

miR-103-3p Regulates the Differentiation and Autophagy of Myoblasts by Targeting MAP4

 and
and
Abstract
1. Introduction
2. Results
2.1. Expression of miR-103-3p
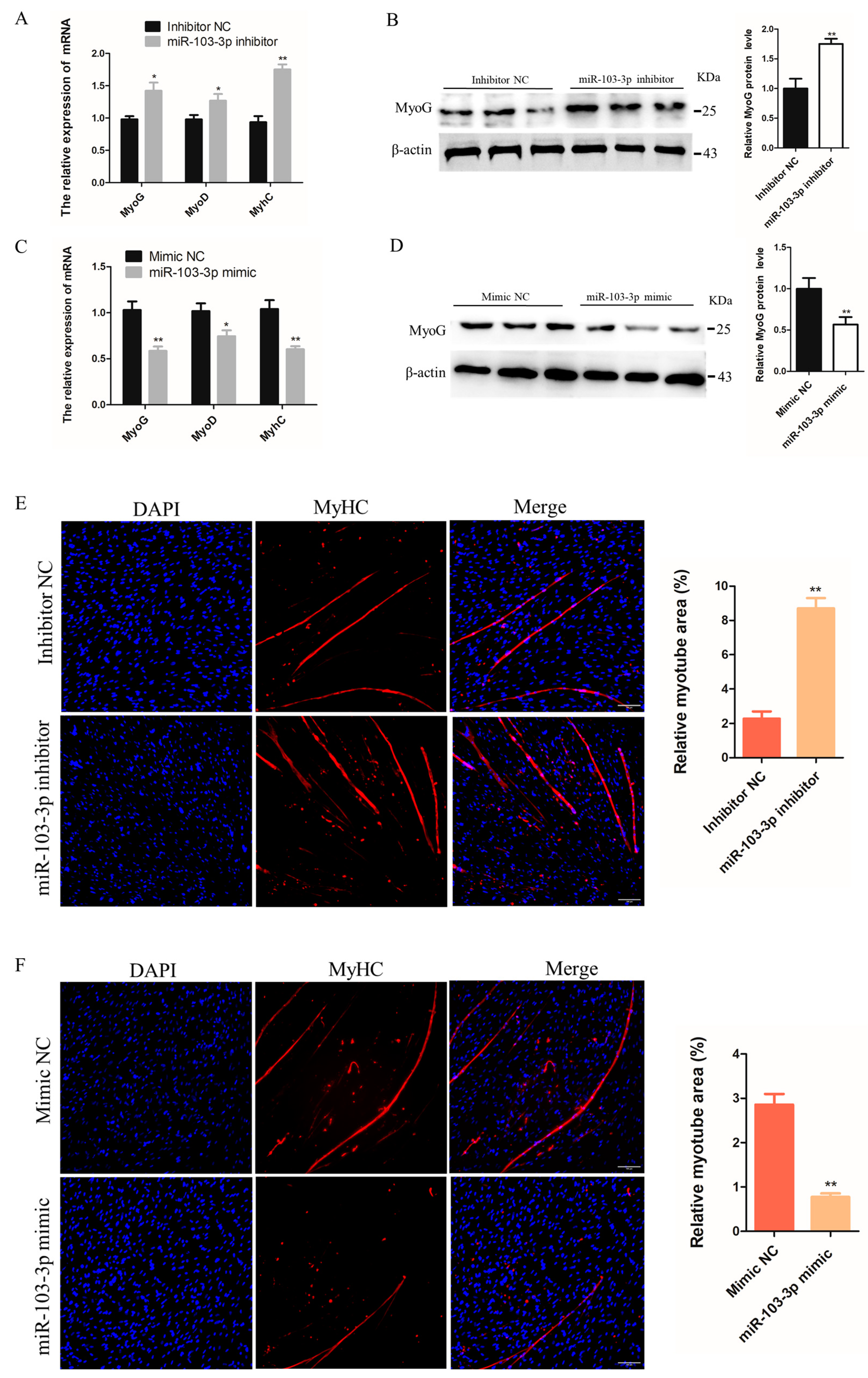
2.2. miR-103-3p Inhibited the Differentiation of C2C12 Cells
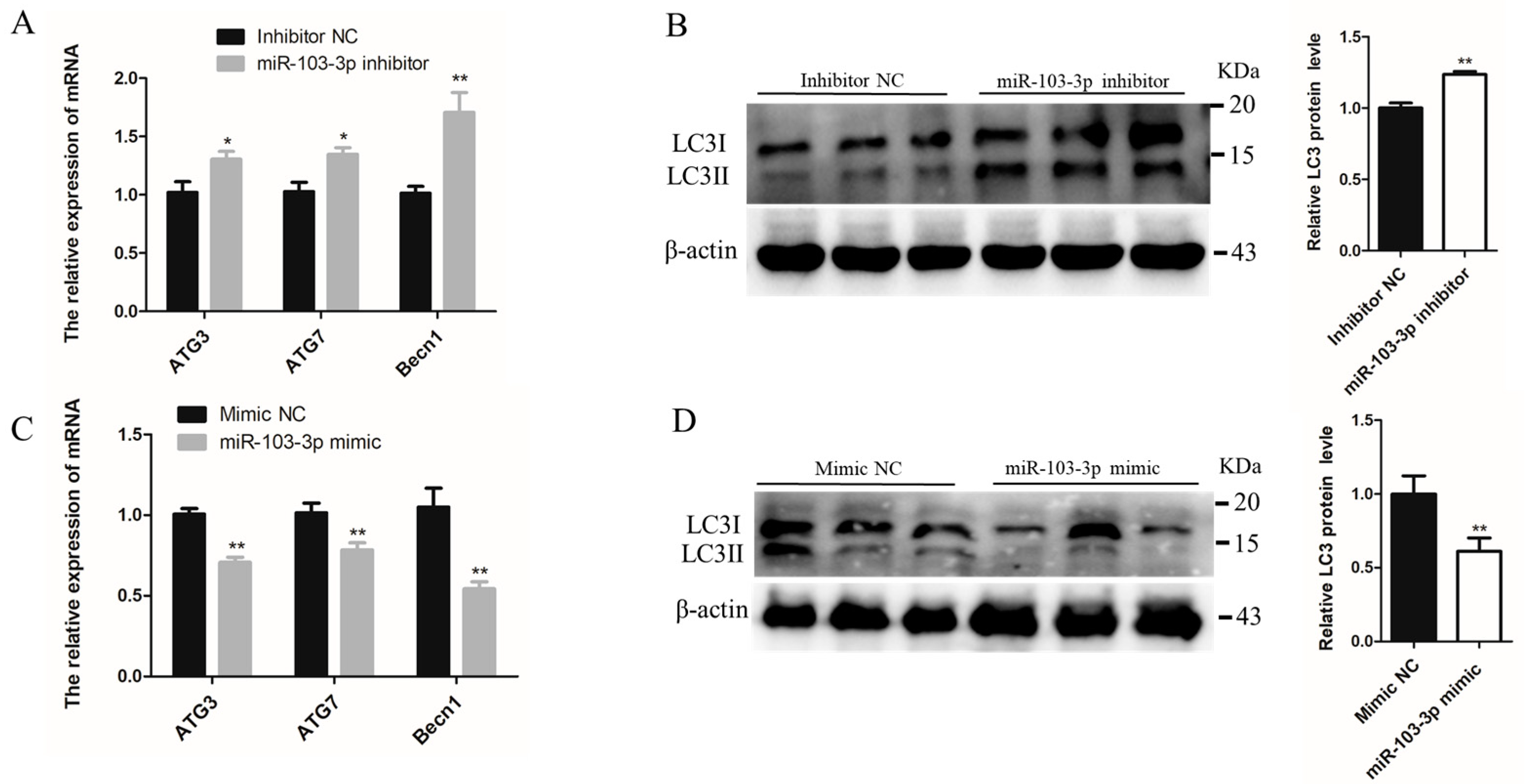
2.3. miR-103-3p Inhibited Autophagy of C2C12 Cells
2.4. miR-103-3p Suppressed MAP4 Expression
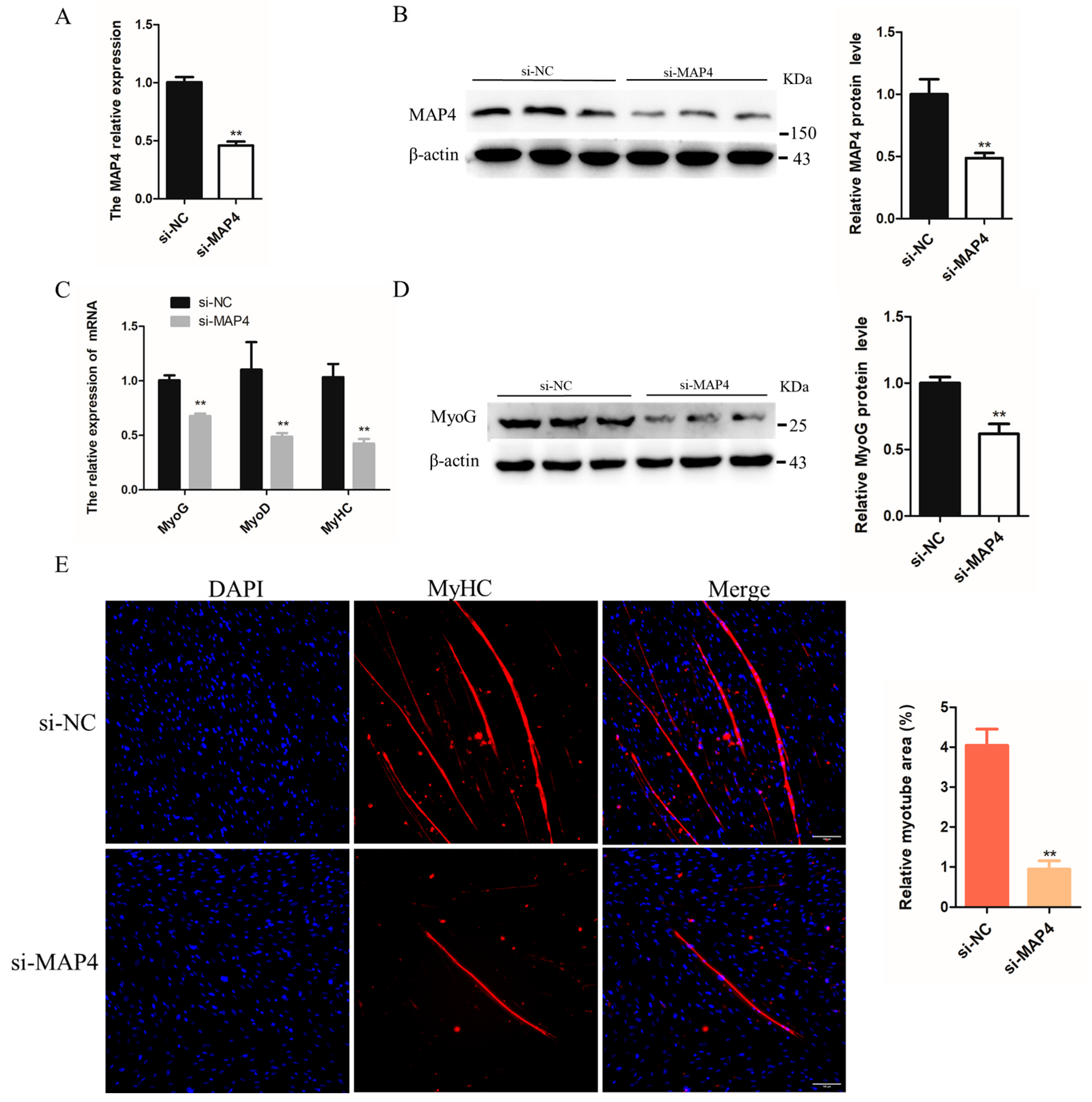
2.5. MAP4 Promoted the Differentiation of C2C12 Cells
2.6. MAP4 Induced Autophagy of C2C12 Cells
2.7. MAP4 Interacted with LC3
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Quantitative Real-Time PCR (qPCR)
4.3. Western Blot and Immunoprecipitation
4.4. Dual-Luciferase Reporter Assay
4.5. LC3-RFP-GFP Reporter Construct and Confocal Microscopy
4.6. Immunofluorescence
4.7. Transmission Electron Microscopy (TEM)
4.8. Animal Experiments
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diniz, P.G.; Wang, D.-Z. Regulation of Skeletal Muscle by microRNAs. Compr. Physiol. 2016, 3, 1279–1294. [Google Scholar]
- Zammit, S.P. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Beylkin, H.D.; Allen, L.D.; Leinwand, A.L. MyoD, Myf5, and the calcineurin pathway activate the developmental myosin heavy chain genes. Dev. Biol. 2006, 294, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Kassar Duchossoy, L.; Gayraud Morel, B.; Gomes, D.; Rocancourt, D.; Buckingham, M.; Shinin, V.; Tajbakhsh, S. Mrf4 determines skeletal muscle identity in Myf5:Myod double-mutant mice. Nature 2004, 431, 466. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Chen, D.; Yang, T.; Liu, G. Role of microRNA-27a in myoblast differentiation. Cell Biol. Int. 2014, 38, 266–271. [Google Scholar] [CrossRef]
