
PPAR Alpha Activation by Clofibrate Alleviates Ischemia/Reperfusion Injury in Metabolic Syndrome Rats by Decreasing Cardiac Inflammation and Remodeling and by Regulating the Atrial Natriuretic Peptide Compensatory Response

 , , ,
, , ,
Abstract
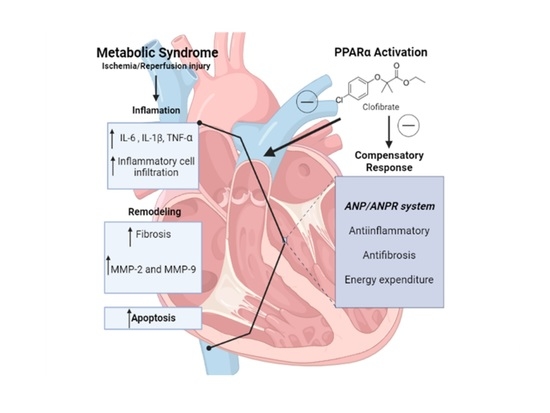
1. Introduction
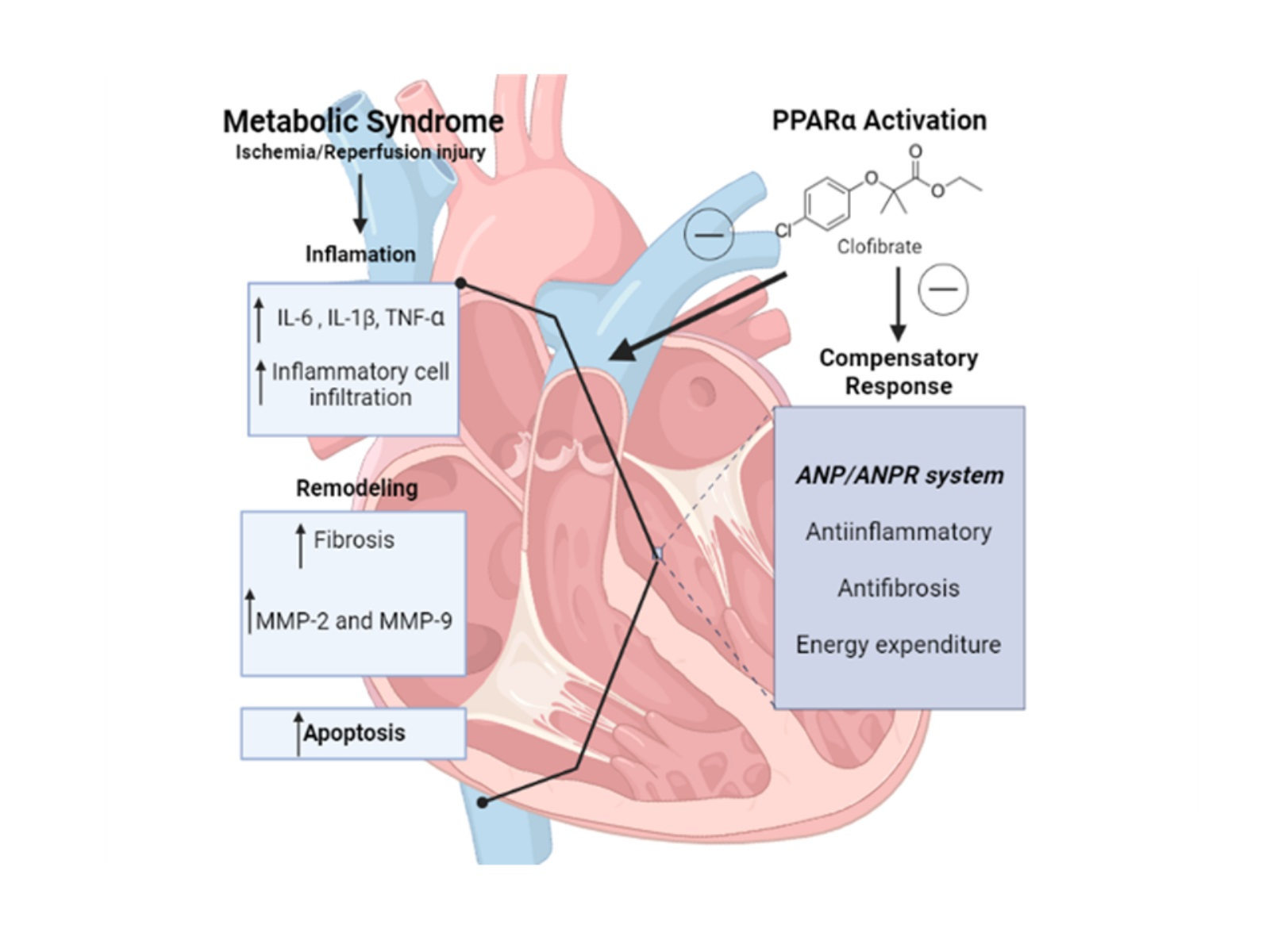
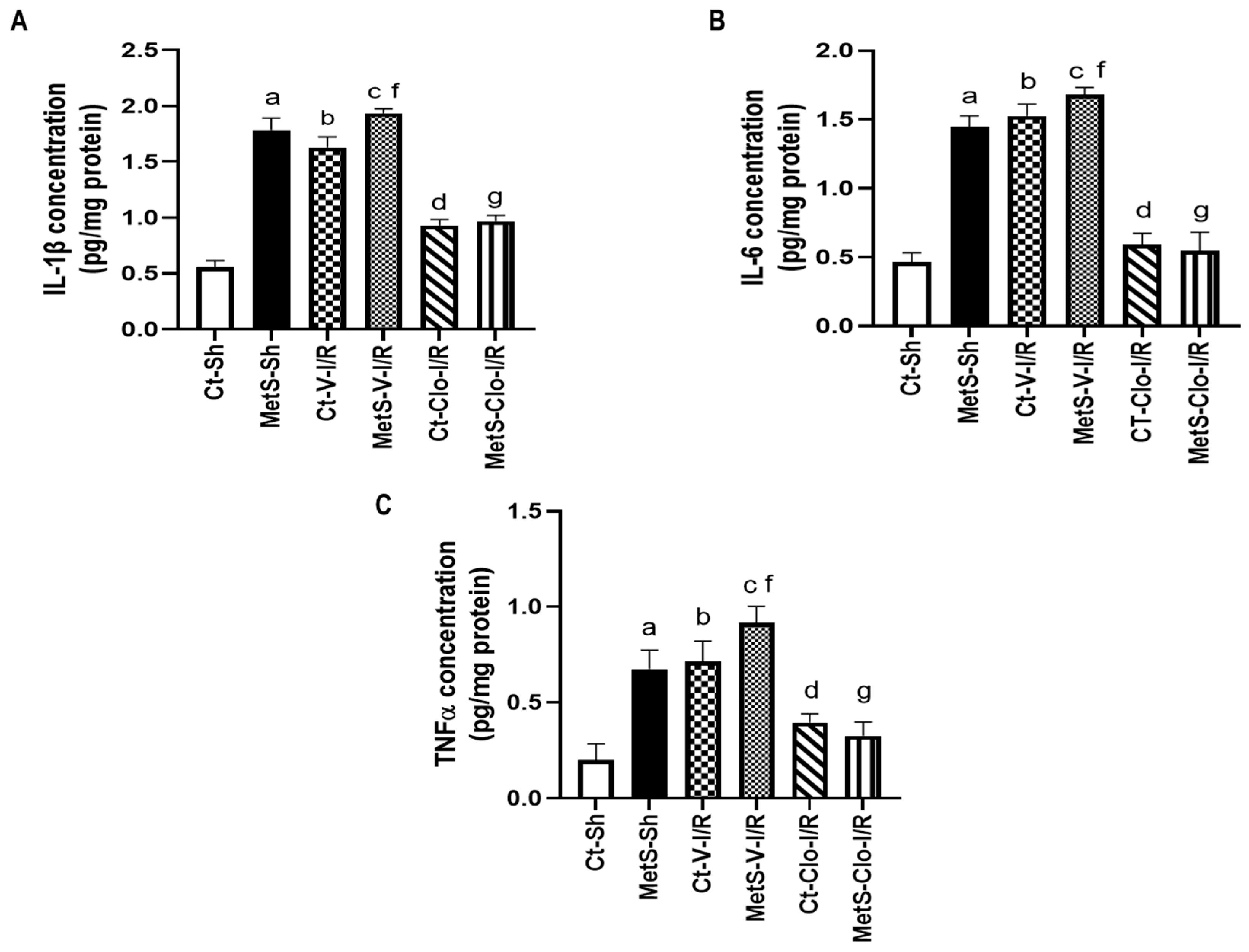
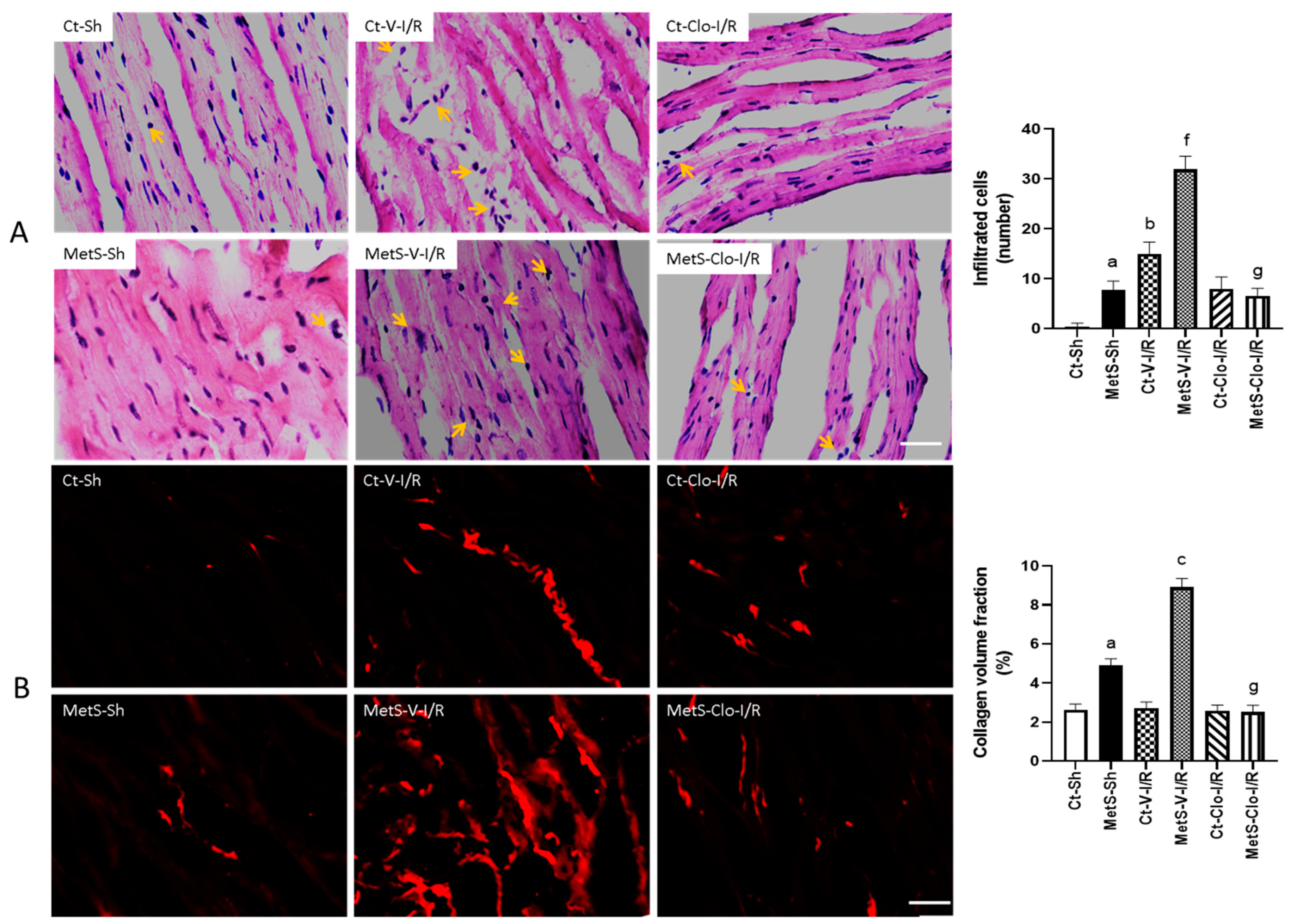
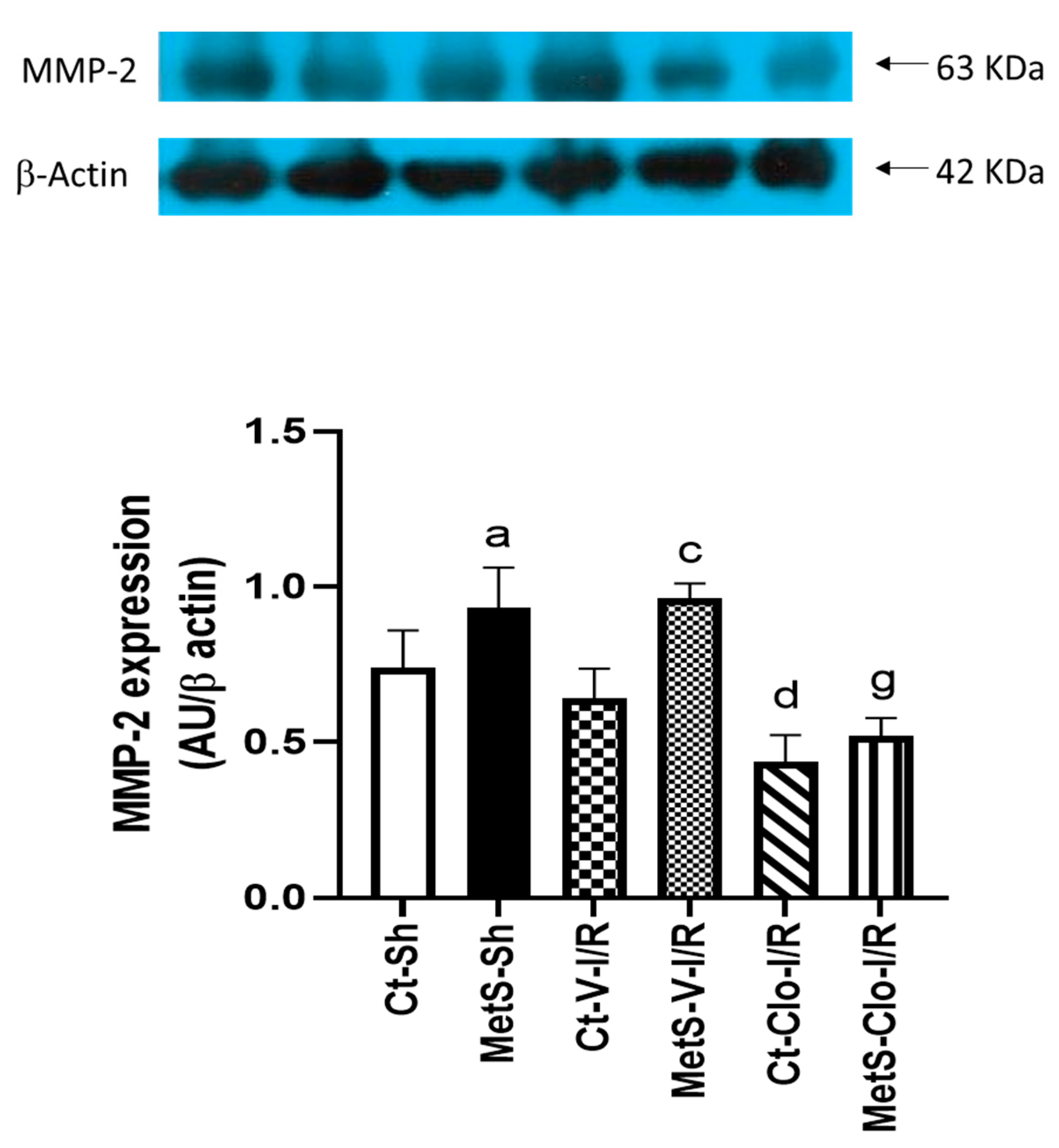
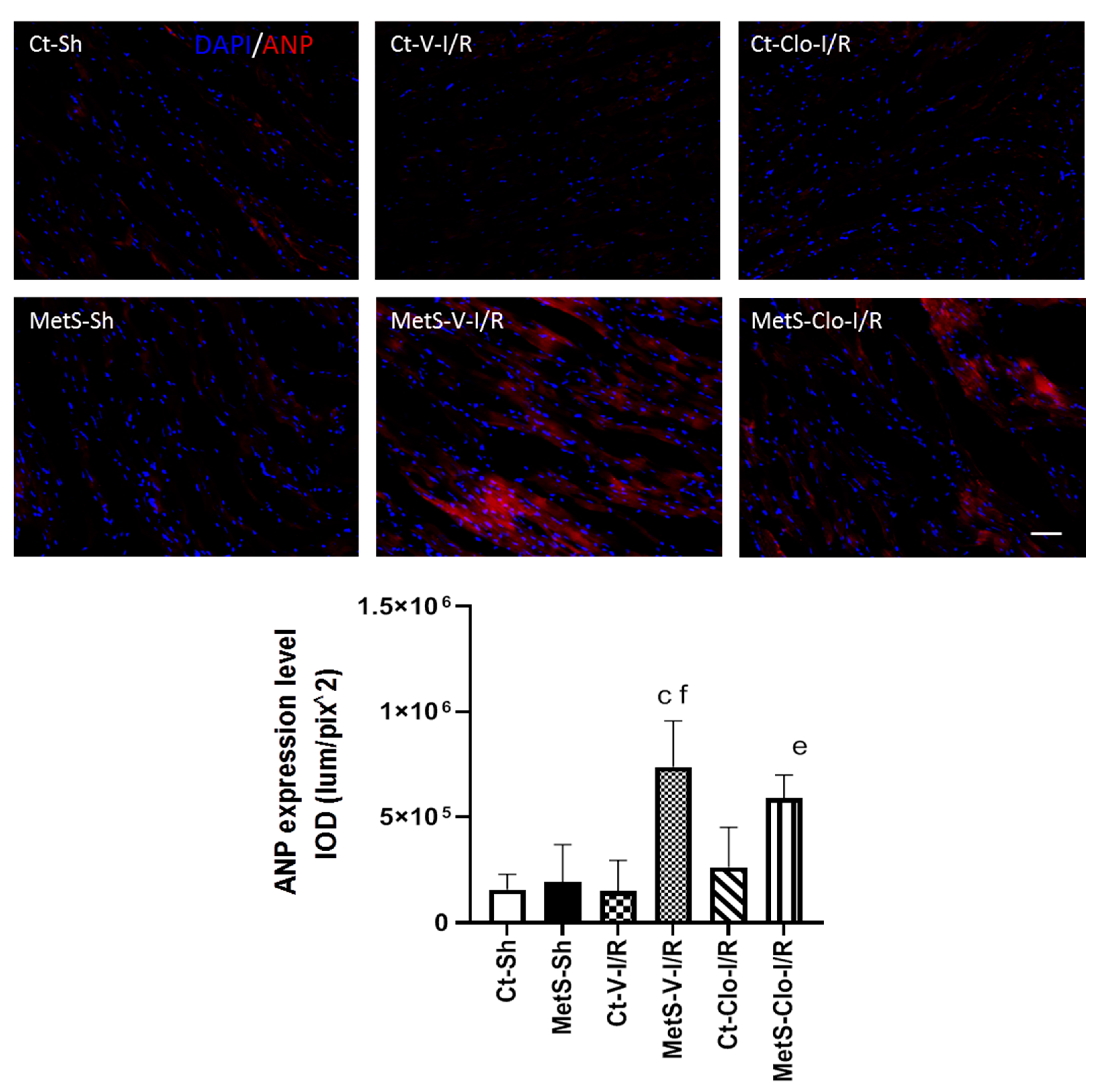
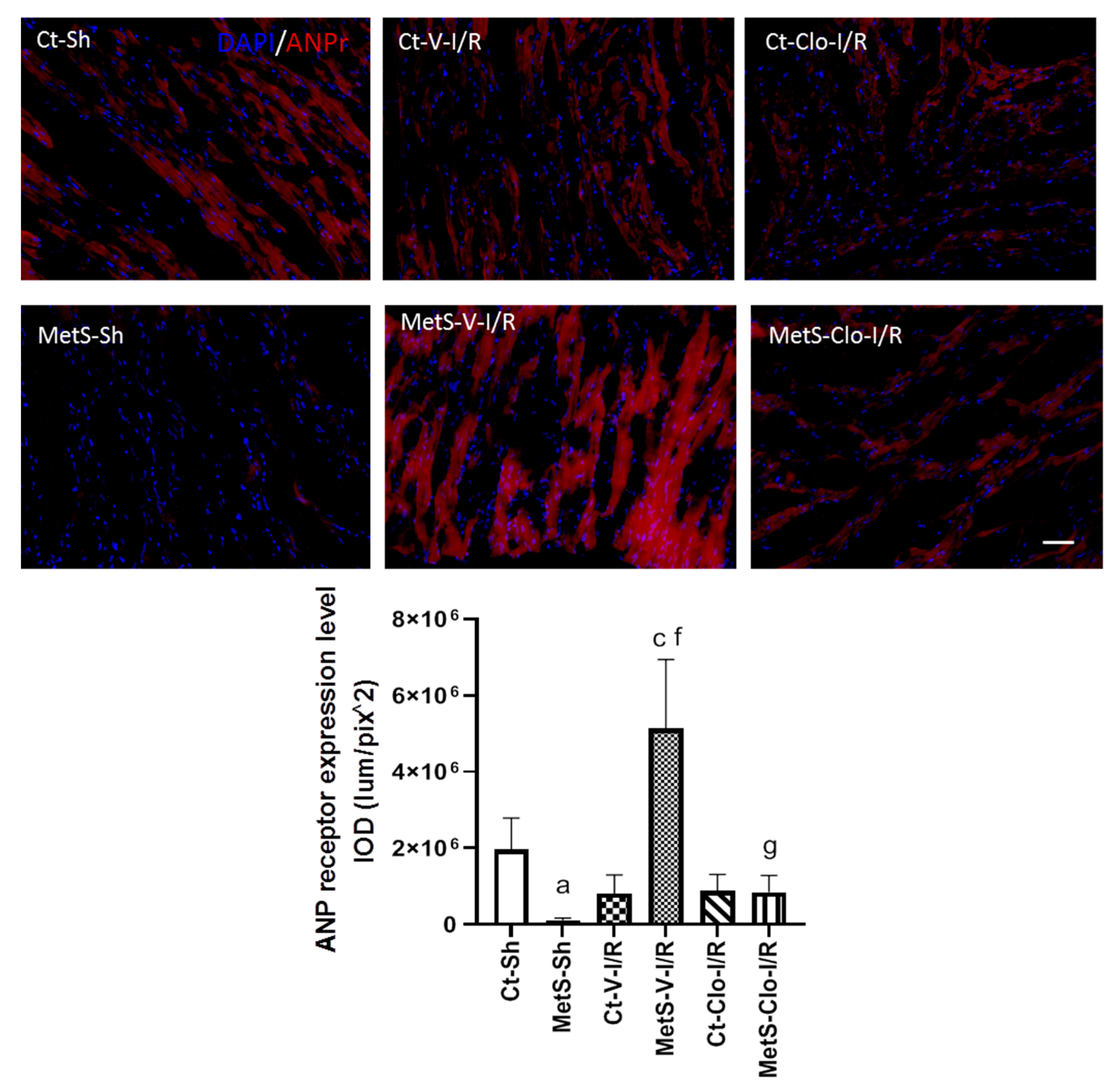
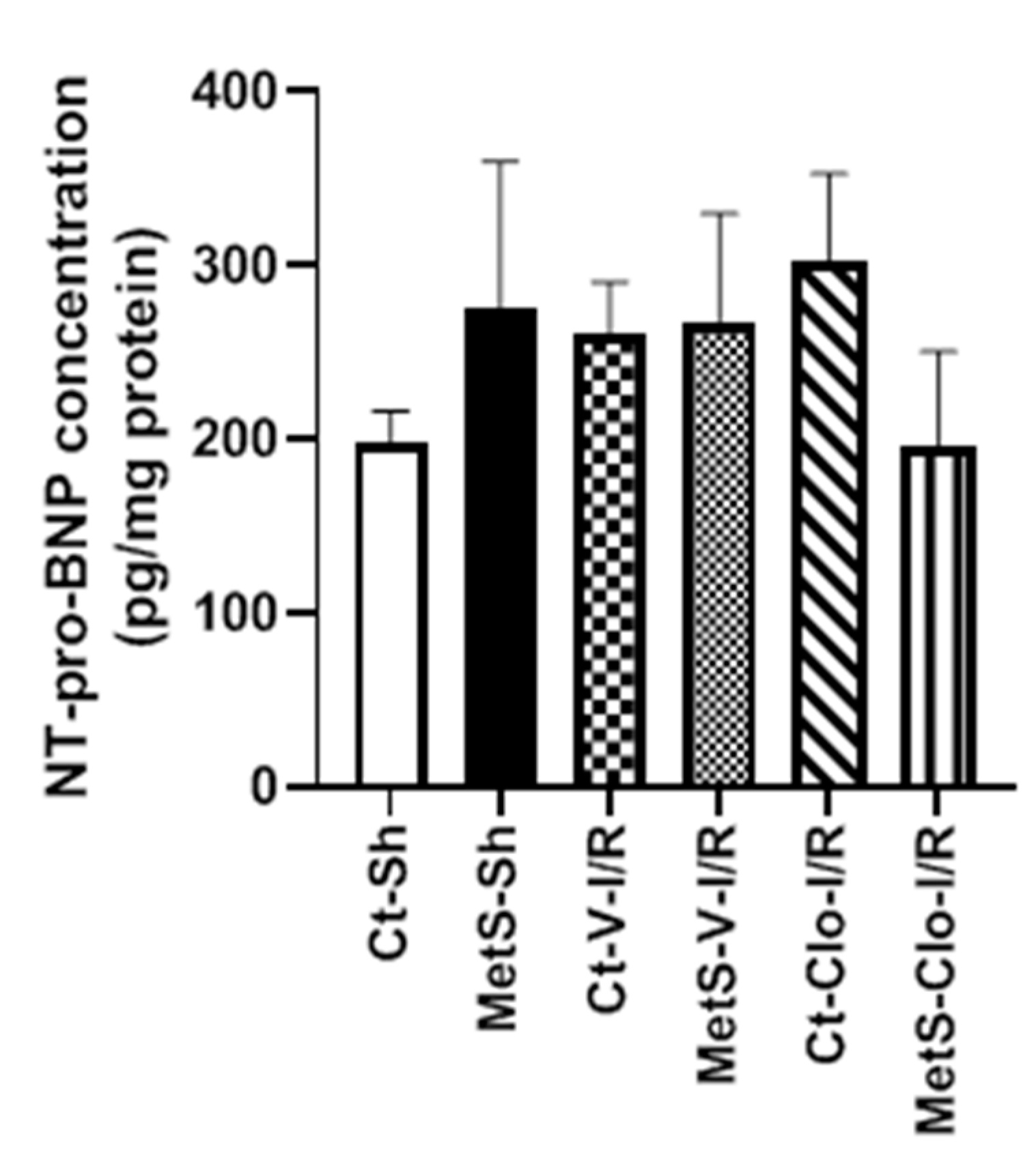
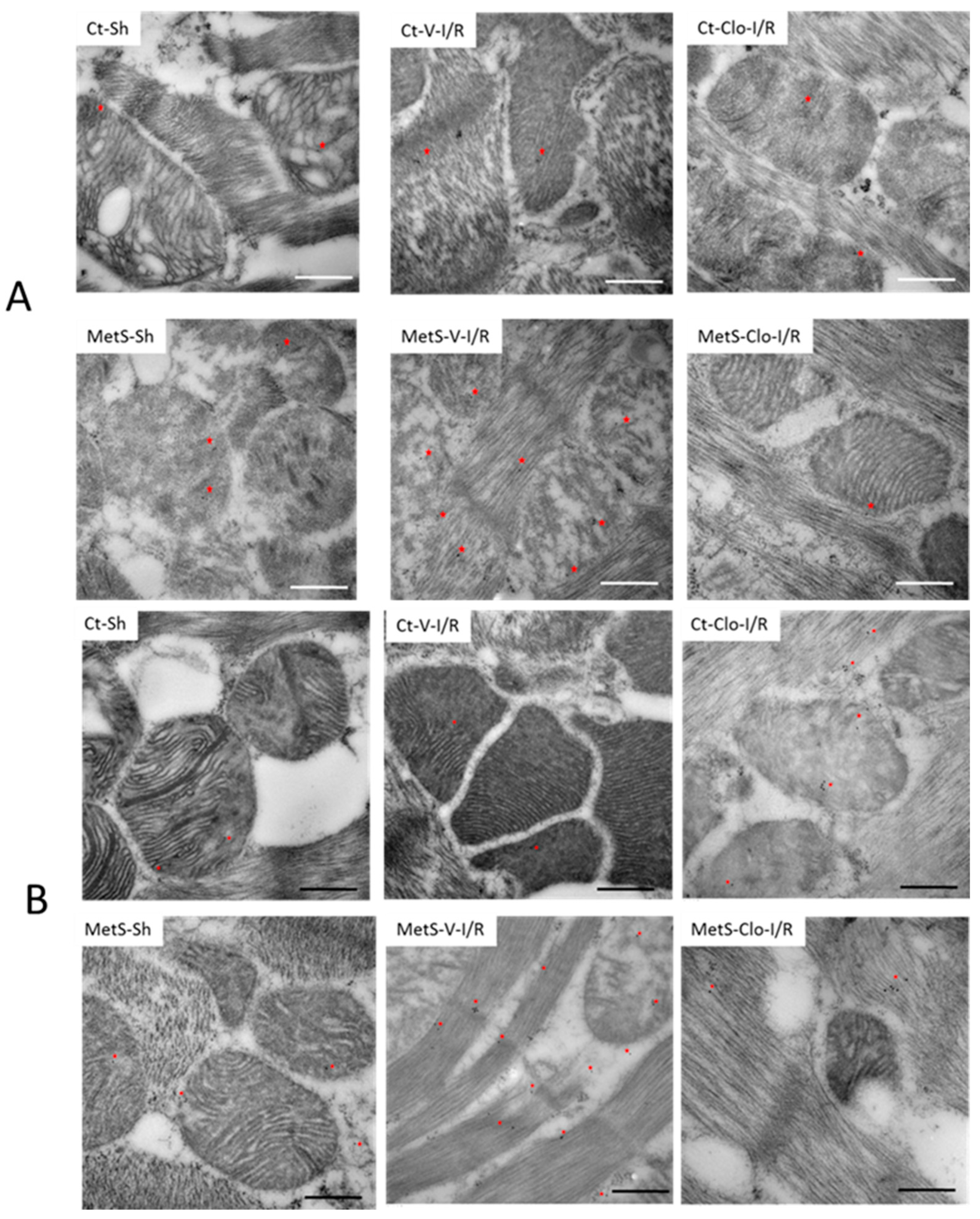
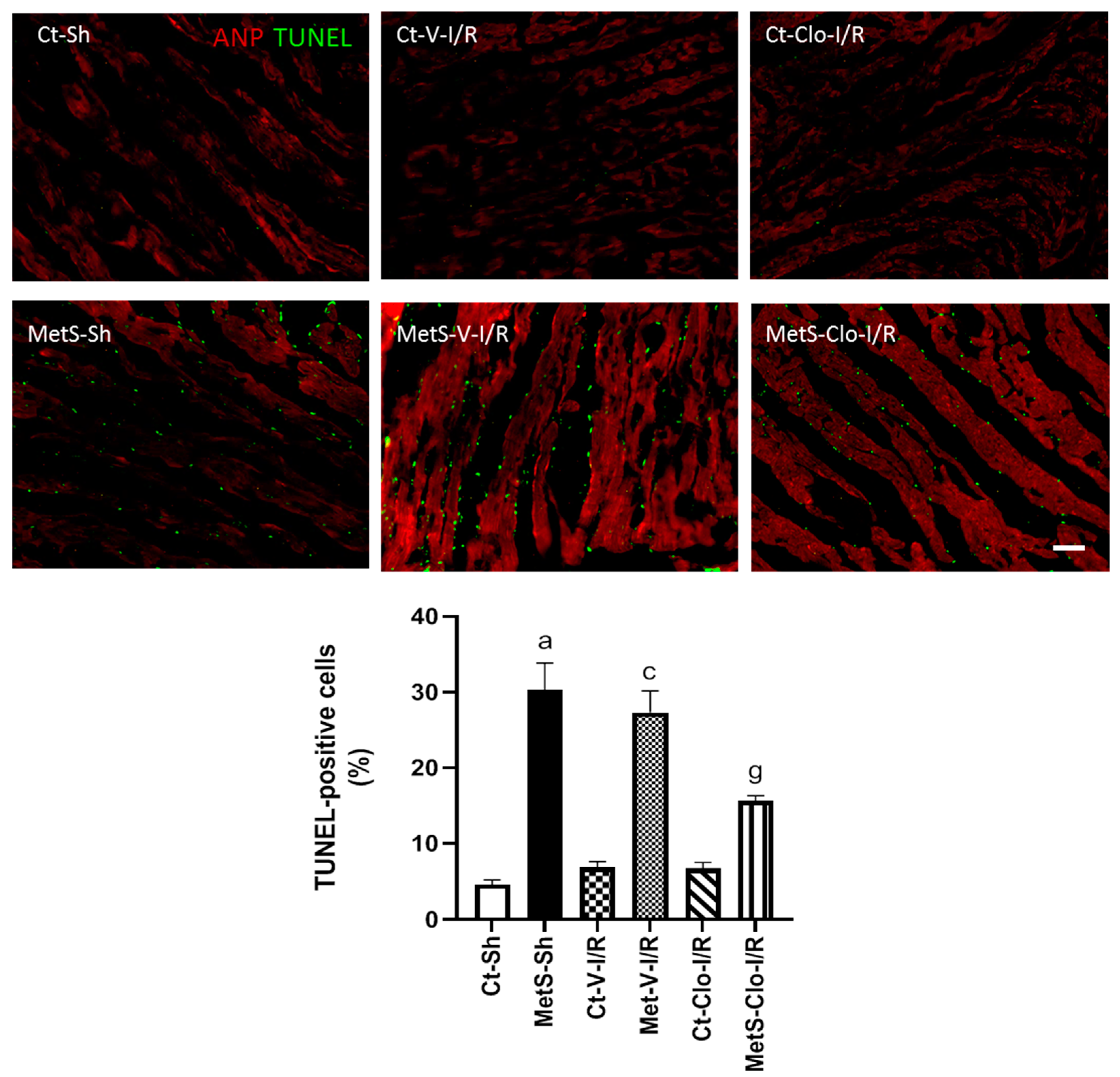
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Ischemia Reperfusion Model
4.3. Serum Biochemical Variables
4.4. Cardiac Cytokines and NT-proBNP Quantification
4.5. Western Blot
4.6. Histology
4.7. Hematoxylin–Eosin Stain (HE) and Picrosirius Red (PSR)
4.8. Immunodetection of Atrial Natriuretic Peptide (ANP) and Atrial Natriuretic Peptide Receptor (ANPr)
4.9. TUNEL Test
4.10. Image Acquisition, Number of Cell Infiltrate, Positive TUNEL Cells, and ANP and ANPr Intensity Quantification
4.11. Immune Colloidal Gold Technique
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shipman, K.E.; Strange, R.C.; Ramachandran, S. Use of fibrates in the metabolic syndrome: A review. World J. Diabetes 2016, 7, 74–88. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Lee, E.Y.; Han, K.; Kim, D.H.; Park, Y.M.; Kwon, H.S.; Yoon, K.H.; Kim, M.K.; Lee, S.H. Exposure-weighted scoring for metabolic syndrome and the risk of myocardial infarction and stroke: A nationwide population-based study. Cardiovasc. Diabetol. 2020, 19, 153. [Google Scholar] [CrossRef]
- Lafuse, W.P.; Wozniak, D.J.; Rajaram, M.V.S. Role of cardiac macrophages on cardiac inflammation, fibrosis and tissue repair. Cells 2020, 10, 51. [Google Scholar] [CrossRef]
- Krylatov, A.V.; Tsibulnikov, S.Y.; Mukhomedzyanov, A.V.; Boshchenko, A.A.; Goldberg, V.E.; Jaggi, A.S.; Erben, R.G.; Maslov, L.N. The role of natriuretic peptides in the regulation of cardiac tolerance to ischemia/reperfusion and postinfarction heart remodeling. J. Cardiovasc. Pharmacol. Ther. 2021, 26, 131–148. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, J.; Rong, J. Pharmacological modulation of cardiac remodeling after myocardial infarction. Oxid. Med. Cell Longev. 2020, 2020, 8815349. [Google Scholar] [CrossRef]
- Ibrahim, N.E.; Januzzi, J.L., Jr. Established and emerging roles of biomarkers in heart failure. Circ. Res. 2018, 123, 614–629. [Google Scholar] [CrossRef]
