
Construction of ceRNA Networks at Different Stages of Somatic Embryogenesis in Garlic


 ,
, 
{kind=link}
{kind=link}
{kind=link}
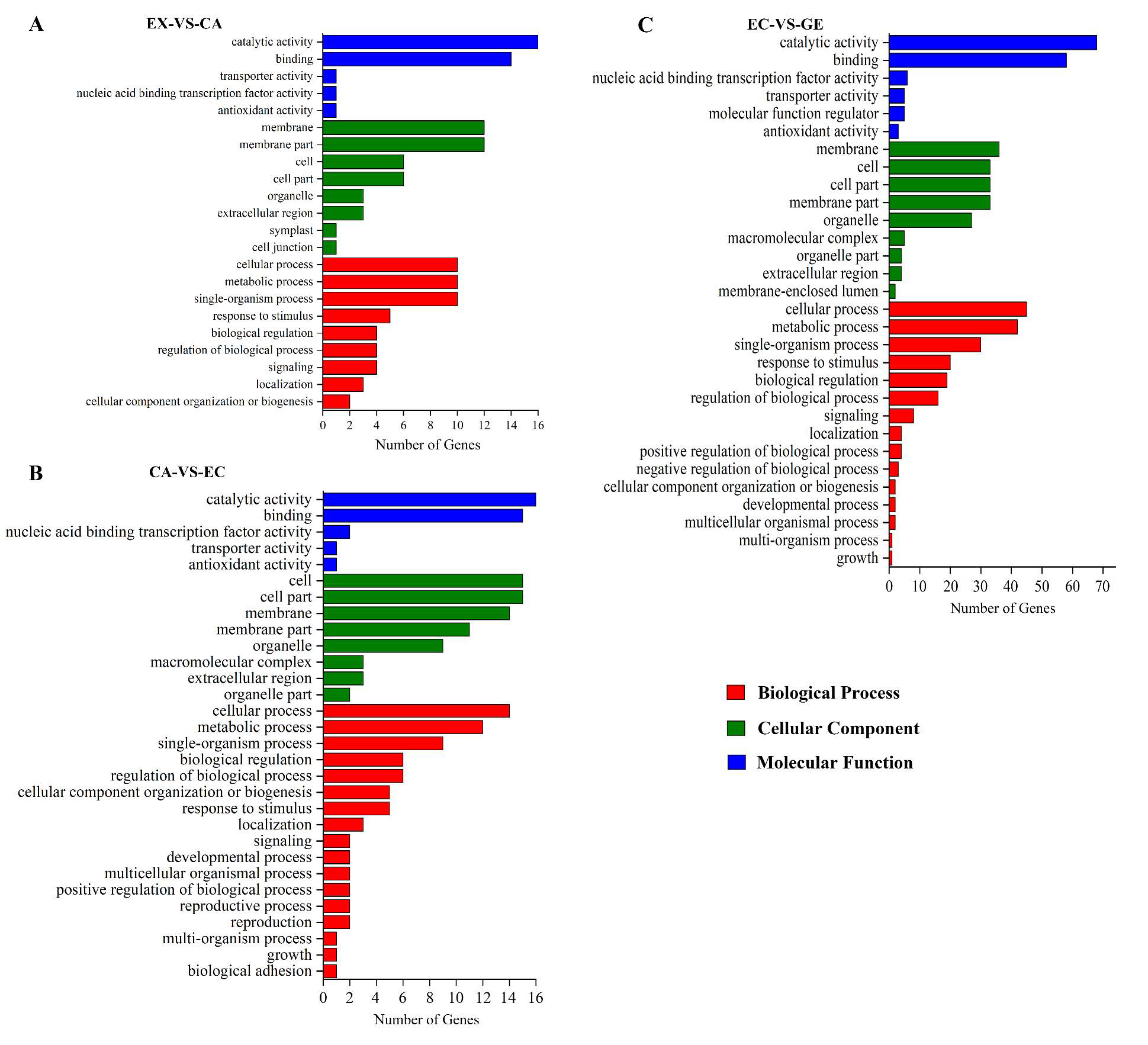{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Morphological and Histological Characteristics of Garlic Somatic Embryos at Different Developmental Stages
2.2. Identification and Functional Analysis of lncRNA and mRNA during Somatic Embryogenesis of Garlic
2.2.1. Identification and Characterization of lncRNA and mRNA during Somatic Embryogenesis of Garlic
2.2.2. Differential Expression Analysis of lncRNA and mRNA during Somatic Embryogenesis of Garlic
2.2.3. Function Prediction of Differentially Expressed lncRNA
2.3. Identification and Functional Analysis of miRNA during Somatic Embryogenesis of Garlic
2.4. lncRNA-miRNA Network Prediction
2.5. Construction of ceRNA Network in Adjacent Stages of Garlic Somatic Embryogenesis
2.6. Expression Identification of miR393s-ceRNA Network during Somatic Embryogenesis
3. Discussion
3.1. Expression Characteristics and Differences of lncRNA and miRNA in Different Stages of Garlic Somatic Embryogenesis
3.2. Correlation and Characteristics of miRNA with lncRNA and mRNA in Different Stages of Garlic Somatic Embryo Development
3.3. Regulatory Network and Potential Functions of ceRNA during Somatic Embryogenesis in Garlic
4. Materials and Methods
4.1. Plant Material and Culture
4.2. Total RNA Extraction
4.3. RNA Library Construction
4.4. Reads Mapping and Identification of lncRNA
4.5. Target Gene Prediction and Functional Enrichment Analysis
4.6. Prediction of the Relationship between lncRNA and miRNA
4.7. Expression Analysis by RT-qPCR
4.8. Data Statistics and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Namazi, H. The role of garlic in the prevention of ischemia-reperfusion injury: A new mechanism. Mol. Nutr. Food Res. 2010, 52, 739. [Google Scholar] [CrossRef] [PubMed]
- Botas, J.; Fernandes, N.; Barros, L.; Alves, M.J.; Carvalho, A.M.; Ferreira, I. A comparative study of black and white Allium sativum L.: Nutritional composition and bioactive properties. Molecules 2019, 24, 2194. [Google Scholar] [CrossRef] [PubMed]
- Einat, S.M.; Tomer, B.M.; Neta, R.; Rabinowitch, H.D.; Adi, D.F.; Arkadiusz, K.; Dawid, P.; Amir, S.; Rina, K. Garlic (Allium sativum L.) fertility: Transcriptome and proteome analyses provide insight into flower and pollen development. Front. Plant. Sci. 2015, 6, 271. [Google Scholar]
- Fan, B.L.; He, R.F.; Shang, Y.T.; Xu, L.W.; Wang, N.N.; Gao, H.; Liu, X.Y.; Wang, Z.Y. System construction of virus-free and rapid-propagation technology of baodi garlic (Allium sativum L.). Sci. Hortic. 2017, 225, 498–504. [Google Scholar] [CrossRef]
