
Safety and Immunogenicity of Recombinant Bacille Calmette-Guérin Strain VPM1002 and Its Derivatives in a Goat Model

 , and
, and
Abstract
1. Introduction
2. Results
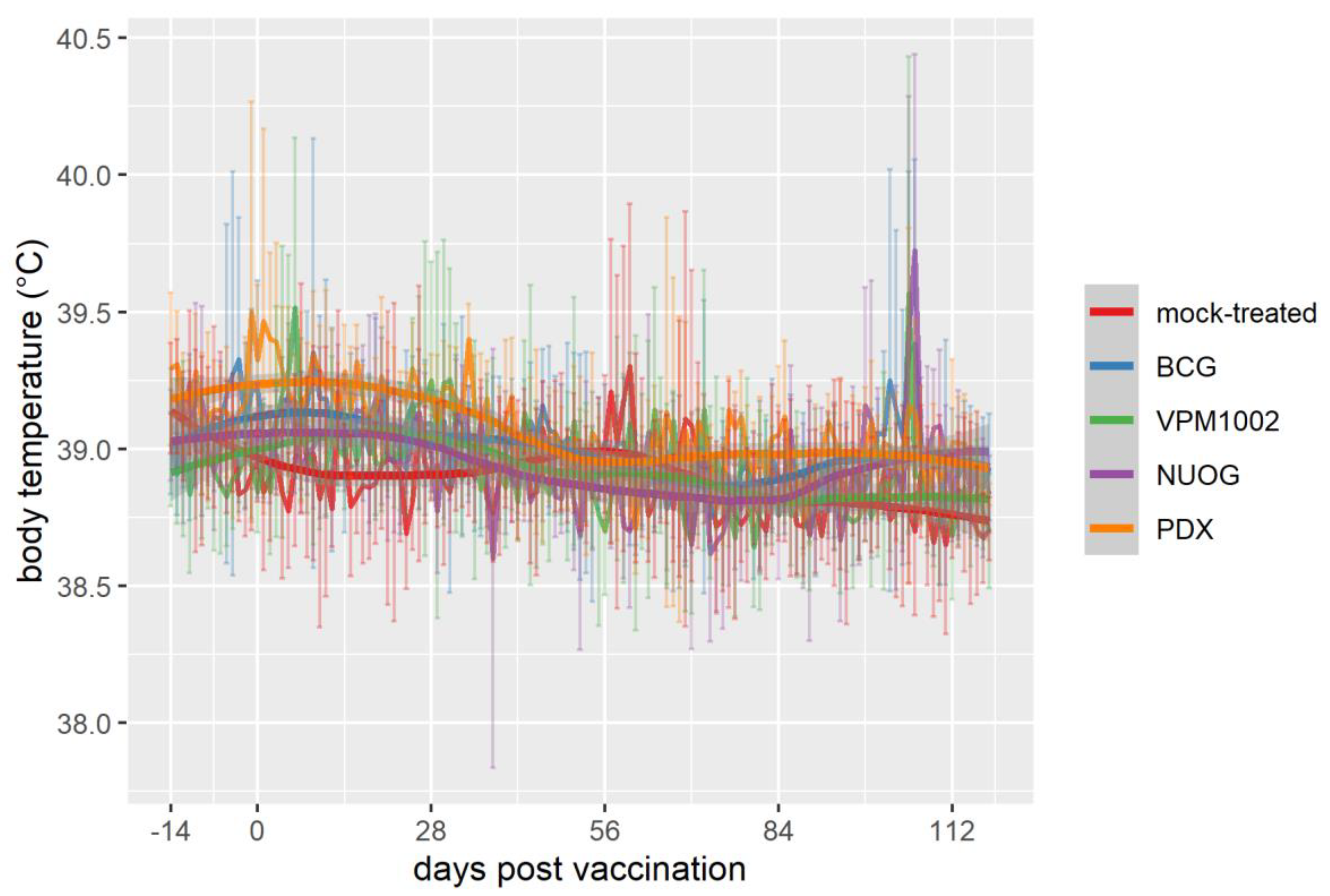
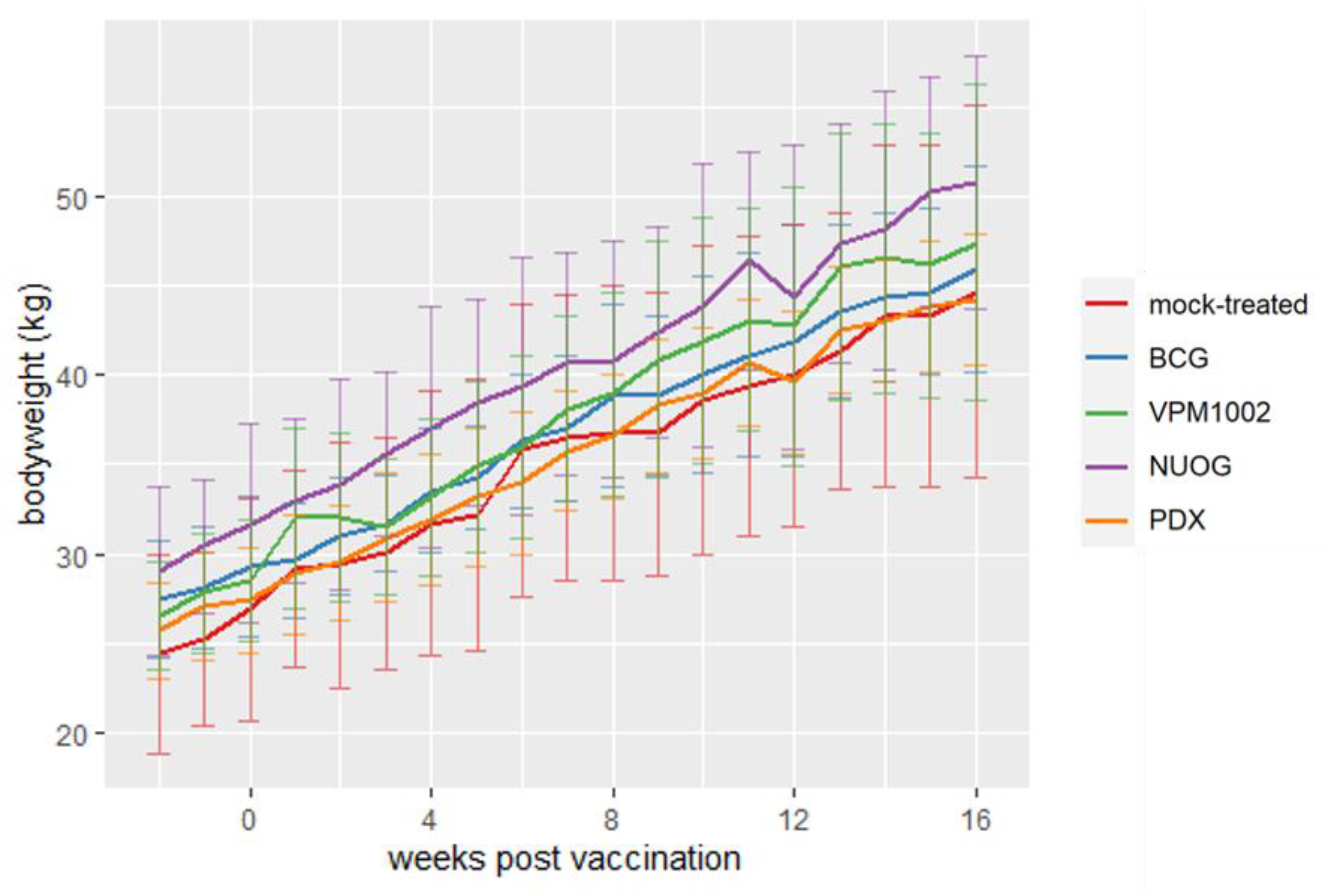
2.1. Clinical, Hematological, and Bacteriological Findings Intra Vitam
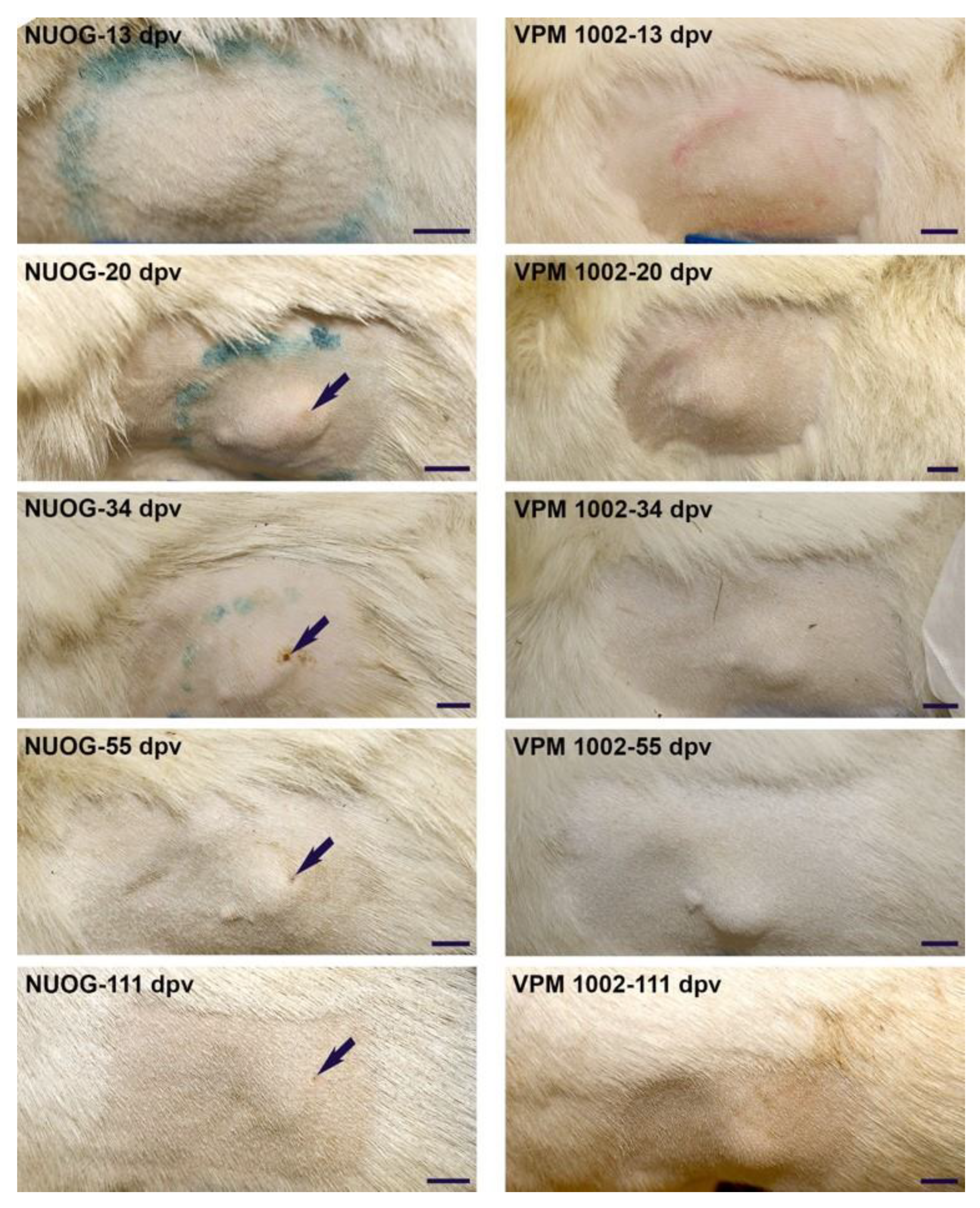
2.2. Post Mortem Findings
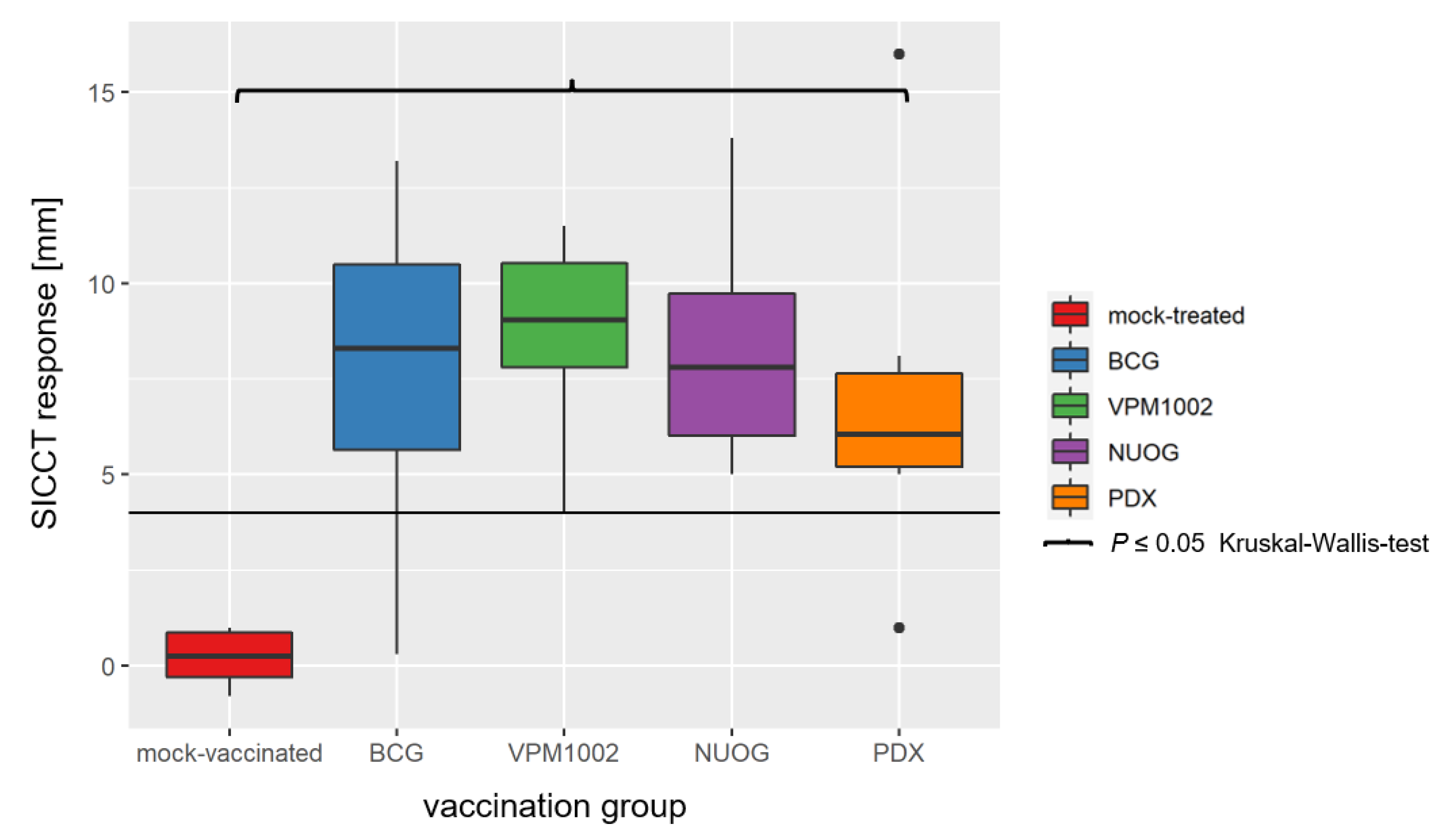
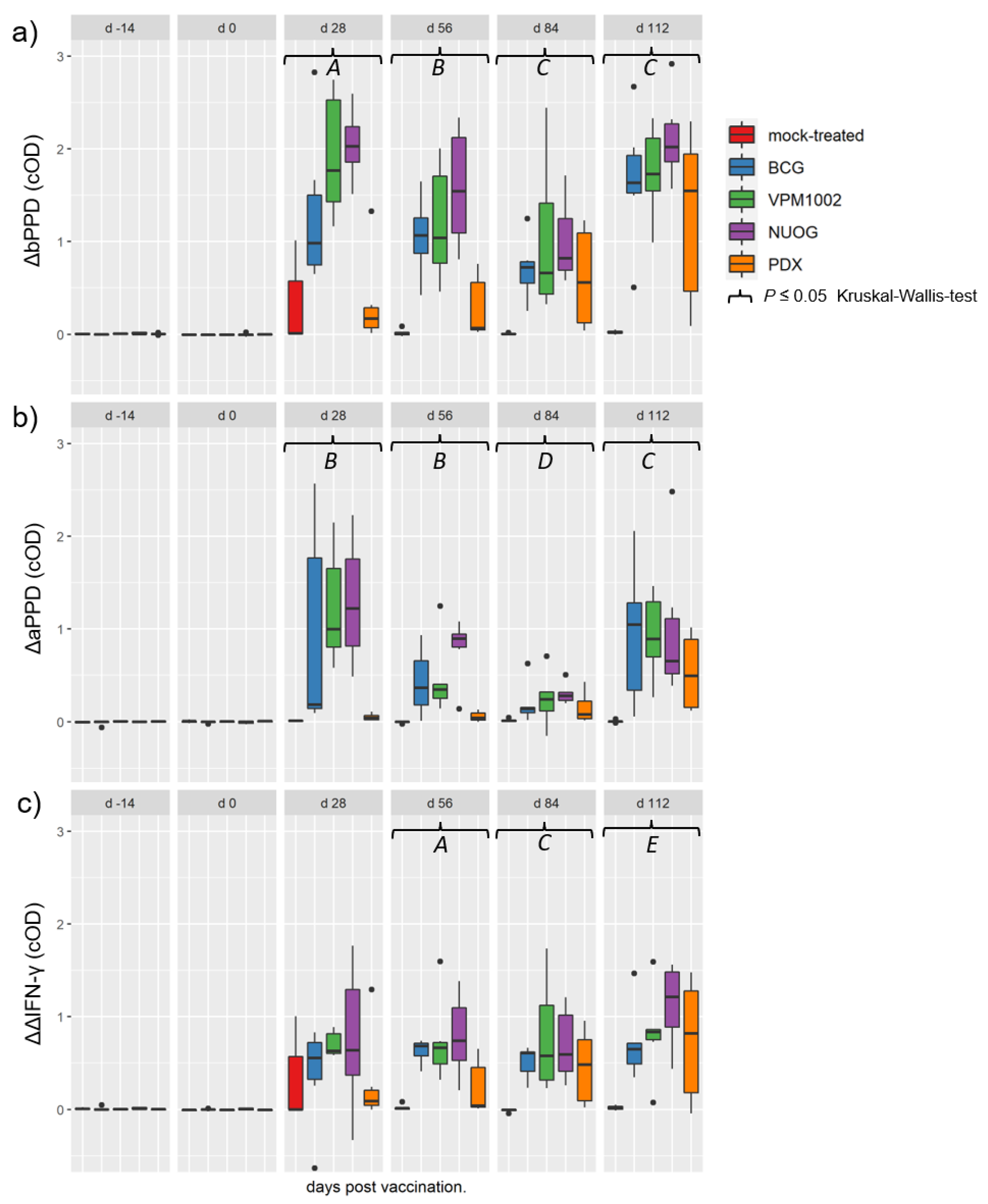
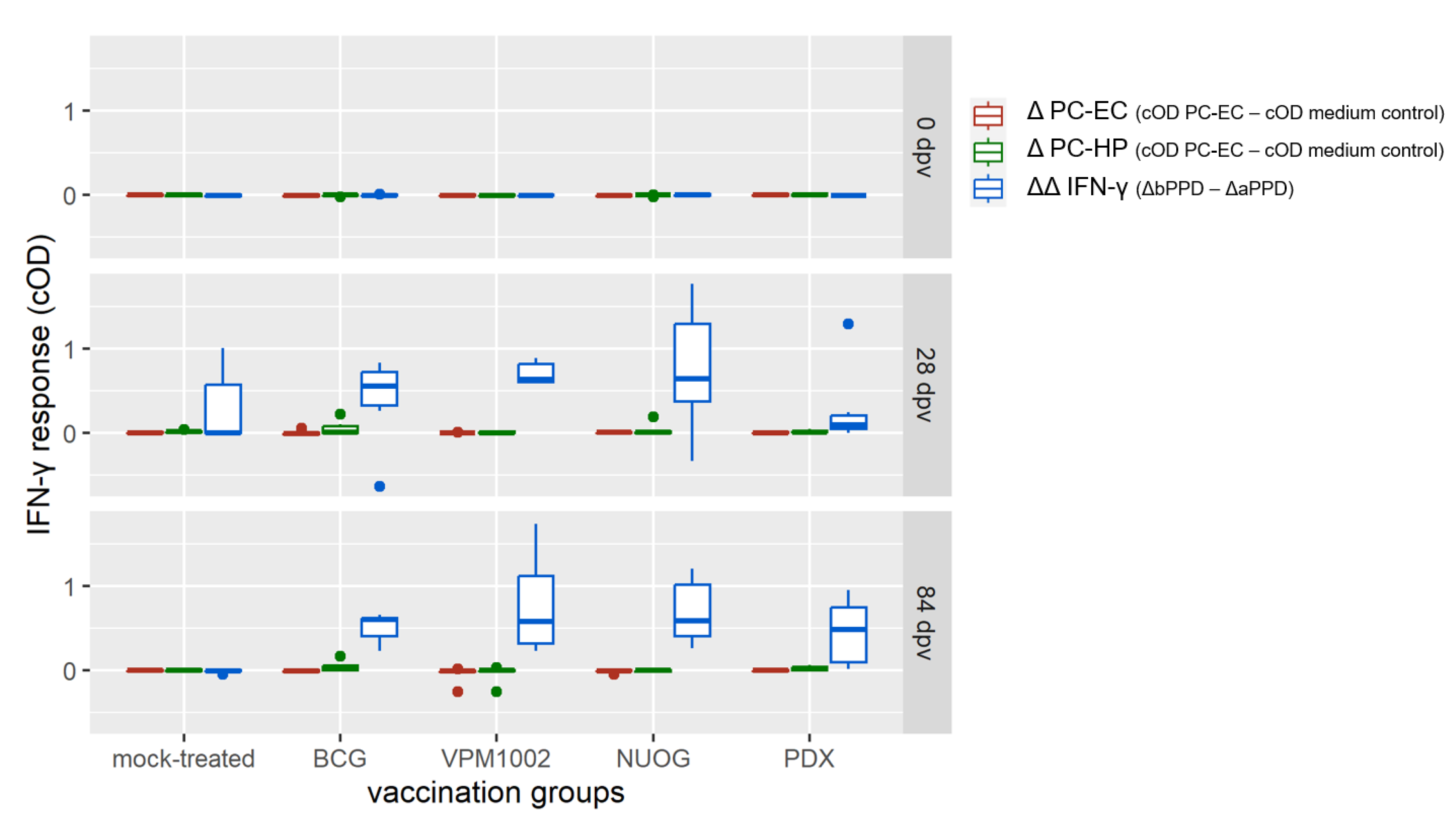
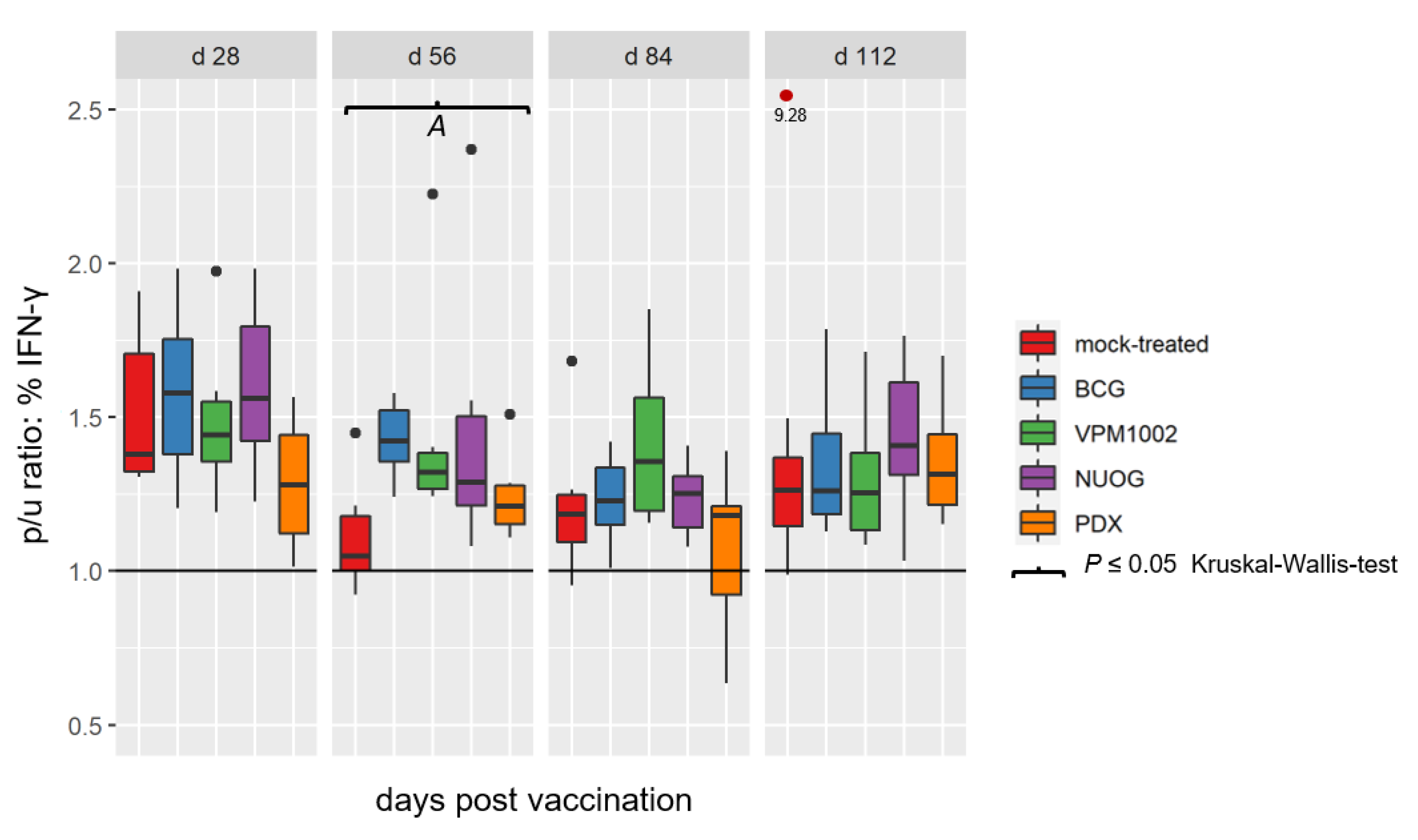
2.3. Cell-Mediated Immune Response Assessed by SICCT and IGRA
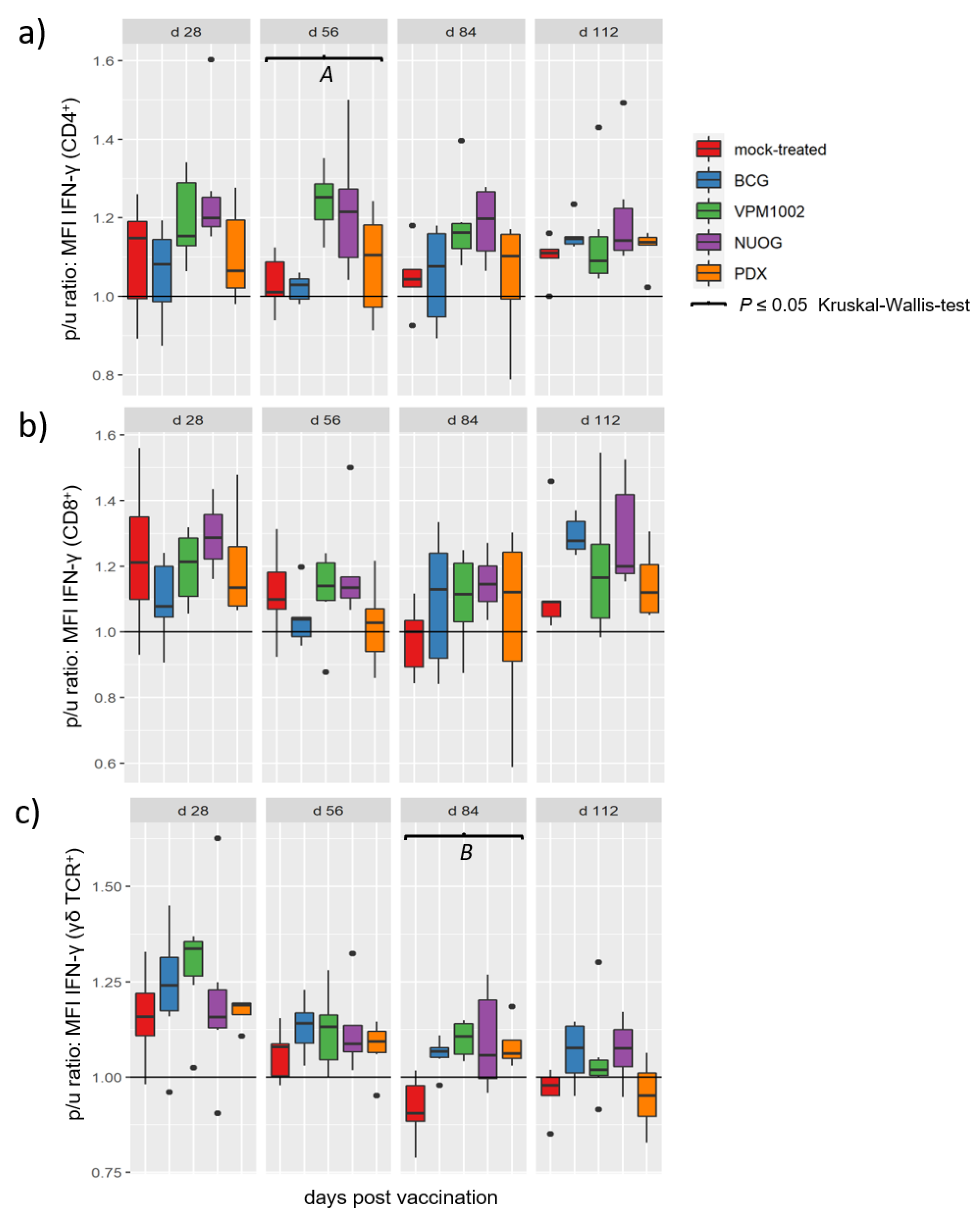
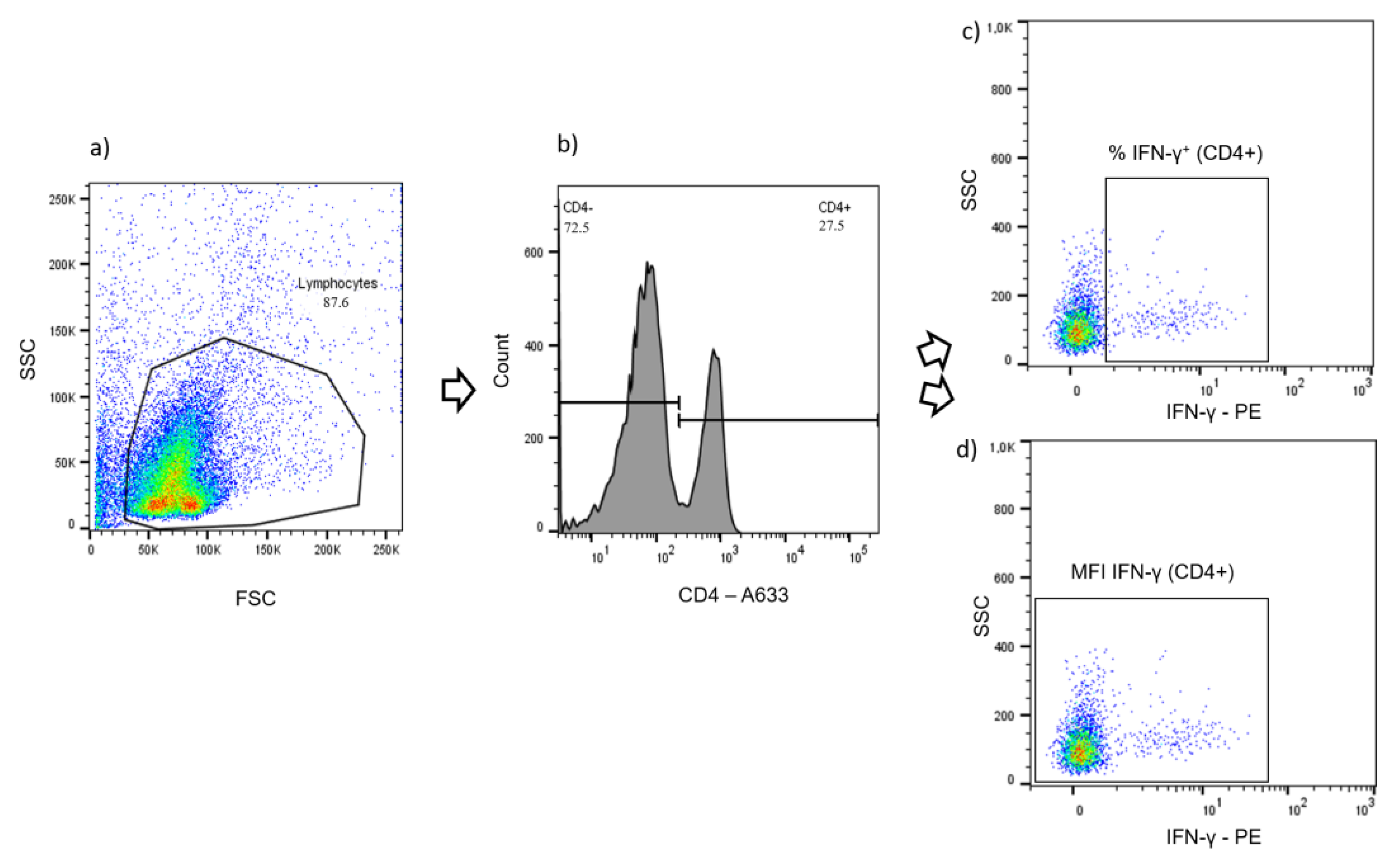
2.4. Cellular Immune Responses Assessed by Quantitation of Intracellular IFN-γ in T Cell Subsets after In Vitro Stimulation
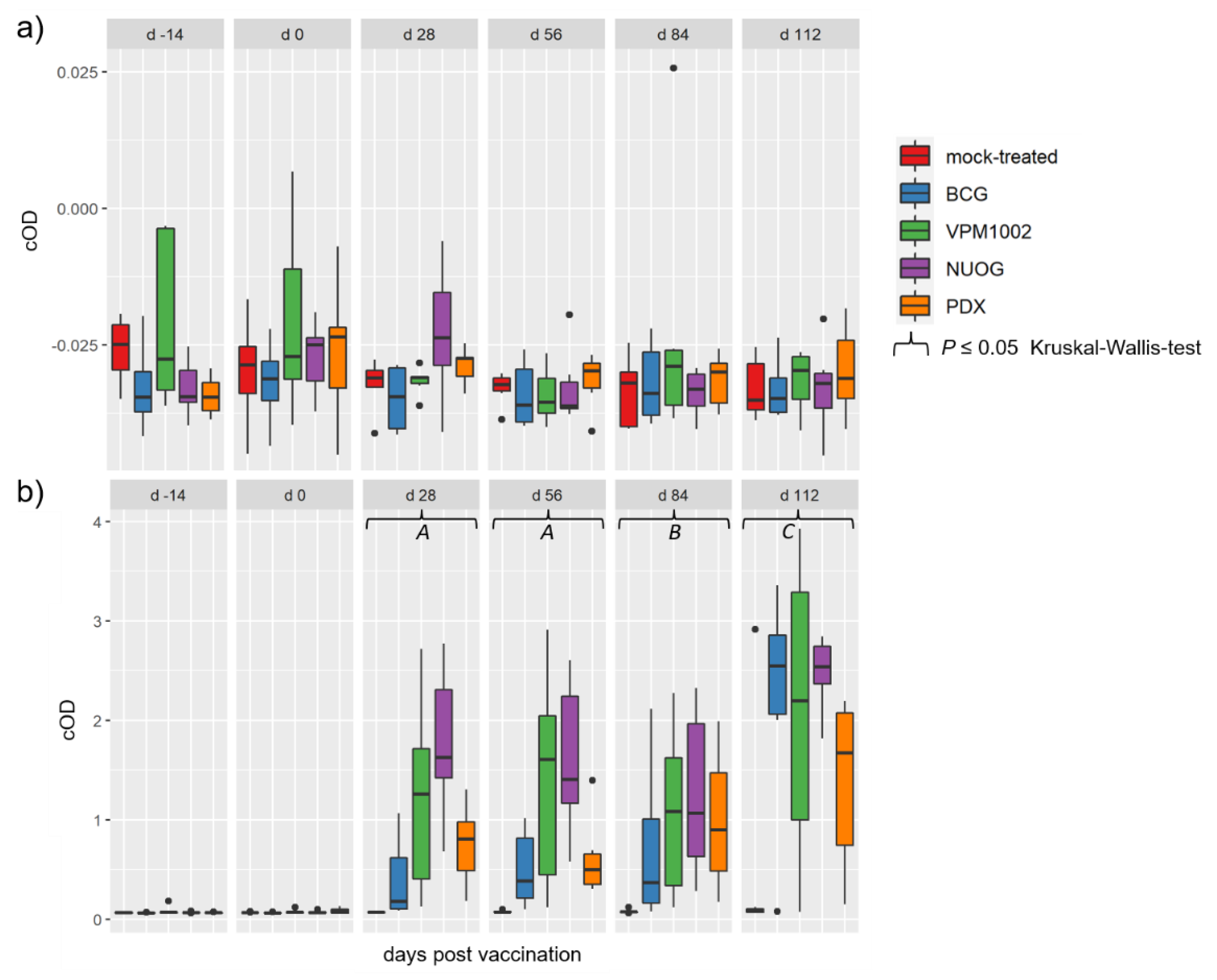
2.5. Humoral Immune Responses
3. Discussion
4. Materials and Methods
4.1. Experimental Animals, Housing, and Health Status
4.2. Vaccination, Clinical Examination, and Intra Vitam Sampling