- Burley, D.S.; Hamid, S.A.; Baxter, G.F. Cardioprotective actions of peptide hormones in myocardial ischemia. Heart Fail. Rev. 2007, 12, 279–291. [Google Scholar] [CrossRef]
- Verboven, K.; Hansen, D.; Jocken, J.W.E.; Blaak, E.E. Natriuretic peptides in the control of lipid metabolism and insulin sensitivity. Obes. Rev. 2017, 18, 1243–1259. [Google Scholar] [CrossRef]
- Schlueter, N.; de Sterke, A.; Willmes, D.M.; Spranger, J.; Jordan, J.; Birkenfeld, A.L. Metabolic actions of natriuretic peptides and therapeutic potential in the metabolic syndrome. Pharmacol. Ther. 2014, 144, 12–27. [Google Scholar] [CrossRef]
- Santhekadur, P.K.; Kumar, D.P.; Seneshaw, M.; Mirshahi, F.; Sanyal, A.J. The multifaceted role of natriuretic peptides in metabolic syndrome. Biomed. Pharmacother. 2017, 92, 826–835. [Google Scholar] [CrossRef]
- D’Souza, S.P.; Davis, M.; Baxter, G.F. Autocrine and paracrine actions of natriuretic peptides in the heart. Pharmacol. Ther. 2004, 101, 113–129. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Nishikimi, T.; Kuwahara, K. Atrial and brain natriuretic peptides: Hormones secreted from the heart. Peptides 2019, 111, 18–25. [Google Scholar] [CrossRef]
- Rubattu, S.; Sciarretta, S.; Ciavarella, G.M.; Venturelli, V.; De Paolis, P.; Tocci, G.; De Biase, L.; Ferrucci, A.; Volpe, M. Reduced levels of N-terminal-proatrial natriuretic peptide in hypertensive patients with metabolic syndrome and their relationship with left ventricular mass. J. Hypertens. 2007, 25, 833–839. [Google Scholar] [CrossRef]
- Ramos, H.R.; Birkenfeld, A.L.; de Bold, A.J. Cardiac natriuretic peptides and obesity: Perspectives from an endocrinologist and a cardiologist. Endocr. Connect. 2015, 4, R25–R36. [Google Scholar] [CrossRef]
- Li, S.; Yang, B.; Du, Y.; Lin, Y.; Liu, J.; Huang, S.; Zhang, A.; Jia, Z.; Zhang, Y. Targeting PPARα for the treatment and understanding of cardiovascular diseases. Cell. Physiol. Biochem. 2018, 51, 2760–2775. [Google Scholar] [CrossRef]
- Ibarra-Lara, L.; Sánchez-Aguilar, M.; Soria-Castro, E.; Vargas-Barrón, J.; Roldán, F.J.; Pavón, N.; Torres-Narváez, J.C.; Cervantes-Pérez, L.G.; Pastelín-Hernández, G.; Sánchez-Mendoza, A. Clofibrate treatment decreases inflammation and reverses myocardial infarction-induced remodelation in a rodent experimental model. Molecules 2019, 24, 270. [Google Scholar] [CrossRef]
- Ibarra-Lara, L.; Sánchez-Aguilar, M.; Sánchez-Mendoza, A.; Del Valle-Mondragón, L.; Soria-Castro, E.; Carreón-Torres, E.; Díaz-Díaz, E.; Vázquez-Meza, H.; Guarner-Lans, V.; Rubio-Ruiz, M.E. Fenofibrate therapy restores antioxidant protection and improves myocardial insulin resistance in a rat model of metabolic syndrome and myocardial ischemia: The role of Angiotensin II. Molecules 2016, 22, 31. [Google Scholar] [CrossRef]
