
Recent Insights into the Functional Role of AMPA Receptors in the Oligodendrocyte Lineage Cells In Vivo


Abstract
1. Introduction
2. Loss-of-Function of AMPAR Subunits in OL Lineage Cells
2.1. In Vivo Experimental Systems


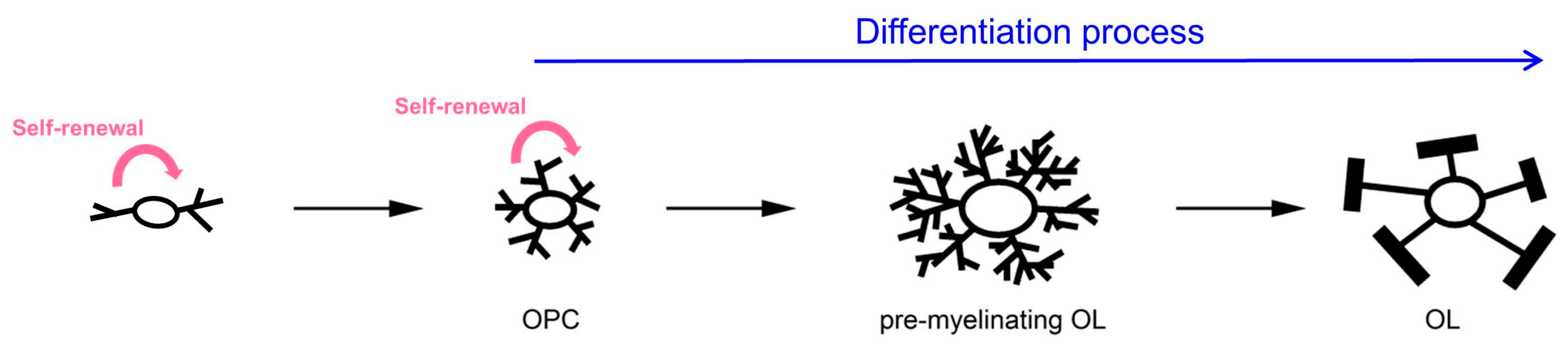{kind=link}
{kind=link}
{kind=link}
| Subunit | How AMPARs Were Targeted | In Which Cells AMPARs Were Targeted | Brain Areas Studied | How Targeting of AMPAR Was Verified | Consequences of AMPA Targeting | Reference |
|---|---|---|---|---|---|---|
| Loss of function of AMPAR subunits in the OL lineage cells | ||||||
| GluA2 | Conditional deletion of GluA2 by crossing Sox10-Cre and GluA2flox/flox mice | OL lineage cells | Subcortical white matter | Electro-physiology: I-V curve of kainate-induced current changed from linear to inwardly rectifying | No change in the density of OPCs and OLs; no change in proliferation rate of OPCs | Kougioumtzidou et al., 2017 [15] |
| GluA4 | Inducible deletion of GluA4 by crossing PLP-Cre and GluA4flox/flox mice | OLs | Optic nerve and the spinal cord white matter | Immuno-labelling: reduced GluA4 expression in CC1+-OLs; Ca2+-imaging: reduction in glutamate-induced somatic Ca2+-responses | No changes in normal mice, but changes during EAE: better preservation of axons, reduced production of amyloid precursor protein, reduced microglia labelling | Evonuk et al., 2020 [34] |
| GluA4 | Frameshift mutation in GluA4A using CRISPR/ Cas9 gene editing in zebrafish | OL lineage cells | Dorsal spinal cord | In situ hybridization: lower levels of GluA4A | Reduced net dorsal migration of OPCs; fewer myelin internodes per somite, and more myelin internodes per OL | Piller et al., 2021 [37] |
