
Development of Novel siRNA Therapeutics: A Review with a Focus on Inclisiran for the Treatment of Hypercholesterolemia



Abstract
1. Introduction
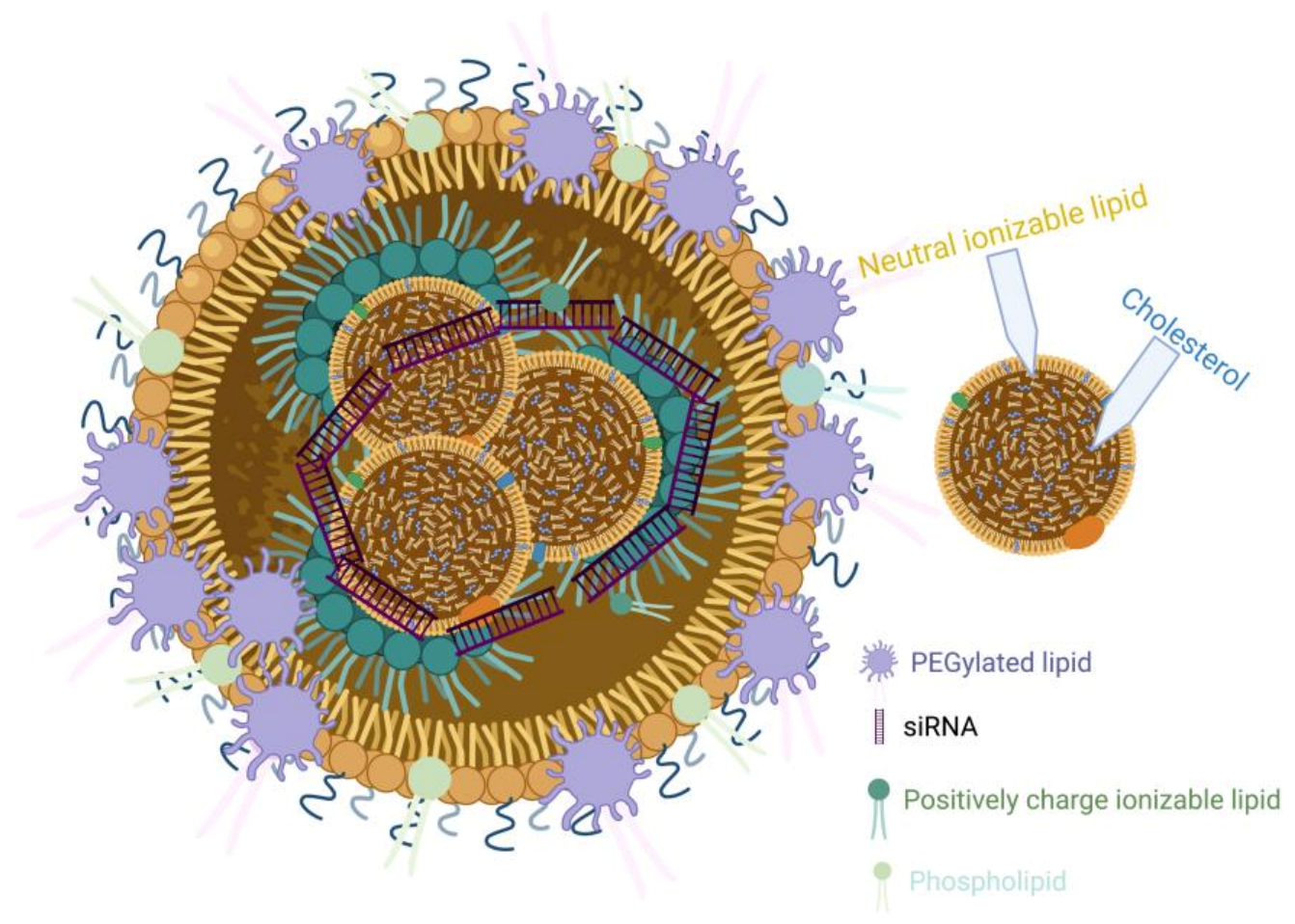
2. Lipid Nanoparticles (LNPs) and GalNAc Conjugates for siRNA Delivery
2.1. Lipid Nanoparticles (LNPs)-siRNA
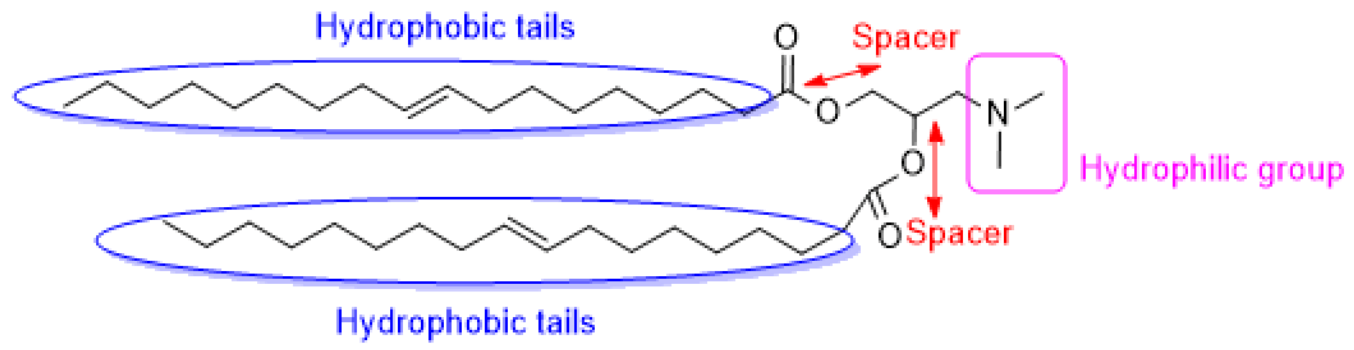
2.1.1. Ionizable Cationic Lipids
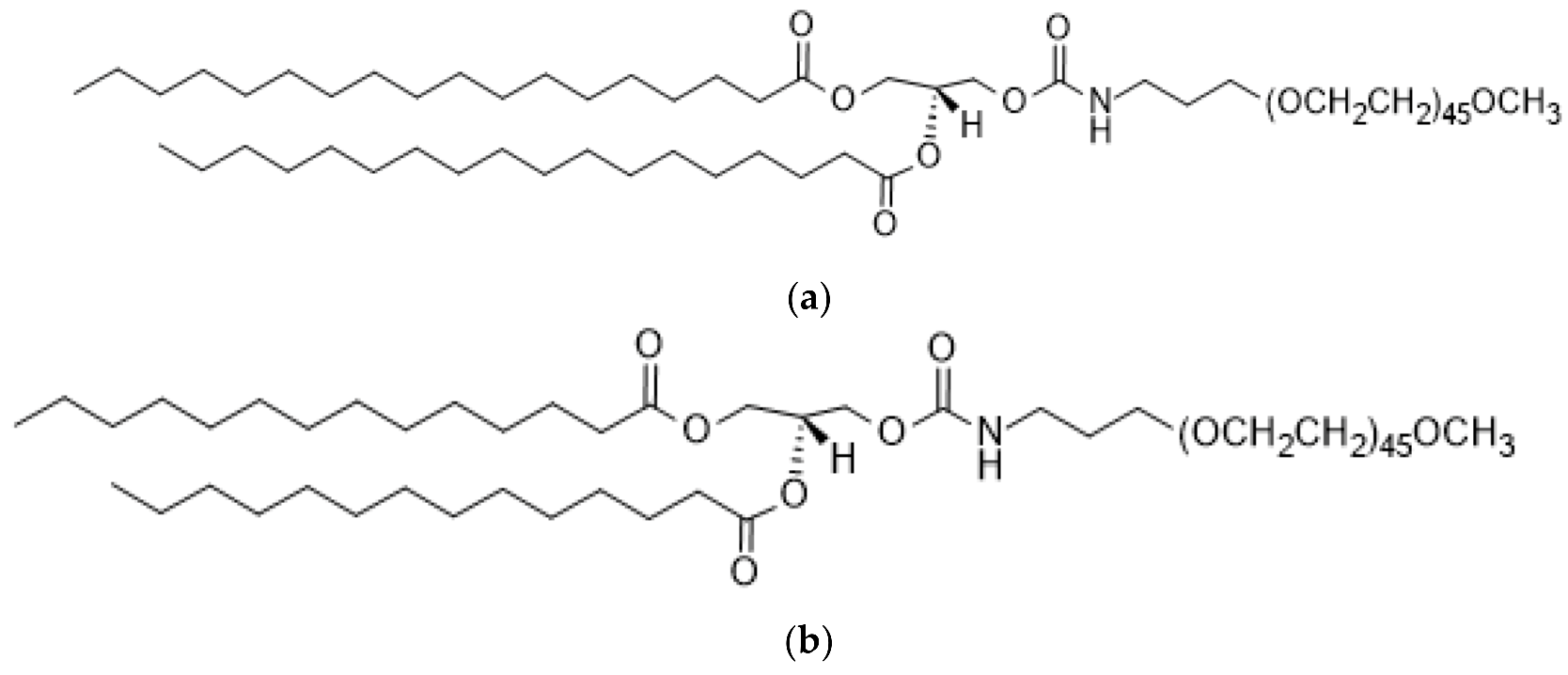
2.1.2. Poly (Ethylene Glycol) (PEG)
2.1.3. Cholesterol and Saturated Phosphatidylcholine (PC)
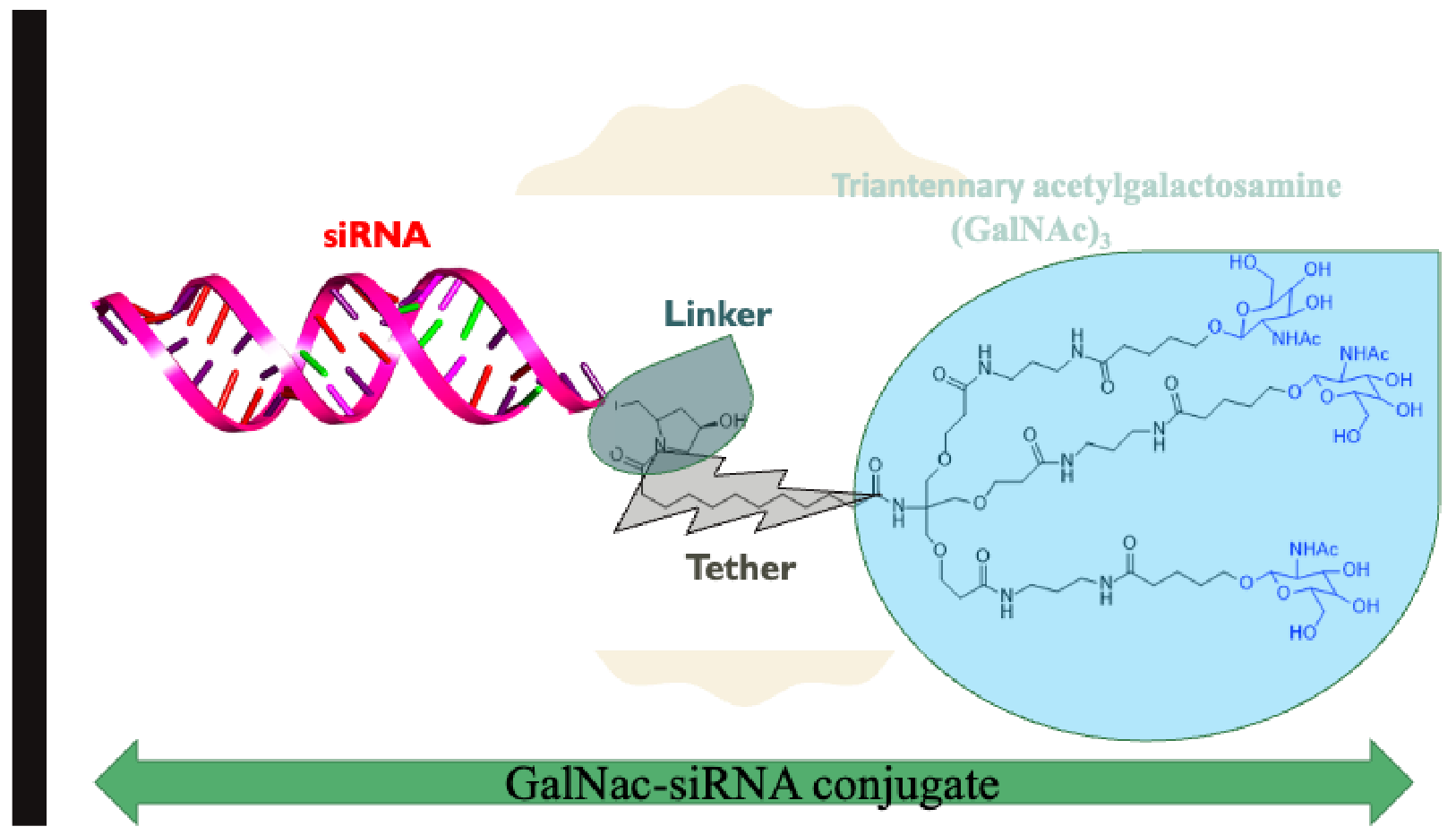
2.2. N-Acetylgalactosamine (GalNAc)-siRNA
3. Inclisiran
3.1. Some Selected Cholesterol-Lowering Drugs before Approval of Inclisiran
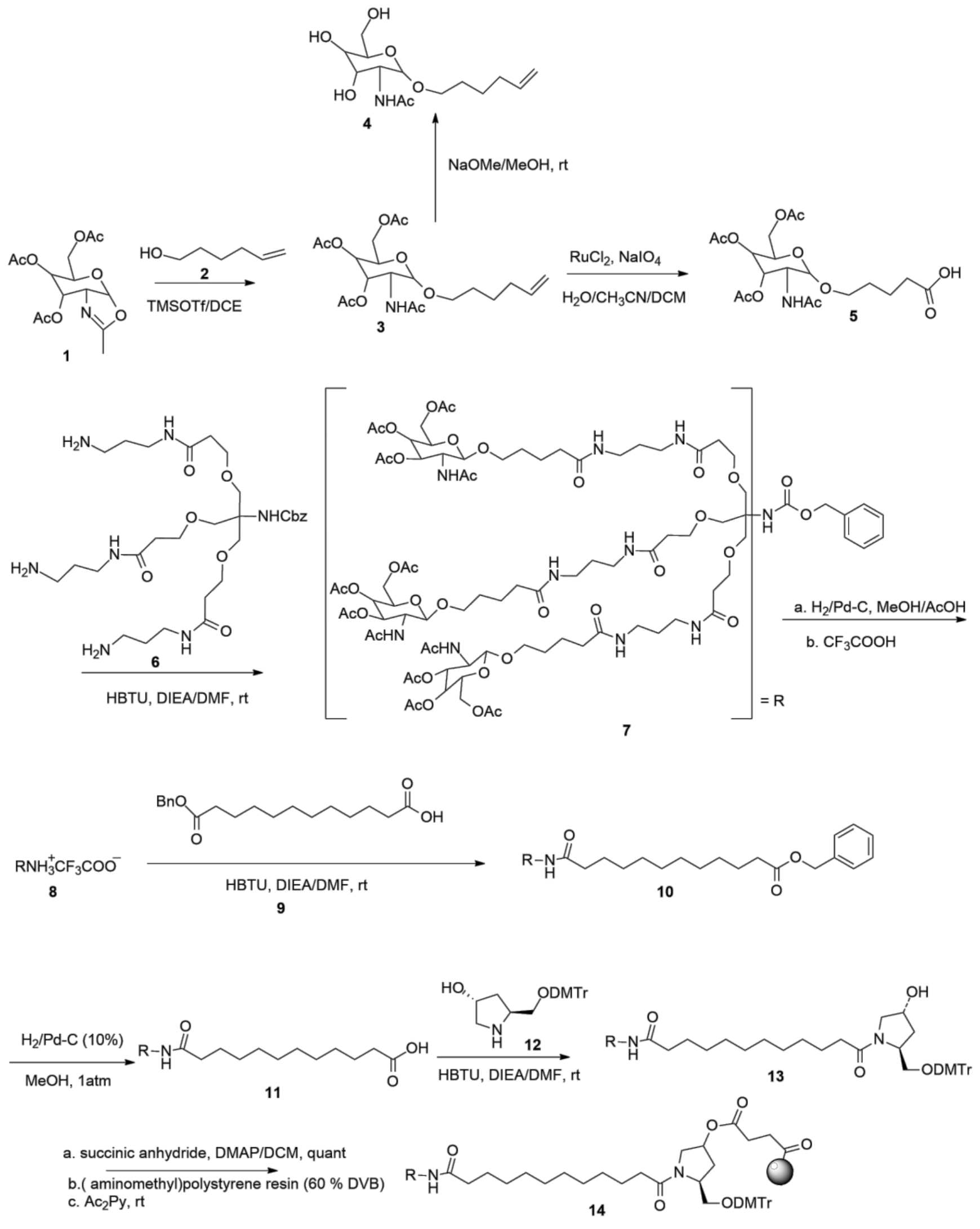
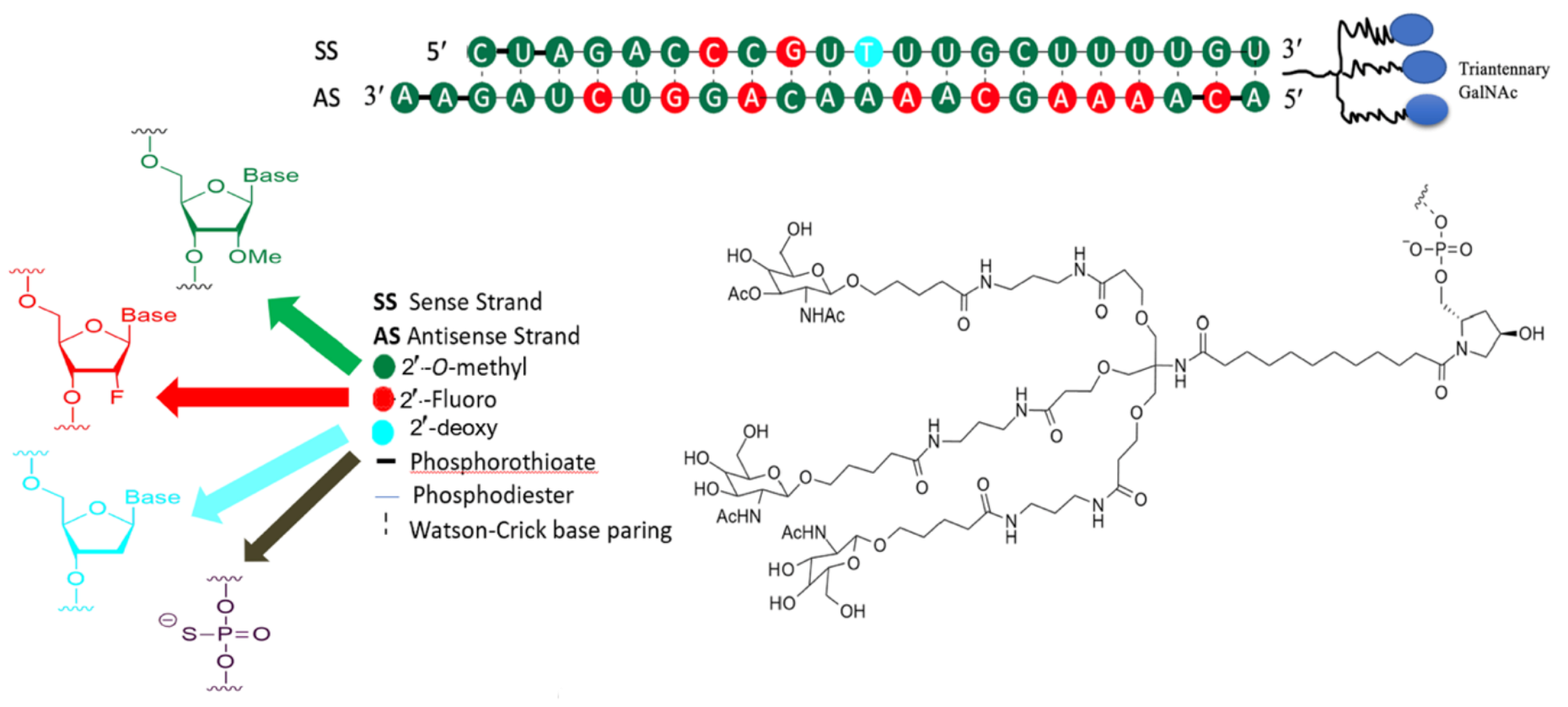
3.2. Development of Inclisiran
3.3. Clinical Trials of Inclisiran
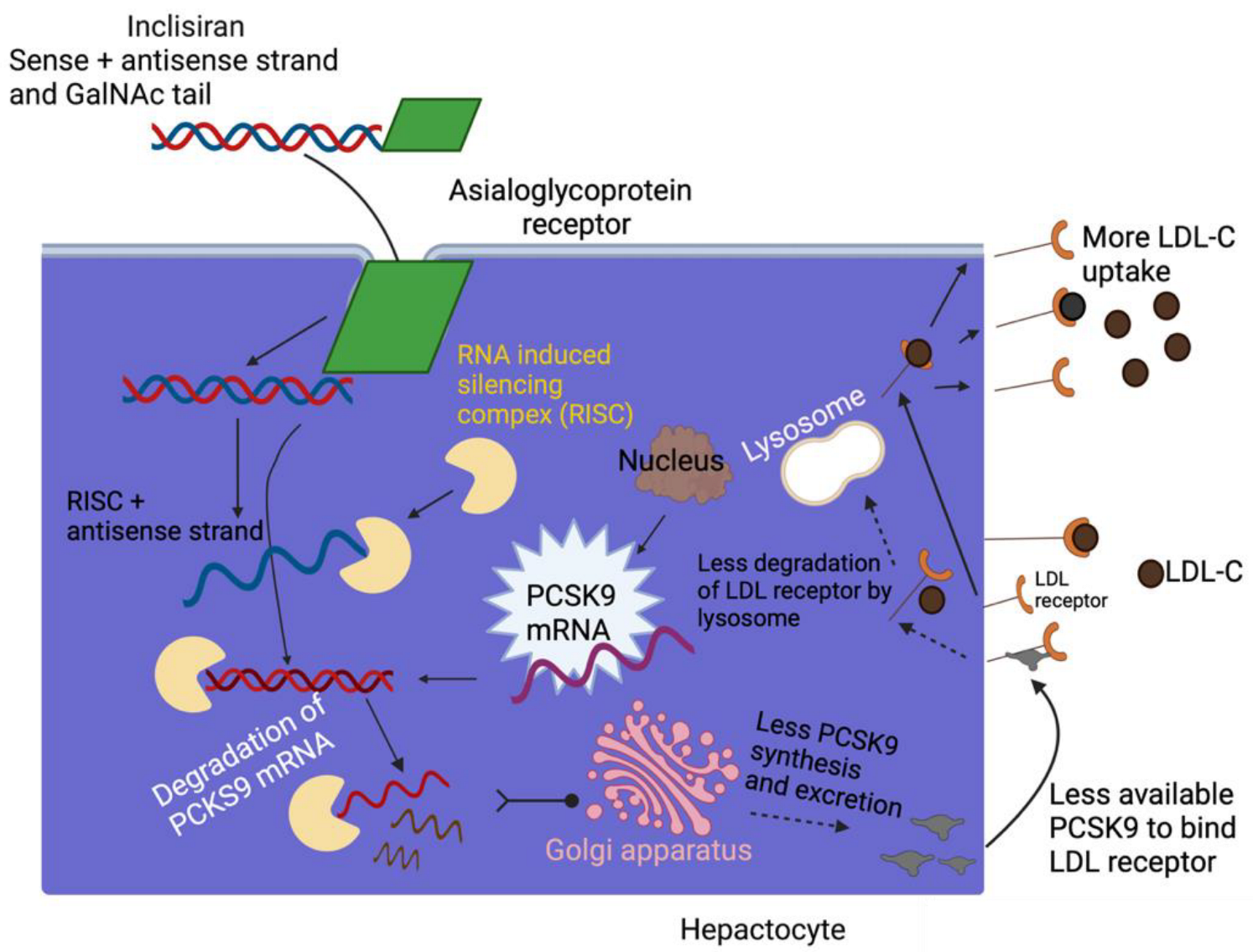
3.4. Inclisiran-Mechanism of Action
3.5. Clinical Benefits of Inclisiran
4. Other Critical Facts on siRNA Therapeutic and Inclisiran
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopkins, A.L.; Groom, C.R. The druggable genome. Nat. Rev. Drug Discov. 2002, 1, 727–730. [Google Scholar] [CrossRef]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.P.; Lampel, S. The druggable genome: An update. Drug Discov. Today 2005, 10, 1607–1610. [Google Scholar] [CrossRef]
- Hambly, K.; Danzer, J.; Muskal, S.; Debe, D.A. Interrogating the druggable genome with structural informatics. Mol. Divers. 2006, 10, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Cimermancic, P.; Weinkam, P.; Rettenmaier, T.J.; Bichmann, L.; Keedy, D.A.; Woldeyes, R.A.; Schneidman-Duhovny, D.; Demerdash, O.N.; Mitchell, J.C.; Wells, J.A.; et al. CryptoSite: Expanding the Druggable Proteome by Characterization and Prediction of Cryptic Binding Sites. J. Mol. Biol. 2016, 428, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewski, R.; Sikorski, A.F. Editorial focus: Understanding off-target effects as the key to successful RNAi therapy. Cell. Mol. Biol. Lett. 2019, 24, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X.B. The growth of siRNA-based therapeutics: Updated clinical studies. Biochem. Pharmacol. 2021, 189, 114432. [Google Scholar] [CrossRef]
