
Structurally Different Yet Functionally Similar: Aptamers Specific for the Ebola Virus Soluble Glycoprotein and GP1,2 and Their Application in Electrochemical Sensing


 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
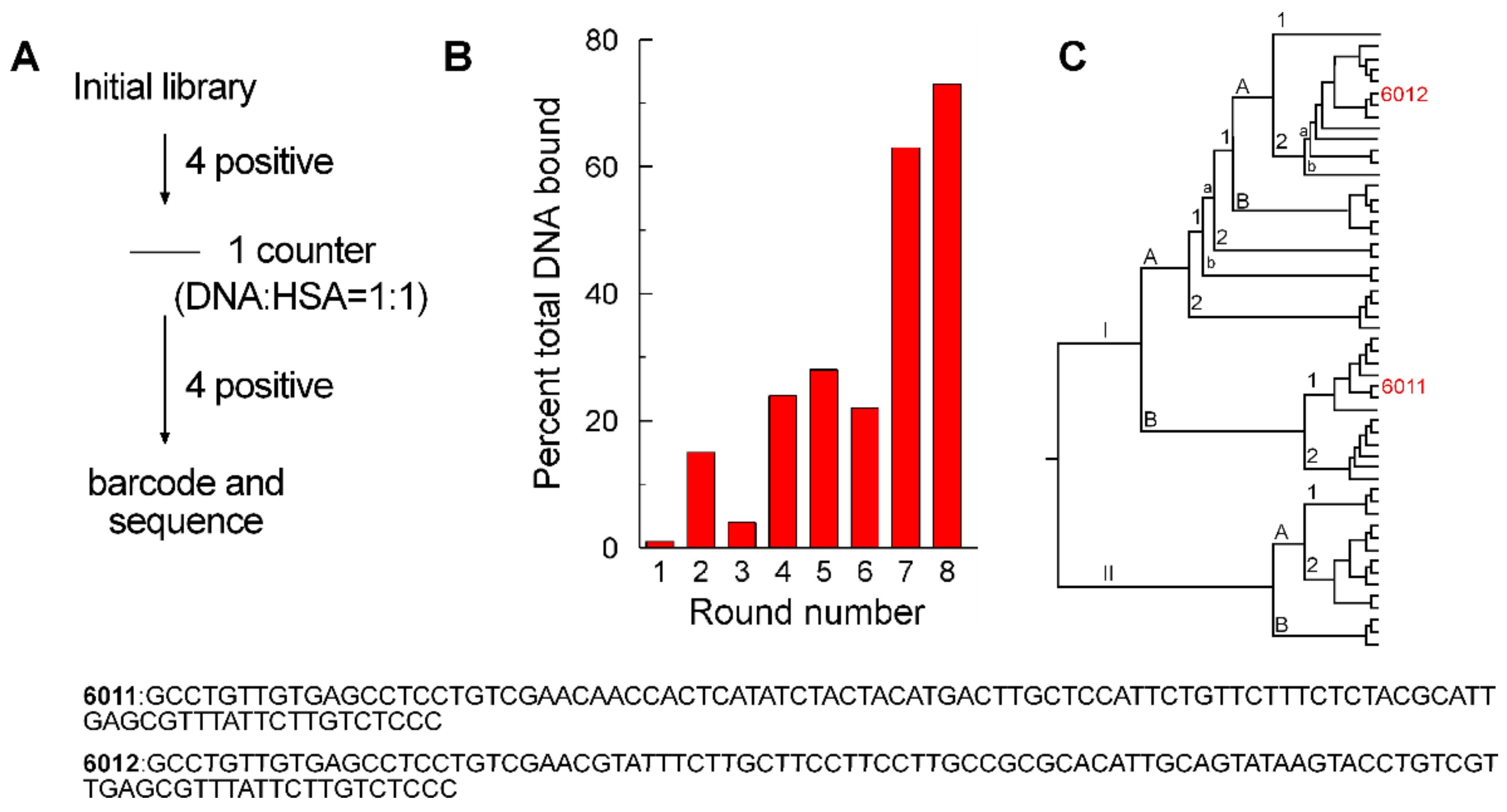
2.1. Aptamer Selection
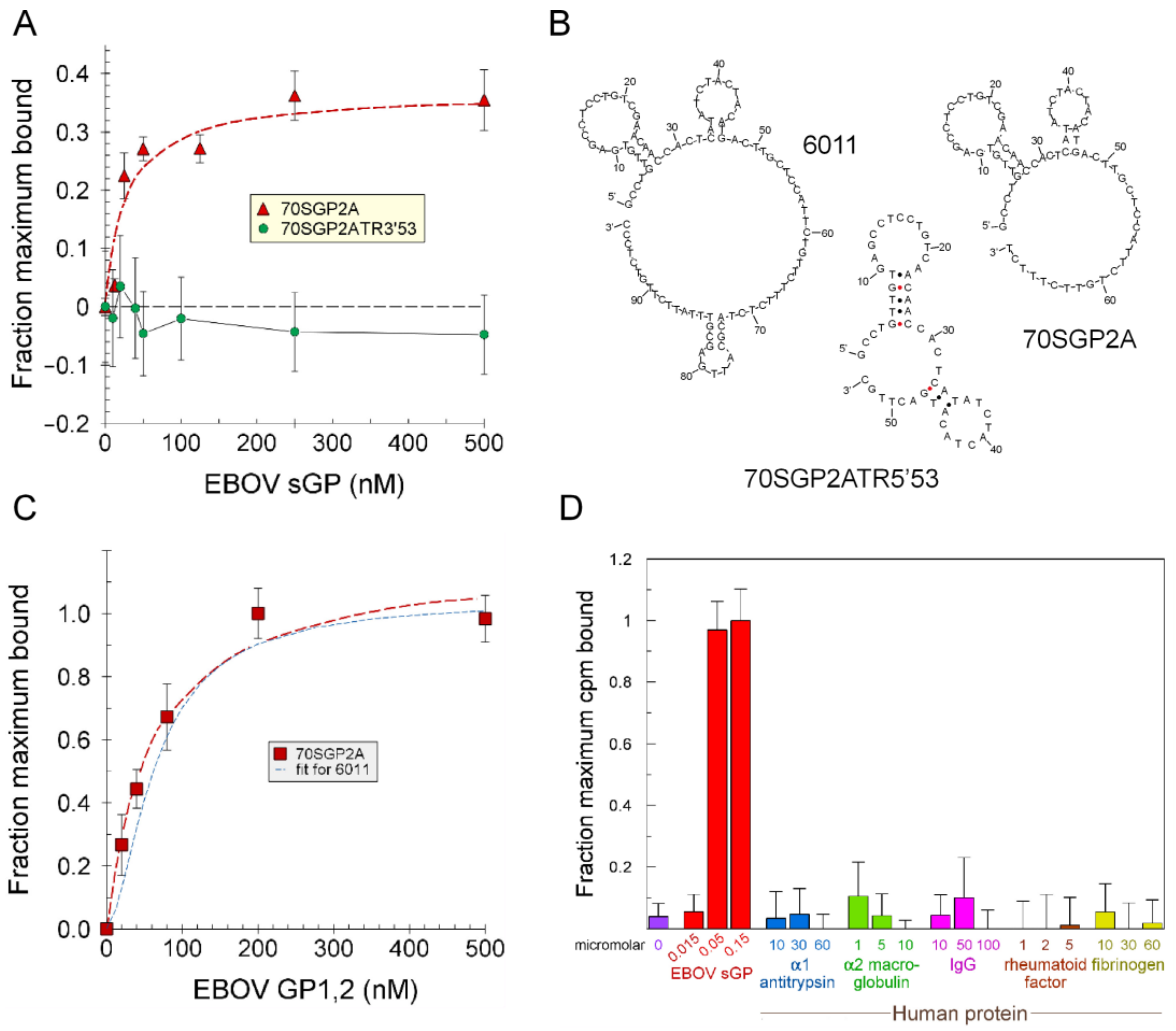
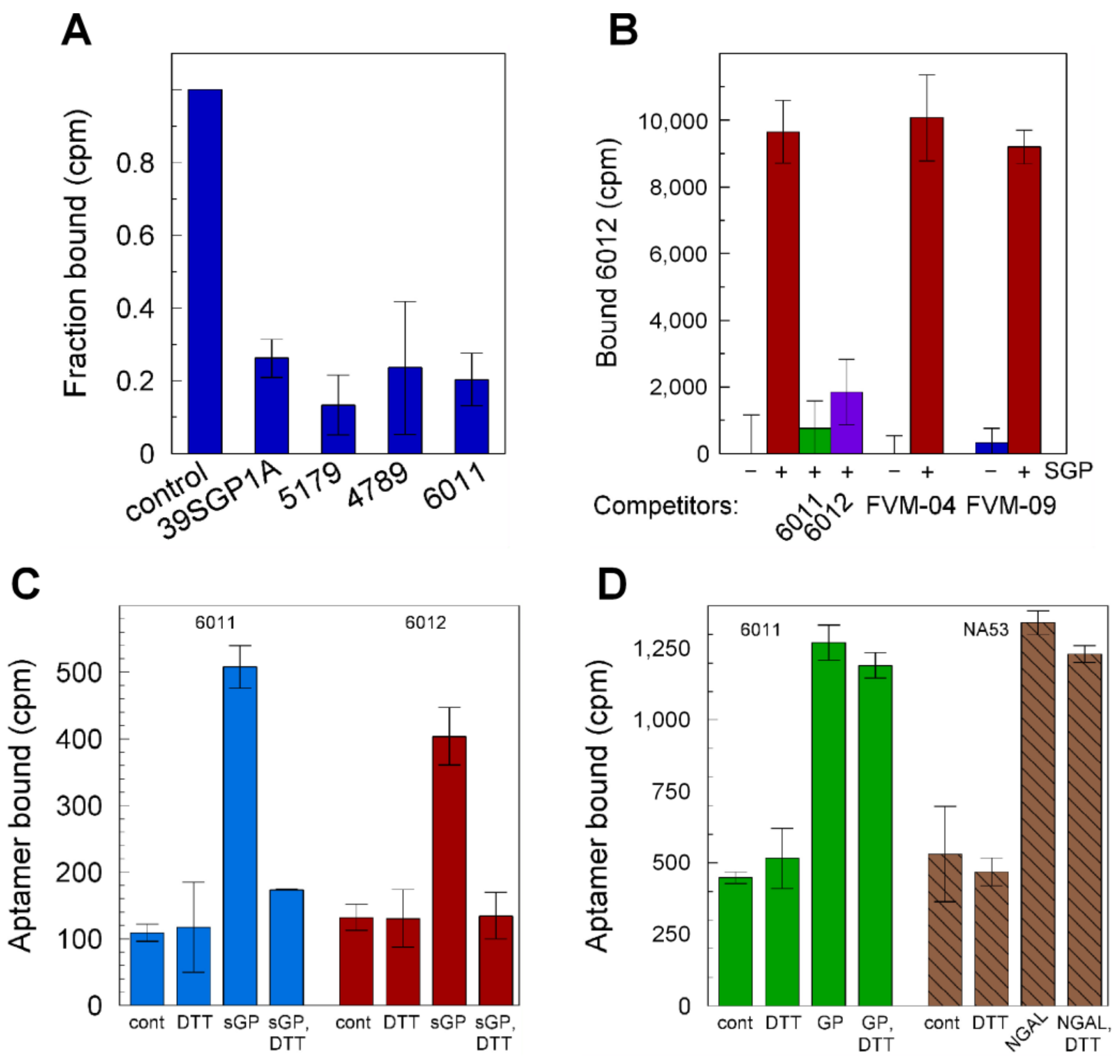
2.2. Aptamer Affinities and Specificities
2.3. Truncation to Produce Aptamer 70SGP2A
2.4. Protein Surface Epitope for Aptamer Binding
2.5. Molecular Modeling to Explore the Probable Aptamer Docking Site on sGP
2.6. Aptamer Structural Stabilities in the Presence of Serum
2.7. Electrochemical Assay and Detection of sGP and GP1,2
3. Discussion
3.1. Aptamer Characteristics
3.2. Application of Aptamers as Recognition Elements on an Electrochemical Sensor
3.3. Protein Site(s) to Which the Aptamers Bind
3.4. Distinction of Antibody and Aptamer Binding Sites on sGP
3.5. Summary
4. Materials and Methods
4.1. Buffers
4.2. Ebola Virus Proteins
4.3. Aptamers and Oligonucleotides
4.4. Ebola Pseudovirions
4.5. In Vitro Selection of DNA Aptamers against sGP (Soluble Glycoprotein)
4.6. End Labelling the ssDNA Oligonucleotides and In Vitro Translation of 2′FY-RNA for Characterizing Aptamers
4.7. Single Well and Multiplex Filter Capture Assays
4.8. Gel Electrophoresis, Western Blots, and ELISA
- PAGE
- Western blots
- ELISA
- Aptamer competition tested by ELISA
4.9. Aptamer and Protein Modeling and Docking
4.10. Preparing the NAAO Membranes for Sensing Experiments
4.11. Electrochemical Sensing (EIS)
4.12. EIS of Infected Monkey Serum
4.13. Analysis of Binding Isotherms and Statistical Evaluations
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verikios, G. The dynamic effects of infectious disease outbreaks: The case of pandemic influenza and human coronavirus. Socio-Econ. Plan. Sci. 2020, 71, 100898. [Google Scholar] [CrossRef] [PubMed]
- Omoleke, S.A.; Mohammed, I.; Saidu, Y. Ebola Viral Disease in West Africa: A Threat to Global Health, Economy and Political Stability. J. Public Health Afr. 2016, 7, 534. [Google Scholar] [CrossRef] [PubMed]
- Kadanali, A.; Karagoz, G. An overview of Ebola virus disease. North. Clin. Istanb. 2015, 2, 81–86. [Google Scholar] [CrossRef]
- WHO. Ebola Virus Disease. Available online: https://www.who.int/en/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 7 May 2021).
- Keita, A.K.; Koundouno, F.R.; Faye, M.; Düx, A.; Hinzmann, J.; Diallo, H.; Ayouba, A.; Le Marcis, F.; Soropogui, B.; Ifono, K.; et al. Resurgence of Ebola virus in 2021 in Guinea suggests a new paradigm for outbreaks. Nature 2021, 597, 539–543. [Google Scholar] [CrossRef]
- Mehedi, M.; Falzarano, D.; Seebach, J.; Hu, X.; Carpenter, M.S.; Schnittler, H.-J.; Feldmann, H. A new Ebola virus nonstructural glycoprotein expressed through RNA editing. J. Virol. 2011, 85, 5406–5414. [Google Scholar] [CrossRef]