| GluA2, GluA3 | Conditional deletion of GluA2 on a Gria3 germline KO background by crossing GluA3 KO, Sox10-Cre, GluA2flox/flox mice | OL lineage cells | Subcortical white matter | Electro-physiology: reduced density of kainate-induced whole-cell current, I-V curve of kainate-induced whole-cell current changed from linear to inwardly rectifying; reduced number of synaptic contacts on OPCs | Reduced density of OLs at P7 and P14; reduced number of myelin figures at P14; increased apoptotic death of OL lineage cells at P7 and P14 | Kougioumtzidou et al., 2017 [15] |
| GluA2, GluA3, GluA4 | Conditional deletion of GluA2 and GluA4 on a Gria3 germline KO background by crossing Sox10-Cre, GluA2flox/flox, GluA4flox/flox mice | OL lineage cells | Subcortical white matter | Electro-physiology: reduced density of kainate-induced whole-cell current; strongly reduced frequency of synaptic axon-glia EPSCs in OPCs | Reduced density of OLs at P14 and P53; increased apoptotic death of OL lineage cells at P14 | Kougioumtzidou et al., 2017 [15] |
| Gain-of-function and modification of AMPAR subunits in the OL cells | ||||||
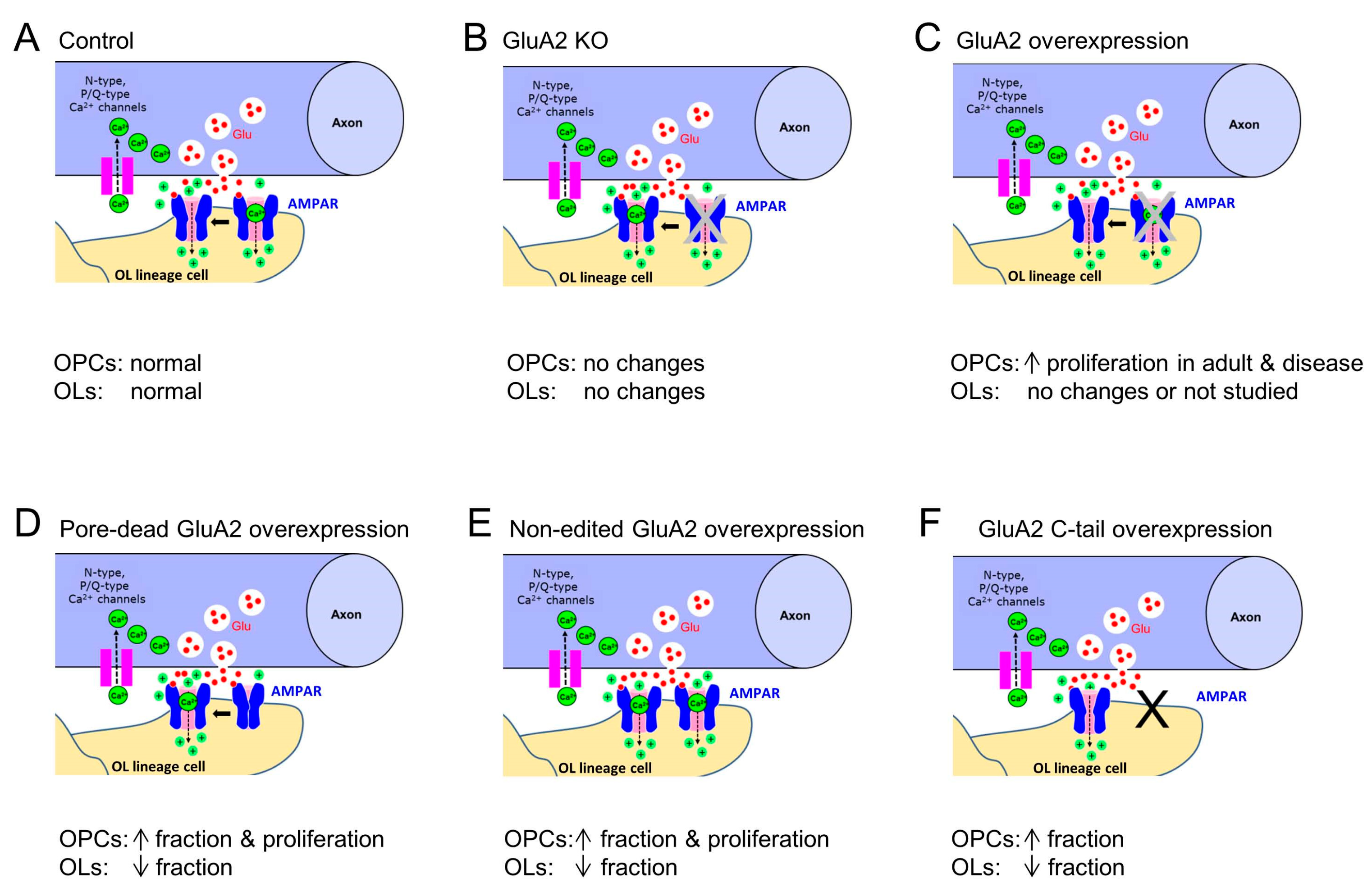
| GluA2 | Expression of the unedited GluA2(R583Q)-GFP subunit using retroviral gene delivery in vivo | Proliferating OPCs | Corpus callosum | Electro-physiology: I-V curve of axon-glia EPSCs changed from linear to inwardly rectifying; slight increase in quantal amplitude and single-channel conductance of AMPARs | Increased fraction of OPCs, decreased fraction of OLs, increased fraction of proliferating OPCs | Chen et al., 2018 [13] |
