

Impact of the Voltage-Gated Calcium Channel Antagonist Nimodipine on the Development of Oligodendrocyte Precursor Cells
 , , ,
, , , 
Abstract
1. Introduction
2. Results
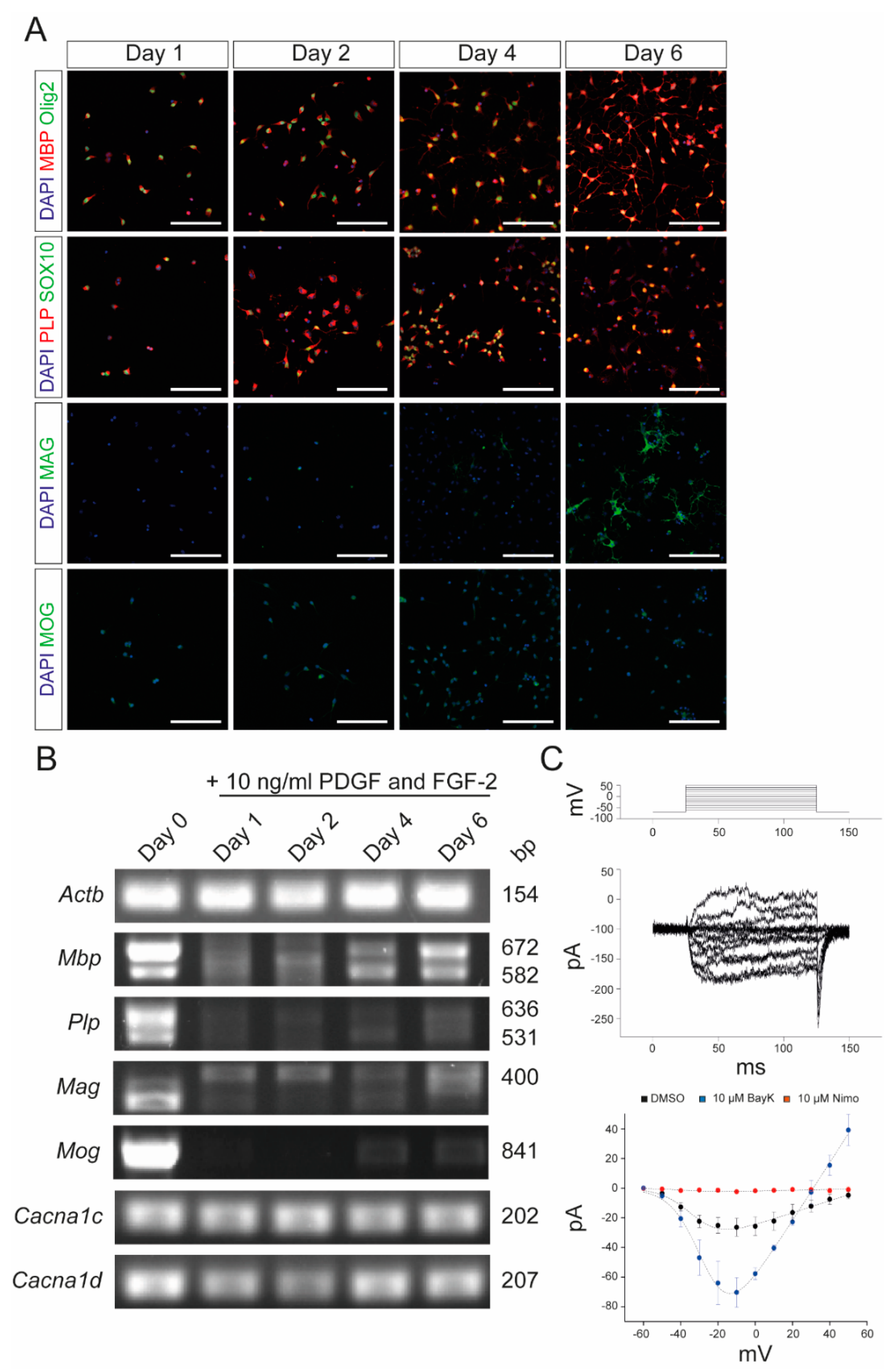
2.1. Oli-Neu Cells Share Common Characteristics with OPCs and Express Functional Cav1.2
2.2. Nimodipine Does Not Have an Effect on Myelin Gene Expression in Oli-Neu Cells
2.3. Nimodipine Does Not Have Any Effect on Myelin Protein Synthesis in Primary OPCs from Neonatal Mice
2.4. Effect of Nimodipine on the mRNA Level in OPCs
2.5. Nimodipine Increases the Number of Oligodendrocytes, but Not OPCs, in an In Vivo Model
3. Discussion
4. Materials and Methods
4.1. Nimodipine and BayK8644
4.2. Oli-Neu Cell Line
4.3. ICC
4.4. RNA Isolation and PCR
4.5. Patch-Clamp Analysis
4.6. Mice
4.7. OPC Isolation and Culture
4.8. Quantitative PCR
4.9. Bulk mRNA-Seq
4.10. Fish
4.11. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lassmann, H. Pathogenic mechanisms associated with different clinical courses of multiple sclerosis. Front. Immunol. 2018, 9, 3116. [Google Scholar] [CrossRef] [PubMed]
- Frischer, J.M.; Bramow, S.; Dal-Bianco, A.; Lucchinetti, C.F.; Rauschka, H.; Schmidbauer, M.; Laursen, H.; Sorensen, P.S.; Lassmann, H. The relation between inflammation and neurodegeneration in multiple sclerosis brains. Brain 2009, 132, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H. Multiple Sclerosis Pathology. Cold Spring Harb. Perspect. Med. 2018, 8, a028936. [Google Scholar] [CrossRef]
- Weber, M.S.; Menge, T.; Lehmann-Horn, K.; Kronsbein, H.C.; Zettl, U.; Sellner, J.; Hemmer, B.; Stüve, O. Current treatment strategies for multiple sclerosis-efficacy versus neurological adverse effects. Curr. Pharm. Des. 2012, 18, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Van Schaik, P.E.M.; Zuhorn, I.S.; Baron, W. Targeting fibronectin to overcome remyelination failure in multiple sclerosis: The need for brain- and lesion-targeted drug delivery. Int. J. Mol. Sci. 2022, 23, 8418. [Google Scholar] [CrossRef]
- Klistorner, A.; Barnett, M. Remyelination trials: Are we expecting the unexpected? Neurol. Neuroimmunol. Neuroinflamm. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Trapp, B.D. Relapsing and progressive forms of multiple sclerosis: Insights from pathology. Curr. Opin. Neurol. 2014, 27, 271–278. [Google Scholar] [CrossRef]
- Calabresi, P.A. Trials and tribulations on the path to remyelination. Lancet Neurol. 2021, 20, 686–687. [Google Scholar] [CrossRef]
- Cunniffe, N.; Coles, A. Promoting remyelination in multiple sclerosis. J. Neurol. 2021, 268, 30–44. [Google Scholar] [CrossRef]
- Hanf, K.J.M.; Arndt, J.W.; Liu, Y.; Gong, B.J.; Rushe, M.; Sopko, R.; Massol, R.; Smith, B.; Gao, Y.; Dalkilic-Liddle, I.; et al. Functional activity of anti-LINGO-1 antibody opicinumab requires target engagement at a secondary binding site. MAbs 2020, 12, 1713648. [Google Scholar] [CrossRef]
- Cadavid, D.; Mellion, M.; Hupperts, R.; Edwards, K.R.; Calabresi, P.A.; Drulović, J.; Giovannoni, G.; Hartung, H.-P.; Arnold, D.L.; Fisher, E.; et al. Safety and efficacy of opicinumab in patients with relapsing multiple sclerosis (SYNERGY): A randomised, placebo-controlled, phase 2 trial. Lancet Neurol. 2019, 18, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Ineichen, B.V.; Kapitza, S.; Bleul, C.; Good, N.; Plattner, P.S.; Seyedsadr, M.S.; Kaiser, J.; Schneider, M.P.; Zörner, B.; Martin, R.; et al. Nogo-A antibodies enhance axonal repair and remyelination in neuro-inflammatory and demyelinating pathology. Acta Neuropathol. 2017, 134, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xiong, J.-Q.; Ren, X.-B.; Sun, W. The role of Nogo-A in neuroregeneration: A review. Brain Res. Bull. 2012, 87, 499–503. [Google Scholar] [CrossRef]