- Flego, M.; Ascione, A.; Cianfriglia, M.; Vella, S. Clinical development of monoclonal antibody-based drugs in HIV and HCV diseases. BMC Med. 2013, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Weng, Y.; Xia, X.H.; Liang, X.J.; Huang, Y. Clinical advances of siRNA therapeutics. J. Gene Med. 2019, 21, e3097. [Google Scholar] [CrossRef] [PubMed]
- Schütze, N. siRNA technology. Mol. Cell. Endocrinol. 2004, 213, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Michler, T.; Merkel, O.M. siRNA Therapeutics against Respiratory Viral Infections-What Have We Learned for Potential COVID-19 Therapies? Adv. Healthc. Mater. 2021, 10, e2001650. [Google Scholar] [CrossRef] [PubMed]
- Ryther, R.C.; Flynt, A.S.; Phillips, J.A., 3rd; Patton, J.G. siRNA therapeutics: Big potential from small RNAs. Gene Ther. 2005, 12, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Dowdy, S.F. Overcoming cellular barriers for RNA therapeutics. Nat. Biotechnol. 2017, 35, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Juliano, R.L. The delivery of therapeutic oligonucleotides. Nucleic Acids Res. 2016, 44, 6518–6548. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Witztum, J.L.; Bennett, C.F.; Baker, B.F. RNA-Targeted Therapeutics. Cell Metab. 2018, 27, 714–739. [Google Scholar] [CrossRef]
- Springer, A.D.; Dowdy, S.F. GalNAc-siRNA Conjugates: Leading the Way for Delivery of RNAi Therapeutics. Nucleic Acid Ther. 2018, 28, 109–118. [Google Scholar] [CrossRef]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nat. Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef]
- Alexis, F.; Pridgen, E.; Molnar, L.K.; Farokhzad, O.C. Factors affecting the clearance and biodistribution of polymeric nanoparticles. Mol. Pharm. 2008, 5, 505–515. [Google Scholar] [CrossRef]
- Petros, R.A.; DeSimone, J.M. Strategies in the design of nanoparticles for therapeutic applications. Nat. Rev. Drug Discov. 2010, 9, 615–627. [Google Scholar] [CrossRef]
- Kanasty, R.L.; Whitehead, K.A.; Vegas, A.J.; Anderson, D.G. Action and reaction: The biological response to siRNA and its delivery vehicles. Mol. Ther. 2012, 20, 513–524. [Google Scholar] [CrossRef]
- Kanasty, R.; Dorkin, J.R.; Vegas, A.; Anderson, D. Delivery materials for siRNA therapeutics. Nat. Mater. 2013, 12, 967–977. [Google Scholar] [CrossRef]