| GluA2 | Expression of the pore-dead GluA2(R583E)-GFP subunit using retroviral gene delivery in vivo | Proliferating OPCs | Corpus callosum | Electro-physiology: I-V curve of axon-glia EPSCs changed from linear to inwardly rectifying; slight increase in quantal amplitude and single-channel conductance of AMPARs | Increased fraction of OPCs, decreased fraction of OLs, increased fraction of proliferating OPCs | Chen et al., 2018 [13] |
| GluA2 | Expression of the cytoplasmic C-terminal (GluA2(813-862)) of GluA2 | Proliferating OPCs | Corpus callosum | Electro-physiology: no changes | Increased fraction of OPCs, decreased fraction of OLs | Chen et al., 2018 [13] |
| GluA2 | Inducible over-expression GluA2 subunit by crossing R26-Gria2 * and Sox10-CreER mice | OL lineage cells | Cortex, corpus callosum, external capsule | Calcium imaging in isolated brain OPCs: Ca2+ increases triggered in OPCs without GluA2 over-expression were much larger than in OPCs over-expressing the GluA2 subunit | No change in myelination in the young brain | Khawaja et al., 2021 [36] |
| GluA2 | Inducible over-expression GluA2 subunit by crossing R26-Gria2 * and Cspg4-CreER | OPCs | Cortex, corpus callosum, external capsule | RT-qPCR: OPCs over-expressing GluA2 subunit express more Gria2 by ~3-fold | Increased proliferation of OPCs in adult mice; increased post-injury proliferation of OPCs and increased OLs lineage progression in a mouse model of neonatal hypoxia-ischemia and upon focal demyelination | Khawaja et al., 2021 [36] |
| GluA2 | Inducible over-expression GluA2 subunit by crossing R26-Gria2 * and Plp1-CreER | OLs | Cortex, corpus callosum, external capsule | GluA2 overexpression in mature OLs does not prevent loss of OLs in a mouse model of neonatal hypoxia-ischemia | Khawaja et al., 2021 [36] | |