- Frank, A.; Bonney, M.; Bonney, S.; Weitzel, L.; Koeppen, M.; Eckle, T. Myocardial ischemia reperfusion injury: From basic science to clinical bedside. Semin. Cardiothorac. Vasc. Anesth. 2012, 16, 123–132. [Google Scholar] [CrossRef]
- Rahmutula, D.; Cui, J.; Chen, S.; Gardner, D.G. Transcriptional regulation of type B human natriuretic Peptide receptor gene promoter: Dependence on Sp1. Hypertension 2004, 44, 283–288. [Google Scholar] [CrossRef]
- Gardner, D.G.; Chen, S.; Glenn, D.J.; Grigsby, C.L. Molecular biology of the natriuretic peptide system: Implications for physiology and hypertension. Hypertension 2007, 49, 419–426. [Google Scholar] [CrossRef]
- Oidor-Chan, V.H.; Hong, E.; Pérez-Severiano, F.; Montes, S.; Torres-Narváez, J.C.; Del Valle-Mondragón, L.; Pastelín-Hernández, G.; Sánchez-Mendoza, A. Fenofibrate plus Metformin produces cardioprotection in a Type 2 diabetes and acute myocardial infarction model. PPAR Res. 2016, 2016, 8237264. [Google Scholar] [CrossRef]
- Sun, B.; Xie, Y.; Jiang, J.; Wang, Y.; Xu, X.; Zhao, C.; Huang, F. Pleiotropic effects of fenofibrate therapy on rats with hypertriglycemia. Lipids. Health Dis. 2015, 14, 27. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Bolli, R.; Canty, J.M., Jr.; Du, X.J.; Frangogiannis, N.G.; Frantz, S.; Gourdie, R.G.; Holmes, J.W.; Jones, S.P.; Kloner, R.A.; et al. Guidelines for experimental models of myocardial ischemia and infarction. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H812–H838. [Google Scholar] [CrossRef]
- Fernández-Sada, E.; Torres-Quintanilla, A.; Silva-Platas, C.; García, N.; Willis, B.C.; Rodríguez-Rodríguez, C.; De la Peña, E.; Bernal-Ramírez, J.; Treviño-Saldaña, N.; Oropeza-Almazán, Y.; et al. Proinflammatory cytokines are soluble mediators linked with ventricular arrhythmias and contractile dysfunction in a rat model of metabolic syndrome. Oxid. Med. Cell Longev. 2017, 2017, 7682569. [Google Scholar] [CrossRef]
- Rubio-Ruiz, M.E.; Guarner-Lans, V. Inflammation and the use of anti-inflammatory agents in signs and cardiovascular consequences of metabolic syndrome. In Handbook on Metabolic Syndrome. Classification, Risk Factors and Health Impact; Lopez-García, C., Pérez-González, P., Eds.; Nova Biomedical, Nova Science Publishers: New York, NY, USA, 2012; pp. 169–188. [Google Scholar]
- Jiang, N.; Li, Y.; Shu, T.; Wang, J. Cytokines and inflammation in adipogenesis: An updated review. Front. Med. 2019, 13, 314–329. [Google Scholar] [CrossRef]
