
Inhibitory Effect of Styrylpyrone Extract of Phellinus linteus on Hepatic Steatosis in HepG2 Cells

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
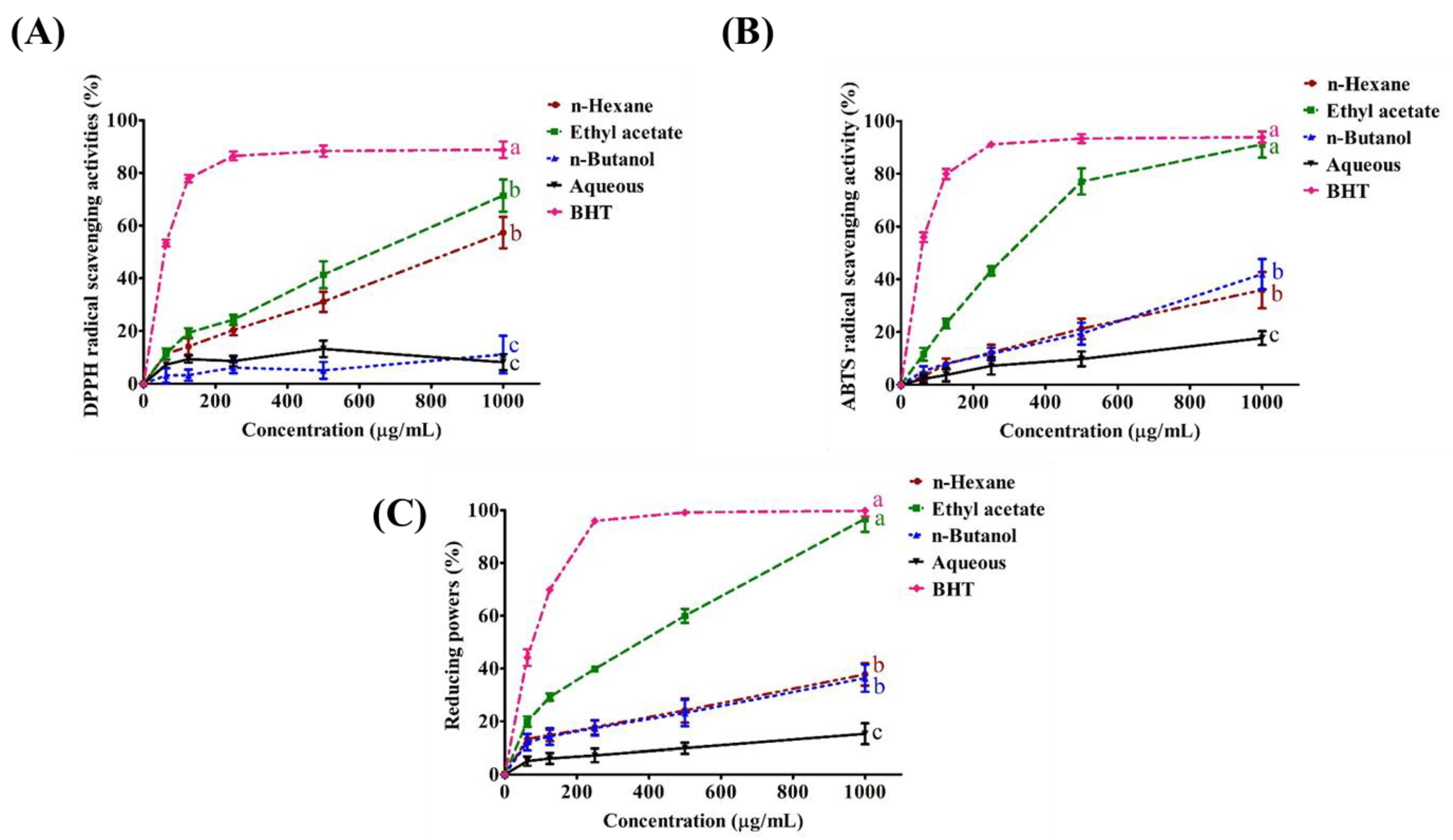
2.1. Yield of Extraction from Different Solvents and the Antioxidant Activity of Each Extract
2.2. Antioxidant Activity of SPEE
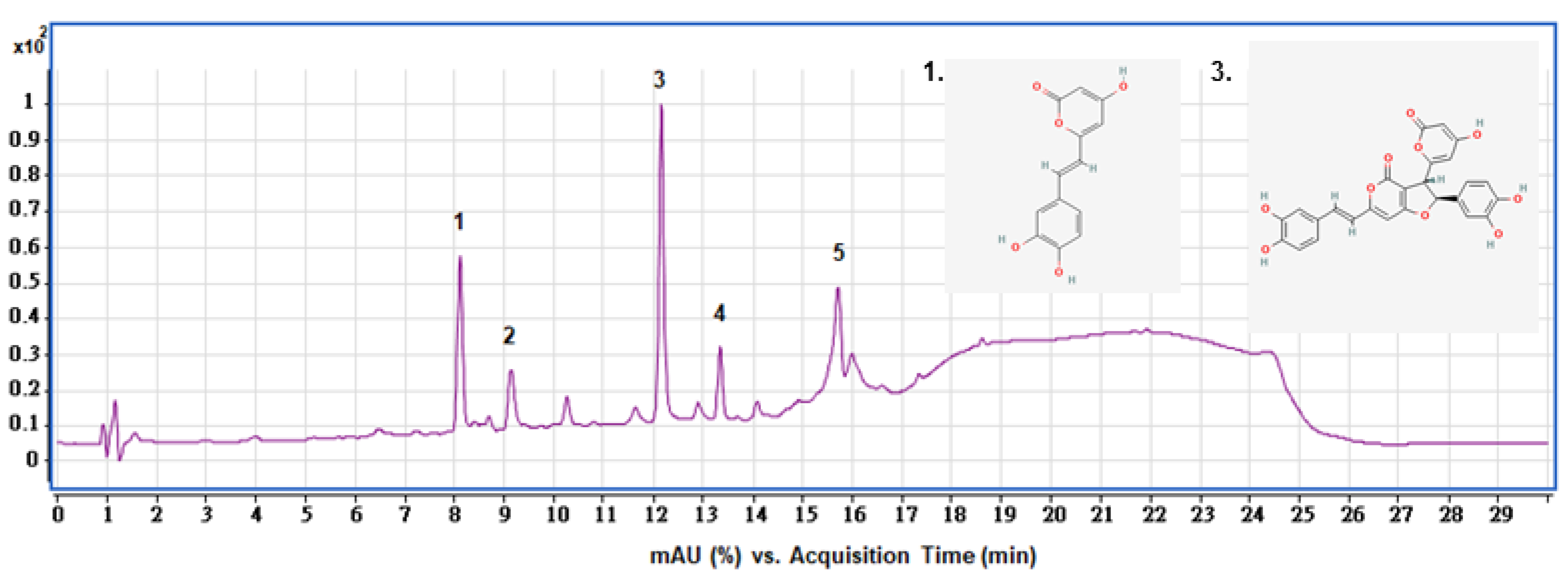
2.3. HPLC Analysis of SPEE
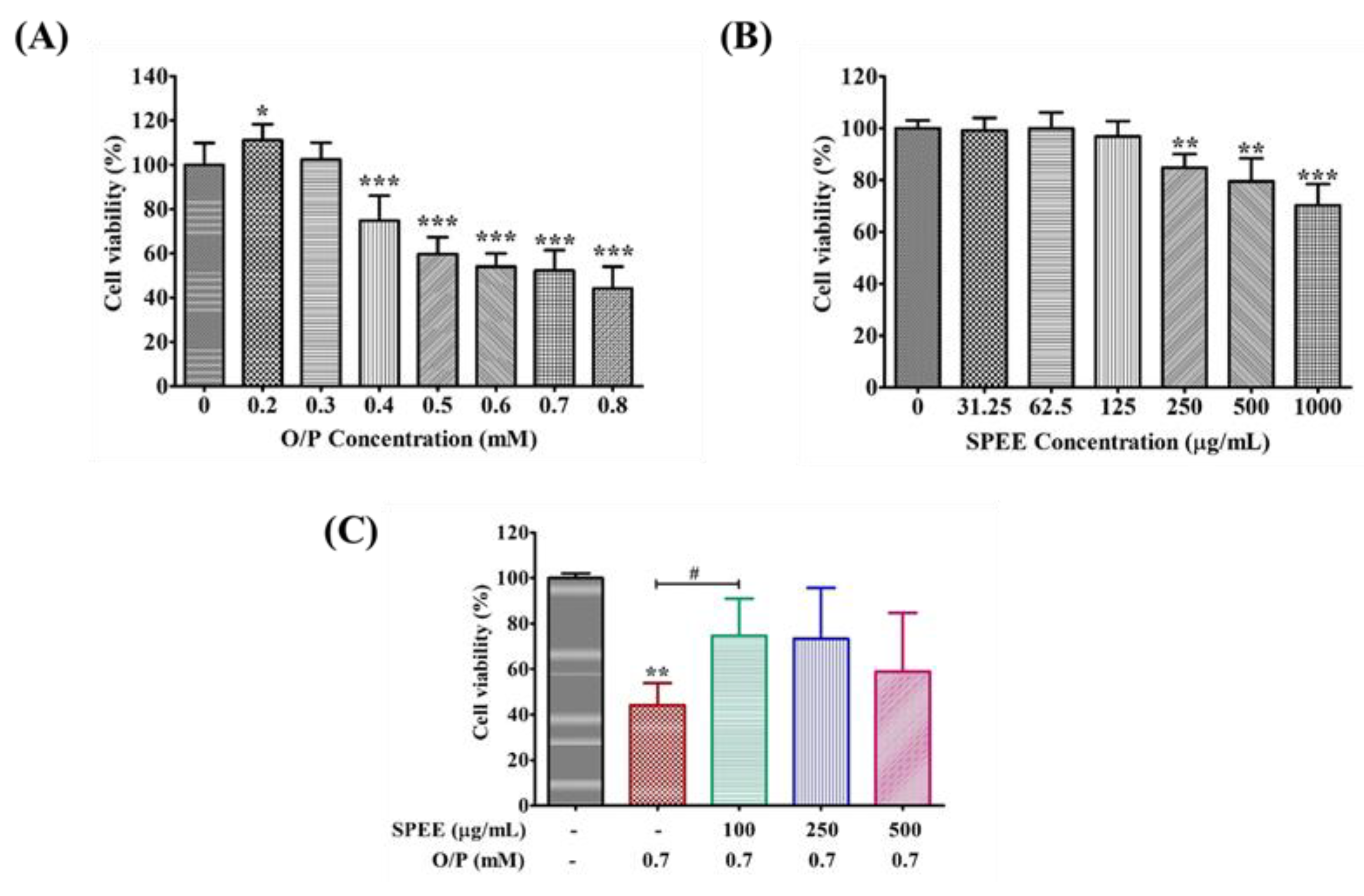
2.4. Effects of SPEE on O/P-Induced Cytotoxicity in HepG2 Cells
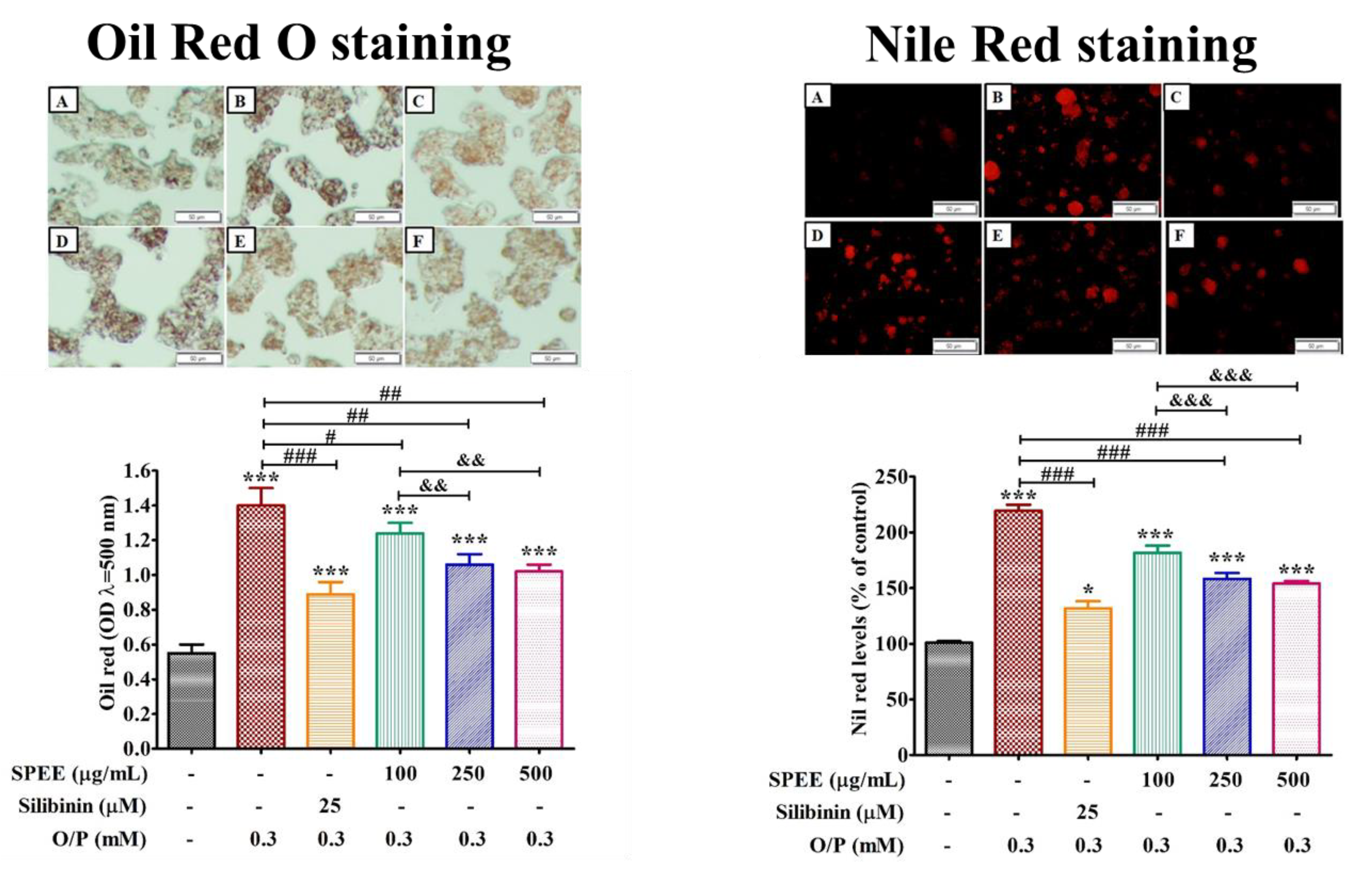
2.5. Effects of SPEE on O/P-Induced Lipid Droplet Deposition
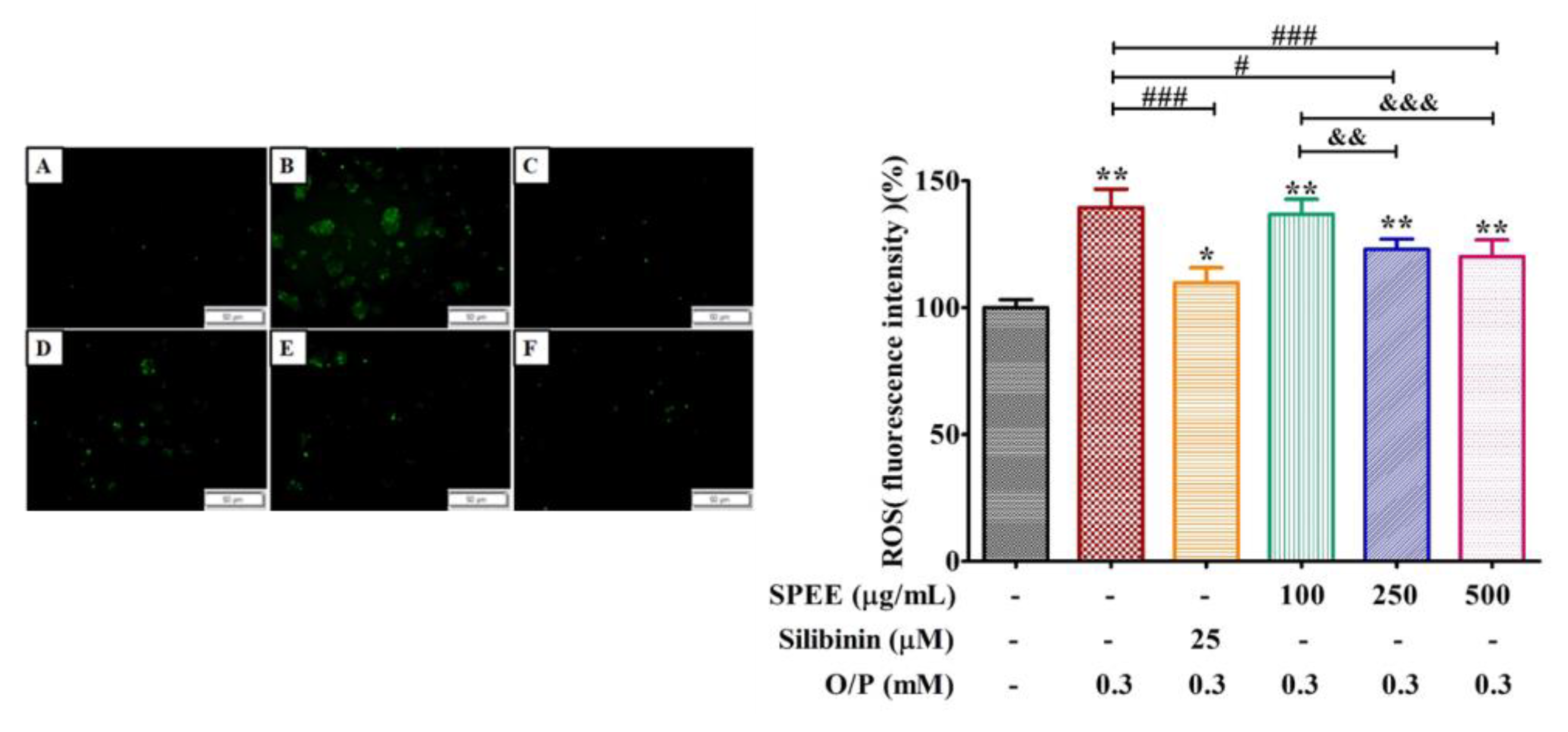
2.6. Effects of SPEE on O/P-Induced Production of Intracellular Reactive Oxygen Species (ROS)
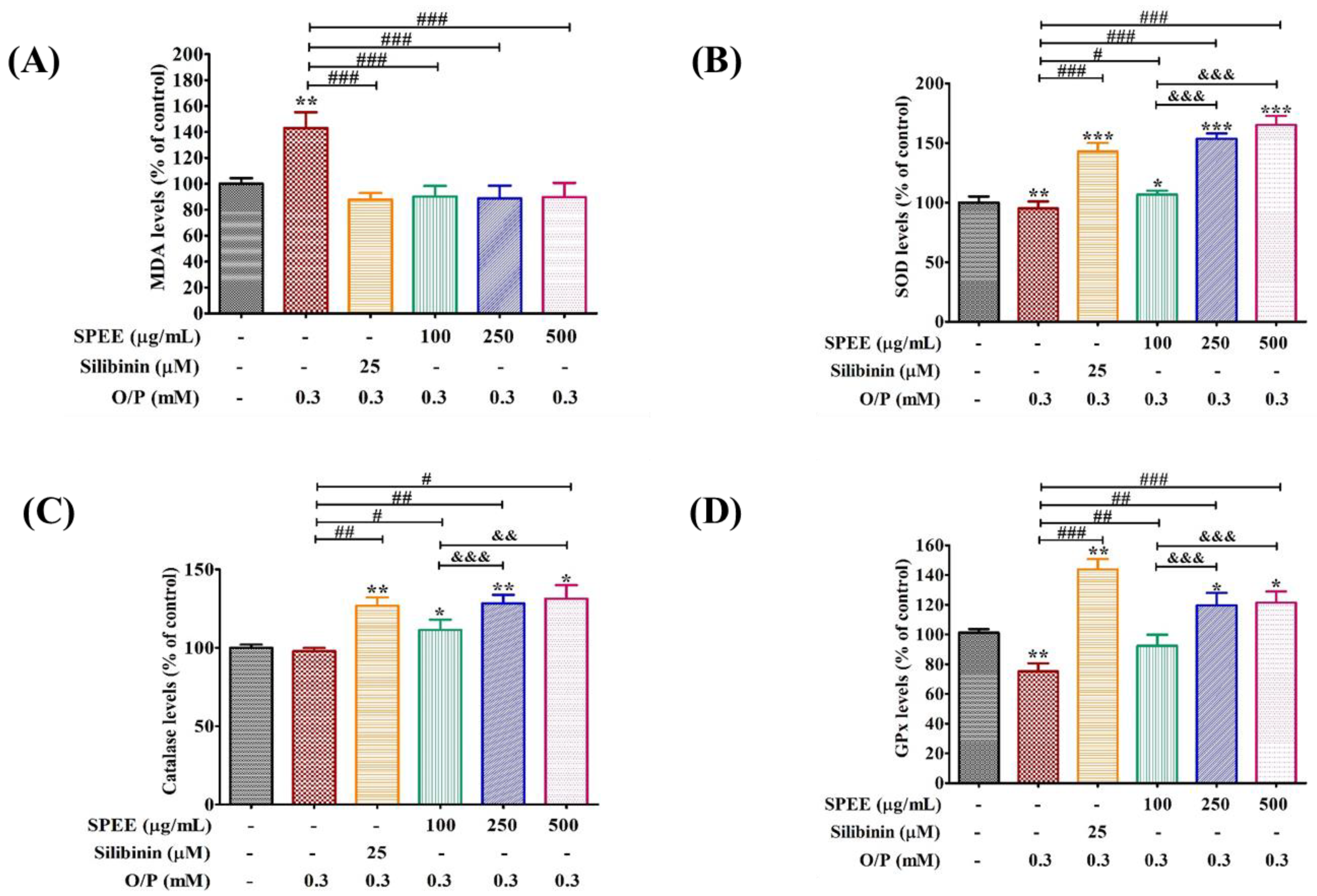
2.7. Effects of SPEE on O/P-Induced Antioxidant Enzyme Activities
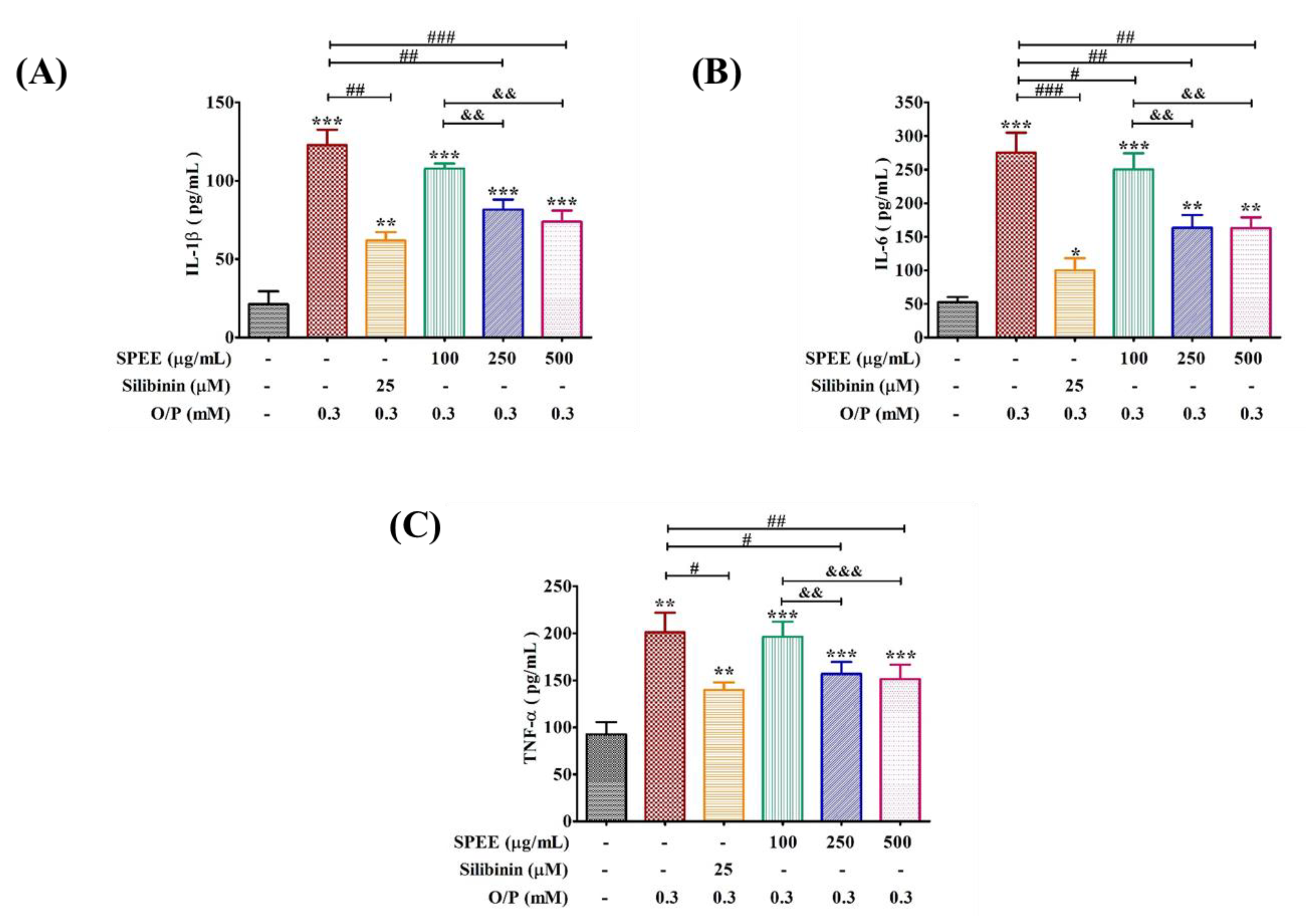
2.8. Effects of SPEE on O/P-Induced Pro-Inflammatory Signals
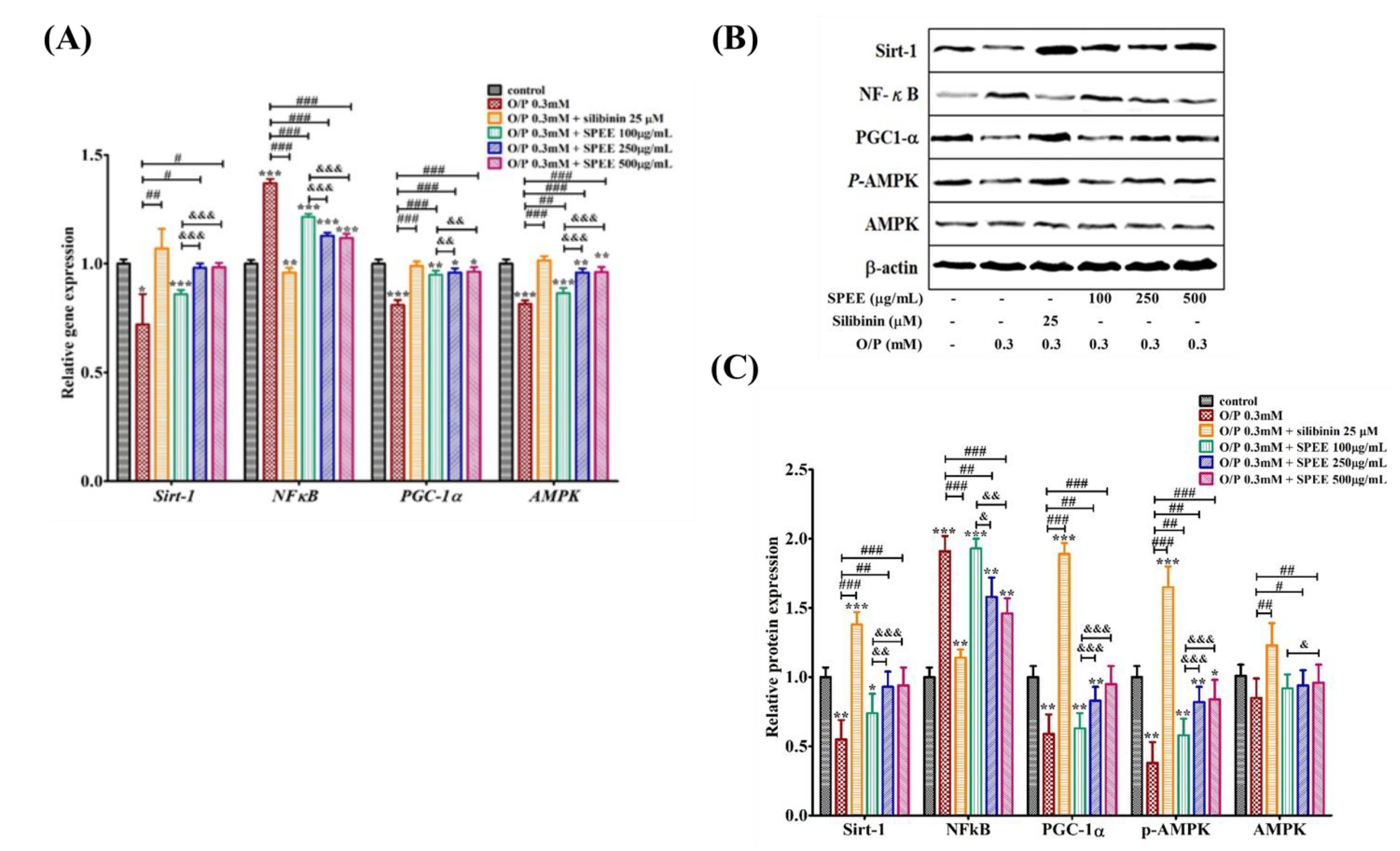
2.9. Effects of SPEE on O/P-Induced Expression of Intracellular mRNAs
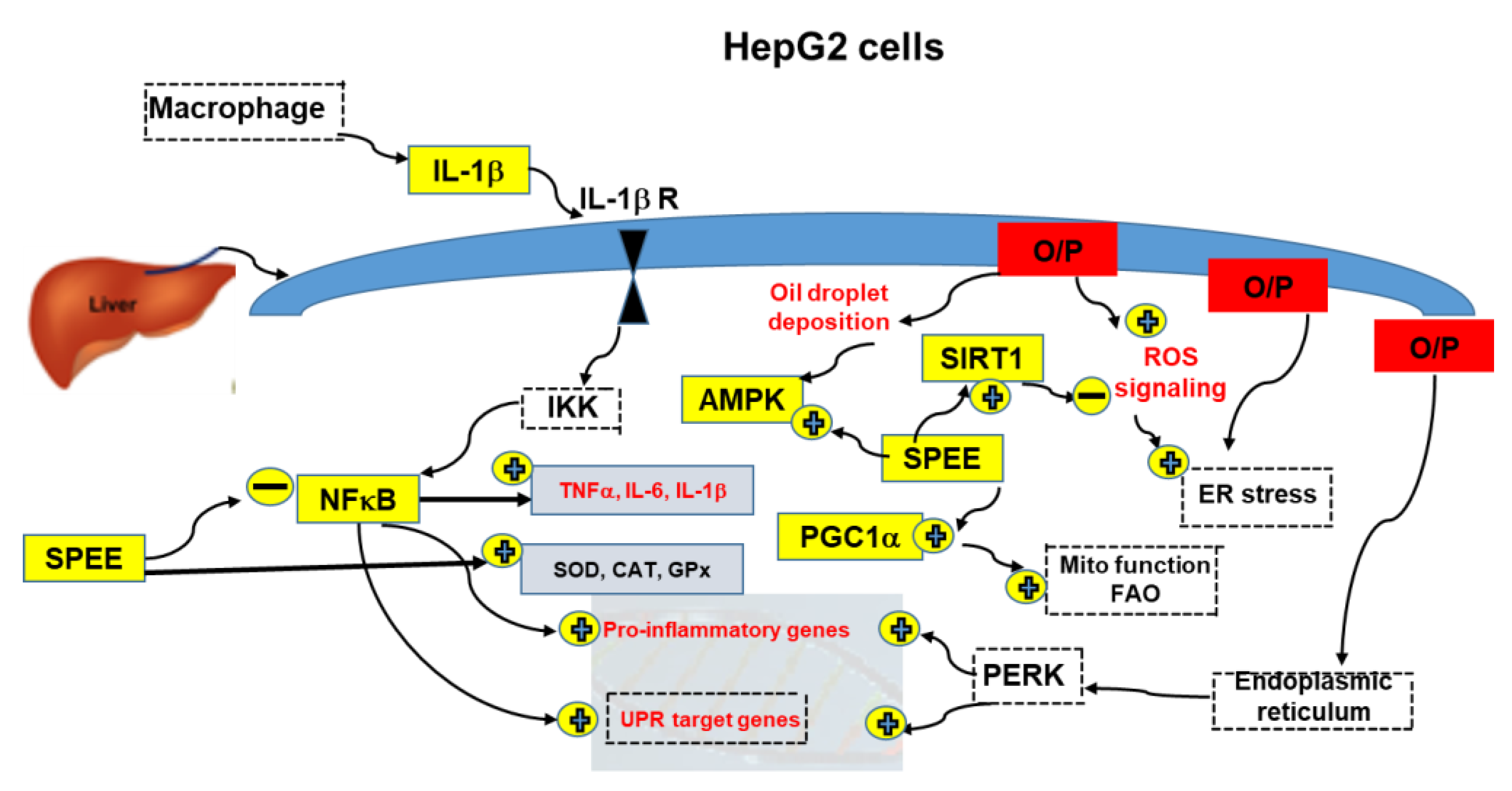
3. Discussion
3.1. The Plausible Strong Antioxidant Activities of SPEE
3.2. How Did SPEE Rescue the HepG2 Cell Viability Insulted by O/P?
3.3. SPEE Dose-Dependently Inhibited the Intracellular Lipid Droplet Deposition
3.4. SPEE Suppressed ROS Signaling and Induced Antioxidant Enzyme Activities
3.5. SPEE Alleviated ROS-Induced Pro-Inflammatory Signals under Insult of O/P
3.6. SPEE Modulated and Alleviated O/P-Induced Intracellular mRNA Expressions
4. Materials and Methods