2.2. What Happens to Functional AMPARs in OL Lineage Cells upon Deletion of One or Multiple AMPAR Subunits?
3. How Does Loss of Function of AMPARs Affect the Behaviour and Function of the OL Lineage Cells?
3.1. Deletion of the GluA2 Subunit in Sox10+ OL Lineage Cells in Mice
3.2. Reduction in GluA4 Subunit Expression in Zebrafish
3.3. Deletion of the GluA4 Subunit in PLP+ Oligodendrocytes in Mice
3.4. Deletion of Multiple AMPAR Subunits in Sox10+ Cells in Mice
4. Gain-of-Function and Modification of AMPAR Subunits in OL Cells
4.1. In Vivo Experimental Systems
4.2. What Happens to Functional AMPARs in OL Lineage Cells upon Overexpression and Modification of AMPAR Subunits?
4.3. How Does the Overexpression and Modification of AMPAR Subunits Affect the Behaviour and Function of OPCs?
4.3.1. Targeting the GluA2 Subunit of AMPARs in Proliferating OPCs
4.3.2. Targeting the GluA2 Subunit of AMPARs in OL Lineage Cells
5. Agreements and Contradictions Regarding the Functional Role of AMPARs in OL Lineage Cells
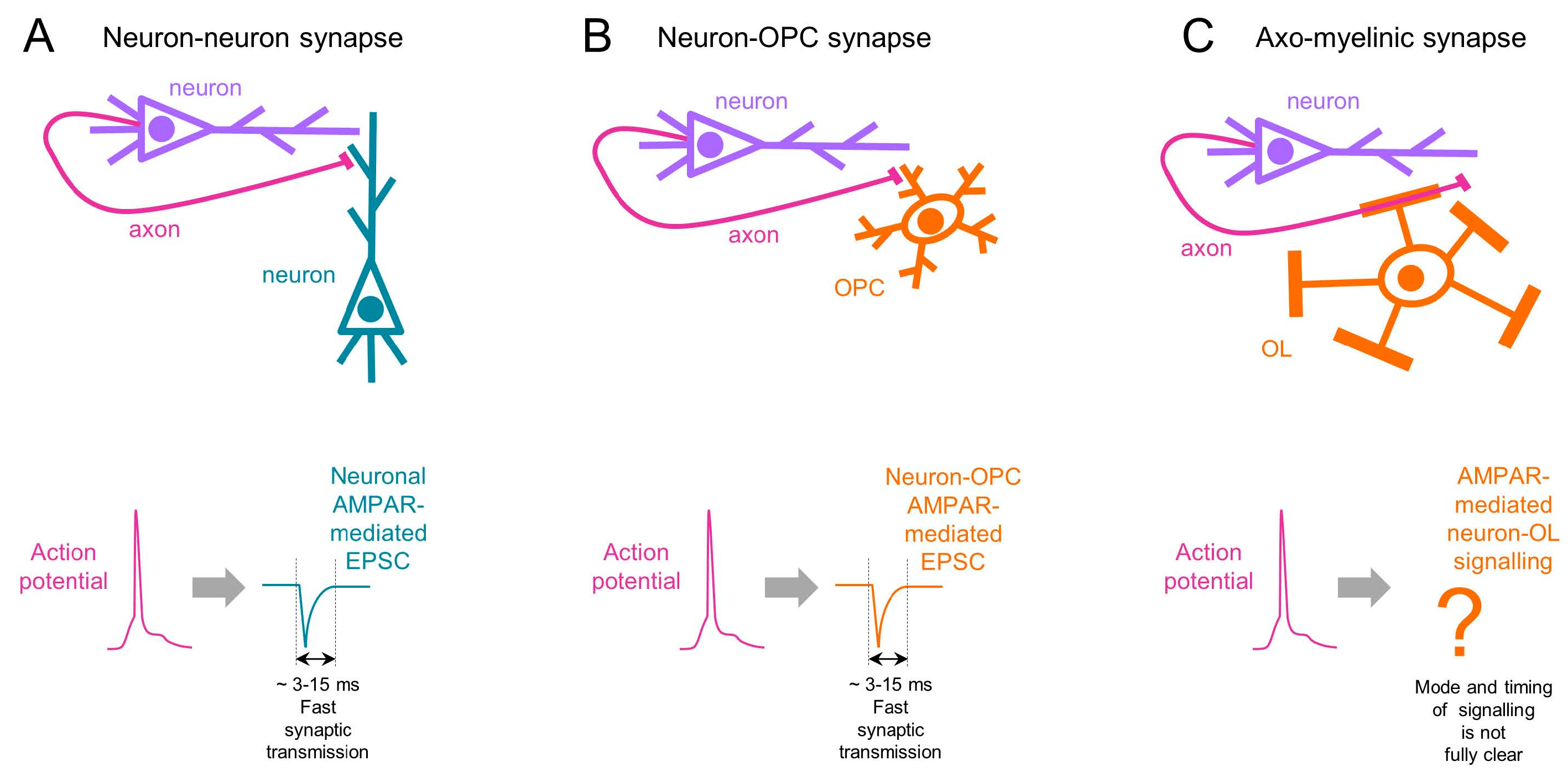
6. Temporal Aspects of AMPAR-Mediated Signalling in OL Lineage Cells
7. Spatial Aspects of AMPAR-Mediated Signalling in OL Lineage Cells
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Gianmarco, L.; Kurt, B.; Kukley, M. Technical approaches and challenges to study AMPA receptors in oligodendrocyte lineage cells: Past, present, and future. Glia 2022. [Google Scholar] [CrossRef] [PubMed]
- Bergles, D.E.; Roberts, J.D.; Somogyi, P.; Jahr, C.E. Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nature 2000, 405, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.P.; Yang, X.J.; Zhang, Z.; Wang, H.K.; Shen, W.; Deng, Q.D.; Duan, S. Long-term potentiation of neuron-glia synapses mediated by Ca2+-permeable AMPA receptors. Science 2006, 312, 1533–1537. [Google Scholar] [CrossRef] [PubMed]
- Kukley, M.; Nishiyama, A.; Dietrich, D. The fate of synaptic input to NG2 glial cells: Neurons specifically downregulate transmitter release onto differentiating oligodendroglial cells. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 8320–8331. [Google Scholar] [CrossRef] [PubMed]
- Passlick, S.; Trotter, J.; Seifert, G.; Steinhauser, C.; Jabs, R. The NG2 Protein Is Not Required for Glutamatergic Neuron-NG2 Cell Synaptic Signaling. Cereb. Cortex 2016, 26, 51–57. [Google Scholar] [CrossRef][Green Version]
- Hardt, S.; Tascio, D.; Passlick, S.; Timmermann, A.; Jabs, R.; Steinhauser, C.; Seifert, G. Auxiliary Subunits Control Function and Subcellular Distribution of AMPA Receptor Complexes in NG2 Glia of the Developing Hippocampus. Front. Cell. Neurosci. 2021, 15, 669717. [Google Scholar] [CrossRef]
- Mangin, J.M.; Li, P.; Scafidi, J.; Gallo, V. Experience-dependent regulation of NG2 progenitors in the developing barrel cortex. Nat. Neurosci. 2012, 15, 1192–1194. [Google Scholar] [CrossRef]
- Lin, S.C.; Huck, J.H.; Roberts, J.D.; Macklin, W.B.; Somogyi, P.; Bergles, D.E. Climbing fiber innervation of NG2-expressing glia in the mammalian cerebellum. Neuron 2005, 46, 773–785. [Google Scholar] [CrossRef]