- Brown, J.D.; Plutzky, J. Peroxisome proliferator-activated receptors as transcriptional nodal points and therapeutic targets. Circulation 2007, 115, 518–533. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef]
- Peterson, E.A.; Sun, J.; Wang, J. Leukocyte-mediated cardiac repair after myocardial infarction in Non-regenerative vs. regenerative systems. J. Cardiovasc. Dev. Dis. 2022, 9, 63. [Google Scholar] [CrossRef]
- Kruszewska, J.; Cudnoch-Jedrzejewska, A.; Czarzasta, K. Remodeling and fibrosis of the cardiac muscle in the course of obesity-pathogenesis and involvement of the extracellular matrix. Int. J. Mol. Sci. 2022, 23, 4195. [Google Scholar] [CrossRef]
- Nishida, K.; Otsu, K. Inflammation and metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 389–398. [Google Scholar] [CrossRef]
- Tuleta, I.; Frangogiannis, N.G. Fibrosis of the diabetic heart: Clinical significance, molecular mechanisms, and therapeutic opportunities. Adv. Drug Deliv. Rev. 2021, 176, 113904. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shan, X.; Chen, H.; Li, Z.; Zhao, P.; Zhang, C.; Guo, W.; Xu, M.; Lu, R. Stachydrine ameliorates cardiac fibrosis through inhibition of Angiotensin II/Transformation Growth Factor 1 fibrogenic axis. Front. Pharmacol. 2019, 10, 538. [Google Scholar] [CrossRef]
- Wilson, E.M.; Spinale, F.G. Myocardial remodelling and matrix metalloproteinases in heart failure: Turmoil within the interstitium. Ann. Med. 2001, 33, 623–634. [Google Scholar] [CrossRef]
- Cheung, P.Y.; Sawicki, G.; Wozniak, M.; Wang, W.; Radomski, M.W.; Schulz, R. Matrix metalloproteinase-2 contributes to ischemia-reperfusion injury in the heart. Circulation 2000, 101, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.R.; Nascimento, L.D.; Gerlach, R.F.; Rodrigues, K.E.; Prado, A.F. Matrix Metalloproteinase 2 as a pharmacological target in heart failure. Pharmaceuticals 2022, 15, 920. [Google Scholar] [CrossRef] [PubMed]
- Tsuruda, T.; Boerrigter, G.; Huntley, B.K.; Noser, J.A.; Cataliotti, A.; Costello-Boerrigter, L.C.; Chen, H.H.; Burnett, J.C., Jr. Brain natriuretic peptide is produced in cardiac fibroblasts and induces matrix metalloproteinases. Circ. Res. 2002, 91, 1127–1134. [Google Scholar] [CrossRef]
- Yue, T.L.; Bao, W.; Jucker, B.M.; Gu, J.L.; Romanic, A.M.; Brown, P.J.; Cui, J.; Thudium, D.T.; Boyce, R.; Burns-Kurtis, C.L.; et al. Activation of peroxisome proliferator-activated receptor-alpha protects the heart from ischemia/reperfusion injury. Circulation 2003, 108, 2393–2399. [Google Scholar] [CrossRef]