- Scriabine, A.; van den Kerckhoff, W. Pharmacology of nimodipine. A review. Ann. N. Y. Acad. Sci. 1988, 522, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.S.; Ahn, H.S.; Preziosi, T.J.; Battye, R.; Boone, S.C.; Chou, S.N.; Kelly, D.L.; Weir, B.K.; Crabbe, R.A.; Lavik, P.J.; et al. Cerebral arterial spasm--a controlled trial of nimodipine in patients with subarachnoid hemorrhage. New Engl. J. Med. 1983, 308, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Höllerhage, H.G.; Gaab, M.R.; Zumkeller, M.; Walter, G.F. The influence of nimodipine on cerebral blood flow autoregulation and blood-brain barrier. J. Neurosurg. 1988, 69, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Dejong, G.; Deweerd, H.; Schuurman, T.; Traber, J.; Luiten, P. Microvascular changes in aged rat forebrain. Effects of chronic nimodipine treatment. Neurobiol. Aging 1990, 11, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Kong, R.M.; Stillman, M.J.; Shukitt-Hale, B.; Kadar, T.; Rauch, T.M.; Lieberman, H.R. Nimodipine improves spatial working memory and elevates hippocampal acetylcholine in young rats. Pharmacol. Biochem. Behav. 1991, 39, 781–786. [Google Scholar] [CrossRef]
- Taya, K.; Watanabe, Y.; Kobayashi, H.; Fujiwara, M. Nimodipine improves the disruption of spatial cognition induced by cerebral ischemia. Physiol. Behav. 2000, 70, 19–25. [Google Scholar] [CrossRef]
- Schampel, A.; Volovitch, O.; Koeniger, T.; Scholz, C.-J.; Jörg, S.; Linker, R.A.; Wischmeyer, E.; Wunsch, M.; Hell, J.W.; Ergün, S.; et al. Nimodipine fosters remyelination in a mouse model of multiple sclerosis and induces microglia-specific apoptosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3295–E3304. [Google Scholar] [CrossRef]
- Ingwersen, J.; Santi, L.D.; Wingerath, B.; Graf, J.; Koop, B.; Schneider, R.; Hecker, C.; Schröter, F.; Bayer, M.; Engelke, A.D.; et al. Nimodipine confers clinical improvement in two models of experimental autoimmune encephalomyelitis. J. Neurochem. 2018, 146, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Leisz, S.; Simmermacher, S.; Prell, J.; Strauss, C.; Scheller, C. Nimodipine-dependent protection of schwann cells, astrocytes and neuronal cells from osmotic, oxidative and heat stress is associated with the activation of AKT and CREB. Int. J. Mol. Sci. 2019, 20, 4578. [Google Scholar] [CrossRef] [PubMed]
- Zech, J.; Leisz, S.; Göttel, B.; Syrowatka, F.; Greiner, A.; Strauss, C.; Knolle, W.; Scheller, C.; Mäder, K. Electrospun Nimodipine-loaded fibers for nerve regeneration: Development and in vitro performance. Eur. J. Pharm. Biopharm. 2020, 151, 116–126. [Google Scholar] [CrossRef]
- Zamora, N.N.; Cheli, V.T.; Santiago González, D.A.; Wan, R.; Paez, P.M. Deletion of voltage-gated calcium channels in astrocytes during demyelination reduces brain inflammation and promotes myelin regeneration in mice. J. Neurosci. 2020, 40, 3332–3347. [Google Scholar] [CrossRef] [PubMed]
- Boltz, F.; Enders, M.; Feigenspan, A.; Kirchner, P.; Ekici, A.; Kuerten, S. Nimodipine exerts beneficial effects on the rat oligodendrocyte cell line OLN-93. Brain Sci. 2022, 12, 476. [Google Scholar] [CrossRef]
- Jung, M.; Krämer, E.; Grzenkowski, M.; Tang, K.; Blakemore, W.; Aguzzi, A.; Khazaie, K.; Chlichlia, K.; Blankenfeld, G.V.; Kettenmann, H. Lines of murine oligodendroglial precursor cells immortalized by an activated neu tyrosine kinase show distinct degrees of interaction with axons in vitro and in vivo. Eur. J. Neurosci. 1995, 7, 1245–1265. [Google Scholar] [CrossRef]
- Simon, K.; Hennen, S.; Merten, N.; Blättermann, S.; Gillard, M.; Kostenis, E.; Gomeza, J. The orphan G protein-coupled receptor GPR17 negatively regulates oligodendrocyte differentiation via Gαi/o and its downstream effector molecules. J. Biol. Chem. 2016, 291, 705–718. [Google Scholar] [CrossRef]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.A.; Ratnayake, U.; Dickinson, H.; Castillo-Melendez, M.; Walker, D.W. Ontogenetic Change in the regional distribution of dehydroepiandrosterone-synthesizing enzyme and the glucocorticoid receptor in the brain of the spiny mouse (Acomys cahirinus). Dev. Neurosci. 2016, 38, 54–73. [Google Scholar] [CrossRef]