- Zonouzi, M.; Renzi, M.; Farrant, M.; Cull-Candy, S.G. Bidirectional plasticity of calcium-permeable AMPA receptors in oligodendrocyte lineage cells. Nat. Neurosci. 2011, 14, 1430–1438. [Google Scholar] [CrossRef]
- Etxeberria, A.; Mangin, J.M.; Aguirre, A.; Gallo, V. Adult-born SVZ progenitors receive transient synapses during remyelination in corpus callosum. Nat. Neurosci. 2010, 13, 287–289. [Google Scholar] [CrossRef]
- Kukley, M.; Capetillo-Zarate, E.; Dietrich, D. Vesicular glutamate release from axons in white matter. Nat. Neurosci. 2007, 10, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Nagy, B.; Hovhannisyan, A.; Barzan, R.; Chen, T.J.; Kukley, M. Different patterns of neuronal activity trigger distinct responses of oligodendrocyte precursor cells in the corpus callosum. PLoS Biol. 2017, 15, e2001993. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.J.; Kula, B.; Nagy, B.; Barzan, R.; Gall, A.; Ehrlich, I.; Kukley, M. In Vivo Regulation of Oligodendrocyte Precursor Cell Proliferation and Differentiation by the AMPA-Receptor Subunit GluA2. Cell Rep. 2018, 25, 852–861.e857. [Google Scholar] [CrossRef]
- Ziskin, J.L.; Nishiyama, A.; Rubio, M.; Fukaya, M.; Bergles, D.E. Vesicular release of glutamate from unmyelinated axons in white matter. Nat. Neurosci. 2007, 10, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Kougioumtzidou, E.; Shimizu, T.; Hamilton, N.B.; Tohyama, K.; Sprengel, R.; Monyer, H.; Attwell, D.; Richardson, W.D. Signalling through AMPA receptors on oligodendrocyte precursors promotes myelination by enhancing oligodendrocyte survival. eLife 2017, 6, e28080. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Reyes-Haro, D.; Pivneva, T.; Nolte, C.; Schaette, R.; Lubke, J.; Kettenmann, H. The principal neurons of the medial nucleus of the trapezoid body and NG2(+) glial cells receive coordinated excitatory synaptic input. J. Gen. Physiol. 2009, 134, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Osterstock, G.; Le Bras, B.; Arulkandarajah, K.H.; Le Corronc, H.; Czarnecki, A.; Mouffle, C.; Bullier, E.; Legendre, P.; Mangin, J.M. Axoglial synapses are formed onto pioneer oligodendrocyte precursor cells at the onset of spinal cord gliogenesis. Glia 2018, 66, 1678–1694. [Google Scholar] [CrossRef]
- Ge, W.P.; Zhou, W.; Luo, Q.; Jan, L.Y.; Jan, Y.N. Dividing glial cells maintain differentiated properties including complex morphology and functional synapses. Proc. Natl. Acad. Sci. USA 2009, 106, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Mangin, J.M.; Kukley, M.; Dietrich, D. Synapses on NG2-expressing progenitors in the brain: Multiple functions? J. Physiol. 2008, 586, 3767–3781. [Google Scholar] [CrossRef] [PubMed]
- Kukley, M.; Kiladze, M.; Tognatta, R.; Hans, M.; Swandulla, D.; Schramm, J.; Dietrich, D. Glial cells are born with synapses. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 2957–2969. [Google Scholar] [CrossRef]