- Chen, J.; Tung, C.H.; Allport, J.R. Near-infrared fluorescent imaging of matrix metalloproteinase activity after myocardial infarction. Circulation 2005, 111, 1800–1805. [Google Scholar] [CrossRef]
- Jian, L.; Fan, Q.; Ding, T.; Sun, L.; Liu, S.; Niu, S. Correlation between matrix metalloproteinases and brain natriuretic peptide in patients with chronic heart failure. Chin. J. Geriatr. 2017, 12, 739–741. [Google Scholar]
- Fish-Trotter, H.; Ferguson, J.F.; Patel, N.; Arora, P.; Allen, N.B.; Bachmann, K.N.; Daniels, L.B.; Reilly, M.P.; Lima, J.A.C.; Wang, T.J.; et al. Inflammation and circulating natriuretic peptide levels. Circ. Heart Fail. 2020, 13, e006570. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, V.; Burnett, J.C., Jr. Natriuretic peptides, inflammation, and sounding the alarm. Circ. Heart Fail. 2020, 13, e007208. [Google Scholar] [CrossRef] [PubMed]
- Brigadeau, F.; Gelé, P.; Wibaux, M.; Marquié, C.; Martin-Nizard, F.; Torpier, G.; Fruchart, J.C.; Staels, B.; Duriez, P.; Lacroix, D. The PPARalpha activator fenofibrate slows down the progression of the left ventricular dysfunction in porcine tachycardia-induced cardiomyopathy. J. Cardiovasc. Pharmacol. 2007, 49, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Ping, P.; Wang, F.; Luo, L. Synthesis, secretion, function, metabolism and application of natriuretic peptides in heart failure. J. Biol. Eng. 2018, 12, 2. [Google Scholar] [CrossRef]
- Kuwahara, K. The natriuretic peptide system in heart failure: Diagnostic and therapeutic implications. Pharmacol. Ther. 2021, 227, 107863. [Google Scholar] [CrossRef]
- Pandey, K.N.; Nguyen, H.T.; Sharma, G.D.; Shi, S.J.; Kriegel, A.M. Ligand-regulated internalization, trafficking, and down-regulation of guanylyl cyclase/atrial natriuretic peptide receptor-A in human embryonic kidney 293 cells. J. Biol. Chem. 2002, 277, 4618–4627. [Google Scholar] [CrossRef]
- Calamera, G.; Moltzau, L.R.; Levy, F.O.; Andressen, K.W. Phosphodiesterases and compartmentation of cAMP and cGMP signaling in regulation of cardiac contractility in normal and failing hearts. Int. J. Mol. Sci. 2022, 23, 2145. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.; Yan, M.; Wu, Y.; Zheng, X. B-type natriuretic peptide-induced cardioprotection against reperfusion is associated with attenuation of mitochondrial permeability transition. Biol. Pharm. Bull. 2009, 32, 1545–1551. [Google Scholar] [CrossRef]
- Domondon, M.; Nikiforova, A.B.; DeLeon-Pennell, K.Y.; Ilatovskaya, D.V. Regulation of mitochondria function by natriuretic peptides. Am. J. Physiol. Renal. Physiol. 2019, 317, F1164–F1168. [Google Scholar] [CrossRef]