- De La Vega, M.-A.; Wong, G.; Kobinger, G.P.; Qiu, X. The Multiple Roles of sGP in Ebola Pathogenesis. Viral Immunol. 2015, 28, 3–9. [Google Scholar] [CrossRef]
- Sanchez, A.; Trappier, S.G.; Mahy, B.W.; Peters, C.J.; Nichol, S.T. The virion glycoproteins of Ebola viruses are encoded in two reading frames and are expressed through transcriptional editing. Proc. Natl. Acad. Sci. USA 1996, 93, 3602–3607. [Google Scholar] [CrossRef]
- Sanchez, A.; Ksiazek, T.G.; Rollin, P.E.; Miranda, M.E.; Trappier, S.G.; Khan, A.S.; Peters, C.J.; Nichol, S.T. Detection and molecular characterization of Ebola viruses causing disease in human and nonhuman primates. J. Infect. Dis. 1999, 179, S164–S169. [Google Scholar] [CrossRef]
- Alfson, K.J.; Goez-Gazi, Y.; Gazi, M.; Staples, H.; Mattix, M.; Ticer, A.; Klaffke, B.; Stanfield, K.; Escareno, P.; Keiser, P.; et al. Development of a Well-Characterized Rhesus Macaque Model of Ebola Virus Disease for Support of Product Development. Microorganisms 2021, 9, 489. [Google Scholar] [CrossRef]
- Shubham, S.; Hoinka, J.; Banerjee, S.; Swanson, E.; Dillard, J.A.; Lennemann, N.J.; Przytycka, T.M.; Maury, W.; Nilsen-Hamilton, M. A 2′FY-RNA Motif Defines an Aptamer for Ebolavirus Secreted Protein. Sci. Rep. 2018, 8, 12373. [Google Scholar] [CrossRef]
- Hong, S.-L.; Xiang, M.-Q.; Tang, M.; Pang, D.-W.; Zhang, Z.-L. Ebola Virus Aptamers: From Highly Efficient Selection to Application on Magnetism-controlled Chips. Anal. Chem. 2019, 91, 3367–3373. [Google Scholar] [CrossRef]
- Kortepeter, M.G.; Bausch, D.G.; Bray, M. Basic clinical and laboratory features of filoviral hemorrhagic fever. J. Infect. Dis. 2011, 204, S810–S816. [Google Scholar] [CrossRef]
- Broadhurst, M.J.; Brooks, T.J.; Pollock, N.R. Diagnosis of Ebola Virus Disease: Past, Present, and Future. Clin. Microbiol. Rev. 2016, 29, 773–793. [Google Scholar] [CrossRef]
- Furuyama, W.; Marzi, A. Development of an Enzyme-Linked Immunosorbent Assay to Determine the Expression Dynamics of Ebola Virus Soluble Glycoprotein during Infection. Microorganisms 2020, 8, 1535. [Google Scholar] [CrossRef] [PubMed]
- Grieshaber, D.; MacKenzie, R.; Vörös, J.; Reimhult, E. Electrochemical Biosensors—Sensor Principles and Architectures. Sensors 2008, 8, 1400–1458. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Wambach, T.; Venkatesh, A.G.; Hall, D.A. A Low-Cost Smartphone-Based Electrochemical Biosensor for Point-of-Care Diagnostics. In Proceedings of the 2014 IEEE Biomedical Circuits and Systems Conference (BioCAS), Lausanne, Switzerland, 22–24 October 2014; pp. 312–315. [Google Scholar]
- Pallesen, J.; Murin, C.D.; de Val, N.; Cottrell, C.A.; Hastie, K.M.; Turner, H.L.; Fusco, M.L.; Flyak, A.I.; Zeitlin, L.; Crowe, J.E.; et al. Structures of Ebola virus GP and sGP in complex with therapeutic antibodies. Nat. Microbiol. 2016, 1, 16128. [Google Scholar] [CrossRef] [PubMed]