4.3. Blood Count and Vitamin B6 Levels
4.4. Interferon Gamma Release Assay (IGRA)—ELISA
4.5. Intracellular IFN-γ of Different T Cell Subsets (Flow Cytometry)
4.6. Antibody Response against M. bovis and M. avium subsp. Paratuberculosis (MAP)
4.7. Single Intradermal Comparative Cervical Test (SICCT)
4.8. Necropsy
4.9. Bacterial Examination of Nasal Swabs and Tissue Samples
4.10. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2022; World Health Organization: Geneva, Switzerland, 2022.
- De Lisle, G.W.; Mackintosh, C.G.; Bengis, R.G. Mycobacterium bovis in free-living and captive wildlife, including farmed deer. Rev. Sci. Et Tech. Int. Off. Epizoot. 2001, 20, 86–111. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Alvarez, J.; Boniotti, M.B.; Cagiola, M.; Di Marco, V.; Marianelli, C.; Pacciarini, M.; Pasquali, P. Tuberculosis in domestic animal species. Res. Vet. Sci. 2014, 97, S78–S85. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; de Val, B.P.; Buddle, B.M.; Villarreal-Ramos, B.; Jones, G.J.; Hewinson, R.G.; Domingo, M. Vaccination of domestic animals against tuberculosis: Review of progress and contributions to the field of the TBSTEP project. Res. Vet. Sci. 2014, 97, S53–S60. [Google Scholar] [CrossRef] [PubMed]
- Buddle, B.M.; Vordermeier, H.M.; Hewinson, R.G. Experimental infection models of tuberculosis in domestic livestock. Microbiol. Spectr. 2016, 4, 66. [Google Scholar] [CrossRef]
- World Health Organization. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020.
- World Health Organization; Food and Agriculture Organization of the United Nations; World Organisation for Animal Health. Roadmap for Zoonotic Tuberculosis; World Health Organization: Geneva, Switzerland, 2017.
- World Health Organization. BCG Immunization Coverage Estimates by WHO Region. Available online: https://apps.who.int/gho/data/view.main.81500?lang=en (accessed on 4 May 2022).
- Andersen, P.; Kaufmann, S.H. Novel vaccination strategies against tuberculosis. Cold Spring Harb. Perspect. Med. 2014, 4, a018523. [Google Scholar] [CrossRef]