- Chung, R.S.; Hidalgo, J.; West, A.K. New insight into the molecular pathways of metallothionein-mediated neuroprotection and regeneration. J. Neurochem. 2008, 104, 14–20. [Google Scholar] [CrossRef]
- Teuber-Hanselmann, S.; Meinl, E.; Junker, A. MicroRNAs in gray and white matter multiple sclerosis lesions: Impact on pathophysiology. J. Pathol. 2020, 250, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.; Teuber-Hanselmann, S.; Soub, D.; Harnisch, K.; Mairinger, F.; Junker, A. MicroRNA profiles of MS gray matter lesions identify modulators of the synaptic protein synaptotagmin-7. Brain Pathol. 2020, 30, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Volsko, C.; Datta, U.; Regev, K.; Dutta, R. Expression of disease-related miRNAs in white-matter lesions of progressive multiple sclerosis brains. Ann. Clin. Transl. Neurol. 2019, 6, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Rahimirad, S.; Navaderi, M.; Alaei, S.; Sanati, M.H. Identification of hsa-miR-106a-5p as an impact agent on promotion of multiple sclerosis using multi-step data analysis. Neurol. Sci. 2021, 42, 3791–3799. [Google Scholar] [CrossRef]
- Seuter, S.; Pehkonen, P.; Heikkinen, S.; Carlberg, C. The gene for the transcription factor BHLHE40/DEC1/stra13 is a dynamically regulated primary target of the vitamin D receptor. J. Steroid Biochem. Mol. Biol. 2013, 136, 62–67. [Google Scholar] [CrossRef]
- Shaked, I.; Hanna, R.N.; Shaked, H.; Chodaczek, G.; Nowyhed, H.N.; Tweet, G.; Tacke, R.; Basat, A.B.; Mikulski, Z.; Togher, S.; et al. Transcription factor Nr4a1 couples sympathetic and inflammatory cues in CNS-recruited macrophages to limit neuroinflammation. Nat. Immunol. 2015, 16, 1228–1234. [Google Scholar] [CrossRef]
- Yu, H.-Z.; Zhu, B.-Q.; Zhu, L.; Li, S.; Wang, L.-M. NR4A1 agonist cytosporone B attenuates neuroinflammation in a mouse model of multiple sclerosis. Neural Regen. Res. 2022, 17, 2765–2770. [Google Scholar] [CrossRef]
- Penkowa, M.; Espejo, C.; Martínez-Cáceres, E.M.; Montalban, X.; Hidalgo, J. Increased demyelination and axonal damage in metallothionein I+II-deficient mice during experimental autoimmune encephalomyelitis. Cell. Mol. Life Sci. 2003, 60, 185–197. [Google Scholar] [CrossRef]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system: Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 489–503. [Google Scholar] [CrossRef]
- Baldassarro, V.A.; Krężel, W.; Fernández, M.; Schuhbaur, B.; Giardino, L.; Calzà, L. The role of nuclear receptors in the differentiation of oligodendrocyte precursor cells derived from fetal and adult neural stem cells. Stem Cell Res. 2019, 37, 101443. [Google Scholar] [CrossRef]
- Kapfhammer, J.P.; Schwab, M.E. Inverse patterns of myelination and GAP-43 expression in the adult CNS: Neurite growth inhibitors as regulators of neuronal plasticity? J. Comp. Neurol. 1994, 340, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, F.; Nedelcu, J.; Leopold, P.; Zhan, J.; Clarner, T.; Nellessen, L.; Beißel, C.; van Heuvel, Y.; Goswami, A.; Weis, J.; et al. Cuprizone-induced graded oligodendrocyte vulnerability is regulated by the transcription factor DNA damage-inducible transcript 3. Glia 2019, 67, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Bodmer, R.; Bier, E.; Karin, M. Sestrins at the crossroad between stress and aging. Aging 2010, 2, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Odabas, F.O.; Uca, A.U.; Akdag, T.; Demirdögen, F.; Altas, M.; Tokgoz, O.S. Possible roles of sestrin2 in multiple sclerosis and its relationships with clinical outcomes. Arq. Neuropsiquiatr. 2022, 80, 399–404. [Google Scholar] [CrossRef]
- Li, P.; Stetler, R.A.; Leak, R.K.; Shi, Y.; Li, Y.; Yu, W.; Bennett, M.V.L.; Chen, J. Oxidative stress and DNA damage after cerebral ischemia: Potential therapeutic targets to repair the genome and improve stroke recovery. Neuropharmacology 2018, 134, 208–217. [Google Scholar] [CrossRef]