4.1. Chemicals and Instruments
4.2. Culture of HepG2 Cells
4.3. Induction of HepG2 Cell Line with O/P
4.4. MTT Assay
4.5. Oil Red O Staining
4.6. Nile Red Staining
4.7. Assay for Intracellular ROS Content
4.8. Assay for Protein Content
4.9. Assay for Malondialdehyde Content
4.10. Assay for the Intracellular Antioxidative Enzymes
4.10.1. Assay for Superoxide Dismutase (SOD) Activity
4.10.2. Assay for the Catalase Activity
4.10.3. Assay for the Glutathione Peroxidase Activity
4.11. Assay for the Inflammatory Factors
4.11.1. Assay for the TNF-α Content
4.11.2. Assay for the IL-6 Content
4.11.3. Assay for the IL-1β Content
4.12. Analysis for Gene Expression
4.12.1. Extraction of RNA
4.12.2. Reverse Transcription to Obtain cDNA
4.12.3. Real-Time Polymerase Chain Reaction
4.13. SDS-PAGE
4.13.1. Separation of Proteins by Electrophoresis
4.13.2. Electrotransfer
4.13.3. Western Blot
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazarus, J.V.; Mark, H.E.; Anstee, Q.M.; Arab, J.P.; Batterham, R.L.; Castera, L.; Cortez-Pinto, H.; Crespo, J.; Cusi, K.; Dirac, M.A.; et al. Advancing the global public health agenda for NAFLD: A consensus statement. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Cotter, T.G.; Rinella, M. Nonalcoholic fatty liver disease 2020: The state of the disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Baranova, A.; Drapkina, O.; Tonevitsky, A. Cell-free circulating nucleic acids as early biomarkers for NAFLD and NAFLD-associated disorders. Front. Physiol. 2018, 9, 1256. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Huh, Y.; Cho, Y.J.; Nam, G.E. Recent epidemiology and risk factors of nonalcoholic fatty liver disease. J. Obes. Metab. Syndr. 2022, 31, 17–27. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease–meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, incidence, and outcome of non-alcoholic fatty liver disease in Asia, 1999–2019: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2019, 4, 389–398. [Google Scholar] [CrossRef]
- Huang, D.Q.; El-Serag, H.B.; Loomba, R. Global epidemiology of NAFLD-related HCC: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 223–238. [Google Scholar] [CrossRef]
- Starley, B.Q.; Calcagno, C.J.; Harrison, S.A. Nonalcoholic fatty liver disease and hepatocellular carcinoma: A weighty connection. Hepatology 2010, 51, 1820–1832. [Google Scholar] [CrossRef]
- Marchesi, E.; Rota, C.; Fann, Y.C.; Chignell, C.F.; Mason, R.P. Photoreduction of the fluorescent dye 2′-7′-dichlorofluorescein: A spin trapping and direct electron spin resonance study with implications for oxidative stress measurements. Free Radic. Biol. Med. 1999, 26, 148–161. [Google Scholar] [CrossRef]
- Chen, W.; Tan, H.; Liu, Q.; Zheng, X.; Zhang, H.; Liu, Y.; Xu, L. The bioactivities and pharmacological applications of Phellinus linteus. A Review. Molecules 2019, 24, 1888. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Kim, S.H.; Chen, C.Y. A medicinal mushroom: Phellinus linteus. Curr. Med. Chem. 2008, 15, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.G.; Exposito, T.S.; Marante, F.J.T. Lanosterol derivatives from Phellinus torulosus. J. Phytochem. 1994, 35, 1523–1526. [Google Scholar] [CrossRef]
