
High VEGFR3 Expression Reduces Doxorubicin Efficacy in Triple-Negative Breast Cancer
 , , , , , , , , and
, , , , , , , , and
Abstract
1. Introduction
2. Results
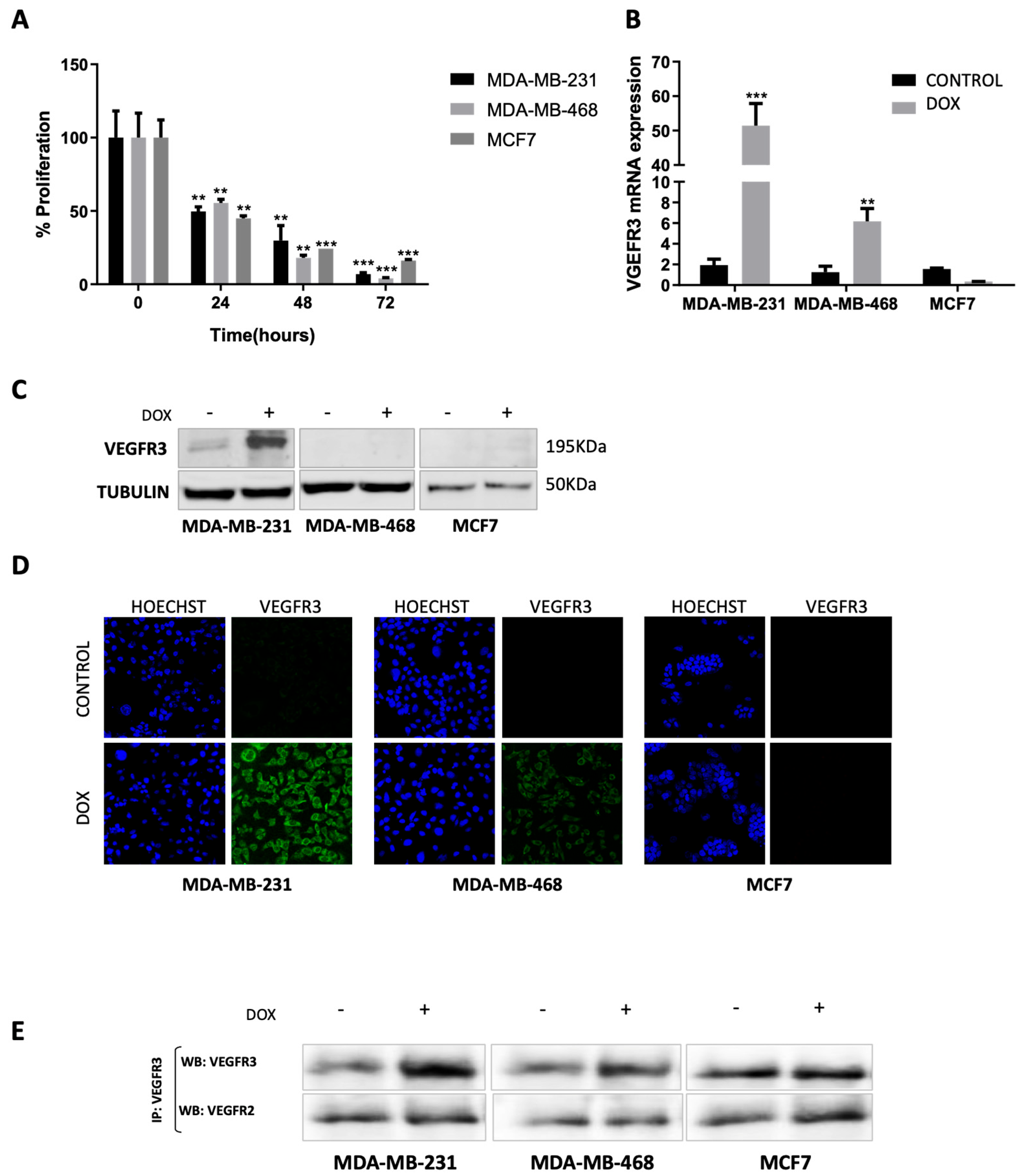
2.1. Doxorubicin Treatment Enhances VEGFR3 Gene Expression in MDA-MB-231 and MDA-MB-468 but Not in MCF7 Cells
2.2. VEGFR3 Protein Is Overexpressed after Doxorubicin Treatment in MDA-MB-231 Cell Line
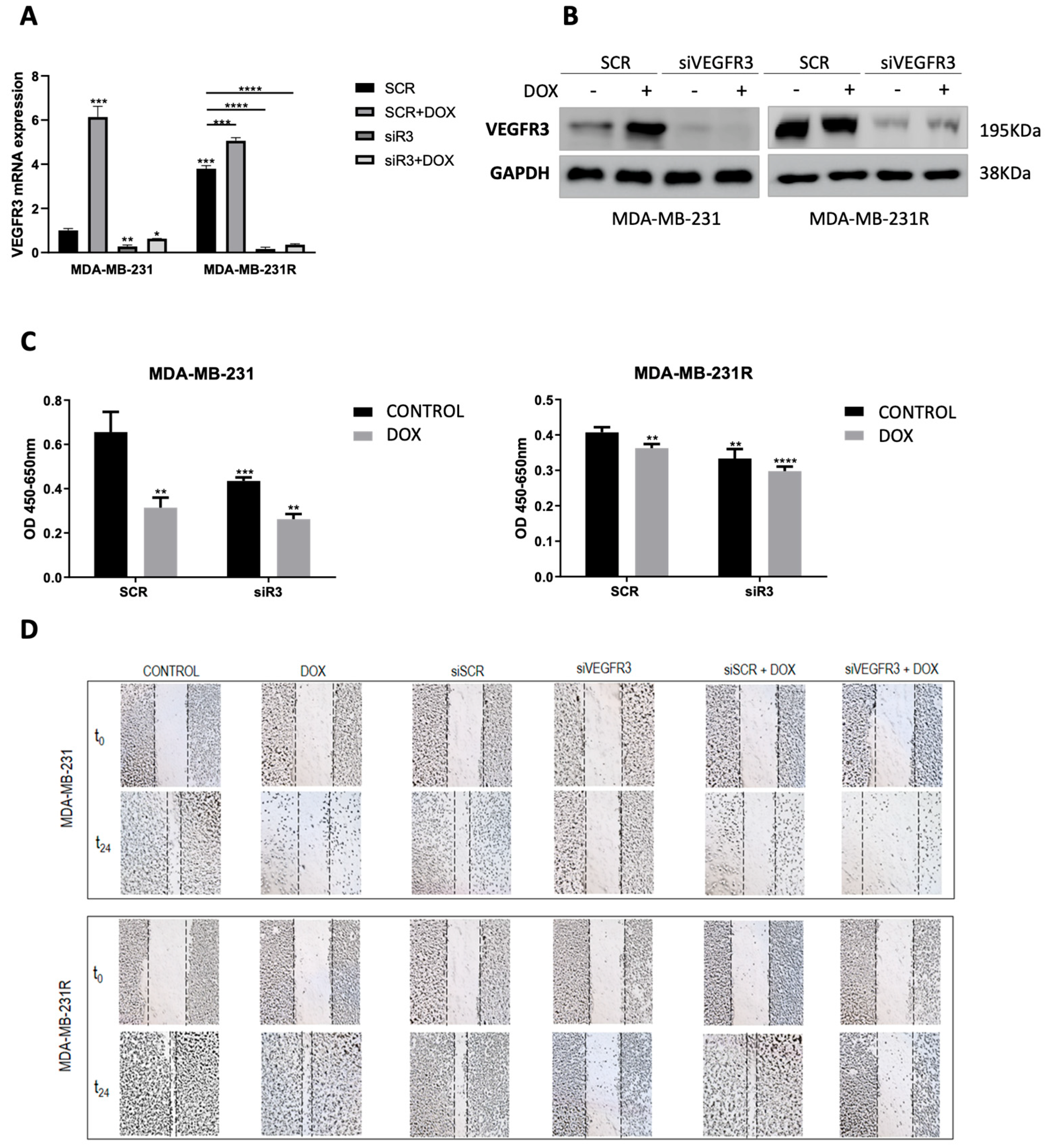
2.3. VEGFR3 Modulates Response to Doxorubicin in Both Sensitive and Resistant TNBC Cell Models
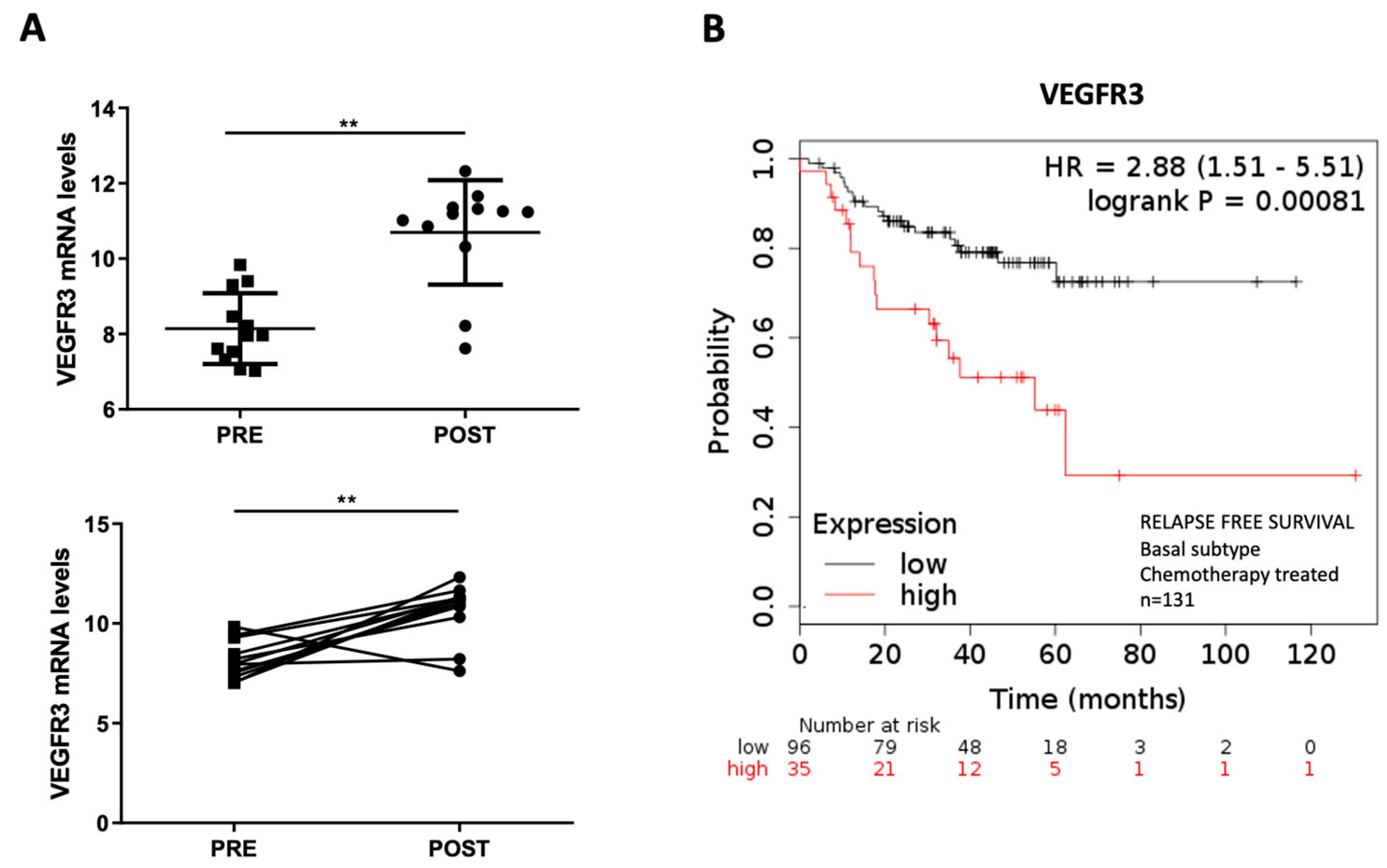
2.4. Overexpression of VEGFR3 as a Potential Prognosis Biomarker
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. MTT Proliferation Assay
4.3. Confocal Immunofluorescence Microscopy
4.4. Real-Time Quantitative PCR Analysis
4.5. Transient Transfection of siRNA
4.6. Wound-Healing Assay
4.7. Western Blot and Immunoprecipitation
4.8. Sample Patients
4.9. Survival Analysis
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohammed, R.A.A.; Ellis, I.O.; Mahmmod, A.M.; Hawkes, E.C.; Green, A.R.; Rakha, E.A.; Martin, S.G. Lymphatic and Blood Vessels in Basal and Triple-Negative Breast Cancers: Characteristics and Prognostic Significance. Mod. Pathol. 