- Albrecht, C.; Russinova, E.; Hecht, V.; Baaijiens, E.; Vires, S. The Arabidopsis thaliana somatic embryogenesis receptor-like kinases 1 and 2 control male sporogenesis. Plant. Cell 2006, 17, 3337–3349. [Google Scholar] [CrossRef]
- Zimmerman, J.L. Somatic embryogenesis: A model for early development in higher plants. Plant. Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef]
- Zheng, Q.L.; Zheng, Y.M.; Ji, H.H.; Burnie, W.; Perry, S.E. Gene regulation by the AGL15 transcription factor reveals hormone interactions in somatic embryogenesis. Plant. Physiol. 2016, 172, 2374–2387. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Bai, B.; Zhang, X.S. Establishment of embryonic shoot–root axis is involved in auxin and cytokinin response during Arabidopsis somatic embryogenesis. Front. Plant. Sci. 2015, 5, 792. [Google Scholar] [CrossRef]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; Vries, S. The Arabidopsis somatic embryogenesis receptor-like kinase1 protein complex includes brassinosteroid-insensitive1. Plant. Cell 2006, 18, 626–638. [Google Scholar] [CrossRef]
- Muhammad, S.H.; Wada, T.; Hattori, k. Efficient plant regeneration in garlic through somatic embryogenesis from root tip explants. Plant. Prod. Sci. 1998, 1, 216–222. [Google Scholar]
- Fereol, L.; Chovelon, V.; Causse, S.; Michaux-Ferriere, N.; Kahane, R. Evidence of a somatic embryogenesis process for plant regeneration in garlic (Allium sativum L.). Plant. Cell Rep. 2002, 21, 197–203. [Google Scholar]
- Wójcikowska, B.; Gaj, M.D. Expression profiling of auxin response factor genes during somatic embryogenesis induction in Arabidopsis. Plant. Cell Rep. 2017, 36, 843–858. [Google Scholar] [CrossRef]
- Luo, Y.C.; Zhou, H.; Li, Y.; Chen, J.Y.; Yang, J.H.; Chen, Y.Q.; Qu, L.H. Rice embryogenic calli express a unique set of microRNAs, suggesting regulatory roles of microRNAs in plant post-embryogenic development. FEBS Lett. 2006, 580, 5111–5116. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Kou, S.J.; Liu, Y.L.; Fang, Y.N.; Xu, Q.; Guo, W.W. Genomewide analysis of small RNAs in nonembryogenic and embryogenic tissues of citrus: microRNA-and siRNA-mediated transcript cleavage involved in somatic embryogenesis. Plant. Biotechnol. J. 2015, 13, 383–394. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lai, Z.X.; Tian, Q.L.; Lin, L.X.; Lai, R.L.; Yang, M.M.; Zhang, D.M.; Chen, Y.K.; Zhang, Z.H. Endogenous target mimics down-regulate miR160 mediation of ARF10, -16, and -17 cleavage during somatic embryogenesis in dimocarpus longan lour. Front. Plant. Sci. 2015, 6, 956. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Wang, L.C.; Yuan, D.J.; Lindsey, K.; Zhang, X.L. Small RNA and degradome sequencing reveal complex miRNA regulation during cotton somatic embryogenesis. J. Exp. Bot. 2013, 64, 1521–1536. [Google Scholar] [CrossRef]