- Liu, H.-K.; Tsai, T.-H. Lanostane-triterpenoids from the fungus Phellinus gilvus. J. Phytochem. 2009, 70, 558–563. [Google Scholar] [CrossRef]
- Wang, G.J.; Tsai, T.H.; Chang, T.T. Lanostanes from Phellinus igniarius and their iNOS inhibitory activities. Planta Med. 2009, 75, 1602–1607. [Google Scholar] [CrossRef]
- Lu, T.-L.; Huang, G.-J.; Lu, T.-J. Hispolon from Phellinus linteus has antiproliferative effects via MDM2-recruited ERK1/2 activity in breast and bladder cancer cells. J. Food Chem. Toxicol. 2009, 47, 2013–2021. [Google Scholar] [CrossRef]
- Shou, D.; Dong, Y.; Wang, N.; Li, H.; Zhang, Y.; Zhu, Y. Simultaneous quantification of antioxidant compoundsin Phellinus igniarius using ultra performance liquid chromatography photodiode array detection-electrospray ionization tandem mass spectrometry. PLoS ONE 2016, 11, e0163797. [Google Scholar] [CrossRef]
- Ali, N.A.A.; Jansen, R.; Pilgrim, H. Hispolon, a yellow pigment from Inonotus hispidus. J. Phytochem. 1996, 41, 927. [Google Scholar] [CrossRef]
- Lee, I.-K.; Lee, M.-S. Styrylpyrones from the medicinal fungus Phellinus baumii and their antioxidant properties. J. Bioorg. Med. Chem. Lett. 2010, 20, 5459–5461. [Google Scholar] [CrossRef]
- Chiu, C.-C.; Chang, C.-C.; Lin, J.-J.; Chen, C.-C.; Chyau, C.-C.; Peng, R.Y. Styrylpyrones from Phellinus linteus mycelia alleviate non-alcoholic fatty liver by modulating lipid and glucose metabolic homeostasis in high-fat and high-fructose diet-fed mice. Antioxidants 2022, 11, 898. [Google Scholar] [CrossRef]