- Cao, Z.; Jia, Y.; Zhu, B. BNP and NT-proBNP as diagnostic biomarkers for cardiac dysfunction in both clinical and forensic medicine. Int. J. Mol. Sci. 2019, 20, 1820. [Google Scholar] [CrossRef]
- Hall, C. Essential biochemistry and physiology of (NT-pro)BNP. Eur. J. Heart Fail. 2004, 6, 257–260. [Google Scholar] [CrossRef]
- Sergeeva, I.A.; Christoffels, V.M. Regulation of expression of atrial and brain natriuretic peptide, biomarkers for heart development and disease. Biochim. Biophys. Acta. 2013, 1832, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.P.; Zhang, Y.; Mi, L.; Luo, X.Y.; Tian, M.H.; Zhu, B.L. The expression of B-Type natriuretic peptide after CaCl2-induced arrhythmias in rats. Am. J. Forensic. Med. Pathol. 2016, 37, 133–140. [Google Scholar] [CrossRef]
- Hasić, S.; Hadzović-Dzuvo, A.; Jadrić, R.; Kiseljakovic, E. B-type natriuretic peptide and adiponectin releases in rat model of myocardial damage induced by isoproterenol administration. Bosn. J. Basic. Med. Sci. 2013, 13, 225–229. [Google Scholar] [CrossRef]
- Potter, L.R.; Yoder, A.R.; Flora, D.R.; Antos, L.K.; Dickey, D.M. Natriuretic peptides: Their structures, receptors, physiologic functions and therapeutic applications. Handb. Exp. Pharmacol. 2009, 191, 341–366. [Google Scholar]
- Richards, A.M. New biomarkers in heart failure: Applications in diagnosis, prognosis and guidance of therapy. Rev. Esp. Cardiol. 2010, 63, 635–639. [Google Scholar] [CrossRef]
- Omran, F.; Kyrou, I.; Osman, F.; Lim, V.G.; Randeva, H.S.; Chatha, K. Cardiovascular biomarkers: Lessons of the past and prospects for the future. Int. J. Mol. Sci. 2022, 23, 5680. [Google Scholar] [CrossRef]
- Hayakawa, K.; Takemura, G.; Kanoh, M.; Li, Y.; Koda, M.; Kawase, Y.; Maruyama, R.; Okada, H.; Minatoguchi, S.; Fujiwara, T.; et al. Inhibition of granulation tissue cell apoptosis during the subacute stage of myocardial infarction improves cardiac remodeling and dysfunction at the chronic stage. Circulation 2003, 108, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Aguilar, M.; Ibarra-Lara, L.; Del Valle-Mondragón, L.; Soria-Castro, E.; Torres-Narváez, J.C.; Carreón-Torres, E.; Sánchez-Mendoza, A.; Rubio-Ruíz, M.E. Nonclassical axis of the renin-angiotensin system and neprilysin: Key mediators that underlie the cardioprotective effect of PPAR-Alpha activation during myocardial ischemia in a metabolic syndrome model. PPAR Res. 2020, 2020, 8894525. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Elami, A.; Gavish, Z.; Korach, A.; Houminer, E.; Schneider, A.; Schwalb, H.; Arav, A. Successful restoration of function of frozen and thawed isolated rat hearts. J. Thorac. Cardiovasc. Surg. 2008, 135, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Marques, L.S.; Fossati, A.A.N.; Rodrigues, R.B.; Da Rosa, H.T.; Izaguirry, A.P.; Ramalho, J.B.; Moreira, J.C.F.; Santos, F.W.; Zhang, T.; Streit, D.P., Jr. Slow freezing versus vitrification for the cryopreservation of zebrafish (Danio rerio) ovarian tissue. Sci. Rep. 2019, 9, 15353. [Google Scholar] [CrossRef] [PubMed]