- Kiba, T. Relationships between ventromedial hypothalamic lesions and the expressions of neuron-related genes in visceral organs. Neurosci. Res. 2012, 74, 1–6. [Google Scholar] [CrossRef]
- He, D.; Wang, J.; Lu, Y.; Deng, Y.; Zhao, C.; Xu, L.; Chen, Y.; Hu, Y.-C.; Zhou, W.; Lu, Q.R. lncRNA Functional Networks in Oligodendrocytes Reveal Stage-Specific Myelination Control by an lncOL1/Suz12 Complex in the CNS. Neuron 2017, 93, 362–378. [Google Scholar] [CrossRef]
- Salehi, Z.; Hadiyan, S.P.; Navidi, R. Ciliary neurotrophic factor role in myelin oligodendrocyte glycoprotein expression in Cuprizone-induced multiple sclerosis mice. Cell. Mol. Neurobiol. 2013, 33, 531–535. [Google Scholar] [CrossRef]
- Mueller, A.M.; Pedré, X.; Stempfl, T.; Kleiter, I.; Couillard-Despres, S.; Aigner, L.; Giegerich, G.; Steinbrecher, A. Novel role for SLPI in MOG-induced EAE revealed by spinal cord expression analysis. J. Neuroinflammation 2008, 5, 20. [Google Scholar] [CrossRef]
- Richter-Landsberg, C.; Heinrich, M. S-100 immunoreactivity in rat brain glial cultures is associated with both astrocytes and oligodendrocytes. J. Neurosci. Res. 1995, 42, 657–665. [Google Scholar] [CrossRef]
- Chaerkady, R.; Letzen, B.; Renuse, S.; Sahasrabuddhe, N.A.; Kumar, P.; All, A.H.; Thakor, N.V.; Delanghe, B.; Gearhart, J.D.; Pandey, A.; et al. Quantitative temporal proteomic analysis of human embryonic stem cell differentiation into oligodendrocyte progenitor cells. Proteomics 2011, 11, 4007–4020. [Google Scholar] [CrossRef] [PubMed]
- Chamling, X.; Kallman, A.; Fang, W.; Berlinicke, C.A.; Mertz, J.L.; Devkota, P.; Pantoja, I.E.M.; Smith, M.D.; Ji, Z.; Chang, C.; et al. Single-cell transcriptomic reveals molecular diversity and developmental heterogeneity of human stem cell-derived oligodendrocyte lineage cells. Nat. Commun. 2021, 12, 652. [Google Scholar] [CrossRef] [PubMed]
- Durand, B.; Gao, F.B.; Raff, M. Accumulation of the cyclin-dependent kinase inhibitor p27/Kip1 and the timing of oligodendrocyte differentiation. EMBO J. 1997, 16, 306–317. [Google Scholar] [CrossRef] [PubMed]
- La Fuente, A.G.D.; Queiroz, R.M.L.; Ghosh, T.; McMurran, C.E.; Cubillos, J.F.; Bergles, D.E.; Fitzgerald, D.C.; Jones, C.A.; Lilley, K.S.; Glover, C.P.; et al. Changes in the oligodendrocyte progenitor cell proteome with ageing. Mol. Cell. Proteomics 2020, 19, 1281–1302. [Google Scholar] [CrossRef]
- Arnold, S.E.; Arvanitakis, Z.; Macauley-Rambach, S.L.; Koenig, A.M.; Wang, H.-Y.; Ahima, R.S.; Craft, S.; Gandy, S.; Buettner, C.; Stoeckel, L.E.; et al. Brain insulin resistance in type 2 diabetes and Alzheimer disease: Concepts and conundrums. Nat. Rev. Neurol. 2018, 14, 168–181. [Google Scholar] [CrossRef]
- Bruinsma, I.B.; van Dijk, M.; Bridel, C.; van de Lisdonk, T.; Haverkort, S.Q.; Runia, T.F.; Steinman, L.; Hintzen, R.Q.; Killestein, J.; Verbeek, M.M.; et al. Regulator of oligodendrocyte maturation, miR-219, a potential biomarker for MS. J. Neuroinflamm. 2017, 14, 235. [Google Scholar] [CrossRef]
- Inamura, N.; Go, S.; Watanabe, T.; Takase, H.; Takakura, N.; Nakayama, A.; Takebayashi, H.; Matsuda, J.; Enokido, Y. Reduction in miR-219 expression underlies cellular pathogenesis of oligodendrocytes in a mouse model of Krabbe disease. Brain Pathol. 2021, 31, e12951. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Ong, W.; Wang, K.; Wang, M.; Nizetic, D.; Chew, S.Y. Effects of miR-219/miR-338 on microglia and astrocyte behaviors and astrocyte-oligodendrocyte precursor cell interactions. Neural Regen. Res. 2020, 15, 739–747. [Google Scholar] [CrossRef]
- Wang, H.; Moyano, A.L.; Ma, Z.; Deng, Y.; Lin, Y.; Zhao, C.; Zhang, L.; Jiang, M.; He, X.; Ma, Z.; et al. miR-219 Cooperates with miR-338 in myelination and promotes myelin repair in the CNS. Dev. Cell 2017, 40, 566–582.e5. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Zhou, F.; Li, J.; Lu, G.; Zhao, Y. MiR-20a-5p Regulates MPP+-induced oxidative stress and neuroinflammation in HT22 cells by targeting IRF9/NF-κB Axis. Evid. Based Complement. Altern. Med. 2021, 2021, 6621206. [Google Scholar] [CrossRef]