- Shedlock, D.J.; Bailey, M.A.; Popernack, P.M.; Cunningham, J.M.; Burton, D.R.; Sullivan, N.J. Antibody-mediated neutralization of Ebola virus can occur by two distinct mechanisms. Virology 2010, 401, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.A.; Nilsen-Hamilton, M. A mathematical analysis of SELEX. Comput. Biol. Chem. 2007, 31, 11–35. [Google Scholar] [CrossRef]
- Keck, Z.-Y.; Enterlein, S.G.; Howell, K.A.; Vu, H.; Shulenin, S.; Warfield, K.L.; Froude, J.W.; Araghi, N.; Douglas, R.; Biggins, J.; et al. Macaque Monoclonal Antibodies Targeting Novel Conserved Epitopes within Filovirus Glycoprotein. J. Virol. 2016, 90, 279–291. [Google Scholar] [CrossRef]
- Zhang, Q.; Gui, M.; Niu, X.; He, S.; Wang, R.; Feng, Y.; Kroeker, A.; Zuo, Y.; Wang, H.; Wang, Y.; et al. Potent neutralizing monoclonal antibodies against Ebola virus infection. Sci. Rep. 2016, 6, 25856. [Google Scholar] [CrossRef]
- Holtsberg, F.W.; Shulenin, S.; Vu, H.; Howell, K.A.; Patel, S.J.; Gunn, B.; Karim, M.; Lai, J.R.; Frei, J.C.; Nyakatura, E.K.; et al. Pan-ebolavirus and Pan-filovirus Mouse Monoclonal Antibodies: Protection against Ebola and Sudan Viruses. J. Virol. 2016, 90, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Yan, H.; Xie, F.; Wang, K.; Wang, Q.; Huang, H.; Yang, K.; Huang, S.; Zhao, T.; Wang, J.; et al. Development of a novel ssDNA aptamer targeting neutrophil gelatinase-associated lipocalin and its application in clinical trials. J. Transl. Med. 2019, 17, 204. [Google Scholar] [CrossRef] [PubMed]
- Gosai, A.; Yeah, B.S.H.; Nilsen-Hamilton, M.; Shrotriya, P. Label Free Thrombin Detection in Presence of High Concentration of Albumin Using an Aptamer-Functionalized Nanoporous Membrane. Biosens. Bioelectron. 2019, 126, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Perez, B.; Volchkova, V.A.; Dolnik, O.; Lawrence, P.; Volchkov, V.E. Shed GP of Ebola virus triggers immune activation and increased vascular permeability. PLoS Pathog. 2014, 10, e1004509. [Google Scholar] [CrossRef] [PubMed]
- Judson, S.D.; Fischer, R.; Judson, A.; Munster, V.J. Ecological Contexts of Index Cases and Spillover Events of Different Ebolaviruses. PLoS Pathog. 2016, 12, e1005780. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, A.; Shimojima, M.; Kawaoka, Y. sGP Serves as a Structural Protein in Ebola Virus Infection. J. Infect. Dis. 2011, 204, S897–S903. [Google Scholar] [CrossRef]
- Layzer, J.M.; McCaffrey, A.P.; Tanner, A.K.; Huang, Z.; Kay, M.A.; Sullenger, B.A. In vivo activity of nuclease-resistant siRNAs. RNA 2004, 10, 766–771. [Google Scholar] [CrossRef]
- Allerson, C.R.; Sioufi, N.; Jarres, R.; Prakash, T.P.; Naik, N.; Berdeja, A.; Wanders, L.; Griffey, R.H.; Swayze, E.E.; Bhat, B. Fully 2′-modified oligonucleotide duplexes with improved in vitro potency and stability compared to unmodified small interfering RNA. J. Med. Chem. 2005, 48, 901–904. [Google Scholar] [CrossRef]