- Buddle, B.M.; Vordermeier, H.M.; Chambers, M.A.; de Klerk-Lorist, L.M. Efficacy and safety of BCG vaccine for control of tuberculosis in domestic livestock and wildlife. Front. Vet. Sci. 2018, 5, 259. [Google Scholar] [CrossRef]
- Parlane, N.A.; Shu, D.; Subharat, S.; Wedlock, D.N.; Rehm, B.H.; de Lisle, G.W.; Buddle, B.M. Revaccination of cattle with bacille Calmette-Guérin two years after first vaccination when immunity has waned, boosted protection against challenge with Mycobacterium bovis. PloS ONE 2014, 9, e106519. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Waters, W.R.; Thacker, T.C. Vaccination of white-tailed deer (Odocoileus virginianus) with Mycobacterium bovis bacille Calmette-Guérin (BCG) results in positive tuberculin skin test results in a dose-dependent fashion. Res. Vet. Sci. 2020, 129, 70–73. [Google Scholar] [CrossRef]
- Schiller, I.; Oesch, B.; Vordermeier, H.M.; Palmer, M.V.; Harris, B.N.; Orloski, K.A.; Buddle, B.M.; Thacker, T.C.; Lyashchenko, K.P.; Waters, W.R. Bovine tuberculosis: A review of current and emerging diagnostic techniques in view of their relevance for disease control and eradication. Transbound. Emerg. Dis. 2010, 57, 205–220. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Whelan, A.; Cockle, P.J.; Farrant, L.; Palmer, N.; Hewinson, R.G. Use of synthetic peptides derived from the antigens ESAT-6 and CFP-10 for differential diagnosis of bovine tuberculosis in cattle. Clin. Diagn. Lab. Immunol. 2001, 8, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Kleynhans, L.; Warren, R.M.; Goosen, W.J.; Miller, M.A. Cell-mediated immunological biomarkers and their diagnostic application in livestock and wildlife infected with Mycobacterium bovis. Front. Immunol. 2021, 12, 483. [Google Scholar] [CrossRef] [PubMed]
- Agriculture and Horticulture Development Board Development of a Deployable Tuberculosis Vaccine for Cattle. Available online: https://tbhub.co.uk/resources/frequently-asked-questions/development-of-a-deployable-tuberculosis-vaccine-for-cattle/ (accessed on 12 June 2022).