- Lee, I.K.; Yun, B.S. Styrylpyrone-class compounds from medicinal fungi Phellinus and Inonotus spp., and their medicinal importance. J. Antibiot. 2011, 64, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Feng, X.L.; Liu, C.; Gao, J.M.; Qi, J. Diverse metabolites and pharmacological effects from the basidiomycetes Inonotus hispidus. Antibiotics 2022, 11, 1097. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, I.K.; Seok, S.J.; Lee, H.J.; Kim, Y.H.; Yun, B.S. Antioxidant polyphenols from the mycelial culture of the medicinal fungi Inonotus xeranticus and Phellinus linteus. J. Appl. Microbiol. 2008, 104, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Sun, W.; Wang, X.; Long, J.; Liu, X.; Feng, Z.; Liu, J. Punicalagin attenuates palmitate-induced lipotoxicity in HepG2 cells by activating the Keap1-Nrf2 antioxidant defense system. Mol. Nutr. Food Res. 2016, 60, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Yano, W.; Kubota, T.; Yamauchi, T.; Itoh, S.; Kumagai, H.; Kozono, H.; Takamoto, I.; Okamoto, S.; Shiuchi, T.; et al. Adiponectin stimulates AMP-activated protein kinase in the hypothalamus and increases food intake. Cell Metab. 2007, 6, 55–68. [Google Scholar] [CrossRef]
- Chou, I.P.; Lin, Y.Y.; Ding, S.T.; Chen, C.Y. Adiponectin receptor 1 enhances fatty acid metabolism and cell survival in palmitate-treated HepG2 cells through the PI3 K/AKT pathway. Eur. J. Nutr. 2014, 53, 907–917. [Google Scholar] [CrossRef]