- Patra, A.; Paolillo, M.; Charisse, K.; Manoharan, M.; Rozners, E.; Egli, M. 2’-Fluoro RNA shows increased Watson-Crick H-bonding strength and stacking relative to RNA: Evidence from NMR and thermodynamic data. Angew. Chem. Int. Ed. 2012, 51, 11863–11866. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Q.; Hu, Z.; Zhang, Y.; Wu, C.; Yang, M.; Wang, P. A novel electrochemical biosensor based on dynamic polymerase-extending hybridization for E. coli O157:H7 DNA detection. Talanta 2009, 78, 647–652. [Google Scholar] [CrossRef]
- Tan, F.; Leung, P.H.M.; Liu, Z.-b.; Zhang, Y.; Xiao, L.; Ye, W.; Zhang, X.; Yi, L.; Yang, M. A PDMS microfluidic impedance immunosensor for E. coli O157:H7 and Staphylococcus aureus detection via antibody-immobilized nanoporous membrane. Sens. Actuators B Chem. 2011, 159, 328–335. [Google Scholar] [CrossRef]
- Ye, W.W.; Shi, J.Y.; Chan, C.Y.; Zhang, Y.; Yang, M. A nanoporous membrane based impedance sensing platform for DNA sensing with gold nanoparticle amplification. Sens. Actuators B Chem. 2014, 193, 877–882. [Google Scholar] [CrossRef]
- Devarakonda, S.; Ganapathysubramanian, B.; Shrotriya, P. Impedance-Based Nanoporous Anodized Alumina/ITO Platforms for Label-Free Biosensors. ACS Appl. Mater. Interfaces 2022, 14, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Wang, T.; Kang, K.; Zhao, Y.; Shrotriya, P.; Nilsen-Hamilton, M. An RNA aptamer-based microcantilever sensor to detect the inflammatory marker, mouse lipocalin-2. Anal. Chem. 2012, 84, 8763–8770. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, N.F.; Holmstrom, E.D.; Nesbitt, D.J. Molecular-crowding effects on single-molecule RNA folding/unfolding thermodynamics and kinetics. Proc. Natl. Acad. Sci. USA 2014, 111, 8464–8469. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Miyoshi, D.; Sugimoto, N. Effects of molecular crowding on the structures, interactions, and functions of nucleic acids. Chem. Rev. 2014, 114, 2733–2758. [Google Scholar] [CrossRef]
- Han, J.H.; Kim, J.H.; Kim, S.K.; Jang, Y.J. Conformational change of a G-quadruplex under molecular crowding conditions. J. Biomol. Struct. Dyn. 2020, 38, 2575–2581. [Google Scholar] [CrossRef]
- Howell, K.A.; Qiu, X.; Brannan, J.M.; Bryan, C.; Davidson, E.; Holtsberg, F.W.; Wec, A.Z.; Shulenin, S.; Biggins, J.E.; Douglas, R.; et al. Antibody treatment of Ebola and Sudan virus infection via a uniquely exposed epitope within the glycoprotein receptor-binding site. Cell Rep. 2016, 15, 1514–1526. [Google Scholar] [CrossRef]
- Howell, K.A.; Brannan, J.M.; Bryan, C.; McNeal, A.; Davidson, E.; Turner, H.L.; Vu, H.; Shulenin, S.; He, S.; Kuehne, A.; et al. Cooperativity Enables Non-neutralizing Antibodies to Neutralize Ebolavirus. Cell Rep. 2017, 19, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Karadag, M.; Arslan, M.; Kaleli, N.E.; Kalyoncu, S. Chapter Four—Physicochemical determinants of antibody-protein interactions. Adv. Protein Chem. Struct. Biol. 2020, 121, 85–114. [Google Scholar] [CrossRef]