- Kaufmann, S.H.E. The TB vaccine development pipeline: Present and future priorities and challenges for research and innovation. In Essential Tuberculosis; Migliori, G.B., Raviglione, M.C., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 395–405. [Google Scholar]
- Grode, L.; Seiler, P.; Baumann, S.; Hess, J.; Brinkmann, V.; Eddine, A.N.; Mann, P.; Goosmann, C.; Bandermann, S.; Smith, D.; et al. Increased vaccine efficacy against tuberculosis of recombinant Mycobacterium bovis bacille Calmette-Guerin mutants that secrete listeriolysin. J. Clin. Investig. 2005, 115, 2472–2479. [Google Scholar] [CrossRef] [PubMed]
- Höner zu Bentrup, K.; Russell, D.G. Mycobacterial persistence: Adaptation to a changing environment. Trends Microbiol. 2001, 9, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuizen, N.E.; Kulkarni, P.S.; Shaligram, U.; Cotton, M.F.; Rentsch, C.A.; Eisele, B.; Grode, L.; Kaufmann, S.H.E. The Recombinant Bacille Calmette-Guerin Vaccine VPM1002: Ready for Clinical Efficacy Testing. Front. Immunol. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Saiga, H.; Nieuwenhuizen, N.; Gengenbacher, M.; Koehler, A.B.; Schuerer, S.; Moura-Alves, P.; Wagner, I.; Mollenkopf, H.J.; Dorhoi, A.; Kaufmann, S.H. The Recombinant BCG DeltaureC::hly Vaccine Targets the AIM2 Inflammasome to Induce Autophagy and Inflammation. J. Infect. Dis. 2015, 211, 1831–1841. [Google Scholar] [CrossRef]
- Grode, L.; Ganoza, C.A.; Brohm, C.; Weiner, J., 3rd; Eisele, B.; Kaufmann, S.H. Safety and immunogenicity of the recombinant BCG vaccine VPM1002 in a phase 1 open-label randomized clinical trial. Vaccine 2013, 31, 1340–1348. [Google Scholar] [CrossRef]
- Loxton, A.G.; Knaul, J.K.; Grode, L.; Gutschmidt, A.; Meller, C.; Eisele, B.; Johnstone, H.; van der Spuy, G.; Maertzdorf, J.; Kaufmann, S.H.E.; et al. Safety and Immunogenicity of the Recombinant Mycobacterium bovis BCG Vaccine VPM1002 in HIV-Unexposed Newborn Infants in South Africa. Clin. Vaccine Immunol. CVI 2017, 24, 00439-16. [Google Scholar] [CrossRef]
- Cotton, M.F.; Madhi, S.A.; Luabeya, A.K.; Tameris, M.; Hesseling, A.C.; Shenje, J.; Schoeman, E.; Hatherill, M.; Desai, S.; Kapse, D.; et al. Safety and immunogenicity of VPM1002 versus BCG in South African newborn babies: A randomised, phase 2 non-inferiority double-blind controlled trial. Lancet Infect. Dis. 2022, 22, 1472–1483. [Google Scholar] [CrossRef]
- Kaufmann, S.H.E. Vaccine development against tuberculosis over the last 140 years: Failure as part of success. Front. Microbiol. 2021, 12, 750124. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Vogelzang, A.; Schuerer, S.; Lazar, D.; Kaiser, P.; Kaufmann, S.H. Dietary pyridoxine controls efficacy of vitamin B6-auxotrophic tuberculosis vaccine bacillus Calmette-Guerin DeltaureC::hly Deltapdx1 in mice. Mbio 2014, 5, e01262-14. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Nieuwenhuizen, N.; Vogelzang, A.; Liu, H.; Kaiser, P.; Schuerer, S.; Lazar, D.; Wagner, I.; Mollenkopf, H.J.; Kaufmann, S.H. Deletion of nuoG from the Vaccine Candidate Mycobacterium bovis BCG DeltaureC::hly Improves Protection against Tuberculosis. Mbio 2016, 7, e00679-16. [Google Scholar] [CrossRef]
- Perez de Val, B.; Villarreal-Ramos, B.; Nofrarias, M.; Lopez-Soria, S.; Romera, N.; Singh, M.; Abad, F.X.; Xing, Z.; Vordermeier, H.M.; Domingo, M. Goats primed with Mycobacterium bovis BCG and boosted with a recombinant adenovirus expressing Ag85A show enhanced protection against tuberculosis. Clin. Vaccine Immunol. CVI 2012, 19, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Liang, Y.; Wu, X. Animal Models of Tuberculosis Vaccine Research: An Important Component in the Fight against Tuberculosis. BioMed Res. Int. 2020, 2020, 4263079. [Google Scholar] [CrossRef]
- Buddle, B.M.; Skinner, M.A.; Wedlock, D.N.; de Lisle, G.W.; Vordermeier, H.M.; Hewinson, R.G. Cattle as a model for development of vaccines against human tuberculosis. Tuberculosis 2005, 85, 19–24. [Google Scholar] [CrossRef]
- Liebler-Tenorio, E.M.; Heyl, J.; Wedlich, N.; Figl, J.; Köhler, H.; Krishnamoorthy, G.; Nieuwenhuizen, N.; Grode, L.; Kaufmann, S.H.E.; Menge, C. Vaccine-induced subcutaneous granulomas in goats reflect differences in host-mycobacterium interactions between BCG- and recombinant BCG-derivative vaccines. Int. J. Mol. Sci. 2022, 23, 10992. [Google Scholar] [CrossRef] [PubMed]
- Desel, C.; Dorhoi, A.; Bandermann, S.; Grode, L.; Eisele, B.; Kaufmann, S.H.E. Recombinant BCG Delta ureC hly plus Induces Superior Protection Over Parental BCG by Stimulating a Balanced Combination of Type 1 and Type 17 Cytokine Responses. J. Infect. Dis. 2011, 204, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Vogelzang, A.; Perdomo, C.; Zedler, U.; Kuhlmann, S.; Hurwitz, R.; Gengenbacher, M.; Kaufmann, S.H. Central memory CD4+ T cells are responsible for the recombinant Bacillus Calmette-Guerin DeltaureC::hly vaccine’s superior protection against tuberculosis. J. Infect. Dis. 2014, 210, 1928–1937. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Kaiser, P.; Schuerer, S.; Lazar, D.; Kaufmann, S.H. Post-exposure vaccination with the vaccine candidate Bacillus Calmette-Guerin DeltaureC::hly induces superior protection in a mouse model of subclinical tuberculosis. Microbes Infect. Inst. Pasteur 2016, 18, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, K.; Grode, L.; Chang, R.; Fitzpatrick, M.; Laddy, D.; Hokey, D.; Derrick, S.; Morris, S.; McCown, D.; Kidd, R.; et al. Nonclinical Development of BCG Replacement Vaccine Candidates. Vaccines 2013, 1, 120–138. [Google Scholar] [CrossRef]
- Pérez de Val, B.; Vidal, E.; López-Soria, S.; Marco, A.; Cervera, Z.; Martín, M.; Mercader, I.; Singh, M.; Raeber, A.; Domingo, M. Assessment of safety and interferon gamma responses of Mycobacterium bovis BCG vaccine in goat kids and milking goats. Vaccine 2016, 34, 881–886. [Google Scholar] [CrossRef]
- Valheim, M.; Hasvold, H.J.; Storset, A.K.; Larsen, H.J.S.; Press, C.M. Localisation of CD25+ cells and MHCII+ cells in lymph nodes draining Mycobacterium avium subsp. paratuberculosis vaccination granuloma and the presence of a systemic immune response. Res. Vet. Sci. 2002, 73, 77–85. [Google Scholar]
- Stau, A.; Ganter, M. Immune reactions to and side effects of a vaccination against paratuberculosis in milk goats. Tierarztl Prax Ausg G Grosstiere Nutztiere 2012, 40, 14–20. [Google Scholar] [PubMed]
- Vortel, V. Pathology of BCG vaccine. Am. Rev. Respir. Dis. 1962, 86, 336–349. [Google Scholar]