- Li, W.F.; Zhang, S.G.; Han, S.Y.; Wu, S.Y.; Zhang, J.H.; Qi, L.W. The post-transcriptional regulation of laSCL6 by miR171 during maintenance of embryogenic potential in Larix kaempferi (lamb.) carr. Tree Genet. Genomes 2014, 10, 223–229. [Google Scholar] [CrossRef]
- Zhu, B.Z.; Yang, Y.F.; Li, R.; Fu, D.Q.; Wen, L.W.; Luo, Y.B.; Zhu, H.L. RNA sequencing and functional analysis implicate the regulatory role of long non-coding RNAs in tomato fruit ripening. J. Exp. Bot. 2015, 66, 4483–4495. [Google Scholar] [CrossRef]
- Yang, T.; Ma, H.Y.; Zhang, J.; Wu, T.; Song, T.T.; Tian, J.; Yao, Y.C. Systematic identification of long noncoding RNAs expressed during light-induced anthocyanin accumulation in apple fruit. Plant. J. 2019, 100, 572–590. [Google Scholar] [CrossRef]
- Liu, S.; Wu, L.; Qi, H.R.; Xu, M. LncRNA/circRNA–miRNA–mRNA networks regulate the development of root and shoot meristems of Populus. Ind. Crops Prod. 2019, 133, 333–347. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.H.; Liang, Q.; Zhang, Y.H.; Kang, K.Q.; Wang, W.T.; Feng, Y.; Wu, S.H.; Yang, C.; Li, Y.Y. Genome-wide analysis of long noncoding RNAs affecting floral bud dormancy in pears in response to cold stress. Tree Physiol. 2020, 41, 771–790. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.Y.; Liu, Z.C. Global identification and analysis of long non-coding RNAs in diploid strawberry Fragaria Vesca during flower and fruit development. BMC Genom. 2015, 16, 815. [Google Scholar] [CrossRef] [PubMed]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fu, D.Q.; Zhu, B.Z.; Luo, Y.B.; Zhu, H.L. CRISPR/Cas9-mediated mutagenesis of lncrna1459 alters tomato fruit ripening. Plant. J. 2018, 97, 795. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.B.; Yang, X.S.; Li, X.L.; Wang, J.L.; Gao, Y.; Shang, R.Z.; Sun, W.; Dou, K.F.; Li, H.M. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; Garcia, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef]
- Pan, T.; Sun, X.Q.; Liu, Y.X.; Li, H.; Deng, G.B.; Lin, H.H.; Wang, S.H. Heat stress alters genome-wide profiles of circular RNAs in Arabidopsis. Plant. Mol. Biol. 2018, 96, 217–229. [Google Scholar] [CrossRef]