- Itoh, T.; Beesley, J.; Itoh, A.; Cohen, A.S.; Kavanaugh, B.; Coulter, D.A.; Grinspan, J.B.; Pleasure, D. AMPA glutamate receptor-mediated calcium signaling is transiently enhanced during development of oligodendrocytes. J. Neurochem. 2002, 81, 390–402. [Google Scholar] [CrossRef]
- Karadottir, R.; Cavelier, P.; Bergersen, L.H.; Attwell, D. NMDA receptors are expressed in oligodendrocytes and activated in ischaemia. Nature 2005, 438, 1162–1166. [Google Scholar] [CrossRef]
- Micu, I.; Plemel, J.R.; Lachance, C.; Proft, J.; Jansen, A.J.; Cummins, K.; van Minnen, J.; Stys, P.K. The molecular physiology of the axo-myelinic synapse. Exp. Neurol. 2016, 276, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A.; Koroshetz, W.J.; Swartz, K.J.; Chun, L.L.; Corey, D.P. Ion channel expression by white matter glia: The O-2A glial progenitor cell. Neuron 1990, 4, 507–524. [Google Scholar] [CrossRef] [PubMed]
- Wyllie, D.J.; Mathie, A.; Symonds, C.J.; Cull-Candy, S.G. Activation of glutamate receptors and glutamate uptake in identified macroglial cells in rat cerebellar cultures. J. Physiol. 1991, 432, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.; Ohlemeyer, C.; Trotter, J.; Kettenmann, H. AMPA/kainate receptor activation in murine oligodendrocyte precursor cells leads to activation of a cation conductance, calcium influx and blockade of delayed rectifying K+ channels. Neuroscience 1994, 63, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.; Kettenmann, H. Blockade of K+ channels induced by AMPA/kainate receptor activation in mouse oligodendrocyte precursor cells is mediated by Na+ entry. J. Neurosci. Res. 1995, 42, 579–593. [Google Scholar] [CrossRef] [PubMed]
- De Biase, L.M.; Nishiyama, A.; Bergles, D.E. Excitability and synaptic communication within the oligodendrocyte lineage. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 3600–3611. [Google Scholar] [CrossRef] [PubMed]
- Pende, M.; Holtzclaw, L.A.; Curtis, J.L.; Russell, J.T.; Gallo, V. Glutamate regulates intracellular calcium and gene expression in oligodendrocyte progenitors through the activation of DL-alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors. Proc. Natl. Acad. Sci. USA 1994, 91, 3215–3219. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Zhou, J.M.; McBain, C.J.; Wright, P.; Knutson, P.L.; Armstrong, R.C. Oligodendrocyte progenitor cell proliferation and lineage progression are regulated by glutamate receptor-mediated K+ channel block. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 2659–2670. [Google Scholar] [CrossRef]
- Yuan, X.; Eisen, A.M.; McBain, C.J.; Gallo, V. A role for glutamate and its receptors in the regulation of oligodendrocyte development in cerebellar tissue slices. Development 1998, 125, 2901–2914. [Google Scholar] [CrossRef] [PubMed]
- Harlow, D.E.; Saul, K.E.; Komuro, H.; Macklin, W.B. Myelin Proteolipid Protein Complexes with alphav Integrin and AMPA Receptors In Vivo and Regulates AMPA-Dependent Oligodendrocyte Progenitor Cell Migration through the Modulation of Cell-Surface GluR2 Expression. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 12018–12032. [Google Scholar] [CrossRef] [PubMed]