- Wang, D.; Wei, Y.; Frye, M.; Gentile, C.L.; Pagliassotti, M.J. Saturated fatty acid-induced cytotoxicity in liver cells does not involve phosphatase and tensin homologue deleted on chromosome 10. J. Nutr. Metab. 2013, 2013, 514206. [Google Scholar] [CrossRef]
- Mubarak, S.A.; Al Otaibi, A.; Al Qarni, A.; Bakillah, A.; Iqbal, J. Reduction in insulin mediated ERK phosphorylation by palmitate in liver cells is independent of fatty acid induced ER stress. Nutrients 2022, 14, 3641. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Topczewski, F.; Pagliassotti, M.J. Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E275–E281. [Google Scholar] [CrossRef]
- Malhi, H.; Bronk, S.F.; Werneburg, N.W.; Gores, G.J. Free fatty acids induce JNK-dependent hepatocyte lipoapoptosis. J. Biol. Chem. 2006, 281, 12093–12101. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.W.; Lee, K.-T.; Lee, M.W.; Ka, K.-H. SIRT1 attenuates palmitate-induced endoplasmic reticulum stress and insulin resistance in HepG2 cells via induction of oxygen-regulated protein 150. Biochem. Biophys. Res. Commun. 2012, 422, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, S.; Wang, Y.; Zhu, T. Silymarin improved diet-induced liver damage and insulin resistance by decreasing inflammation in mice. Pharm. Biol. 2016, 54, 2995–3000. [Google Scholar] [CrossRef] [PubMed]
- Mbanefo, E.C.; Le, L.; Zee, R.; Banskota, N.; Ishida, K.; Pennington, L.F.; Odegaard, J.I.; Jardetzky, T.S.; Alouffi, A.; Falcone, F.H.; et al. IPSE, a urogenital parasite derived immunomodulatory protein, ameliorates ifosfamide induced hemorrhagic cystitis through downregulation of proinfammatory pathways. Sci. Rep. 2019, 9, 1586. [Google Scholar] [CrossRef]