- Cao, X.; Tang, S.; Du, F.; Li, H.; Shen, X.; Li, D.; Wang, Y.; Zhang, Z.; Xia, L.; Zhu, Q.; et al. miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken. Genes 2020, 11, 369. [Google Scholar] [CrossRef]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.Y. MicroRNA in Skeletal Muscle: Its Crucial Roles in Signal Proteins, Muscle Fiber Type, and Muscle Protein Synthesis. Curr. Protein Pept. Sci. 2017, 18, 579–588. [Google Scholar] [CrossRef]
- John, E.; Wienecke-Baldacchino, A.; Liivrand, M.; Heinäniemi, M.; Carlberg, C.; Sinkkonen, L. Dataset integration identifies transcriptional regulation of microRNA genes by PPARγ in differentiating mouse 3T3-L1 adipocytes. Nucleic Acids Res. 2012, 40, 4446–4460. [Google Scholar] [CrossRef]
- Han, B.; Lian, L.; Li, X.; Zhao, C.; Qu, L.; Liu, C.; Song, J.; Yang, N. Chicken gga-miR-103-3p Targets CCNE1 and TFDP2 and Inhibits MDCC-MSB1 Cell Migration. G3-Genes Genomes Genet. 2016, 6, 1277–1285. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, P.; Sun, L.; Lu, Y.; Zhu, W.; Zhang, J.; Xiang, C.; Mao, Y.; Chen, Q.; Zhang, F. Overexpression of circRNA SNRK targets miR-103-3p to reduce apoptosis and promote cardiac repair through GSK3β/β-catenin pathway in rats with myocardial infarction. Cell Death Discov. 2021, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Chen, T.; Zhu, Y.; Xiao, X.; Bu, J.; Huang, Z. MiR-103 alleviates autophagy and apoptosis by regulating SOX2 in LPS-injured PC12 cells and SCI rats. Iran. J. Basic Med. Sci. 2018, 21, 292–300. [Google Scholar] [PubMed]