- Fannon, J.; Tarmier, W.; Fulton, D. Neuronal activity and AMPA-type glutamate receptor activation regulates the morphological development of oligodendrocyte precursor cells. Glia 2015, 63, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Evonuk, K.S.; Doyle, R.E.; Moseley, C.E.; Thornell, I.M.; Adler, K.; Bingaman, A.M.; Bevensee, M.O.; Weaver, C.T.; Min, B.; DeSilva, T.M. Reduction of AMPA receptor activity on mature oligodendrocytes attenuates loss of myelinated axons in autoimmune neuroinflammation. Sci. Adv. 2020, 6, eaax5936. [Google Scholar] [CrossRef] [PubMed]
- Stolt, C.C.; Rehberg, S.; Ader, M.; Lommes, P.; Riethmacher, D.; Schachner, M.; Bartsch, U.; Wegner, M. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes Dev. 2002, 16, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Khawaja, R.R.; Agarwal, A.; Fukaya, M.; Jeong, H.K.; Gross, S.; Gonzalez-Fernandez, E.; Soboloff, J.; Bergles, D.E.; Kang, S.H. GluA2 overexpression in oligodendrocyte progenitors promotes postinjury oligodendrocyte regeneration. Cell Rep. 2021, 35, 109147. [Google Scholar] [CrossRef]
- Piller, M.; Werkman, I.L.; Brown, E.A.; Latimer, A.J.; Kucenas, S. Glutamate Signaling via the AMPAR Subunit GluR4 Regulates Oligodendrocyte Progenitor Cell Migration in the Developing Spinal Cord. J. Neurosci. Off. J. Soc. Neurosci. 2021, 41, 5353–5371. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Vissel, B. The essential role of AMPA receptor GluR2 subunit RNA editing in the normal and diseased brain. Front. Mol. Neurosci. 2012, 5, 34. [Google Scholar] [CrossRef]
- Gudz, T.I.; Komuro, H.; Macklin, W.B. Glutamate stimulates oligodendrocyte progenitor migration mediated via an alphav integrin/myelin proteolipid protein complex. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 2458–2466. [Google Scholar] [CrossRef]
- Paez, P.M.; Fulton, D.J.; Spreur, V.; Handley, V.; Campagnoni, A.T. Multiple kinase pathways regulate voltage-dependent Ca2+ influx and migration in oligodendrocyte precursor cells. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 6422–6433. [Google Scholar] [CrossRef]
- Fulton, D.; Paez, P.M.; Fisher, R.; Handley, V.; Colwell, C.S.; Campagnoni, A.T. Regulation of L-type Ca++ currents and process morphology in white matter oligodendrocyte precursor cells by golli-myelin proteins. Glia 2010, 58, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Hayashi, Y.; Esteban, J.A.; Malinow, R. Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell 2001, 105, 331–343. [Google Scholar] [CrossRef]
- Swanson, G.T.; Kamboj, S.K.; Cull-Candy, S.G. Single-channel properties of recombinant AMPA receptors depend on RNA editing, splice variation, and subunit composition. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Matthews, E.A.; Nicolas, V.; Schoch, S.; Dietrich, D. NG2 glial cells integrate synaptic input in global and dendritic calcium signals. eLife 2016, 5, e16262. [Google Scholar] [CrossRef]