- Balkan, E.; Bilge, N. Expression levels of IL-17/IL-23 cytokine-targeting microRNAs 20, 21, 26, 155, and Let-7 in patients with relapsing-remitting multiple sclerosis. Neurol. Res. 2021, 43, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Bazrgar, M.; Khodabakhsh, P.; Prudencio, M.; Mohagheghi, F.; Ahmadiani, A. The role of microRNA-34 family in Alzheimer’s disease: A potential molecular link between neurodegeneration and metabolic disorders. Pharmacol. Res. 2021, 172, 105805. [Google Scholar] [CrossRef] [PubMed]
- Junker, A.; Krumbholz, M.; Eisele, S.; Mohan, H.; Augstein, F.; Bittner, R.; Lassmann, H.; Wekerle, H.; Hohlfeld, R.; Meinl, E. MicroRNA profiling of multiple sclerosis lesions identifies modulators of the regulatory protein CD47. Brain 2009, 132, 3342–3352. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-T.; Fu, Y.-H. miR-23 regulation of lamin B1 is crucial for oligodendrocyte development and myelination. Dis. Model. Mech. 2009, 2, 178–188. [Google Scholar] [CrossRef]
- Lin, S.-T.; Huang, Y.; Zhang, L.; Heng, M.Y.; Ptácek, L.J.; Fu, Y.-H. MicroRNA-23a promotes myelination in the central nervous system. Proc. Natl. Acad. Sci. USA 2013, 110, 17468–17473. [Google Scholar] [CrossRef]
- Adusumilli, L.; Facchinello, N.; Teh, C.; Busolin, G.; Le, M.T.; Yang, H.; Beffagna, G.; Campanaro, S.; Tam, W.L.; Argenton, F.; et al. miR-7 controls the dopaminergic/oligodendroglial fate through Wnt/β-catenin signaling regulation. Cells 2020, 9, 711. [Google Scholar] [CrossRef]
- Katsushima, K.; Shinjo, K.; Natsume, A.; Ohka, F.; Fujii, M.; Osada, H.; Sekido, Y.; Kondo, Y. Contribution of microRNA-1275 to Claudin11 protein suppression via a polycomb-mediated silencing mechanism in human glioma stem-like cells. J. Biol. Chem. 2012, 287, 27396–27406. [Google Scholar] [CrossRef]
- Li, A.; Song, W.; Qian, J.; Li, Y.; He, J.; Zhang, Q.; Li, W.; Zhai, A.; Kao, W.; Hu, Y.; et al. MiR-122 modulates type I interferon expression through blocking suppressor of cytokine signaling 1. Int. J. Biochem. Cell Biol. 2013, 45, 858–865. [Google Scholar] [CrossRef]
- Lewkowicz, P.; Cwiklińska, H.; Mycko, M.P.; Cichalewska, M.; Domowicz, M.; Lewkowicz, N.; Jurewicz, A.; Selmaj, K.W. Dysregulated RNA-induced silencing complex (RISC) assembly within CNS corresponds with abnormal miRNA expression during autoimmune demyelination. J. Neurosci. 2015, 35, 7521–7537. [Google Scholar] [CrossRef]
- Münzel, E.J.; Schaefer, K.; Obirei, B.; Kremmer, E.; Burton, E.A.; Kuscha, V.; Becker, C.G.; Brösamle, C.; Williams, A.; Becker, T. Claudin k is specifically expressed in cells that form myelin during development of the nervous system and regeneration of the optic nerve in adult zebrafish. Glia 2012, 60, 253–270. [Google Scholar] [CrossRef]
- Tiwari-Woodruff, S.K.; Buznikov, A.G.; Vu, T.Q.; Micevych, P.E.; Chen, K.; Kornblum, H.I.; Bronstein, J.M. OSP/claudin-11 forms a complex with a novel member of the tetraspanin super family and beta1 integrin and regulates proliferation and migration of oligodendrocytes. J. Cell Biol. 2001, 153, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Long, K.L.P.; Breton, J.M.; Barraza, M.K.; Perloff, O.S.; Kaufer, D. Hormonal Regulation of Oligodendrogenesis I: Effects across the Lifespan. Biomolecules 2021, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef]
- Cannell, I.G.; Kong, Y.W.; Bushell, M. How do microRNAs regulate gene expression? Biochem. Soc. Trans. 2008, 36, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-W.; Ren, S.-H.; Ren, J.-R.; Zhen, Z.-G.; Li, L.-R.; Hao, X.-D.; Ji, H.-M. Nimodipine improves cognitive impairment after subarachnoid hemorrhage in rats through IncRNA NEAT1/miR-27a/MAPT axis. Drug Des. Devel. Ther. 2020, 14, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Chen, Y.; Xie, W.; Liu, X.; Zhu, Y.; Zhu, Y. Nimodipine attenuates tau phosphorylation at Ser396 via miR-132/GSK-3β pathway in chronic cerebral hypoperfusion rats. Eur. J. Pharmacol. 2018, 819, 1–8. [Google Scholar] [CrossRef]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueff, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef]