- Zhang, X.-H.; Jin, G.-H.; Li, W.; Wang, S.-S.; Shan, B.-Q.; Qin, J.-B.; Zhao, H.-Y.; Tian, M.-L.; He, H.; Cheng, X. miR-103-3p targets Ndel1 to regulate neural stem cell proliferation and differentiation. Neural Regen. Res. 2022, 17, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Lu, C.; Shi, J.; Li, H.; Si, J.; Shen, G. Satb2 expression in Foxci-promoted osteogenic differentiation of MC3T3-E1 cells is negatively regulated by microRNA-103-3p. Acta Biochim. Biophys. Sin. 2019, 51, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Faruki, S.; Karsenti, E. Purification of microtubule proteins from Xenopus egg extracts: Identification of a 230K MAP4-like protein. Cell Motil. Cytoskelet. 1994, 28, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Mogessie, B.; Roth, D.; Rahil, Z.; Straube, A. A novel isoform of MAP4 organises the paraxial microtubule array required for muscle cell differentiation. Elife 2015, 4, e05697. [Google Scholar] [CrossRef]
- Mangan, E.M.; Olmsted, B.J. Muscle-specific variant of microtubule-associated protein 4 (MAP4) is required in myogenesis. Development 1996, 122, 771–781. [Google Scholar] [CrossRef]
- Straube, A.; Merdes, A. EB3 regulates microtubule dynamics at the cell cortex and is required for myoblast elongation and fusion. Curr. Biol. CB 2007, 17, 1318–1325. [Google Scholar] [CrossRef]
- Yang, Z.; Klionsky, J.D. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, H.; Qi, Y.; Kan, Y.; Ge, Z. Effects of miR-103a-3p on the autophagy and apoptosis of cardiomyocytes by regulating Atg5. Int. J. Mol. Med. 2019, 43, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Daniel, K.J. The Atg8 and Atg12 ubiquitin-like conjugation systems in macroautophagy. ‘Protein Modifications: Beyond the Usual Suspects’ Review Series. EMBO Rep. 2008, 9, 859–864. [Google Scholar] [CrossRef]