- Al-Bhlal, L.A. Pathologic findings for bacille Calmette-Guérin infections in immunocompetent and immunocompromised patients. Am. J. Clin. Pathol. 2000, 113, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.; Cotton, M.F.; Eisele, B.; Gengenbacher, M.; Grode, L.; Hesseling, A.C.; Walzl, G. The BCG replacement vaccine VPM1002: From drawing board to clinical trial. Expert Rev. Vaccines 2014, 13, 619–630. [Google Scholar] [CrossRef]
- Rao, M.; Vogelzang, A.; Kaiser, P.; Schuerer, S.; Kaufmann, S.H.; Gengenbacher, M. The tuberculosis vaccine candidate Bacillus Calmette-Guerin DeltaureC::hly coexpressing human interleukin-7 or -18 enhances antigen-specific T cell responses in mice. PloS ONE 2013, 8, e78966. [Google Scholar] [CrossRef] [PubMed]
- Rentsch, C.A.; Thalmann, G.N.; Lucca, I.; Kwiatkowski, M.; Wirth, G.J.; Strebel, R.T.; Engeler, D.; Pedrazzini, A.; Hüttenbrink, C.; Schultze-Seemann, W.; et al. A Phase 1/2 Single-arm Clinical Trial of Recombinant Bacillus Calmette-Guérin (BCG) VPM1002BC Immunotherapy in Non-muscle-invasive Bladder Cancer Recurrence After Conventional BCG Therapy: SAKK 06/14. Eur. Urol. Oncol. 2022, 5, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wedlock, D.N.; Denis, M.; Vordermeier, H.M.; Hewinson, R.G.; Buddle, B.M. Vaccination of cattle with Danish and Pasteur strains of Mycobacterium bovis BCG induce different levels of IFNgamma post-vaccination, but induce similar levels of protection against bovine tuberculosis. Vet. Immunol. Immunopathol. 2007, 118, 50–58. [Google Scholar] [CrossRef]
- Hope, J.C.; Thom, M.L.; McAulay, M.; Mead, E.; Vordermeier, H.M.; Clifford, D.; Hewinson, R.G.; Villarreal-Ramos, B. Identification of surrogates and correlates of protection in protective immunity against Mycobacterium bovis infection induced in neonatal calves by vaccination with M. bovis BCG Pasteur and M. bovis BCG Danish. Clin. Vaccine Immunol. CVI 2011, 18, 373–379. [Google Scholar] [CrossRef]
- Waters, W.R.; Palmer, M.V.; Nonnecke, B.J.; Thacker, T.C.; Scherer, C.F.; Estes, D.M.; Hewinson, R.G.; Vordermeier, H.M.; Barnes, S.W.; Federe, G.C.; et al. Efficacy and immunogenicity of Mycobacterium bovis DeltaRD1 against aerosol M. bovis infection in neonatal calves. Vaccine 2009, 27, 1201–1209. [Google Scholar] [CrossRef][Green Version]
- Blanco, F.C.; Soria, M.; Gravisaco, M.J.; Bianco, M.V.; Meikle, V.; Garbaccio, S.; Vagnoni, L.; Cataldi, A.A.; Bigi, F. Assessment of the immune responses induced in cattle after inoculation of a Mycobacterium bovis strain deleted in two mce2 genes. J. Biomed. Biotechnol. 2012, 2012, 258353. [Google Scholar] [CrossRef] [PubMed]
- Vordermeier, H.M.; Villarreal-Ramos, B.; Cockle, P.J.; McAulay, M.; Rhodes, S.G.; Thacker, T.; Gilbert, S.C.; McShane, H.; Hill, A.V.; Xing, Z.; et al. Viral booster vaccines improve Mycobacterium bovis BCG-induced protection against bovine tuberculosis. Infect. Immun. 2009, 77, 3364–3373. [Google Scholar] [CrossRef] [PubMed]
- Buddle, B.M.; Hewinson, R.G.; Vordermeier, H.M.; Wedlock, D.N. Subcutaneous administration of a 10-fold-lower dose of a commercial human tuberculosis vaccine, Mycobacterium bovis bacillus Calmette-Guerin Danish, induced levels of protection against bovine tuberculosis and responses in the tuberculin intradermal test similar to those induced by a standard cattle dose. Clin. Vaccine Immunol. CVI 2013, 20, 1559–1562. [Google Scholar] [PubMed]
- Griffin, J.F.; Mackintosh, C.G.; Slobbe, L.; Thomson, A.J.; Buchan, G.S. Vaccine protocols to optimise the protective efficacy of BCG. Tuber. Lung Dis. Off. J. Int. Union Against Tuberc. Lung Dis. 1999, 79, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Panton, K.K.; Farup, P.G.; Sagen, E.; Sirum, U.F.; Asberg, A. Vitamin B6 in plasma—Sample stability and the reference limits. Scand. J. Clin. Lab. Investig. 2013, 73, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Cheng, C.-H.; Hsu, C.-L.; Lee, W.-J.; Huang, S.-C.; Huang, Y.-C. Role of vitamin B6 status on antioxidant defenses, glutathione, and related enzyme activities in mice with homocysteine-induced oxidative stress. Food Nutr. Res. 2015, 59, 25702. [Google Scholar] [CrossRef]
- Waters, W.R.; Palmer, M.V.; Thacker, T.C.; Davis, W.C.; Sreevatsan, S.; Coussens, P.; Meade, K.G.; Hope, J.C.; Estes, D.M. Tuberculosis immunity: Opportunities from studies with cattle. Clin. Dev. Immunol. 2011, 2011, 768542. [Google Scholar] [CrossRef]
- Maggioli, M.F.; Palmer, M.V.; Thacker, T.C.; Vordermeier, H.M.; Waters, W.R. Characterization of effector and memory T cell subsets in the immune response to bovine tuberculosis in cattle. PloS ONE 2015, 10, e0122571. [Google Scholar] [CrossRef]
- McGill, J.L.; Sacco, R.E.; Baldwin, C.L.; Telfer, J.C.; Palmer, M.V.; Waters, W.R. The role of gamma delta T cells in immunity to Mycobacterium bovis infection in cattle. Vet. Immunol. Immunopathol. 2014, 159, 133–143. [Google Scholar] [CrossRef]
- Yirsaw, A.; Baldwin, C.L. Goat γδ T cells. Dev. Comp. Immunol. 2021, 114, 103809. [Google Scholar] [CrossRef]
- Caro, M.R.; Gallego, M.C.; Buendía, A.J.; Del Rio, L.; Seva, J.; Navarro, J.A. Differences in lymphocyte subpopulations from peripheral blood and lymphoid organs in natural caprine tuberculosis infection. J. Vet. Medicine. B Infect. Dis. Vet. Public Health 2001, 48, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Seva, J.; Hernández, D.; Bernabé, A.; Pallarés, F.J.; Navarro, J.A. Immunophenotypical characterization of the lymphocyte infiltrate in caprine pulmonary tuberculosis. J. Comp. Pathol. 2000, 123, 96–103. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE) Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2021. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access (accessed on 20 June 2022).
- Wiker, H.G. MPB70 and MPB83--major antigens of Mycobacterium bovis. Scand. J. Immunol. 2009, 69, 492–499. [Google Scholar] [CrossRef]
- Tanner, R.; Villarreal-Ramos, B.; Vordermeier, H.M.; McShane, H. The Humoral Immune Response to BCG Vaccination. Front. Immunol. 2019, 10, 1317. [Google Scholar] [CrossRef] [PubMed]
- Köhler, H.; Soschinka, A.; Meyer, M.; Kather, A.; Reinhold, P.; Liebler-Tenorio, E. Characterization of a caprine model for the subclinical initial phase of Mycobacterium avium subsp. paratuberculosis infection. BMC Vet. Res. 2015, 11, 74. [Google Scholar]
- Wangoo, A.; Johnson, L.; Gough, J.; Ackbar, R.; Inglut, S.; Hicks, D.; Spencer, Y.; Hewinson, G.; Vordermeier, M. Advanced granulomatous lesions in Mycobacterium bovis-infected cattle are associated with increased expression of type I procollagen, gammadelta (WC1+) T cells and CD 68+ cells. J. Comp. Pathol. 2005, 133, 223–234. [Google Scholar] [CrossRef]
- Friedrich-Loeffler-Institut Tuberkulose der Rinder: Mycobacterium bovis und Mycobacterium caprae. Amtliche Methode und Falldefinition. Available online: https://www.openagrar.de/receive/openagrar_mods_00054079 (accessed on 22 June 2022).