- De Gregorio, E.; Colell, A.; Morales, A.; Marí, M. Relevance of SIRT1-NF-κB axis as therapeutic target to ameliorate inflammation in liver disease. Int. J. Mol. Sci. 2020, 21, 3858. [Google Scholar] [CrossRef]
- Fournier, M.; Paulson, A.; Pavelka, N.; Mosley, A.L.; Gaudenz, K.; Bradford, W.D.; Glynn, E.; Li, H.; Sardiu, M.E.; Fleharty, B.; et al. Delayed correlation of mRNA and protein expression in rapamycin-treated cells and a role for Ggc1 in cellular sensitivity to rapamycin. Mol. Cell Proteom. 2010, 9, 271–284. [Google Scholar] [CrossRef]
- Lyons, C.L.; Roche, H.M. Nutritional modulation of AMPK-impact upon metabolic-inflammation. Int. J. Mol. Sci. 2018, 19, 3092. [Google Scholar] [CrossRef]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef]
- Yao, H.R.; Liu, J.; Cao, Y.B. Lipotoxicity in HepG2 cells triggered by free fatty acids. Am. J. Transl. Res. 2011, 3, 284–291. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Beaudoin, A. New technique for revealing latent fingerprints on wet, porous surfaces: Oil red O. J. Forensic Identif. 2004, 54, 413–421. [Google Scholar]
- Phillip, G.; Mayer, E.P.; Fowler, S.D. Nile red: A selective fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar]
- Crow, J.P. Dichlorodihydrofluorescein and dihydrorhodamine 123 are sensitive indicators of peroxynitrite in vitro: Implications for intracellular measurement of reactive nitrogen and oxygen species. Nitric Oxide 1997, 1, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1996, 72, 248–254. [Google Scholar] [CrossRef]