- Bergles, D.E.; Dzubay, J.A.; Jahr, C.E. Glutamate transporter currents in bergmann glial cells follow the time course of extrasynaptic glutamate. Proc. Natl. Acad. Sci. USA 1997, 94, 14821–14825. [Google Scholar] [CrossRef] [PubMed]
- Heflin, J.K.; Sun, W. Novel Toolboxes for the Investigation of Activity-Dependent Myelination in the Central Nervous System. Front. Cell. Neurosci. 2021, 15, 769809. [Google Scholar] [CrossRef]
- Pruvost, M.; Moyon, S. Oligodendroglial Epigenetics, from Lineage Specification to Activity-Dependent Myelination. Life 2021, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, B.L.; Regehr, W.G. Timing of neurotransmission at fast synapses in the mammalian brain. Nature 1996, 384, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, H.S.; Morishita, W.; Geraghty, A.C.; Silverbush, D.; Gillespie, S.M.; Arzt, M.; Tam, L.T.; Espenel, C.; Ponnuswami, A.; Ni, L.; et al. Electrical and synaptic integration of glioma into neural circuits. Nature 2019, 573, 539–545. [Google Scholar] [CrossRef]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Korber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef]
- Spruston, N.; Jaffe, D.B.; Williams, S.H.; Johnston, D. Voltage- and space-clamp errors associated with the measurement of electrotonically remote synaptic events. J. Neurophysiol. 1993, 70, 781–802. [Google Scholar] [CrossRef] [PubMed]
- Micu, I.; Plemel, J.R.; Caprariello, A.V.; Nave, K.A.; Stys, P.K. Axo-myelinic neurotransmission: A novel mode of cell signalling in the central nervous system. Nat. Rev. Neurosci. 2018, 19, 49–58. [Google Scholar] [CrossRef]
- Hill, R.A.; Patel, K.D.; Goncalves, C.M.; Grutzendler, J.; Nishiyama, A. Modulation of oligodendrocyte generation during a critical temporal window after NG2 cell division. Nat. Neurosci. 2014, 17, 1518–1527. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Dietrich, D. Synaptic integration by NG2 cells. Front. Cell. Neurosci. 2013, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Humeau, Y.; Choquet, D. The next generation of approaches to investigate the link between synaptic plasticity and learning. Nat. Neurosci. 2019, 22, 1536–1543. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kukley, M. Recent Insights into the Functional Role of AMPA Receptors in the Oligodendrocyte Lineage Cells In Vivo. Int. J. Mol. Sci. 2023, 24, 4138. https://doi.org/10.3390/ijms24044138
Kukley M. Recent Insights into the Functional Role of AMPA Receptors in the Oligodendrocyte Lineage Cells In Vivo. International Journal of Molecular Sciences. 2023; 24(4):4138. https://doi.org/10.3390/ijms24044138
Chicago/Turabian StyleKukley, Maria. 2023. "Recent Insights into the Functional Role of AMPA Receptors in the Oligodendrocyte Lineage Cells In Vivo" International Journal of Molecular Sciences 24, no. 4: 4138. https://doi.org/10.3390/ijms24044138
APA StyleKukley, M. (2023). Recent Insights into the Functional Role of AMPA Receptors in the Oligodendrocyte Lineage Cells In Vivo. International Journal of Molecular Sciences, 24(4), 4138. https://doi.org/10.3390/ijms24044138