- Ocaña, C.; Hayat, A.; Mishra, R.; Vasilescu, A.; del Valle, M.; Marty, J.-L. A novel electrochemical aptamer–antibody sandwich assay for lysozyme detection. Analyst 2015, 140, 4148–4153. [Google Scholar] [CrossRef]
- Dreymann, N.; Sabrowski, W.; Danso, J.; Menger, M.M. Aptamer-Based Sandwich Assay Formats for Detection and Discrimination of Human High- and Low-Molecular-Weight uPA for Cancer Prognosis and Diagnosis. Cancers 2022, 14, 5222. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kang, M.; Ban, C. Aptamer-antibody hybrid ELONA that uses hybridization chain reaction to detect a urinary biomarker EN2 for bladder and prostate cancer. Sci. Rep. 2022, 12, 11523. [Google Scholar] [CrossRef] [PubMed]
- Seiler, L.K.; Phung, N.L.; Nikolin, C.; Immenschuh, S.; Erck, C.; Kaufeld, J.; Haller, H.; Falk, C.S.; Jonczyk, R.; Lindner, P.; et al. An Antibody-Aptamer-Hybrid Lateral Flow Assay for Detection of CXCL9 in Antibody-Mediated Rejection after Kidney Transplantation. Diagnostics 2022, 12, 308. [Google Scholar] [CrossRef]
- Svigelj, R.; Dossi, N.; Grazioli, C.; Toniolo, R. Paper-based aptamer-antibody biosensor for gluten detection in a deep eutectic solvent (DES). Anal. Bioanal. Chem. 2022, 414, 3341–3348. [Google Scholar] [CrossRef]
- Uversky, V.N.; Van Regenmortel, M.H.V. Mobility and disorder in antibody and antigen binding sites do not prevent immunochemical recognition. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 149–156. [Google Scholar] [CrossRef]
- Poongavanam, M.-V.; Kisley, L.; Kourentzi, K.; Landes, C.F.; Willson, R.C. Ensemble and single-molecule biophysical characterization of D17.4 DNA aptamer–IgE interactions. Biochim. Biophys. Acta Proteins Proteom. 2016, 1864, 154–164. [Google Scholar] [CrossRef]
- Nguyen, H.; Lan, P.D.; Nissley, D.A.; O’Brien, E.P.; Li, M.S. Electrostatic Interactions Explain the Higher Binding Affinity of the CR3022 Antibody for SARS-CoV-2 than the 4A8 Antibody. J. Phys. Chem. B 2021, 125, 7368–7379. [Google Scholar] [CrossRef]
- Tuncbag, N.; Gursoy, A.; Keskin, O. Identification of computational hot spots in protein interfaces: Combining solvent accessibility and inter-residue potentials improves the accuracy. Bioinformatics 2009, 25, 1513–1520. [Google Scholar] [CrossRef]
- Bjerregaard, N.; Andreasen, P.A.; Dupont, D.M. Expected and unexpected features of protein-binding RNA aptamers. Wiley Interdiscip. Rev. RNA 2016, 7, 744–757. [Google Scholar] [CrossRef]
- Wilson, K.A.; Holland, D.J.; Wetmore, S.D. Topology of RNA-protein nucleobase-amino acid pi-pi interactions and comparison to analogous DNA-protein pi-pi contacts. RNA 2016, 22, 696–708. [Google Scholar] [CrossRef]