- Deng, F.N.; Zhang, X.P.; Wang, W.; Yuan, R.; Shen, F.F. Identification of Gossypium hirsutum long non-coding RNAs (lncRNAs) under salt stress. BMC Plant. Biol. 2018, 18, 23. [Google Scholar] [CrossRef]
- Li, H.; Heng, D.; Dong, Z.; Ming, L.; Liu, Y.H.; Zhang, F.; Feng, Y.Y.; Yu, D.L.; Liu, S.; Cao, J.S. Systematic identification of long non-coding RNAs during pollen development and fertilization in Brassica rapa. Plant. J. 2018, 96, 203–222. [Google Scholar]
- Williams, L.; Grigg, S.P.; Xie, M.T.; Christensen, S.; Fletcher, J.C. Regulation of Arabidopsis shoot apical meristem and lateral organ formation by microRNA miR166g and its AtHD-ZIP target genes. Development 2005, 132, 3657–3668. [Google Scholar] [CrossRef] [PubMed]
- Ákos, Á.; Zoltán, H. Analysis of gradient-like expression of mir167 in Arabidopsis thaliana embryonic tissue. J. Plant. Biol. 2013, 56, 336–344. [Google Scholar]
- Vernoux, T.; Benfey, P.N. Signals that regulate stem cell activity during plant development. Curr. Opin. Genet. Dev. 2005, 15, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Ohtani, M.; Mitsuda, N.; Kubo, M.; Ohme-Takagi, M.; Fukuda, H.; Demura, T. VND-interacting2, a NAC domain transcription factor, negatively regulates xylem vessel formation in Arabidopsis. Plant. Cell 2010, 22, 1249–1263. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhang, S.G.; Han, S.Y.; Wu, T.; Li, X.M.; Li, W.F.; Qi, L.W. Genome-wide identification of microRNAs in Larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 2012, 236, 647–657. [Google Scholar] [CrossRef]
- Kuang, B.; Zhao, X.; Zhou, C.; Zeng, W.; Ren, J.; Ebert, B.; Beahan, C.T.; Deng, X.; Zeng, Q.; Zhou, G.; et al. Role of UDP-glucuronic acid decarboxylase in Xylan biosynthesis in Arabidopsis. Mol. Plant. 2016, 9, 1119–1131. [Google Scholar] [CrossRef]
- Ruan, N.; Dang, Z.; Wang, M.; Cao, L.; Wang, Y.; Liu, S.; Tang, Y.; Huang, Y.; Zhang, Q.; Xu, Q.; et al. Fragile culm 18 encodes a UDP-glucuronic acid decarboxylase required for xylan biosynthesis and plant growth in rice. J. Exp. Bot. 2022, 73, 2320–2335. [Google Scholar] [CrossRef]
- Juárez-González, V.T.; López-Ruiz, B.A.; Baldrich, P.; Eduardo, L.S.; Meyers, B.C.; Dinkova, T.D. The explant developmental stage profoundly impacts small RNA-mediated regulation at the dedifferentiation step of maize somatic embryogenesis. Sci. Rep. 2019, 9, 14511. [Google Scholar] [CrossRef]
- Wierzbicki, A.T. The role of long non-coding RNA in transcriptional gene silencing. Curr. Opin. in Plant. Biol. 2012, 15, 517–522. [Google Scholar] [CrossRef]