- Sin, J.; Andres, A.M.; Taylor, D.J.; Weston, T.; Hiraumi, Y.; Stotland, A.; Kim, B.J.; Huang, C.; Doran, K.S.; Gottlieb, R.A. Mitophagy is required for mitochondrial biogenesis and myogenic differentiation of C2C12 myoblasts. Autophagy 2016, 12, 369–380. [Google Scholar] [CrossRef]
- Baechler, B.L.; Bloemberg, D.; Quadrilatero, J. Mitophagy regulates mitochondrial network signaling, oxidative stress, and apoptosis during myoblast differentiation. Autophagy 2019, 15, 1606–1619. [Google Scholar] [CrossRef]
- Fortini, P.; Ferretti, C.; Iorio, E.; Cagnin, M.; Garribba, L.; Pietraforte, D.; Falchi, M.; Pascucci, B.; Baccarini, S.; Morani, F.; et al. The fine tuning of metabolism, autophagy and differentiation during in vitro myogenesis. Cell Death Dis. 2016, 7, e2168. [Google Scholar] [CrossRef]
- Tang, H.A.; Rando, A.T. Induction of autophagy supports the bioenergetic demands of quiescent muscle stem cell activation. EMBO J. 2014, 33, 2782–2797. [Google Scholar] [CrossRef]
- Chen, W.; Chen, Y.; Liu, Y.; Wang, X. Autophagy in muscle regeneration: Potential therapies for myopathies. J. Cachexia Sarcopenia Muscle 2022, 13, 1673–1685. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA Translation and Stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Lei, X.; Huang, Y.; Hu, J. MAP4 as a New Candidate in Cardiovascular Disease. Front. Physiol. 2020, 11, 1044. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Zhang, X.; Zhang, J.; Wang, X.; Ren, J.; Jia, J.; Zhang, D.; Jiang, X.; Zhang, J.; et al. Microtubule associated protein 4 phosphorylation leads to pathological cardiac remodeling in mice. EBioMedicine 2018, 37, 221–235. [Google Scholar] [CrossRef]