- Qiu, C.; Dutcher, R.C.; Porter, D.F.; Arava, Y.; Wickens, M.; Hall, T.M.T. Distinct RNA-binding modules in a single PUF protein cooperate to determine RNA specificity. Nucleic Acids Res. 2019, 47, 8770–8784. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.A.; Kung, R.W.; D’Souza, S.; Wetmore, S.D. Anatomy of noncovalent interactions between the nucleobases or ribose and pi-containing amino acids in RNA-protein complexes. Nucleic Acids Res. 2021, 49, 2213–2225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Chen, Z.H.; Liu, D.D.; Jiang, H.W.; Zhang, Z.K.; Lu, A.P.; Zhang, B.T.; Yu, Y.Y.; Zhang, G. Structural Biology for the Molecular Insight between Aptamers and Target Proteins. Int. J. Mol. Sci. 2021, 22, 4093. [Google Scholar] [CrossRef] [PubMed]
- Brouillette, R.B.; Maury, W. Production of Filovirus Glycoprotein-Pseudotyped Vesicular Stomatitis Virus for Study of Filovirus Entry Mechanisms. Methods Mol. Biol. 2017, 1628, 53–63. [Google Scholar] [CrossRef]
- Moller-Tank, S.; Albritton, L.M.; Rennert, P.D.; Maury, W. Characterizing functional domains for TIM-mediated enveloped virus entry. J. Virol. 2014, 88, 6702–6713. [Google Scholar] [CrossRef]
- Rhein, B.A.; Brouillette, R.B.; Schaack, G.A.; Chiorini, J.A.; Maury, W. Characterization of Human and Murine T-Cell Immunoglobulin Mucin Domain 4 (TIM-4) IgV Domain Residues Critical for Ebola Virus Entry. J. Virol. 2016, 90, 6097–6111. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Nilsen-Hamilton, M.; Shapiro, J.M.; Massoglia, S.L.; Hamilton, R.T. Selective stimulation by mitogens of incorporation of 35S-methionine into a family of proteins released into the medium by 3T3 cells. Cell 1980, 20, 19–28. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- PDB. Available online: https://doi.org/10.2210/pdb5kem/pdb (accessed on 6 October 2022).
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef] [PubMed]
- PDB. Available online: https://doi.org/10.2210/pdb5ken/pdb (accessed on 20 October 2022).
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Markham, N.R.; Zuker, M. UNAFold. In Bioinformatics: Structure, Function and Applications; Keith, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 3–31. [Google Scholar]
- Wang, J.; Wang, J.; Huang, Y.; Xiao, Y. 3dRNA v2.0: An Updated Web Server for RNA 3D Structure Prediction. Int. J. Mol. Sci. 2019, 20, 4116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiong, Y.; Xiao, Y. 3dDNA: A Computational Method of Building DNA 3D Structures. Molecules 2022, 27, 5936. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Zou, X. A knowledge-based scoring function for protein-RNA interactions derived from a statistical mechanics-based iterative method. Nucleic Acids Res. 2014, 42, e55. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, D.; Zhou, P.; Li, B.; Huang, S.Y. HDOCK: A web server for protein-protein and protein-DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res. 2017, 45, W365–W373. [Google Scholar] [CrossRef]
- The PyMol Molecular Graphics System V.S., LLC. Available online: https://pymol.org/2/ (accessed on 16 September 2022).