- Chiozzi, P.; Sarti, A.C.; Sanz, J.M.; Giuliani, A.L.; Adinolfi, E.; Vultaggio-Poma, V.; Falzoni, S.; Di Virgilio, F. Amyloid β-dependent mitochondrial toxicity in mouse microglia requires P2X7 receptor expression and is prevented by nimodipine. Sci. Rep. 2019, 9, 6475. [Google Scholar] [CrossRef]
- Matute, C. P2X7 receptors in oligodendrocytes: A novel target for neuroprotection. Mol. Neurobiol. 2008, 38, 123–128. [Google Scholar] [CrossRef]
- Langley, M.S.; Sorkin, E.M. Nimodipine. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic potential in cerebrovascular disease. Drugs 1989, 37, 669–699. [Google Scholar] [CrossRef]
- Rämsch, K.D.; Ahr, G.; Tettenborn, D.; Auer, L.M. Overview on pharmacokinetics of nimodipine in healthy volunteers and in patients with subarachnoid hemorrhage. Neurochirurgia 1985, 28 (Suppl. 1), 74–78. [Google Scholar] [CrossRef] [PubMed]
- Podbielska, M.; Banik, N.L.; Kurowska, E.; Hogan, E.L. Myelin recovery in multiple sclerosis: The challenge of remyelination. Brain Sci. 2013, 3, 1282–1324. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche, S.; Strauss, C.; Scheller, C.; Leisz, S. Nimodipine Treatment protects auditory hair cells from cisplatin-induced cell death accompanied by upregulation of LMO4. Int. J. Mol. Sci. 2022, 23, 5780. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, X.; Liu, Y.; Bao, Y.; An, L. Nimodipine protects dopaminergic neurons against inflammation-mediated degeneration through inhibition of microglial activation. Neuropharmacology 2009, 56, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Verma, P.; Balaji, G.; Samantaray, S.; Mohanakumar, K.P. Nimodipine, an L-type calcium channel blocker attenuates mitochondrial dysfunctions to protect against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinsonism in mice. Neurochem. Int. 2016, 99, 221–232. [Google Scholar] [CrossRef]
- Herzfeld, E.; Strauss, C.; Simmermacher, S.; Bork, K.; Horstkorte, R.; Dehghani, F.; Scheller, C. Investigation of the neuroprotective impact of nimodipine on Neuro2a cells by means of a surgery-like stress model. Int. J. Mol. Sci. 2014, 15, 18453–18465. [Google Scholar] [CrossRef]
- Herzfeld, E.; Speh, L.; Strauss, C.; Scheller, C. Nimodipine but not nifedipine promotes expression of fatty acid 2-hydroxylase in a surgical stress model based on Neuro2a cells. Int. J. Mol. Sci. 2017, 18, 964. [Google Scholar] [CrossRef]
- Cheli, V.T.; Santiago González, D.A.; Namgyal Lama, T.; Spreuer, V.; Handley, V.; Murphy, G.G.; Paez, P.M. Conditional Deletion of the L-Type Calcium Channel Cav1.2 in Oligodendrocyte Progenitor Cells Affects Postnatal Myelination in Mice. J. Neurosci. 2016, 36, 10853–10869. [Google Scholar] [CrossRef]
- Cheli, V.T.; Santiago González, D.A.; Spreuer, V.; Paez, P.M. Voltage-gated Ca2+ entry promotes oligodendrocyte progenitor cell maturation and myelination in vitro. Exp. Neurol. 2015, 265, 69–83. [Google Scholar] [CrossRef]
- Babai, N.; Gierke, K.; Müller, T.; Regus-Leidig, H.; Brandstätter, J.H.; Feigenspan, A. Signal transmission at invaginating cone photoreceptor synaptic contacts following deletion of the presynaptic cytomatrix protein Bassoon in mouse retina. Acta Physiol. 2019, 226, e13241. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. Royal Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Nagarajan, B.; Harder, A.; Japp, A.; Häberlein, F.; Mingardo, E.; Kleinert, H.; Yilmaz, Ö.; Zoons, A.; Rau, B.; Christ, A.; et al. CNS myelin protein 36K regulates oligodendrocyte differentiation through Notch. Glia 2020, 68, 509–527. [Google Scholar] [CrossRef]