- Hu, J.; Chu, Z.; Han, J.; Zhang, Q.; Zhang, D.; Dang, Y.; Ren, J.; Chan, H.C.; Zhang, J.; Huang, Y. Phosphorylation-dependent mitochondrial translocation of MAP4 is an early step in hypoxia-induced apoptosis in cardiomyocytes. Cell Death Dis. 2014, 5, e1424. [Google Scholar] [CrossRef]
- Kumar, A.; Dhawan, A.; Kadam, A.; Shinde, A. Autophagy and Mitochondria: Targets in Neurodegenerative Disorders. CNS Neurol. Disord. Drug Targets 2018, 17, 696–705. [Google Scholar] [CrossRef]
- Tanida, I.; Tanida-Miyake, E.; Ueno, T.; Kominami, E. The human homolog of Saccharomyces cerevisiae Apg7p is a Protein-activating enzyme for multiple substrates including human Apg12p, GATE-16, GABARAP, and MAP-LC3. J. Biol. Chem. 2001, 276, 1701–1706. [Google Scholar] [CrossRef]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef]
- Köchl, R.; Hu, W.X.; Chan, E.Y.W.; Tooze, A.S. Microtubules facilitate autophagosome formation and fusion of autophagosomes with endosomes. Traffic 2006, 2, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Balcells, I.; Cirera, S.; Busk, P.K. Specific and sensitive quantitative RT-PCR of miRNAs with DNA primers. BMC Biotechnol. 2011, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Lee, A.; Sitharam, R.; Kesner, J.; Rabadan, R.; Shapira, S.D. Germ-Cell-Specific Inflammasome Component NLRP14 Negatively Regulates Cytosolic Nucleic Acid Sensing to Promote Fertilization. Immunity 2017, 46, 621–634. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′⟶3′) | Reverse Primer (5′⟶3′) |
|---|---|---|