- Chen, R.Z.; Shen, X.; Zhang, S.T.; Zhao, H.; Chen, X.H.; Xu, X.P.; Huo, W.; Zhang, Z.H.; Lin, Y.L.; Lai, Z.X. Genome-wide identification and expression analysis of Argonaute gene family from longan embryogenic callus. J. Integr. Agric. 2021, 20, 2138–2155. [Google Scholar] [CrossRef]
- Wójcikowska, B.; Wójcik, A.M.; Gaj, M.D. Epigenetic regulation of auxin-induced somatic embryogenesis in plants. Int. J. Mol. Sci. 2020, 21, 2307. [Google Scholar] [CrossRef]
- Lemaitre, R.N.; King, I.B.; Sotoodehnia, N. Red blood cell membrane α-linolenic acid and the risk of sudden cardiac arrest. Metab. Clin. Exp. 2009, 58, 534–540. [Google Scholar] [CrossRef]
- Mueller, M.J. Enzymes involved in jasmonic acid biosynthesis. Physiol. Plant. 1997, 100, 653–663. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Shi, Y.S.; Yang, G.L.; Zhou, X.X.; Hou, X.X.; Meng, J.; Luan, Y.S. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate NBS-LRR genes by decoying miR482b in the tomato-Phytophthora infestans interaction. Hort. Res. 2019, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Shen, J.; Xu, Q.J.; Dong, J.; Song, L.R.; Wang, W.; Shen, F.F. Long noncoding RNA lncrna354 functions as a competing endogenous RNA of mir160b to regulate ARF genes in response to salt stress in upland cotton. Plant. Cell Environ. 2021, 44, 3302–3321. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, T.; Hamada, M. RIblast: An ultrafast RNA-RNA interaction prediction system based on a seed-and-extension approach. Bioinformatics 2017, 33, 2666–2674. [Google Scholar] [CrossRef]
- Dai, X.B.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Liu, M.; Zhou, R.; Jiang, F.; Li, P.; Li, M.; Zhang, M.; Wei, H.; Wu, Z. Construction of ceRNA Networks at Different Stages of Somatic Embryogenesis in Garlic. Int. J. Mol. Sci. 2023, 24, 5311. https://doi.org/10.3390/ijms24065311
Bai Y, Liu M, Zhou R, Jiang F, Li P, Li M, Zhang M, Wei H, Wu Z. Construction of ceRNA Networks at Different Stages of Somatic Embryogenesis in Garlic. International Journal of Molecular Sciences. 2023; 24(6):5311. https://doi.org/10.3390/ijms24065311
Chicago/Turabian StyleBai, Yunhe, Min Liu, Rong Zhou, Fangling Jiang, Ping Li, Mengqian Li, Meng Zhang, Hanyu Wei, and Zhen Wu. 2023. "Construction of ceRNA Networks at Different Stages of Somatic Embryogenesis in Garlic" International Journal of Molecular Sciences 24, no. 6: 5311. https://doi.org/10.3390/ijms24065311
APA StyleBai, Y., Liu, M., Zhou, R., Jiang, F., Li, P., Li, M., Zhang, M., Wei, H., & Wu, Z. (2023). Construction of ceRNA Networks at Different Stages of Somatic Embryogenesis in Garlic. International Journal of Molecular Sciences, 24(6), 5311. https://doi.org/10.3390/ijms24065311