- Shin, J.; Park, H.-C.; Topczewska, J.M.; Mawdsley, D.J.; Appel, B. Neural cell fate analysis in zebrafish using olig2 BAC transgenics. Methods Cell Sci. 2003, 25, 7–14. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Key Functions of the Gene with Its Relevance to This Study | Impact of Nimodipine | Sources |
|---|---|---|---|
| Bhlhe40 | This growth factor increases the production of cytokines in T cells. The factor itself is regulated by the vitamin D receptor. | Increased expression | [35] |
| Nr4a1 | NR41A is expressed in several cell types. Nr41a is known to prevent immune cell infiltration into the CNS. | Increased expression | [36,37] |
| Mt1 | MT1 is capable of binding to free radicals and heavy metal ions and has neuroprotective potential in the CNS; Mt1−/− and Mt2−/− EAE mice display an increased number of CNS lesions. | Increased expression | [30,38,39] |
| Cyb5r1 | CYB5R1 expression has been described to increase the brain volume in the early phases of development. | Increased expression | [29] |
| Nr1d1 | This nuclear receptor displays higher expression in neuronal stem cells than fetal cells. | Increased expression | [40] |
| Gap43 | The level of GAP43 correlates negatively with the formation of myelin in gray matter. | Decreased expression | [41] |
| Ddit3 | DDIT3 is activated when demyelination occurs. In the cuprizone mouse model, Ddit3-/- knockout mice show reduced demyelination. | Increased expression | [42] |
| Sesn2 | SESN2 has been described as a universal growth factor. In MS patients, serum levels of SESN2 are decreased. | Increased expression | [43,44] |
| Gadd45b | The DNA methylase, GADD45B, increases transcription and stimulates axonal regeneration. | Increased expression | [45] |
| Serpini1 | The direct effect of SERPINI1 is unknown. It is speculated that there is a negative correlation between neuronal growth and SERPINI1 expression. | Decreased expression | [46] |
| Pik3r3 | PIK3R3 regulates lipid metabolism and is part of a myelin synthesis pathway. | Increased expression | [47] |
| Cntfr | CNTFR controls the expression of MOG. | Increased expression | [48] |
| Gene | Key Functions of the Gene with Its Relevance to This Study. | Impact of Nimodipine | Sources |
|---|---|---|---|
| Magi2 | MAGI2 has been described to be involved in synaptic transmission. MAGI2 expression was decreased in EAE. | Increased expression | [49] |
| S100A2 | This calcium sensor is crucial for the organization of the cytoskeleton and the microtubule network in oligodendrocytes. | Increased expression | [50] |
| Oxsr1 | OXSR1 is upregulated when embryonic stem cells change to OPCs. | Increased expression | [51] |
| Tsc22d1 | TSC22D1 is a transcription factor that is upregulated in oligodendrocytes and downregulated in astrocytes. | Increased expression | [52] |
| Cdc2 | CDC2, which regulates the cell cycle, is present in OPCs but absent in mature oligodendrocytes. | Increased expression | [53] |
| Pfdn5 | PFDN5 is a molecular chaperone that stabilizes newly-synthesized proteins, allowing them to fold correctly. It is decreased in aging OPCs. | Decreased expression | [54] |
| Mknk | This kinase is responsible for lipid synthesis in oligodendrocytes. | Increased expression | [55] |
| miRNA | Expression Pattern | Known Interacting Partner | Impact of Nimodipine | Source |
|---|---|---|---|---|
| miR-106 | Lower expression in blood samples and in gray matter lesions of MS patients | miR-106 is a negative regulator for the amyloid precursor protein | Increased expression | [34] |
| miR-219-2-3p | Lower expression in the cerebrospinal fluid of MS patients and in gray matter lesions | miR-219 is crucial for developing OPCs via, e.g., the suppression of caspase 3 expression | Increased expression | [56,57,58,59] |
| miR-219b-3p | ||||
| miR-20a | Lower expression in gray matter lesions in MS patients | miR-20a suppresses the expression of NF-κB and IL-17/IL-23 | Increased expression | [60,61] |