- Chen, X.; Mangala, L.S.; Rodriguez-Aguayo, C.; Kong, X.; Lopez-Berestein, G.; Sood, A.K. RNA interference-based therapy and its delivery systems. Cancer Metastasis Rev. 2018, 37, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, K.; Rossi, J.J. RNAi-based therapeutics-current status, challenges and prospects. EMBO Mol. Med. 2009, 1, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, Z.; Wientjes, M.G.; Au, J.L. Delivery of siRNA therapeutics: Barriers and carriers. AAPS J. 2010, 12, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi Chalbatani, G.; Dana, H.; Gharagouzloo, E.; Grijalvo, S.; Eritja, R.; Logsdon, C.D.; Memari, F.; Miri, S.R.; Rad, M.R.; Marmari, V. Small interfering RNAs (siRNAs) in cancer therapy: A nano-based approach. Int. J. Nanomed. 2019, 14, 3111–3128. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.C.; Rossi, J.J.; Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnol. J. 2011, 6, 1130–1146. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://investors.alnylam.com/press-release?id=26816 (accessed on 26 September 2022).
- Landen, C.N., Jr.; Chavez-Reyes, A.; Bucana, C.; Schmandt, R.; Deavers, M.T.; Lopez-Berestein, G.; Sood, A.K. Therapeutic EphA2 gene targeting in vivo using neutral liposomal small interfering RNA delivery. Cancer Res. 2005, 65, 6910–6918. [Google Scholar] [CrossRef]
- Pal, A.; Ahmad, A.; Khan, S.; Sakabe, I.; Zhang, C.; Kasid, U.N.; Ahmad, I. Systemic delivery of RafsiRNA using cationic cardiolipin liposomes silences Raf-1 expression and inhibits tumor growth in xenograft model of human prostate cancer. Int. J. Oncol. 2005, 26, 1087–1091. [Google Scholar] [CrossRef]
- Oh, Y.K.; Park, T.G. siRNA delivery systems for cancer treatment. Adv. Drug Deliv. Rev. 2009, 61, 850–862. [Google Scholar] [CrossRef]
- Balwani, M.; Sardh, E.; Ventura, P.; Peiro, P.A.; Rees, D.C.; Stolzel, U.; Bissell, D.M.; Bonkovsky, H.L.; Windyga, J.; Anderson, K.E.; et al. Phase 3 Trial of RNAi Therapeutic Givosiran for Acute Intermittent Porphyria. N. Engl. J. Med. 2020, 382, 2289–2301. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.fda.gov/drugs/news-events-human-drugs/fda-approves-add-therapy-lower-cholesterol-among-certain-high-risk-adults (accessed on 26 September 2022).
- Available online: https://www.novartis.com/news/media-releases/fda-approves-novartis-leqvio-inclisiran-first-class-sirna-lower-cholesterol-and-keep-it-low-two-doses-year (accessed on 4 February 2013).
