%) |
| MSS/MSI Status | HHLA2 Expression | ||
|---|---|---|---|
| Negative | Positive | p | |
| MSS | 1 (1.49%) | 66 (98.51%) | 0.2 |
| MSI | 1 (10%) | 9 (90%) | |
| Variable | Factor1 | Factor2 |
|---|---|---|
| logHGF | 0.686 | −0.169 |
| logMCSF | 0.415 | |
| logbNGF | 0.565 | 0.172 |
| logSCGF-b | 0.451 | |
| logIL-7 | 0.597 | −0.332 |
| logBasic-FGF | 0.722 | |
| logG-CSF | 0.880 | −0.173 |
| logPDGF-bb | 0.513 | 0.506 |
| logVEGFA | 0.437 | 0.139 |
| logGM-CSF | 0.590 | 0.463 |
| eigenvalue | 3.488 | 1.876 |
| variance.percent | 34.883 | 18.761 |
| cumulative.variance.percent | 34.883 | 53.645 |
| Variable | Factor1 | Factor2 |
|---|---|---|
| log(IL-1a_tumor/protein) | 0.890 | −0.211 |
| log(IL-12p40_tumor/protein) | 0.718 | 0.580 |
| logIL-13_tumor/protein) | 0.018 | 0.944 |
| log(TNF-a-tumor/protein) | 0.880 | 0.335 |
| eigenvalue | 2.084 | 1.386 |
| variance.percent | 52.11 | 34.67 |
| cumulative.variance.percent | 52.11 | 87.78 |
| Female | Male | All Cases | |
|---|---|---|---|
| 76 (45.51%) | 91 (54.49%) | 167 (100%) | |
| Age | 66.63 ± 9.48 | 63.75 ± 9.46 | 65.09 ± 9.55 |
| Tumor localization | |||
| Left-sided | 49 (66.22%) | 65 (73.86%) | 114 (70.37%) |
| Right-sided | 25 (33.78%) | 23 (26.13%) | 48 (29.63%) |
| T parameter | |||
| T1 | 1 (1.33%) | 7 (8.05%) | 8 (4.94%) |
| T2 | 16 (21.33%) | 12 (13.79%) | 28 (22.22%) |
| T3 | 47 (62.67%) | 54 (62.07%) | 101 (62.35%) |
| T4 | 11 (14.67%) | 14 (16.09%) | 25 (15.43%) |
| N parameter | |||
| N0 | 32 (42.67%) | 37 (42.05%) | 69 (42.33%) |
| N1 | 30 (40.00%) | 37 (42.05%) | 67 (41.10%) |
| N2 | 13 (17.33%) | 14 (15.91%) | 27 (16.56%) |
| M parameter | |||
| M0 | 66 (88.00%) | 69 (79.31%) | 135 (83.33%) |
| M1 | 9 (12.00%) | 18 (20.69%) | 27 (16.67%) |
| TNM stage | |||
| I | 13 (17.33%) | 13 (14.77%) | 26 (15.95%) |
| II | 19 (25.33%) | 21 (28.86%) | 40 (24.54%) |
| III | 34 (45.33%) | 37 (42.05%) | 71 (43.55%) |
| IV | 9 (12.00%) | 17 (19.32%) | 26 (15.95%) |
| Grading | |||
| Low | 63 (85.14%) | 75 (85.23%) | 138 (85.16%) |
| High | 11 (14.86%) | 13 (14.777%) | 24 (14.81%) |
| HHLA2 IHC expression | |||
| Positive | 34 (97.14%) | 41 (97.62%) | 77 (97.40%) |
| Negative | 1 (2.86%) | 1 (2.38%) | 2 (2.60%) |
| MSS/MSI status (N = 101) | |||
| MSS tumors | 36 (76.60%) | 45 (83.33%) | 81 (80.20%) |
| MSI tumors | 11 (23.40%) | 9 (16.67%) | 20 (19.80%) |
| TILs (N = 102) | |||
| 0–5% | 16 (34.04%) | 30 (53.5%) | 46 (44.66%) |
| 6–25% | 15 (31.91%) | 13 (23.21%) | 28 (27.18%) |
| 26–50% | 14 (29.79%) | 8 (14.29%) | 22 (21.36%) |
| 51–75% | 2 (4.26%) | 4 (7.14%) | 6 (5.83%) |
| 76–100% | 0 (0.00%) | 1 (1.79%) | 1 (0.97%) |
| CD8+ Lymphocytes (N = 76) | |||
| 0–5% | 14 (40.00%) | 20 (47.62%) | 34 (44.16%) |
| 6–25% | 8 (22.86%) | 13 (30.95%) | 21 (71.43%) |
| 26–50% | 7 (20.00%) | 5 (11.90%) | 12 (87.01%) |
| 51–75% | 4 (11.43%) | 2 (4.76%) | 6 (7.79%) |
| 76–100% | 2 (5.71%) | 2 (4.76%) | 4 (5.19%) |
| Budding (N = 101) | |||
| 0–4 | 28 (59.57%) | 29 (53.70%) | 57 (56.44%) |
| 5–9 | 13 (27.66%) | 11 (20.37%) | 24 (23.76%) |
| >9 | 6 (12.77%) | 14 (25.93%) | 20 (19.80%) |
| Adjuvant treatment | |||
| yes | 7 (9.21%) | 14 (15.38%) | 21 (12.35%) |
| no | 69 (90.79%) | 77 (84.62%) | 149 (87.65%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kula, A.; Dawidowicz, M.; Mielcarska, S.; Kiczmer, P.; Skiba, H.; Krygier, M.; Chrabańska, M.; Piecuch, J.; Szrot, M.; Robotycka, J.; et al. Overexpression and Role of HHLA2, a Novel Immune Checkpoint, in Colorectal Cancer. Int. J. Mol. Sci. 2023, 24, 5876. https://doi.org/10.3390/ijms24065876
Kula A, Dawidowicz M, Mielcarska S, Kiczmer P, Skiba H, Krygier M, Chrabańska M, Piecuch J, Szrot M, Robotycka J, et al. Overexpression and Role of HHLA2, a Novel Immune Checkpoint, in Colorectal Cancer. International Journal of Molecular Sciences. 2023; 24(6):5876. https://doi.org/10.3390/ijms24065876
Chicago/Turabian StyleKula, Agnieszka, Miriam Dawidowicz, Sylwia Mielcarska, Paweł Kiczmer, Hanna Skiba, Małgorzata Krygier, Magdalena Chrabańska, Jerzy Piecuch, Monika Szrot, Julia Robotycka, and et al. 2023. "Overexpression and Role of HHLA2, a Novel Immune Checkpoint, in Colorectal Cancer" International Journal of Molecular Sciences 24, no. 6: 5876. https://doi.org/10.3390/ijms24065876
APA StyleKula, A., Dawidowicz, M., Mielcarska, S., Kiczmer, P., Skiba, H., Krygier, M., Chrabańska, M., Piecuch, J., Szrot, M., Robotycka, J., Ochman, B., Strzałkowska, B., Czuba, Z., Świętochowska, E., & Waniczek, D. (2023). Overexpression and Role of HHLA2, a Novel Immune Checkpoint, in Colorectal Cancer. International Journal of Molecular Sciences, 24(6), 5876. https://doi.org/10.3390/ijms24065876