- Alía, M.; Mateos, R.; Ramos, S.; Lecumberri, E.; Bravo, L.; Goya, L. Influence of quercetin and rutin on growth and antioxidant defense system of a human hepatoma cell line (HepG2). Eur J Nutr. 2006, 45, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Habig, W.H. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-H.; Chen, M.-Y.; Lieu, J.-J.; Chen, C.-C.; Chang, C.-C.; Chyau, C.-C.; Peng, R.Y. Inhibitory Effect of Styrylpyrone Extract of Phellinus linteus on Hepatic Steatosis in HepG2 Cells. Int. J. Mol. Sci. 2023, 24, 3672. https://doi.org/10.3390/ijms24043672
Chiu C-H, Chen M-Y, Lieu J-J, Chen C-C, Chang C-C, Chyau C-C, Peng RY. Inhibitory Effect of Styrylpyrone Extract of Phellinus linteus on Hepatic Steatosis in HepG2 Cells. International Journal of Molecular Sciences. 2023; 24(4):3672. https://doi.org/10.3390/ijms24043672
Chicago/Turabian StyleChiu, Chun-Hung, Ming-Yao Chen, Jun-Jie Lieu, Chin-Chu Chen, Chun-Chao Chang, Charng-Cherng Chyau, and Robert Y. Peng. 2023. "Inhibitory Effect of Styrylpyrone Extract of Phellinus linteus on Hepatic Steatosis in HepG2 Cells" International Journal of Molecular Sciences 24, no. 4: 3672. https://doi.org/10.3390/ijms24043672
APA StyleChiu, C.-H., Chen, M.-Y., Lieu, J.-J., Chen, C.-C., Chang, C.-C., Chyau, C.-C., & Peng, R. Y. (2023). Inhibitory Effect of Styrylpyrone Extract of Phellinus linteus on Hepatic Steatosis in HepG2 Cells. International Journal of Molecular Sciences, 24(4), 3672. https://doi.org/10.3390/ijms24043672