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [PubMed]
- Fenton, O.S.; Olafson, K.N.; Pillai, P.S.; Mitchell, M.J.; Langer, R. Advances in Biomaterials for Drug Delivery. Adv. Mater. 2018, 30, e1705328. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Santos, B.; Gremiao, M.P.; Chorilli, M. Nanotechnology-based drug delivery systems for the treatment of Alzheimer’s disease. Int. J. Nanomed. 2015, 10, 4981–5003. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef]
- Mukherjee, S.; Ray, S.; Thakur, R.S. Solid lipid nanoparticles: A modern formulation approach in drug delivery system. Indian J. Pharm. Sci. 2009, 71, 349–358. [Google Scholar] [CrossRef]
- Ghasemiyeh, P.; Mohammadi-Samani, S. Solid lipid nanoparticles and nanostructured lipid carriers as novel drug delivery systems: Applications, advantages and disadvantages. Res. Pharm. Sci. 2018, 13, 288–303. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Hajj, K.A.; Whitehead, K.A. Tools for translation: Non-viral materials for therapeutic mRNA delivery. Nat. Rev. Mater. 2017, 2, 17056. [Google Scholar] [CrossRef]
- Meng, C.; Chen, Z.; Li, G.; Welte, T.; Shen, H. Nanoplatforms for mRNA therapeutics. Adv. Ther. 2021, 4, 2000099. [Google Scholar] [CrossRef]
- Bailey, A.L.; Cullis, P.R. Modulation of membrane fusion by asymmetric transbilayer distributions of amino lipids. Biochemistry 1994, 33, 12573–12580. [Google Scholar] [CrossRef] [PubMed]
- Schlich, M.; Palomba, R.; Costabile, G.; Mizrahy, S.; Pannuzzo, M.; Peer, D.; Decuzzi, P. Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles. Bioeng. Transl. Med. 2021, 6, e10213. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Oberli, M.A.; Reichmuth, A.M.; Dorkin, J.R.; Mitchell, M.J.; Fenton, O.S.; Jaklenec, A.; Anderson, D.G.; Langer, R.; Blankschtein, D. Lipid Nanoparticle Assisted mRNA Delivery for Potent Cancer Immunotherapy. Nano Lett. 2017, 17, 1326–1335. [Google Scholar] [CrossRef]
- Liang, F.; Lindgren, G.; Lin, A.; Thompson, E.A.; Ols, S.; Rohss, J.; John, S.; Hassett, K.; Yuzhakov, O.; Bahl, K.; et al. Efficient Targeting and Activation of Antigen-Presenting Cells In Vivo after Modified mRNA Vaccine Administration in Rhesus Macaques. Mol. Ther. 2017, 25, 2635–2647. [Google Scholar] [CrossRef]
- Patel, S.; Ashwanikumar, N.; Robinson, E.; DuRoss, A.; Sun, C.; Murphy-Benenato, K.E.; Mihai, C.; Almarsson, O.; Sahay, G. Boosting Intracellular Delivery of Lipid Nanoparticle-Encapsulated mRNA. Nano Lett. 2017, 17, 5711–5718. [Google Scholar] [CrossRef]
- Leung, A.K.; Tam, Y.Y.; Cullis, P.R. Lipid nanoparticles for short interfering RNA delivery. Adv. Genet. 2014, 88, 71–110. [Google Scholar] [CrossRef]
- Jayaraman, M.; Ansell, S.M.; Mui, B.L.; Tam, Y.K.; Chen, J.; Du, X.; Butler, D.; Eltepu, L.; Matsuda, S.; Narayanannair, J.K.; et al. Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew. Chem. Int. Ed. Engl. 2012, 51, 8529–8533. [Google Scholar] [CrossRef]
- Zimmermann, T.S.; Lee, A.C.; Akinc, A.; Bramlage, B.; Bumcrot, D.; Fedoruk, M.N.; Harborth, J.; Heyes, J.A.; Jeffs, L.B.; John, M.; et al. RNAi-mediated gene silencing in non-human primates. Nature 2006, 441, 111–114. [Google Scholar] [CrossRef]
- Semple, S.C.; Klimuk, S.K.; Harasym, T.O.; Dos Santos, N.; Ansell, S.M.; Wong, K.F.; Maurer, N.; Stark, H.; Cullis, P.R.; Hope, M.J.; et al. Efficient encapsulation of antisense oligonucleotides in lipid vesicles using ionizable aminolipids: Formation of novel small multilamellar vesicle structures. Biochim. Biophys. Acta 2001, 1510, 152–166. [Google Scholar] [CrossRef]
- Maurer, N.; Wong, K.F.; Stark, H.; Louie, L.; McIntosh, D.; Wong, T.; Scherrer, P.; Semple, S.C.; Cullis, P.R. Spontaneous entrapment of polynucleotides upon electrostatic interaction with ethanol-destabilized cationic liposomes. Biophys. J. 2001, 80, 2310–2326. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticle Technology for Clinical Translation of siRNA Therapeutics. Acc. Chem. Res. 2019, 52, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Heyes, J.; Palmer, L.; Bremner, K.; MacLachlan, I. Cationic lipid saturation influences intracellular delivery of encapsulated nucleic acids. J. Control. Release 2005, 107, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Semple, S.C.; Akinc, A.; Chen, J.; Sandhu, A.P.; Mui, B.L.; Cho, C.K.; Sah, D.W.; Stebbing, D.; Crosley, E.J.; Yaworski, E.; et al. Rational design of cationic lipids for siRNA delivery. Nat. Biotechnol. 2010, 28, 172–176. [Google Scholar] [CrossRef]
- Coelho, T.; Adams, D.; Silva, A.; Lozeron, P.; Hawkins, P.N.; Mant, T.; Perez, J.; Chiesa, J.; Warrington, S.; Tranter, E.; et al. Safety and efficacy of RNAi therapy for transthyretin amyloidosis. N. Engl. J. Med. 2013, 369, 819–829. [Google Scholar] [CrossRef]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat. Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef]
- Zhi, D.; Bai, Y.; Yang, J.; Cui, S.; Zhao, Y.; Chen, H.; Zhang, S. A review on cationic lipids with different linkers for gene delivery. Adv. Colloid Interface Sci. 2018, 253, 117–140. [Google Scholar] [CrossRef]
- Cullis, P.R.; Hope, M.J. Lipid Nanoparticle Systems for Enabling Gene Therapies. Mol. Ther. 2017, 25, 1467–1475. [Google Scholar] [CrossRef]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-assembled mRNA vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112. [Google Scholar] [CrossRef]
- Knop, K.; Hoogenboom, R.; Fischer, D.; Schubert, U.S. Poly(ethylene glycol) in drug delivery: Pros and cons as well as potential alternatives. Angew. Chem. Int. Ed. Engl. 2010, 49, 6288–6308. [Google Scholar] [CrossRef]
- Akinc, A.; Goldberg, M.; Qin, J.; Dorkin, J.R.; Gamba-Vitalo, C.; Maier, M.; Jayaprakash, K.N.; Jayaraman, M.; Rajeev, K.G.; Manoharan, M.; et al. Development of lipidoid-siRNA formulations for systemic delivery to the liver. Mol. Ther. 2009, 17, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tao, W.; Liu, D.; Wu, J.; Guo, Z.; Ji, X.; Bharwani, Z.; Zhao, L.; Zhao, X.; Farokhzad, O.C.; et al. Surface De-PEGylation Controls Nanoparticle-Mediated siRNA Delivery In Vitro and In Vivo. Theranostics 2017, 7, 1990–2002. [Google Scholar] [CrossRef] [PubMed]
- Heyes, J.; Hall, K.; Tailor, V.; Lenz, R.; MacLachlan, I. Synthesis and characterization of novel poly(ethylene glycol)-lipid conjugates suitable for use in drug delivery. J. Control. Release 2006, 112, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Ambegia, E.; Ansell, S.; Cullis, P.; Heyes, J.; Palmer, L.; MacLachlan, I. Stabilized plasmid-lipid particles containing PEG-diacylglycerols exhibit extended circulation lifetimes and tumor selective gene expression. Biochim. Biophys. Acta 2005, 1669, 155–163. [Google Scholar] [CrossRef]
- Belliveau, N.M.; Huft, J.; Lin, P.J.; Chen, S.; Leung, A.K.; Leaver, T.J.; Wild, A.W.; Lee, J.B.; Taylor, R.J.; Tam, Y.K.; et al. Microfluidic Synthesis of Highly Potent Limit-size Lipid Nanoparticles for In Vivo Delivery of siRNA. Mol. Ther. Nucleic Acids 2012, 1, e37. [Google Scholar] [CrossRef]
- Mui, B.L.; Tam, Y.K.; Jayaraman, M.; Ansell, S.M.; Du, X.; Tam, Y.Y.; Lin, P.J.; Chen, S.; Narayanannair, J.K.; Rajeev, K.G.; et al. Influence of Polyethylene Glycol Lipid Desorption Rates on Pharmacokinetics and Pharmacodynamics of siRNA Lipid Nanoparticles. Mol. Ther. Nucleic Acids 2013, 2, e139. [Google Scholar] [CrossRef]
- Parr, M.J.; Ansell, S.M.; Choi, L.S.; Cullis, P.R. Factors influencing the retention and chemical stability of poly(ethylene glycol)-lipid conjugates incorporated into large unilamellar vesicles. Biochim. Biophys. Acta 1994, 1195, 21–30. [Google Scholar] [CrossRef]
- Chen, S.; Tam, Y.Y.C.; Lin, P.J.C.; Sung, M.M.H.; Tam, Y.K.; Cullis, P.R. Influence of particle size on the in vivo potency of lipid nanoparticle formulations of siRNA. J. Control. Release 2016, 235, 236–244. [Google Scholar] [CrossRef]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef]
- Riley, R.S.; Kashyap, M.V.; Billingsley, M.M.; White, B.; Alameh, M.G.; Bose, S.K.; Zoltick, P.W.; Li, H.; Zhang, R.; Cheng, A.Y.; et al. Ionizable lipid nanoparticles for in utero mRNA delivery. Sci. Adv. 2021, 7, eaba1028. [Google Scholar] [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles-From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef]
- Daraee, H.; Etemadi, A.; Kouhi, M.; Alimirzalu, S.; Akbarzadeh, A. Application of liposomes in medicine and drug delivery. Artif. Cells Nanomed. Biotechnol. 2016, 44, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef]
- Nakhaei, P.; Margiana, R.; Bokov, D.O.; Abdelbasset, W.K.; Jadidi Kouhbanani, M.A.; Varma, R.S.; Marofi, F.; Jarahian, M.; Beheshtkhoo, N. Liposomes: Structure, Biomedical Applications, and Stability Parameters with Emphasis on Cholesterol. Front. Bioeng. Biotechnol. 2021, 9, 705886. [Google Scholar] [CrossRef] [PubMed]
- Çağdaş, M.; Sezer, A.D.; Bucak, S. Liposomes as potential drug carrier systems for drug delivery. Appl. Nanotechnol. Drug Deliv. 2014, 1, 1–50. [Google Scholar]
- Nisini, R.; Poerio, N.; Mariotti, S.; De Santis, F.; Fraziano, M. The Multirole of Liposomes in Therapy and Prevention of Infectious Diseases. Front. Immunol. 2018, 9, 155. [Google Scholar] [CrossRef]
- Lombardo, D.; Kiselev, M.A.; Caccamo, M.T. Smart nanoparticles for drug delivery application: Development of versatile nanocarrier platforms in biotechnology and nanomedicine. J. Nanomater. 2019, 2019, e3702518. [Google Scholar] [CrossRef]
- Shafiei, M.; Ansari, M.N.M.; Razak, S.I.A.; Khan, M.U.A. A Comprehensive Review on the Applications of Exosomes and Liposomes in Regenerative Medicine and Tissue Engineering. Polymers 2021, 13, 2529. [Google Scholar] [CrossRef]
- Monteiro, N.; Martins, A.; Reis, R.L.; Neves, N.M. Liposomes in tissue engineering and regenerative medicine. J. R. Soc. Interface 2014, 11, 20140459. [Google Scholar] [CrossRef]