- Warren, R.M.; Gey van Pittius, N.C.; Barnard, M.; Hesseling, A.; Engelke, E.; de Kock, M.; Gutierrez, M.C.; Chege, G.K.; Victor, T.C.; Hoal, E.G.; et al. Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int. J. Tuberc. Lung Dis. Off. J. Int. Union Against Tuberc. Lung Dis. 2006, 10, 818–822. [Google Scholar]
- Guerrero, C.; Bernasconi, C.; Burki, D.; Bodmer, T.; Telenti, A. A novel insertion element from Mycobacterium avium, IS1245, is a specific target for analysis of strain relatedness. J. Clin. Microbiol. 1995, 33, 304–307. [Google Scholar] [CrossRef]
- Kunze, Z.M.; Portaels, F.; McFadden, J.J. Biologically distinct subtypes of Mycobacterium avium differ in possession of insertion sequence IS901. J. Clin. Microbiol. 1992, 30, 2366–2372. [Google Scholar] [CrossRef]
- Englund, S.; Ballagi-Pordány, A.; Bölske, G.; Johansson, K.E. Single PCR and nested PCR with a mimic molecule for detection of Mycobacterium avium subsp. paratuberculosis. Diagn. Microbiol. Infect. Dis. 1999, 33, 163–171. [Google Scholar] [CrossRef]
- Kirschner, P.; Rosenau, J.; Springer, B.; Teschner, K.; Feldmann, K.; Böttger, E.C. Diagnosis of mycobacterial infections by nucleic acid amplification: 18-month prospective study. J. Clin. Microbiol. 1996, 34, 304–312. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination Group | |||||
|---|---|---|---|---|---|
| Weeks Post Vaccination (pv) | Mock-Treated | BCG | VPM1002 | NUOG | PDX |
| −1 | 0.0 (0; 0) | 0.0 (0; 0) | 0.0 (0; 0) | 0.0 (0; 0) | 0.0 (0; 0) |
| 1 | 0.0 (0; 0) | 2.0 (1; 3) | 2.2 (0; 4) | 2.2 (0; 4) | 2.2 (1; 3) |
| 2 | 0.0 (0; 0) | 3.2 (2; 5) | 5.0 (3; 7) | 5.7 (5; 6) | 3.8 (3; 5) |
| 3 | 0.0 (0; 0) | 3.5 (2; 5) | 5.5 (4; 7) | 5.7 (5; 6) | 3.5 (0; 6) |
| 4 | 0.0 (0; 0) | 3.7 (2; 5) | 5.2 (4; 6) | 5.8 (5; 7) | 3.3 (0; 6) |
| 5 | 0.0 (0; 0) | 4.0 (3; 6) | 5.3 (4; 6) | 5.8 (5; 7) | 3.2 (0; 5) |
| 6 | 0.0 (0; 0) | 4.0 (3; 6) | 5.0 (3; 6) | 6.0 (4; 8) | 3.7 (2; 5) |
| 7 | 0.0 (0; 0) | 3.5 (2; 5) | 4.5 (3; 5) | 5.7 (4; 8) | 3.3 (2; 5) |
| 8 | 0.0 (0; 0) | 3.2 (2; 5) | 4.0 (3; 5) | 5.2 (4; 7) | 2.8 (2; 5) |
| 9 | 0.0 (0; 0) | 3.0 (2; 4) | 4.2 (3; 5) | 4.3 (4; 5) | 2.7 (2; 4) |
| 10 | 0.0 (0; 0) | 3.0 (2; 5) | 3.8 (3; 5) | 4.5 (4; 5) | 2.5 (2; 3) |
| 11 | 0.0 (0; 0) | 3.2 (2; 5) | 4.2 (3; 5) | 4.3 (4; 5) | 2.7 (2; 4) |
| 12 | 0.0 (0; 0) | 3.2 (2; 5) | 4.0 (3; 5) | 3.5 (3; 4) | 2.7 (2; 4) |
| 13 | 0.0 (0; 0) | 3.0 (2; 5) | 3.7 (3; 5) | 3.5 (3; 4) | 2.8 (2; 4) |
| 14 | 0.0 (0; 0) | 3.0 (2; 5) | 3.8 (3; 5) | 3.3 (3; 4) | 3.2 (2; 5) |
| 15 | 0.0 (0; 0) | 3.2 (2; 5) | 3.5 (3; 5) | 3.3 (3; 4) | 3.0 (2; 5) |
| 16 | 0.0 (0; 0) | 3.2 (2; 5) | 3.5 (3; 5) | 3.5 (3; 4) | 2.7 (1; 5) |
| 17 | 0.0 (0; 0) | 3.0 (2; 5) | 3.5 (3; 5) | 3.7 (3; 5) | 2.3 (0; 5) |
| 18 | 0.0 (0; 0) | 3.0 (2; 5) | 3.2 (2; 5) | 3.5 (3; 4) | 1.8 (0; 4) |
| Number of Positive Goats/Number of Goats Per Group | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Vaccination Site | Ln. cervicalis superficialis sin. | Ln. axillaris profundus sin. | Lnn. mediastinales | ||||||||||
| Granuloma | AFB | Cultivation of Vaccine Strains | Granuloma | AFB | Cultivation of Vaccine Strains | Granuloma | AFB | Cultivation of Vaccine Strains | Granuloma/AFB | Cultivation of Vaccine Strains | ||||
| mock-treated | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | |||
| BCG | 6/6 | 6/6 | 3/6 | 1/6 | 1/6 | 0/6 | 1/6 | 1/6 | 0/6 | 0/6 | 1/6 | |||
| VPM1002 | 6/6 | 6/6 | 2/6 | 1/6 | 1/6 | 0/6 | 1/6 | 1/6 | 0/6 | 0/6 | 0/6 | |||
| NUOG | 6/6 | 5/6 | 1/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | 0/6 | |||
| PDX | 5/6 | 4/6 | 0/6 | 2/6 | 2/6 | 0/6 | 1/6 | 0/6 | 0/6 | 0/6 | 0/6 | |||
| Symptom | Description | Score |
|---|---|---|
| size of the lesion | no lesion | 0 |
| <2 cm diameter | 1 | |
| 2–5 cm diameter | 2 | |
| 5–10 cm diameter | 3 | |
| >10 cm diameter | 4 | |
| edematous swelling | no | 0 |
| yes | 1 | |
| color | physiological | 0 |
| redness, other color | 1 | |
| texture | physiological | 0 |
| alteration | 1 | |
| pain | no | 0 |
| yes | 1 | |
| local temperature | normal | 0 |
| increased | 1 | |
| necrosis | no | 0 |
| yes | 5 | |
| suppuration | no | 0 |
| yes | 5 | |
| total score | no reaction | 0 |
| mild reaction | 1–4 | |
| medium reaction | 4–8 | |
| strong reaction | 9–14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figl, J.; Köhler, H.; Wedlich, N.; Liebler-Tenorio, E.M.; Grode, L.; Parzmair, G.; Krishnamoorthy, G.; Nieuwenhuizen, N.E.; Kaufmann, S.H.E.; Menge, C. Safety and Immunogenicity of Recombinant Bacille Calmette-Guérin Strain VPM1002 and Its Derivatives in a Goat Model. Int. J. Mol. Sci. 2023, 24, 5509. https://doi.org/10.3390/ijms24065509
Figl J, Köhler H, Wedlich N, Liebler-Tenorio EM, Grode L, Parzmair G, Krishnamoorthy G, Nieuwenhuizen NE, Kaufmann SHE, Menge C. Safety and Immunogenicity of Recombinant Bacille Calmette-Guérin Strain VPM1002 and Its Derivatives in a Goat Model. International Journal of Molecular Sciences. 2023; 24(6):5509. https://doi.org/10.3390/ijms24065509
Chicago/Turabian StyleFigl, Julia, Heike Köhler, Nadine Wedlich, Elisabeth M. Liebler-Tenorio, Leander Grode, Gerald Parzmair, Gopinath Krishnamoorthy, Natalie E. Nieuwenhuizen, Stefan H. E. Kaufmann, and Christian Menge. 2023. "Safety and Immunogenicity of Recombinant Bacille Calmette-Guérin Strain VPM1002 and Its Derivatives in a Goat Model" International Journal of Molecular Sciences 24, no. 6: 5509. https://doi.org/10.3390/ijms24065509
APA StyleFigl, J., Köhler, H., Wedlich, N., Liebler-Tenorio, E. M., Grode, L., Parzmair, G., Krishnamoorthy, G., Nieuwenhuizen, N. E., Kaufmann, S. H. E., & Menge, C. (2023). Safety and Immunogenicity of Recombinant Bacille Calmette-Guérin Strain VPM1002 and Its Derivatives in a Goat Model. International Journal of Molecular Sciences, 24(6), 5509. https://doi.org/10.3390/ijms24065509