| MAP4 | AGCTTTTGTTTCCAGCCTCA | TCTGAAGGATTGTCTCGCTCTG |
| β-actin | GTGACGTTGACATCCGTAAAGA | GCCGGACTCATCGTACTCC |
| MyoG | GGTGCCCAGTGAATGCAAC | AGATTGTGGGCGTCTGTAGG |
| MyoD | CGCCTGAGCAAAGTGAATGA | GCAGACCTTCGATGTAGCGG |
| MyHC | AAAATGAAGGGGACGCTGGAG | GTGGTGCCAAAATGGATGCG |
| LC3 | AGATCCCAGTGATTATAGAGCGA | CATGTTCACGTGGTCAGGCA |
| ATG3 | TCCCACCACCTCCTATGTGT | TATGAACACCAAGCTCTCCCC |
| ATG7 | CCCCATGCTCCTCAACAAGT | CTTCGGCTCGACACAGATCA |
| Becn1 | TGAATGAGGATGACAGTGAGCA | CACCTGGTTCTCCACACTCTTG |
| miR-103-3p | CAGCAUUGUACAGGGCUAUGA | TCCAGTTTTTTTTTTTTTTTCAAACAC |
| U6 | CTCGCTTCGGCAGCACA | TCCAGTTTTTTTTTTTTTTTCAAACAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Huang, S.; Niu, X.; Li, S.; Wang, J.; Ran, X. miR-103-3p Regulates the Differentiation and Autophagy of Myoblasts by Targeting MAP4. Int. J. Mol. Sci. 2023, 24, 4130. https://doi.org/10.3390/ijms24044130
Zhang X, Huang S, Niu X, Li S, Wang J, Ran X. miR-103-3p Regulates the Differentiation and Autophagy of Myoblasts by Targeting MAP4. International Journal of Molecular Sciences. 2023; 24(4):4130. https://doi.org/10.3390/ijms24044130
Chicago/Turabian StyleZhang, Xianxian, Shihui Huang, Xi Niu, Sheng Li, Jiafu Wang, and Xueqin Ran. 2023. "miR-103-3p Regulates the Differentiation and Autophagy of Myoblasts by Targeting MAP4" International Journal of Molecular Sciences 24, no. 4: 4130. https://doi.org/10.3390/ijms24044130
APA StyleZhang, X., Huang, S., Niu, X., Li, S., Wang, J., & Ran, X. (2023). miR-103-3p Regulates the Differentiation and Autophagy of Myoblasts by Targeting MAP4. International Journal of Molecular Sciences, 24(4), 4130. https://doi.org/10.3390/ijms24044130