- Xu, Q.; Tanaka, Y.; Czernuszka, J.T. Encapsulation and release of a hydrophobic drug from hydroxyapatite coated liposomes. Biomaterials 2007, 28, 2687–2694. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.A.; Reynolds, L.; Deitcher, S.R. Pharmacokinetics and pharmacodynamics of vincristine sulfate liposome injection (VSLI) in adults with acute lymphoblastic leukemia. J. Clin. Pharmacol. 2013, 53, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Tenchov, B.G.; MacDonald, R.C.; Siegel, D.P. Cubic phases in phosphatidylcholine-cholesterol mixtures: Cholesterol as membrane “fusogen”. Biophys. J. 2006, 91, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Siegel, D. The relationship between bicontinuous inverted cubic phases and membrane fusion. Surfactant Sci. Ser. 2005, 127, 59. [Google Scholar]
- Ashwell, G.; Harford, J. Carbohydrate-specific receptors of the liver. Annu. Rev. Biochem. 1982, 51, 531–554. [Google Scholar] [CrossRef] [PubMed]
- Grewal, P.K. The Ashwell-Morell receptor. Methods Enzym. 2010, 479, 223–241. [Google Scholar] [CrossRef]
- Grozovsky, R.; Begonja, A.J.; Liu, K.; Visner, G.; Hartwig, J.H.; Falet, H.; Hoffmeister, K.M. The Ashwell-Morell receptor regulates hepatic thrombopoietin production via JAK2-STAT3 signaling. Nat. Med. 2015, 21, 47–54. [Google Scholar] [CrossRef]
- Rigopoulou, E.I.; Roggenbuck, D.; Smyk, D.S.; Liaskos, C.; Mytilinaiou, M.G.; Feist, E.; Conrad, K.; Bogdanos, D.P. Asialoglycoprotein receptor (ASGPR) as target autoantigen in liver autoimmunity: Lost and found. Autoimmun. Rev. 2012, 12, 260–269. [Google Scholar] [CrossRef]
- Morell, A.G.; Gregoriadis, G.; Scheinberg, I.H.; Hickman, J.; Ashwell, G. The role of sialic acid in determining the survival of glycoproteins in the circulation. J. Biol. Chem. 1971, 246, 1461–1467. [Google Scholar] [CrossRef]
- Taylor, M.E.; Drickamer, K.; Imberty, A.; van Kooyk, Y.; Schnaar, R.L.; Etzler, M.E.; Varki, A. Discovery and Classification of Glycan-Binding Proteins. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Mohnen, D., Kinoshita, T., Packer, N.H., Eds.; Cold Spring Harbor: Woodbury, NY, USA, 2022; pp. 375–386. [Google Scholar]
- Wu, J.; Nantz, M.H.; Zern, M.A. Targeting hepatocytes for drug and gene delivery: Emerging novel approaches and applications. Front. Biosci. 2002, 7, d717–d725. [Google Scholar] [CrossRef]
- Lee, Y.C. Biochemistry of carbohydrate-protein interaction. FASEB J. 1992, 6, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Spiess, M. The asialoglycoprotein receptor: A model for endocytic transport receptors. Biochemistry 1990, 29, 10009–10018. [Google Scholar] [CrossRef] [PubMed]
- Stockert, R.J. The asialoglycoprotein receptor: Relationships between structure, function, and expression. Physiol. Rev. 1995, 75, 591–609. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Michalak, T.I. Composition, antigenic properties and hepatocyte surface expression of the woodchuck asialoglycoprotein receptor. J. Recept. Signal Transduct. 1996, 16, 243–271. [Google Scholar] [CrossRef] [PubMed]
- Morell, A.G.; Scheinberg, I.H. Solubilization of hepatic binding sites for asialo-glycoproteins. Biochem. Biophys. Res. Commun. 1972, 48, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Kuhlenschmidt, M.; Roseman, S.; Lee, Y.C. Synthesis of some cluster galactosides and their effect on the hepatic galactose-binding system. Arch. Biochem. Biophys. 1980, 205, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Bider, M.D.; Malashkevich, V.N.; Spiess, M.; Burkhard, P. Crystal structure of the carbohydrate recognition domain of the H1 subunit of the asialoglycoprotein receptor. J. Mol. Biol. 2000, 300, 857–865. [Google Scholar] [CrossRef]
- D’Souza, A.A.; Devarajan, P.V. Asialoglycoprotein receptor mediated hepatocyte targeting-strategies and applications. J. Control. Release 2015, 203, 126–139. [Google Scholar] [CrossRef]
- Roggenbuck, D.; Mytilinaiou, M.G.; Lapin, S.V.; Reinhold, D.; Conrad, K. Asialoglycoprotein receptor (ASGPR): A peculiar target of liver-specific autoimmunity. Autoimmun. Highlights 2012, 3, 119–125. [Google Scholar] [CrossRef]
- Baenziger, J.U.; Fiete, D. Galactose and N-acetylgalactosamine-specific endocytosis of glycopeptides by isolated rat hepatocytes. Cell 1980, 22, 611–620. [Google Scholar] [CrossRef]
- Biessen, E.A.; Sliedregt-Bol, K.; PA, T.H.; Prince, P.; Van der Bilt, E.; Valentijn, A.R.; Meeuwenoord, N.J.; Princen, H.; Bijsterbosch, M.K.; Van der Marel, G.A.; et al. Design of a targeted peptide nucleic acid prodrug to inhibit hepatic human microsomal triglyceride transfer protein expression in hepatocytes. Bioconjugate Chem. 2002, 13, 295–302. [Google Scholar] [CrossRef]
- Rensen, P.C.; van Leeuwen, S.H.; Sliedregt, L.A.; van Berkel, T.J.; Biessen, E.A. Design and synthesis of novel N-acetylgalactosamine-terminated glycolipids for targeting of lipoproteins to the hepatic asialoglycoprotein receptor. J. Med. Chem. 2004, 47, 5798–5808. [Google Scholar] [CrossRef]
- Seymour, L.W.; Ferry, D.R.; Anderson, D.; Hesslewood, S.; Julyan, P.J.; Poyner, R.; Doran, J.; Young, A.M.; Burtles, S.; Kerr, D.J.; et al. Hepatic drug targeting: Phase I evaluation of polymer-bound doxorubicin. J. Clin. Oncol. 2002, 20, 1668–1676. [Google Scholar] [CrossRef]
- Fiume, L.; Mattioli, A.; Balboni, P.G.; Tognon, M.; Barbanti-Brodano, G.; de Vries, J.; Wieland, T. Enhanced inhibition of virus DNA synthesis in hepatocytes by trifluorothymidine coupled to asialofetuin. FEBS Lett. 1979, 103, 47–51. [Google Scholar] [CrossRef]
- Fiume, L.; Mattioli, A.; Busi, C.; Balboni, P.G.; Barbanti-Brodano, G.; De Vries, J.; Altmann, R.; Wieland, T. Selective inhibition of Ectromelia virus DNA synthesis in hepatocytes by adenine-9-beta-D-arabinofuranoside (ara-A) and adenine-9-beta-D-arabinofuranoside 5’-monophosphate (ara-AMP) conjugated to asialofetuin. FEBS Lett. 1980, 116, 185–188. [Google Scholar] [CrossRef]
- Wu, G.Y.; Wu, C.H. Receptor-mediated in vitro gene transformation by a soluble DNA carrier system. J. Biol. Chem. 1987, 262, 4429–4432. [Google Scholar] [CrossRef] [PubMed]
- Plank, C.; Zatloukal, K.; Cotten, M.; Mechtler, K.; Wagner, E. Gene transfer into hepatocytes using asialoglycoprotein receptor mediated endocytosis of DNA complexed with an artificial tetra-antennary galactose ligand. Bioconjugate Chem. 1992, 3, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Hangeland, J.J.; Levis, J.T.; Lee, Y.C.; Ts’o, P.O. Cell-type specific and ligand specific enhancement of cellular uptake of oligodeoxynucleoside methylphosphonates covalently linked with a neoglycopeptide, YEE(ah-GalNAc)3. Bioconjugate Chem. 1995, 6, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Prakash, T.P.; Graham, M.J.; Yu, J.; Carty, R.; Low, A.; Chappell, A.; Schmidt, K.; Zhao, C.; Aghajan, M.; Murray, H.F.; et al. Targeted delivery of antisense oligonucleotides to hepatocytes using triantennary N-acetyl galactosamine improves potency 10-fold in mice. Nucleic Acids Res. 2014, 42, 8796–8807. [Google Scholar] [CrossRef]
- Debacker, A.J.; Voutila, J.; Catley, M.; Blakey, D.; Habib, N. Delivery of Oligonucleotides to the Liver with GalNAc: From Research to Registered Therapeutic Drug. Mol. Ther. 2020, 28, 1759–1771. [Google Scholar] [CrossRef]
- Shen, X.; Corey, D.R. Chemistry, mechanism and clinical status of antisense oligonucleotides and duplex RNAs. Nucleic Acids Res. 2018, 46, 1584–1600. [Google Scholar] [CrossRef]
- Nair, J.K.; Willoughby, J.L.; Chan, A.; Charisse, K.; Alam, M.R.; Wang, Q.; Hoekstra, M.; Kandasamy, P.; Kel’in, A.V.; Milstein, S.; et al. Multivalent N-acetylgalactosamine-conjugated siRNA localizes in hepatocytes and elicits robust RNAi-mediated gene silencing. J. Am. Chem. Soc. 2014, 136, 16958–16961. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.F.; Swayze, E.E. RNA targeting therapeutics: Molecular mechanisms of antisense oligonucleotides as a therapeutic platform. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Dhuri, K.; Bechtold, C.; Quijano, E.; Pham, H.; Gupta, A.; Vikram, A.; Bahal, R. Antisense Oligonucleotides: An Emerging Area in Drug Discovery and Development. J. Clin. Med. 2020, 9, 2004. [Google Scholar] [CrossRef]
- Prakash, T.P.; Yu, J.; Migawa, M.T.; Kinberger, G.A.; Wan, W.B.; Østergaard, M.E.; Carty, R.L.; Vasquez, G.; Low, A.; Chappell, A.; et al. Comprehensive Structure-Activity Relationship of Triantennary N-Acetylgalactosamine Conjugated Antisense Oligonucleotides for Targeted Delivery to Hepatocytes. J. Med. Chem. 2016, 59, 2718–2733. [Google Scholar] [CrossRef]
- Castellanos-Rizaldos, E.; Brown, C.R.; Dennin, S.; Kim, J.; Gupta, S.; Najarian, D.; Gu, Y.; Aluri, K.; Enders, J.; Brown, K. Reverse transcription quantitative polymerase chain reaction methods to support pharmacokinetics and drug mechanism of action to advance development of RNA interference therapeutics. Nucleic Acid Ther. 2020, 30, 133–142. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, M.; Hu, B.; Huang, Y. siRNA Design and GalNAc-Empowered Hepatic Targeted Delivery. Methods Mol. Biol. 2021, 2282, 77–100. [Google Scholar] [CrossRef]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L.J.; et al. Two Phase 3 Trials of Inclisiran in Patients with Elevated LDL Cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef]
- Fitzgerald, K.; White, S.; Borodovsky, A.; Bettencourt, B.R.; Strahs, A.; Clausen, V.; Wijngaard, P.; Horton, J.D.; Taubel, J.; Brooks, A.; et al. A Highly Durable RNAi Therapeutic Inhibitor of PCSK9. N. Engl. J. Med. 2017, 376, 41–51. [Google Scholar] [CrossRef]
- Kinberger, G.A.; Prakash, T.P.; Yu, J.; Vasquez, G.; Low, A.; Chappell, A.; Schmidt, K.; Murray, H.M.; Gaus, H.; Swayze, E.E.; et al. Conjugation of mono and di-GalNAc sugars enhances the potency of antisense oligonucleotides via ASGR mediated delivery to hepatocytes. Bioorganic Med. Chem. Lett. 2016, 26, 3690–3693. [Google Scholar] [CrossRef]
- Zhou, Y.; Teng, P.; Montgomery, N.T.; Li, X.; Tang, W. Development of Triantennary N-Acetylgalactosamine Conjugates as Degraders for Extracellular Proteins. ACS Cent. Sci. 2021, 7, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Sardh, E.; Harper, P.; Balwani, M.; Stein, P.; Rees, D.; Bissell, D.M.; Desnick, R.; Parker, C.; Phillips, J.; Bonkovsky, H.L.; et al. Phase 1 Trial of an RNA Interference Therapy for Acute Intermittent Porphyria. N. Engl. J. Med. 2019, 380, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.gov.uk/government/news/uk-government-tackles-heart-disease-with-new-partnership (accessed on 29 September 2022).
- Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 15 August 2022).
- Investigators, A.-H.; Boden, W.E.; Probstfield, J.L.; Anderson, T.; Chaitman, B.R.; Desvignes-Nickens, P.; Koprowicz, K.; McBride, R.; Teo, K.; Weintraub, W. Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N. Engl. J. Med. 2011, 365, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Fazio, S.; Guyton, J.R.; Polis, A.B.; Adewale, A.J.; Tomassini, J.E.; Ryan, N.W.; Tershakovec, A.M. Long-term safety and efficacy of triple combination ezetimibe/simvastatin plus extended-release niacin in patients with hyperlipidemia. Am. J. Cardiol. 2010, 105, 487–494. [Google Scholar] [CrossRef]
- Bays, H. Safety of niacin and simvastatin combination therapy. Am. J. Cardiol. 2008, 101, 3B–8B. [Google Scholar] [CrossRef]
- Staels, B.; Dallongeville, J.; Auwerx, J.; Schoonjans, K.; Leitersdorf, E.; Fruchart, J.C. Mechanism of action of fibrates on lipid and lipoprotein metabolism. Circulation 1998, 98, 2088–2093. [Google Scholar] [CrossRef]
- Insua, A.; Massari, F.; Rodriguez Moncalvo, J.J.; Ruben Zanchetta, J.; Insua, A.M. Fenofibrate of gemfibrozil for treatment of types IIa and IIb primary hyperlipoproteinemia: A randomized, double-blind, crossover study. Endocr. Pract. 2002, 8, 96–101. [Google Scholar] [CrossRef]
- Aldridge, M.A.; Ito, M.K. Colesevelam hydrochloride: A novel bile acid-binding resin. Ann. Pharmacother. 2001, 35, 898–907. [Google Scholar] [CrossRef]
- Heel, R.C.; Brogden, R.N.; Pakes, G.E.; Speight, T.M.; Avery, G.S. Colestipol: A review of its pharmacological properties and therapeutic efficacy in patients with hypercholesterolaemia. Drugs 1980, 19, 161–180. [Google Scholar] [CrossRef]
- Insull, W., Jr. Clinical utility of bile acid sequestrants in the treatment of dyslipidemia: A scientific review. South. Med. J. 2006, 99, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. Cholesterol Lowering Drugs. [Updated 2021 Mar 30]. In Endotext [Internet]; Feingold, K.R., Boyce, A.B.A., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK395573/ (accessed on 29 September 2022).
- Chung, B.H.Y.; Chau, J.F.T.; Wong, G.K. Rare versus common diseases: A false dichotomy in precision medicine. NPJ Genom. Med. 2021, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Garg, J.; Shah, N.; Sumner, A. PCSK9 inhibitors: A new era of lipid lowering therapy. World J. Cardiol. 2017, 9, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Colon, E.; Daum, A.; Yosefy, C. Statins and PCSK9 inhibitors: A new lipid-lowering therapy. Eur. J. Pharmacol. 2020, 878, 173114. [Google Scholar] [CrossRef]
- Olsson, A. PCSK9 inhibition—A new era in cholesterol treatment. Lakartidningen 2015, 112, DAAF. [Google Scholar]
- Cicero, A.F.; Fogacci, F.; Zambon, A.; Toth, P.P.; Borghi, C. Efficacy and safety of inclisiran a newly approved FDA drug: A systematic review and pooled analysis of available clinical studies. Am. Heart J. Plus Cardiol. Res. Pract. 2022, 13, 100127. [Google Scholar] [CrossRef]
- Abifadel, M.; Varret, M.; Rabes, J.P.; Allard, D.; Ouguerram, K.; Devillers, M.; Cruaud, C.; Benjannet, S.; Wickham, L.; Erlich, D.; et al. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat. Genet. 2003, 34, 154–156. [Google Scholar] [CrossRef]
- Allard, D.; Amsellem, S.; Abifadel, M.; Trillard, M.; Devillers, M.; Luc, G.; Krempf, M.; Reznik, Y.; Girardet, J.P.; Fredenrich, A.; et al. Novel mutations of the PCSK9 gene cause variable phenotype of autosomal dominant hypercholesterolemia. Hum. Mutat. 2005, 26, 497. [Google Scholar] [CrossRef]
- Santos, R.D.; Rocha, V.Z. Cholesterol Lowering with Inclisiran: A New Chapter in the PCSK9 Story Book; Oxford University Press US: Oxford, UK, 2023; Volume 44, pp. 139–141. [Google Scholar]
- Ruscica, M.; Ferri, N.; Macchi, C.; Meroni, M.; Lanti, C.; Ricci, C.; Maggioni, M.; Fracanzani, A.L.; Badiali, S.; Fargion, S.; et al. Liver fat accumulation is associated with circulating PCSK9. Ann. Med. 2016, 48, 384–391. [Google Scholar] [CrossRef]
- Guo, Y.L.; Zhang, W.; Li, J.J. PCSK9 and lipid lowering drugs. Clin. Chim. Acta 2014, 437, 66–71. [Google Scholar] [CrossRef]
- Handelsman, Y.; Lepor, N.E. PCSK9 Inhibitors in Lipid Management of Patients With Diabetes Mellitus and High Cardiovascular Risk: A Review. J. Am. Heart Assoc. 2018, 7, e008953. [Google Scholar] [CrossRef] [PubMed]
- McKenney, J.M.; Koren, M.J.; Kereiakes, D.J.; Hanotin, C.; Ferrand, A.C.; Stein, E.A. Safety and efficacy of a monoclonal antibody to proprotein convertase subtilisin/kexin type 9 serine protease, SAR236553/REGN727, in patients with primary hypercholesterolemia receiving ongoing stable atorvastatin therapy. J. Am. Coll. Cardiol. 2012, 59, 2344–2353. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.M.; McKenney, J.M.; Hanotin, C.; Asset, G.; Stein, E.A. Atorvastatin with or without an antibody to PCSK9 in primary hypercholesterolemia. N. Engl. J. Med. 2012, 367, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.A.; Gipe, D.; Bergeron, J.; Gaudet, D.; Weiss, R.; Dufour, R.; Wu, R.; Pordy, R. Effect of a monoclonal antibody to PCSK9, REGN727/SAR236553, to reduce low-density lipoprotein cholesterol in patients with heterozygous familial hypercholesterolaemia on stable statin dose with or without ezetimibe therapy: A phase 2 randomised controlled trial. Lancet 2012, 380, 29–36. [Google Scholar] [CrossRef]
- Stroes, E.; Colquhoun, D.; Sullivan, D.; Civeira, F.; Rosenson, R.S.; Watts, G.F.; Bruckert, E.; Cho, L.; Dent, R.; Knusel, B.; et al. Anti-PCSK9 antibody effectively lowers cholesterol in patients with statin intolerance: The GAUSS-2 randomized, placebo-controlled phase 3 clinical trial of evolocumab. J. Am. Coll. Cardiol. 2014, 63, 2541–2548. [Google Scholar] [CrossRef]
- Nissen, S.E.; Stroes, E.; Dent-Acosta, R.E.; Rosenson, R.S.; Lehman, S.J.; Sattar, N.; Preiss, D.; Bruckert, E.; Ceska, R.; Lepor, N.; et al. Efficacy and Tolerability of Evolocumab vs Ezetimibe in Patients with Muscle-Related Statin Intolerance: The GAUSS-3 Randomized Clinical Trial. JAMA 2016, 315, 1580–1590. [Google Scholar] [CrossRef]
- Roth, E.M.; Taskinen, M.R.; Ginsberg, H.N.; Kastelein, J.J.; Colhoun, H.M.; Robinson, J.G.; Merlet, L.; Pordy, R.; Baccara-Dinet, M.T. Monotherapy with the PCSK9 inhibitor alirocumab versus ezetimibe in patients with hypercholesterolemia: Results of a 24 week, double-blind, randomized Phase 3 trial. Int. J. Cardiol. 2014, 176, 55–61. [Google Scholar] [CrossRef]
- Roth, E.M.; Diller, P. Alirocumab for hyperlipidemia: Physiology of PCSK9 inhibition, pharmacodynamics and Phase I and II clinical trial results of a PCSK9 monoclonal antibody. Future Cardiol. 2014, 10, 183–199. [Google Scholar] [CrossRef]
- Matzno, S.; Tazuya-Murayama, K.; Tanaka, H.; Yasuda, S.; Mishima, M.; Uchida, T.; Nakabayashi, T.; Matsuyama, K. Evaluation of the synergistic adverse effects of concomitant therapy with statins and fibrates on rhabdomyolysis. J. Pharm. Pharmacol. 2003, 55, 795–802. [Google Scholar] [CrossRef]
- Banerjee, Y.; Pantea Stoian, A.; Cicero, A.F.G.; Fogacci, F.; Nikolic, D.; Sachinidis, A.; Rizvi, A.A.; Janez, A.; Rizzo, M. Inclisiran: A small interfering RNA strategy targeting PCSK9 to treat hypercholesterolemia. Expert Opin. Drug Saf. 2022, 21, 9–20. [Google Scholar] [CrossRef]
- Brar, K.S. Ezetimibe (Zetia). Med. J. Armed Forces India 2004, 60, 388–389. [Google Scholar] [CrossRef] [PubMed]
- Colletti, R.B.; Neufeld, E.J.; Roff, N.K.; McAuliffe, T.L.; Baker, A.L.; Newburger, J.W. Niacin treatment of hypercholesterolemia in children. Pediatrics 1993, 92, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.ema.europa.eu/en/documents/assessment-report/leqvio-epar-public-assessment-report_en.pdf (accessed on 29 September 2022).