- Rivas Leonel, E.C.; Lucci, C.M.; Amorim, C.A. Cryopreservation of human ovarian tissue: A review. Transfus. Med. Hemother. 2019, 46, 173–181. [Google Scholar] [CrossRef]
- Lu, M.; Qin, Q.; Yao, J.; Sun, L.; Qin, X. Induction of LOX by TGF-β1/Smad/AP-1 signaling aggravates rat myocardial fibrosis and heart failure. IUBMB Life 2019, 71, 1729–1739. [Google Scholar] [CrossRef]
- Wegner Kyle, A.; Keikhosravi, A.; Eliceiri, K.W.; Vezina, C.M. Fluorescence of picrosirius red multiplexed with immuno-histochemistry for the quantitative assessment of collagen in tissue sections. J Histochem Cytochem. 2017, 65(8), 479–490. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.L.; Barham, B.J.; Heidenreich, B.A. Comparison of indirect peroxidase and avidin-biotin-peroxidase complex (ABC) immunohistochemical staining procedures for c-fos in rat brain. J. Anat. 2019, 234, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Honvo-Houéto, E.; Truchet, S. Indirect immunofluorescence on frozen sections of mouse mammary gland. J. Vis. Exp. 2015, 106, e53179. [Google Scholar] [CrossRef]
- Im, K.; Mareninov, S.; Diazm, M.F.P.; Yong, W.H. An introduction to performing immunofluorescence staining. Methods Mol. Biol. 2019, 1897, 299–311. [Google Scholar] [CrossRef]
- Oberhaus, S.M. TUNEL and immunofluorescence double-labeling assay for apoptotic cells with specific antigen(s). Methods Mol. Biol. 2003, 218, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Kyrylkova, K.; Kyryachenko, S.; Biehs, B.; Klein, O.; Kioussi, C.; Leid, M. BCL11B regulates epithelial proliferation and asymmetric development of the mouse mandibular incisor. PLoS ONE 2012, 7, e37670. [Google Scholar] [CrossRef]
- Soria-Castro, E.; Soto, M.E.; Guarner-Lans, V.; Rojas, G.; Perezpeña-Diazconti, M.; Críales-Vera, S.A.; Manzano Pech, L.; Pérez-Torres, I. The kidnapping of mitochondrial function associated with the SARS-CoV-2 infection. Histol. Histopathol. 2021, 36, 947–965. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ct-V | Ct-Clo | MetS-V | MetS-Clo | |
|---|---|---|---|---|
| Body weight (g) | 536.5 ± 16.0 | 486.8 ± 16.9 | 559.6 ± 19.4 | 440.5 ± 16.9 b |
| Systolic blood pressure (mm Hg) | 98.9 ± 3.6 | 97.8 ± 2.4 | 143.6 ± 4.9 a | 139.7 ± 9.3 a |