- InterfaceResidues. Available online: https://pymolwiki.org/index.php/InterfaceResidues (accessed on 17 October 2022).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Daniel, C.; Roupioz, Y.; Gasparutto, D.; Livache, T.; Buhot, A. Solution-Phase vs Surface-Phase Aptamer-Protein Affinity from a Label-Free Kinetic Biosensor. PLoS ONE 2013, 8, e75419. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, N.; Guo, W.; Xia, F.; Jiang, L. Highly-Efficient Gating of Solid-State Nanochannels by DNA Supersandwich Structure Containing ATP Aptamers: A Nanofluidic IMPLICATION Logic Device. J. Am. Chem. Soc. 2012, 134, 15395–15401. [Google Scholar] [CrossRef]
- Kumeria, T.; Santos, A.; Losic, D. Nanoporous Anodic Alumina Platforms: Engineered Surface Chemistry and Structure for Optical Sensing Applications. Sensors 2014, 14, 11878–11918. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, J.C.; Yue, M.; Castelino, K.; Chakraborty, A.; Majumdar, A. Chemomechanics of Surface Stresses Induced by DNA Hybridization. Langmuir 2006, 22, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gosai, A.; Balasubramanian, G.; Shrotriya, P. Aptamer based electrostatic-stimuli responsive surfaces for on-demand binding/unbinding of a specific ligand. J. Mater. Chem. B 2017, 5, 3675–3685. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhai, T.; Gao, P.; Cheng, H.; Hou, R.; Lou, X.; Xia, F. Role of outer surface probes for regulating ion gating of nanochannels. Nat. Commun. 2018, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Devarakonda, S.; Kim, S.; Ganapathysubramanian, B.; Shrotriya, P. Designing asymmetrically modified nanochannel sensors using virtual EIS. Electrochim. Acta 2021, 403, 139694. [Google Scholar] [CrossRef]
- Gosai, A. Engineering of Nano-Bio Interfaces Towards the Development of Portable Biosensors. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar]
- Duan, D.; Fan, K.; Zhang, D.; Tan, S.; Liang, M.; Liu, Y.; Zhang, J.; Zhang, P.; Liu, W.; Qiu, X.; et al. Nanozyme-strip for rapid local diagnosis of Ebola. Biosens. Bioelectron. 2015, 74, 134–141. [Google Scholar] [CrossRef]
- Yen, C.-W.; de Puig, H.; Tam, J.O.; Gómez-Márquez, J.; Bosch, I.; Hamad-Schifferli, K.; Gehrke, L. Multicolored silver nanoparticles for multiplexed disease diagnostics: Distinguishing dengue, yellow fever, and Ebola viruses. Lab-On-A-Chip 2015, 15, 1638–1641. [Google Scholar] [CrossRef]
- Daaboul, G.G.; Lopez, C.A.; Chinnala, J.; Goldberg, B.B.; Connor, J.H.; Ünlü, M.S. Digital Sensing and Sizing of Vesicular Stomatitis Virus Pseudotypes in Complex Media: A Model for Ebola and Marburg Detection. ACS Nano 2014, 8, 6047–6055. [Google Scholar] [CrossRef]
- Yanik, A.A.; Huang, M.; Kamohara, O.; Artar, A.; Geisbert, T.W.; Connor, J.H.; Altug, H. An Optofluidic Nanoplasmonic Biosensor for Direct Detection of Live Viruses from Biological Media. Nano Lett. 2010, 10, 4962–4969. [Google Scholar] [CrossRef]
- Cai, H.; Parks, J.W.; Wall, T.A.; Stott, M.A.; Stambaugh, A.; Alfson, K.; Griffiths, A.; Mathies, R.A.; Carrion, R.; Patterson, J.L.; et al. Optofluidic analysis system for amplification-free, direct detection of Ebola infection. Sci. Rep. 2015, 5, 14494. [Google Scholar] [CrossRef]
- Chen, Y.; Ren, R.; Pu, H.; Guo, X.; Chang, J.; Zhou, G.; Mao, S.; Kron, M.; Chen, J. Field-Effect Transistor Biosensor for Rapid Detection of Ebola Antigen. Sci. Rep. 2017, 7, 10974. [Google Scholar] [CrossRef] [PubMed]
- Benzine, J.W.; Brown, K.M.; Agans, K.N.; Godiska, R.; Mire, C.E.; Gowda, K.; Converse, B.; Geisbert, T.W.; Mead, D.A.; Chander, Y. Molecular Diagnostic Field Test for Point-of-Care Detection of Ebola Virus Directly From Blood. J. Infect. Dis. 2016, 214, S234–S242. [Google Scholar] [CrossRef] [PubMed]