| miR-34a-3p | Increased expression in patients with Alzheimer’s disease | miR-34a-3p acts as a target for CD47. CD47 is a “do not eat me” surface marker | Decreased expression | [62,63] |
| miR-34a-5p | Decreased expression in lesions of MS patients | miR-34a-5p reduces the expression of, e.g., sirtuin 1, which increases differentiation in neuronal cells | Increased expression | [62,63] |
| miR-23 | Expression is reduced in gray matter lesions in MS patients | miR-23 is a negative regulator of lamin B1, which is essential for oligodendrogenesis | Increased Expression | [64,65] |
| miR-338 | Decreased expression in white and gray matter lesions of MS patients | Interaction partner of miR-219; increases the expression of myelin | Increased expression | [58,59] |
| miR-7 | Downregulation in white matter lesions of MS patients | miR-7 inhibits Wnt and activates Shh signaling, both of which increase myelination | Increased expression | [66] |
| miR-1275 | Upregulation in white matter lesions of MS patents | Inhibition of miR-1275 increases the expression of claudin 11 | Decreased expression | [67] |
| miR-122 | Expression is increased in white matter lesions of MS patients | miR-122 expression increases the expression of proinflammatory type I interferons | Decreased expression | [68,69] |
| Gene | Forward Primer | Reverse Primer | Annealing Temperature | Length (bp) |
|---|---|---|---|---|
| Actb | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT | 55 °C | 154 |
| Mbp | CGGACCCAAGATGAAAACCC | AAAGGAAGCCTGGACCACACAG | 56 °C | 582, 672 |
| Plp | GAAAAGCTAATTGAGACCTA | GAGCAGGGAAACTAGTGTGG | 45 °C | 531, 636 |
| Mag | CTCTATGGCACCCAGAGCCT | TGTCCTTGGTGGGTCGTTTT | 56 °C | 355, 400 |
| Mog | GACCTCAGCTTGGCCTGACCC | TGCTGGGCTCTCCTTCCGC | 63 °C | 841 |
| Cacna1s | TGTGGTATGTCGTCACTTCCTCC | CGTCAATGATGCTGCCGATG | 57 °C | 258 |
| Cacna1c | CAGCTCATGCCAACATGAAT | TGCTTCTTGGGTTTCCCATA | 53 °C | 202 |
| Cacna1d | AATGGCACGGAATGTAGGAG | GACGAAAAATGAGCCAAGGA | 52 °C | 207 |
| Cacna1f | GACGAATGCACAAGACATGC | CAAGCACAAGGTTGAGGACA | 55 °C | 324 |
| Gene | Forward Primer | Reverse Primer | Ann. Temp. | Length (bp) |
|---|---|---|---|---|
| Actb | ACGATATCGCTGCGCTGG | AGCATCGTCGCCCGC | 60 °C | 71 |
| Mbp | GGCACGCTTTCCAAAATCT | GGAGGGCTCTCAGCGTCT | 60 °C | 81 |
| Plp | TCAGTCTATTGCCTTCCCTAGC | AGCATTCCATGGGAGAACAC | 60 °C | 91 |
| Mag | CCAGTACACCTTCTCGGAGC | TCCGGCACCATACAACTGAC | 60 °C | 114 |
| Mog | ATGCTTACATGGAGGTTGGGC | TTCCTCCAAGAAGCCCGAAG | 60 °C | 70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enders, M.; Weier, A.; Chunder, R.; An, Y.; Bremm, F.; Feigenspan, A.; Buettner, C.; Ekici, A.B.; Mingardo, E.; Odermatt, B.; et al. Impact of the Voltage-Gated Calcium Channel Antagonist Nimodipine on the Development of Oligodendrocyte Precursor Cells. Int. J. Mol. Sci. 2023, 24, 3716. https://doi.org/10.3390/ijms24043716
Enders M, Weier A, Chunder R, An Y, Bremm F, Feigenspan A, Buettner C, Ekici AB, Mingardo E, Odermatt B, et al. Impact of the Voltage-Gated Calcium Channel Antagonist Nimodipine on the Development of Oligodendrocyte Precursor Cells. International Journal of Molecular Sciences. 2023; 24(4):3716. https://doi.org/10.3390/ijms24043716
Chicago/Turabian StyleEnders, Michael, Alicia Weier, Rittika Chunder, Young An, Franziska Bremm, Andreas Feigenspan, Christian Buettner, Arif Bülent Ekici, Enrico Mingardo, Benjamin Odermatt, and et al. 2023. "Impact of the Voltage-Gated Calcium Channel Antagonist Nimodipine on the Development of Oligodendrocyte Precursor Cells" International Journal of Molecular Sciences 24, no. 4: 3716. https://doi.org/10.3390/ijms24043716
APA StyleEnders, M., Weier, A., Chunder, R., An, Y., Bremm, F., Feigenspan, A., Buettner, C., Ekici, A. B., Mingardo, E., Odermatt, B., & Kuerten, S. (2023). Impact of the Voltage-Gated Calcium Channel Antagonist Nimodipine on the Development of Oligodendrocyte Precursor Cells. International Journal of Molecular Sciences, 24(4), 3716. https://doi.org/10.3390/ijms24043716