| Intra-abdominal fat (g) | 5.33 ± 1.3 | 3.6 ± 0.4 | 16.6 ± 3.4 a | 7.7 ± 0.9 b |
| Triglycerides (mg/dL) | 71.2 ± 16.0 | 59.1 ± 7.8 | 150.3 ± 12.2 a | 84.5 ± 6.1 b |
| HDL-c (mg/dL) | 40.1 ± 5.3 | 38.6 ± 4.9 | 25.8 ± 2.8 a | 27.6 ± 3.2 |
| Non-HDL-c (mg/dL) | 19.8 ± 1.7 | 10.5 ± 1.2 b | 36.7 ± 3.6 a | 11.9 ± 3.2 b |
| Total cholesterol (mg/dL) | 53.0 ± 5.7 | 54.2 ± 5.2 | 51.7 ± 5.4 | 49.3 ± 6.9 |
| Glucose (mM) | 5.1 ± 0.22 | 4.9 ± 0.14 | 5.8 ± 0.08 | 5.3 ± 0.4 |
| Insulin (ng/mL) | 0.063 ± 0.003 | 0.076 ± 0.001 | 0.36 ± 0.09 a | 0.12 ± 0.02 b |
| HOMA-IR | 0.87 ± 0.2 | 0.67 ± 0.09 | 3.7 ± 0.5 a | 1.0 ± 0.8 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Aguilar, M.; Ibarra-Lara, L.; Cano-Martínez, A.; Soria-Castro, E.; Castrejón-Téllez, V.; Pavón, N.; Osorio-Yáñez, C.; Díaz-Díaz, E.; Rubio-Ruíz, M.E. PPAR Alpha Activation by Clofibrate Alleviates Ischemia/Reperfusion Injury in Metabolic Syndrome Rats by Decreasing Cardiac Inflammation and Remodeling and by Regulating the Atrial Natriuretic Peptide Compensatory Response. Int. J. Mol. Sci. 2023, 24, 5321. https://doi.org/10.3390/ijms24065321
Sánchez-Aguilar M, Ibarra-Lara L, Cano-Martínez A, Soria-Castro E, Castrejón-Téllez V, Pavón N, Osorio-Yáñez C, Díaz-Díaz E, Rubio-Ruíz ME. PPAR Alpha Activation by Clofibrate Alleviates Ischemia/Reperfusion Injury in Metabolic Syndrome Rats by Decreasing Cardiac Inflammation and Remodeling and by Regulating the Atrial Natriuretic Peptide Compensatory Response. International Journal of Molecular Sciences. 2023; 24(6):5321. https://doi.org/10.3390/ijms24065321
Chicago/Turabian StyleSánchez-Aguilar, María, Luz Ibarra-Lara, Agustina Cano-Martínez, Elizabeth Soria-Castro, Vicente Castrejón-Téllez, Natalia Pavón, Citlalli Osorio-Yáñez, Eulises Díaz-Díaz, and María Esther Rubio-Ruíz. 2023. "PPAR Alpha Activation by Clofibrate Alleviates Ischemia/Reperfusion Injury in Metabolic Syndrome Rats by Decreasing Cardiac Inflammation and Remodeling and by Regulating the Atrial Natriuretic Peptide Compensatory Response" International Journal of Molecular Sciences 24, no. 6: 5321. https://doi.org/10.3390/ijms24065321
APA StyleSánchez-Aguilar, M., Ibarra-Lara, L., Cano-Martínez, A., Soria-Castro, E., Castrejón-Téllez, V., Pavón, N., Osorio-Yáñez, C., Díaz-Díaz, E., & Rubio-Ruíz, M. E. (2023). PPAR Alpha Activation by Clofibrate Alleviates Ischemia/Reperfusion Injury in Metabolic Syndrome Rats by Decreasing Cardiac Inflammation and Remodeling and by Regulating the Atrial Natriuretic Peptide Compensatory Response. International Journal of Molecular Sciences, 24(6), 5321. https://doi.org/10.3390/ijms24065321