- Tsui, N.B.; Ng, E.K.; Lo, Y.M. Stability of endogenous and added RNA in blood specimens, serum, and plasma. Clin. Chem. 2002, 48, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Czauderna, F.; Fechtner, M.; Dames, S.; Aygun, H.; Klippel, A.; Pronk, G.J.; Giese, K.; Kaufmann, J. Structural variations and stabilising modifications of synthetic siRNAs in mammalian cells. Nucleic Acids Res. 2003, 31, 2705–2716. [Google Scholar] [CrossRef]
- Choung, S.; Kim, Y.J.; Kim, S.; Park, H.O.; Choi, Y.C. Chemical modification of siRNAs to improve serum stability without loss of efficacy. Biochem. Biophys. Res. Commun. 2006, 342, 919–927. [Google Scholar] [CrossRef]
- Khvorova, A. Oligonucleotide Therapeutics-A New Class of Cholesterol-Lowering Drugs. N. Engl. J. Med. 2017, 376, 4–7. [Google Scholar] [CrossRef]
- Scicchitano, P.; Milo, M.; Mallamaci, R.; De Palo, M.; Caldarola, P.; Massari, F.; Gabrielli, D.; Colivicchi, F.; Ciccone, M.M. Inclisiran in lipid management: A Literature overview and future perspectives. Biomed. Pharmacother. 2021, 143, 112227. [Google Scholar] [CrossRef]
- Frank-Kamenetsky, M.; Grefhorst, A.; Anderson, N.N.; Racie, T.S.; Bramlage, B.; Akinc, A.; Butler, D.; Charisse, K.; Dorkin, R.; Fan, Y.; et al. Therapeutic RNAi targeting PCSK9 acutely lowers plasma cholesterol in rodents and LDL cholesterol in nonhuman primates. Proc. Natl. Acad. Sci. USA 2008, 105, 11915–11920. [Google Scholar] [CrossRef]
- Fitzgerald, K.; Frank-Kamenetsky, M.; Shulga-Morskaya, S.; Liebow, A.; Bettencourt, B.R.; Sutherland, J.E.; Hutabarat, R.M.; Clausen, V.A.; Karsten, V.; Cehelsky, J.; et al. Effect of an RNA interference drug on the synthesis of proprotein convertase subtilisin/kexin type 9 (PCSK9) and the concentration of serum LDL cholesterol in healthy volunteers: A randomised, single-blind, placebo-controlled, phase 1 trial. Lancet 2014, 383, 60–68. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/ct2/results?cond=&term=inclisiran&cntry=&state=&city=&dist= (accessed on 29 September 2022).
- Ray, K.K.; Landmesser, U.; Leiter, L.A.; Kallend, D.; Dufour, R.; Karakas, M.; Hall, T.; Troquay, R.P.; Turner, T.; Visseren, F.L.; et al. Inclisiran in Patients at High Cardiovascular Risk with Elevated LDL Cholesterol. N. Engl. J. Med. 2017, 376, 1430–1440. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Munoz Estrella, A.; Skavdis, A.; Pena Genao, E.; Martinez, I.; Guzman, E. Inclisiran for the Treatment of Cardiovascular Disease: A Short Review on the Emerging Data and Therapeutic Potential. Ther. Clin. Risk Manag. 2020, 16, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Raal, F.J.; Kallend, D.; Ray, K.K.; Turner, T.; Koenig, W.; Wright, R.S.; Wijngaard, P.L.J.; Curcio, D.; Jaros, M.J.; Leiter, L.A.; et al. Inclisiran for the Treatment of Heterozygous Familial Hypercholesterolemia. N. Engl. J. Med. 2020, 382, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.K.; Kallend, D.; Leiter, L.A.; Raal, F.J.; Koenig, W.; Jaros, M.J.; Schwartz, G.G.; Landmesser, U.; Garcia Conde, L.; Wright, R.S. Effect of inclisiran on lipids in primary prevention: The ORION-11 trial. Eur. Heart J. 2022, 43, 5047–5057. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, J.; Lodish, H.F. Two asialoglycoprotein receptor polypeptides in human hepatoma cells. J. Biol. Chem. 1987, 262, 11825–11832. [Google Scholar] [CrossRef] [PubMed]
- Witzigmann, D.; Quagliata, L.; Schenk, S.H.; Quintavalle, C.; Terracciano, L.M.; Huwyler, J. Variable asialoglycoprotein receptor 1 expression in liver disease: Implications for therapeutic intervention. Hepatol. Res. 2016, 46, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, J.L.S.; Chan, A.; Sehgal, A.; Butler, J.S.; Nair, J.K.; Racie, T.; Shulga-Morskaya, S.; Nguyen, T.; Qian, K.; Yucius, K.; et al. Evaluation of GalNAc-siRNA Conjugate Activity in Pre-clinical Animal Models with Reduced Asialoglycoprotein Receptor Expression. Mol. Ther. 2018, 26, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Cupido, A.J.; Kastelein, J.J.P. Inclisiran for the treatment of hypercholesterolaemia: Implications and unanswered questions from the ORION trials. Cardiovasc. Res. 2020, 116, e136–e139. [Google Scholar] [CrossRef]
- Dyrbuś, K.; Gąsior, M.; Penson, P.; Ray, K.K.; Banach, M. Inclisiran-New hope in the management of lipid disorders? J. Clin. Lipidol. 2020, 14, 16–27. [Google Scholar] [CrossRef]
- Hardy, J.; Niman, S.; Pereira, E.; Lewis, T.; Reid, J.; Choksi, R.; Goldfaden, R.F. A Critical Review of the Efficacy and Safety of Inclisiran. Am. J. Cardiovasc. Drugs 2021, 21, 629–642. [Google Scholar] [CrossRef]
- Pirillo, A.; Catapano, A.L. Inclisiran: How Widely and When Should We Use It? Curr. Atheroscler. Rep. 2022, 24, 803–811. [Google Scholar] [CrossRef]
- Damase, T.R.; Sukhovershin, R.; Boada, C.; Taraballi, F.; Pettigrew, R.I.; Cooke, J.P. The Limitless Future of RNA Therapeutics. Front. Bioeng. Biotechnol. 2021, 9, 628137. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. mRNA-based therapeutics: Powerful and versatile tools to combat diseases. Signal Transduct. Target. Ther. 2022, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.M.; Donner, A.J.; Blank, E.E.; Egger, A.W.; Kellar, B.M.; Østergaard, M.E.; Seth, P.P.; Harris, E.N. Stabilin-1 and Stabilin-2 are specific receptors for the cellular internalization of phosphorothioate-modified antisense oligonucleotides (ASOs) in the liver. Nucleic Acids Res. 2016, 44, 2782–2794. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Name | Delivery Platform/Targeting Ligand | Disease/Targeting Gene | Company | Updated Status |
|---|---|---|---|---|
| Onpattro (Patisiran) | LNP-siRNA | TTR-mediated amyloidosis (Transthyretin) | Alnylam | FDA. approval (10/10/2018) |
| Givlaari (Givosiran) | GalNAc-siRNA | Acute hepatic porphyria (delta-aminolevulinate synthase 1) | Alnylam | FDA approval (11/20/2019) |
| Oxlumo (Lumasiran) | GalNAc-siRNA | Primary hyperoxaluria type 1 (hydroxy acid oxidase 1) | Alnylam | FDA approval (11/23/2020) |
| Leqvio (Inclisiran) | GalNAc-siRNA | Hypercholesterolemia (proprotein convertase subtilisin/Kexin type 9) | Alnylam Novartis | FDA approval (12/22/2021) |
| Vutrisiran (ALN-TTRSC02) | GalNAc-siRNA | TTR-mediated amyloidosis (Transthyretin) | Alnylam | Phase III |
| Fitusiran (ALN-AT3SC) | GalNAc-siRNA | Haemophilia A and B and rare blood disorders (antithrombin) | Alnylam, Sanofi, Genzyme | Phase III |
| Nedosiran (DCR-PHXC) | GalNAc-siRNA | Primary hyperoxaluria (lactate dehydrogenase A) | Alnylam, Dicerna | Phase III |
| Teprasiran (QPI-1002) | None | Acute kidney injury (tumor protein) | Quark, Novartis | Phase III |
| Cosdosiran (QPI-1007) | None | NAION and glaucoma (Caspase 2) | Quark, | Phase III |
| Tivanisiran (SYL1001) | None | Ocular pain and dry eye disease (TRPV1) | Sylentis | Phase III |
| Chemical Structure of Ionizable Lipids | Name | Reference | |
|---|---|---|---|
 | DODAP | 1,2-dioleoyl-3-dimethylammonium propane | [53] |
 | DLinDMA | 1,2-dilinoleyloxy-N,N-dimethyl-3-aminopropane | [52,56] |
 | DLin-KC2-DMA | 2,2-dilinoleyl-4-dimethylaminoethyl-[1,3]-dioxolane | [57] |
 | DLin-MC3-DMA | (6Z,9Z,28Z,31Z)-heptatriaconta-6,9,28,31-tetraen-19-yl 4-(dimethylamino) butanoate | [51,58,59] |
| NCT Number/Other IDs | Study Title | Status/Study Type/Phases | Study Design | Start/ Completion Date | NCT Number/Other IDs | Study Title | Status/Study Type/Phases | Study Design | Start/ Completion Date |
|---|---|---|---|---|---|---|---|---|---|
| NCT04929249/ CKJX839A1US02 | Randomized study to evaluate the effect of an “inclisiran first” implementation strategy compared to usual care in patients with atherosclerotic cardiovascular disease and elevated LDL-C despite receiving maximally tolerated Statin Therapy (VICTORION-INITIATE) | * Recruiting/Interventional/Phase 3 | None (Open Label), Primary Purpose: Treatment | 25-Jun-2021 to 30-Jun-2023 | NCT03814187/ MDCO-PCS-17-05/CKJX839A12306B/2017-003092-55 | Trial to assess the effect of long-term dosing of inclisiran in subjects with High CV risk and elevated LDL-C (ORION 8) | ** Active, not recruiting/Interventional/Phase 3 | None (Open Label), Primary Purpose: Treatment | 16-Apr-2019 to 31-Dec-2023 |
| NCT05118230/ CKJX839A1CN01 | Study to assess the real-world effectiveness of inclisiran in Chinese adult patients with primary hypercholesterolemia or mixed dyslipidemia | * Recruiting/Observational/NA | Observational Model: Cohort, Time Perspective: Prospective | 9-Dec-2021 to 30-Sep-2024 | NCT05362903/ CKJX839A1DE01 | A non-interventional implementation study to evaluate treatment with inclisiran (Leqvio) and other lipid lowering treatments in a real-world setting | * Recruiting/Observational/NA | Observational Model: Cohort, Time Perspective: Prospective | 28-Jan-2023 to 31-Jan-2025 |