- Zang, F.; Su, Z.; Zhou, L.; Konduru, K.; Kaplan, G.; Chou, S.Y. Ultrasensitive Ebola Virus Antigen Sensing via 3D Nanoantenna Arrays. Adv. Mater. 2019, 31, 1902331. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Zhang, H.; Li, Y.-T.; Xiao, M.-M.; Zhang, Z.-L.; Pang, D.-W.; Wong, G.; Zhang, Z.-Y.; Zhang, G.-J. A field effect transistor modified with reduced graphene oxide for immunodetection of Ebola virus. Microchim. Acta 2019, 186, 223. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aptamer | Nucleic Acid Type | sGP, EBOV (Kd, nM) | sGP, SUDV (Kd, nM) | GP1,2, EBOV (Kd, nM) |
|---|---|---|---|---|
| 6011 | DNA | 8.5 ± 3.2 | 165 ± 44 | 48 ± 32 |
| 6012 | DNA | 27 ± 12 | 151 ± 44 | 54 ± 23 |
| 39SGP1A | 2′FY-RNA | * 13 ± 5 | * 147 ± 59 | 104 ± 31 |
| 70SGP2A | DNA | 27 ± 15 | ND | 58 ± 17 |
| Oligonucleotide | Nucleic Acid Type | Sequence |
|---|---|---|
| SELEX library | DNA | GCCTGTTGTGAGCCTCCTGTCGAA (53N) TTGAGCGTTTATTCTTGTCTCCC |
| 6011 | DNA | GCCTGTTGTGAGCCTCCTGTCGAACAACCACTCATATCTACTACATGACTTGCTCCATTCTGTTCTTTCTCTACGCATTGAGCGTTTATTCTTGTCTCCC |
| 6012 | DNA | GCCTGTTGTGAGCCTCCTGTCGAACGTATTTCTTGCTTCCTTCCTTGCCGCGCACATTGCAGTATAAGTACCTGTCGTTGAGCGTTTATTCTTGTCTCCC |
| 39SGP1A [11] | 2′FY-RNA | GGGCGCUCAAUUUUUUAUUGCAUUUUUCUUUGAGCGCCC |
| 70SGP2A | DNA | GCCTGTTGTGAGCCTCCTGTCGAACAACCACTCATATCTACTACATGACTTGCTCCATTCTGTTCTTTCT |
| 6020 | DNA | GCCTGTTGTGAGCCTCCTGTCGAACATACCGTTCCACCCACATTTCAACCTTCATCCATCCTATTATTAGCCCACTCTTGAGCGTTTATTCTTGTCTCCC |
| 6022 | DNA | GCCTGTTGTGAGCCTCCTGTCGAACCCTATCTTGTTCATGCTATTCTCAATATTTTCGGTTCACTTACCGTCTGCCTTTGAGCGTTTATTCTTGTCTCCC |
| 485 | 5′ DNA primer for ssDNA library | GCCTGTTGTGAGCCTCCTGTCGAA |
| 5617 | 3′ DNA primer for ssDNA library | GGGAGACAAGAATAAACGCTC (this primer was biotinylated for SELEX) |
| 5197 | DNA template for IVT | TAATACGACTCACTATAGGGCGCTCAATTTTTTATTGCATTTTTCTTTGAGCGCCC |
| 5198 | DNA complement for IVT | GGGCGCTCAAAGAAAAATGCAATAA AAAATTGAGCGCCCTATAGTGAGTCGTATTA |
| NA53 [24] | DNA | AGCAGCACAGAGGTCAGATGCCGTGCGGATGTACAGGGACTTGGATAGTTTCTGACCTATGCGTGCTACCGTGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banerjee, S.; Hemmat, M.A.; Shubham, S.; Gosai, A.; Devarakonda, S.; Jiang, N.; Geekiyanage, C.; Dillard, J.A.; Maury, W.; Shrotriya, P.; et al. Structurally Different Yet Functionally Similar: Aptamers Specific for the Ebola Virus Soluble Glycoprotein and GP1,2 and Their Application in Electrochemical Sensing. Int. J. Mol. Sci. 2023, 24, 4627. https://doi.org/10.3390/ijms24054627
Banerjee S, Hemmat MA, Shubham S, Gosai A, Devarakonda S, Jiang N, Geekiyanage C, Dillard JA, Maury W, Shrotriya P, et al. Structurally Different Yet Functionally Similar: Aptamers Specific for the Ebola Virus Soluble Glycoprotein and GP1,2 and Their Application in Electrochemical Sensing. International Journal of Molecular Sciences. 2023; 24(5):4627. https://doi.org/10.3390/ijms24054627
Chicago/Turabian StyleBanerjee, Soma, Mahsa Askary Hemmat, Shambhavi Shubham, Agnivo Gosai, Sivaranjani Devarakonda, Nianyu Jiang, Charith Geekiyanage, Jacob A. Dillard, Wendy Maury, Pranav Shrotriya, and et al. 2023. "Structurally Different Yet Functionally Similar: Aptamers Specific for the Ebola Virus Soluble Glycoprotein and GP1,2 and Their Application in Electrochemical Sensing" International Journal of Molecular Sciences 24, no. 5: 4627. https://doi.org/10.3390/ijms24054627
APA StyleBanerjee, S., Hemmat, M. A., Shubham, S., Gosai, A., Devarakonda, S., Jiang, N., Geekiyanage, C., Dillard, J. A., Maury, W., Shrotriya, P., Lamm, M. H., & Nilsen-Hamilton, M. (2023). Structurally Different Yet Functionally Similar: Aptamers Specific for the Ebola Virus Soluble Glycoprotein and GP1,2 and Their Application in Electrochemical Sensing. International Journal of Molecular Sciences, 24(5), 4627. https://doi.org/10.3390/ijms24054627