| NCT03159416/ MDCO-PCS-16-03 | A study of inclisiran in participants with renal impairment compared to participants with normal renal function (ORION-7) | *** Completed/Interventional/Phase 1 | None (Open Label), Primary | 22-Jun-2017 to 24-Mar-2018 | NCT04873934/ CKJX839A1US01 | Management of LDL-cholesterol with inclisiran + usual care compared to usual care alone in participants with a recent acute coronary syndrome | * Recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: | 24-Jun-2021 to 20-Mar-2024 |
| NCT03060577/ MDCO-PCS-16-01 | An extension trial of inclisiran compared to evolocumab in participants with cardiovascular disease and high cholesterol (ORION-3) | *** Completed/Interventional/Phase 2 | Allocation: Non-Randomized, Intervention Model: Parallel Assignment, Masking: None (Open Label), Primary Purpose: Treatment | 24-Mar-2017 to 17-Dec-2021 | NCT05399992/ CKJX839A12401 | Study evaluating effectiveness and adherence of inclisiran plus standard of care (SoC) lipid-lowering therapy compared to SoC in ASCVD | **** Not-recruiting/Observational/NA | Observational Model: Cohort, Time Perspective: Prospective | 30-Aug-2022 to 1-Apr-2022 |
| NCT04774003/ CKJX839A12105 | Study of pharmacokinetics, pharmacodynamics, safety, and tolerability of inclisiran in Chinese participants with elevated serum LDL-C (ORION-14) | *** Completed/Interventional/Phase 1 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Treatment | 26-Feb-2021 to 18-Oct-2021 | NCT04659863/ CKJX839C12302/2020-002755-38 | Study to evaluate efficacy and safety of inclisiran in adolescents with homozygous familial hypercholesterolemia (ORION-13) | * Recruiting/Interventional/Phase 3 | Allocation: Randomize, Intervention Model: Parallel Assignment, Masking: Double (Participant, Investigator), Primary Purpose: Treatment | 15-Feb-2021 to 9-Dec-2024 |
| NCT04652726/ CKJX839C12301/2020-002757-18 | Study to evaluate efficacy and safety of inclisiran in adolescents with heterozygous familial hypercholesterolemia (ORION-16) | * Recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Investigator), Primary Purpose: Treatment | 27-Jan-2021 to 9-Dec-2024 | NCT04807400/ CKJX839A1GB01/2020-004401-31 | Study in primary care evaluating inclisiran delivery implementation + enhanced support | ** Active, not recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: None (Open Label), Primary Purpose: Health Services Research | 7-Jul-2021 to 30-Jan-2023 |
| NCT05192941/ CKJX839A12402/2021-003759-40 | Study of efficacy, safety, tolerability, and quality of life of inclisiran (kjx839) vs. placebo, on top of on-going individually optimized lipid-lowering therapy, in participants with hypercholesterolemia | * Recruiting/Interventional/Phase 4 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Treatment | 8-Apr-2022 to 4-Feb-2025 | NCT04666298/ CKJX839A11201 | Study of efficacy and safety of inclisiran in Japanese participants with high cardiovascular risk and elevated LDL-C | ** Active, not recruiting/Interventional/Phase 1 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Treatment | 29-Jan-2021 to 16-Oct-2021 |
| NCT03851705/ MDCO-PCS-17-02/CKJX839A12302 | A study of inclisiran in participants with homozygous familial hypercholesterolemia (HoFH) (ORION-5) | *** Completed/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Investigator), Primary Purpose: Treatment | 6-Feb-2019 to 9-Sep-2021 | NCT03399370/ MDCO-PCS-17-04 | Inclisiran for participants with atherosclerotic cardiovascular disease and elevated low-density lipoprotein Cholesterol (ORION-10) | *** Completed/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Care Provider), Primary Purpose: Treatment | 21-Dec-2017 to 17-Sep-2019 |
| NCT03400800/MDCO-PCS-17-08 | Inclisiran for subjects with ASCVD or ASCVD-risk equivalents and elevated low-density lipoprotein cholesterol | *** Completed/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Care Provider), Primary | 1-Nov-2017 to 27-Aug-2019 | NCT03397121/ MDCO-PCS-17-03/2017-002472-30 | Trial to evaluate the effect of inclisiran treatment on low density lipoprotein cholesterol (LDL-C) in subjects with heterozygous familial hypercholesterolemia (HeFH) (ORION-9) | *** Completed/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Care Provider), Primary Purpose: Treatment | 28-Nov-2017 to 17-Sep-2019 |
| NCT04765657/ CKJX839A12307 | Study of efficacy and safety of inclisiran in asian participants with atherosclerotic cardiovascular disease (ASCVD) or ASCVD high risk and elevated low density lipoprotein cholesterol (LDL-C) | ** Active, not recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Treatment | 1-Mar-2021 to 30-Sep-2022 | NCT03705234/ CTSU_MDCO_PCS-17-01 | A randomized trial assessing the effects of inclisiran on clinical outcomes among people with cardiovascular disease (ORION-4) | * Recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Prevention | 30-Oct-2018 to Dec 2049 |
| NCT05360446/ CKJX839D12303, 2021-004601-47 | Coronary computed tomography study to assess the effect of inclisiran in addition to maximally tolerated statin therapy on atherosclerotic plaque progression in participants with a diagnosis of non-obstructive coronary artery disease | * Recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Triple (Participant, Care Provider, Investigator), Primary Purpose: Treatment | 8-Jul-2022 to 5-Jun-2025 | NCT05004675/ LIB003-012 | Trial to evaluate efficacy and safety of lib003 and inclisiran in high-risk CVD Patients | ***** Enrolling by invitation/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor), Primary Purpose: Treatment | 20-Jun-2022 to31-0ct-2023 |
| NCT05030428/ CKJX839B12302 | Study of inclisiran to prevent cardiovascular (CV) events in participants with established cardiovascular disease | * Recruiting/Interventional/Phase 3 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Investigator), Primary Purpose: Treatment | 23-Nov-2021 to 13-Oct-2027 | NCT02314442/ ALN-PCSSC-001 | A phase 1 study of an investigational drug, aln-pcssc, in subjects with elevated low density lipoprotein cholesterol (LDL-C) | *** Completed/Interventional/Phase 1 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Single (Participant), Primary Purpose: Treatment | Dec-2014 to Nov-2015 |
| NCT02597127/ MDCO-PCS-15-01 | Trial to evaluate the effect of ALN-PCSSC treatment on low density lipoprotein cholesterol (LDL-C) (ORION-1) | *** Completed/Interventional/Phase 2 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Double (Participant, Investigator), Primary Purpose: Treatment | Jan-2016 to 7-Jun-2017 | NCT05438069/ 2021-2429 | German inclisiran network: retrospective registry of patients being treated with the siRNA inclisiran in Germany | * Recruiting/Observational | Observational Model: Cohort, Time Perspective: Retrospective | 11-Dec-2020 to 11-Dec-2025 |
| NCT01437059/ ALN-PCS02-001 | Trial to evaluate safety and tolerability of ALN-PCS02 in subjects with elevated LDL-Cholesterol (LDL-C) (ORION 2) | *** Completed/Interventional/Phase 1 | Allocation: Randomized, Intervention Model: Parallel Assignment, Masking: Single (Participant), Primary Purpose: Treatment | Sep 2011-Sep 2012 | NCT02963311/ MDCO-PCS-16-02 | A study of ALN-PCSSC in participants with homozygous familial hypercholesterolemia (HoFH) | *** Completed/Interventional/Phase 2 | Allocation: N/A, Intervention Model: Single Group Assignment, Masking: None (Open Label), Primary Purpose: Treatment | 13-Dec to 8-Oct-2018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebenezer, O.; Comoglio, P.; Wong, G.K.-S.; Tuszynski, J.A. Development of Novel siRNA Therapeutics: A Review with a Focus on Inclisiran for the Treatment of Hypercholesterolemia. Int. J. Mol. Sci. 2023, 24, 4019. https://doi.org/10.3390/ijms24044019
Ebenezer O, Comoglio P, Wong GK-S, Tuszynski JA. Development of Novel siRNA Therapeutics: A Review with a Focus on Inclisiran for the Treatment of Hypercholesterolemia. International Journal of Molecular Sciences. 2023; 24(4):4019. https://doi.org/10.3390/ijms24044019
Chicago/Turabian StyleEbenezer, Oluwakemi, Pietro Comoglio, Gane Ka-Shu Wong, and Jack A. Tuszynski. 2023. "Development of Novel siRNA Therapeutics: A Review with a Focus on Inclisiran for the Treatment of Hypercholesterolemia" International Journal of Molecular Sciences 24, no. 4: 4019. https://doi.org/10.3390/ijms24044019
APA StyleEbenezer, O., Comoglio, P., Wong, G. K.-S., & Tuszynski, J. A. (2023). Development of Novel siRNA Therapeutics: A Review with a Focus on Inclisiran for the Treatment of Hypercholesterolemia. International Journal of Molecular Sciences, 24(4), 4019. https://doi.org/10.3390/ijms24044019