2011, 24, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Anders, C.K.; Zagar, T.M.; Carey, L.A. The Management of Early-Stage and Metastatic Triple-Negative Breast Cancer. Hematol./Oncol. Clin. N. Am. 2013, 27, 737–749. [Google Scholar] [CrossRef]
- Chavez, K.J.; Garimella, S.V.; Lipkowitz, S. Triple Negative Breast Cancer Cell Lines: One Tool in the Search for Better Treatment of Triple Negative Breast Cancer. Breast Dis. 2011, 32, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of Human Triple-Negative Breast Cancer Subtypes and Preclinical Models for Selection of Targeted Therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef]
- Ferrari, P.; Scatena, C.; Ghilli, M.; Bargagna, I.; Lorenzini, G.; Nicolini, A. Molecular Mechanisms, Biomarkers and Emerging Therapies for Chemotherapy Resistant TNBC. Int. J. Mol. Sci. 2022, 23, 1665. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, L.E.; Camorani, S.; Agnello, L.; Pedone, E.; Pirone, L.; Chesta, C.A.; Palacios, R.E.; Fedele, M.; Cerchia, L. Selective Photo-Assisted Eradication of Triple-Negative Breast Cancer Cells through Aptamer Decoration of Doped Conjugated Polymer Nanoparticles. Pharmaceutics 2022, 14, 626. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.-M.; Lee, K. Immune Checkpoint Blockades in Triple-Negative Breast Cancer: Current State and Molecular Mechanisms of Resistance. Biomedicines 2022, 10, 1130. [Google Scholar] [CrossRef] [PubMed]
- Raica, M.; Cimpean, A.M.; Ceausu, R.; Ribatti, D. Lymphatic Microvessel Density, VEGF-C, and VEGFR-3 Expression in Different Molecular Types of Breast Cancer. Anticancer Res. 2011, 31, 1757–1764. [Google Scholar]
- Wang, J.; Guo, Y.; Wang, B.; Bi, J.; Li, K.; Liang, X.; Chu, H.; Jiang, H. Lymphatic Microvessel Density and Vascular Endothelial Growth Factor-C and -D as Prognostic Factors in Breast Cancer: A Systematic Review and Meta-Analysis of the Literature. Mol. Biol. Rep. 2012, 39, 11153–11165. [Google Scholar] [CrossRef]
- Karpanen, T.; Alitalo, K. Lymphatic Vessels as Targets of Tumor Therapy? J. Exp. Med. 2001, 194, F37–F42. [Google Scholar] [CrossRef]
- Skobe, M.; Hawighorst, T.; Jackson, D.G.; Prevo, R.; Janes, L.; Velasco, P.; Riccardi, L.; Alitalo, K.; Claffey, K.; Detmar, M. Induction of Tumor Lymphangiogenesis by VEGF-C Promotes Breast Cancer Metastasis. Nat. Med. 2001, 7, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Su, J.-L.; Yen, C.-J.; Chen, P.-S.; Chuang, S.-E.; Hong, C.-C.; Kuo, I.-H.; Chen, H.-Y.; Hung, M.-C.; Kuo, M.-L. The Role of the VEGF-C/VEGFR-3 Axis in Cancer Progression. Br. J. Cancer 2007, 96, 541–545. [Google Scholar] [CrossRef]
- Kurenova, E.V.; Hunt, D.L.; He, D.; Fu, A.D.; Massoll, N.A.; Golubovskaya, V.M.; Garces, C.A.; Cance, W.G. Vascular Endothelial Growth Factor Receptor-3 Promotes Breast Cancer Cell Proliferation, Motility and Survival in Vitro and Tumor Formation in Vivo. Cell Cycle 2009, 8, 2266–2280. [Google Scholar] [CrossRef]
- Valtola, R.; Salven, P.; Heikkilä, P.; Taipale, J.; Joensuu, H.; Rehn, M.; Pihlajaniemi, T.; Weich, H.; deWaal, R.; Alitalo, K. VEGFR-3 and Its Ligand VEGF-C Are Associated with Angiogenesis in Breast Cancer. Am. J. Pathol. 1999, 154, 1381–1390. [Google Scholar] [CrossRef]
- Harris, A.R.; Perez, M.J.; Munson, J.M. Docetaxel Facilitates Lymphatic-Tumor Crosstalk to Promote Lymphangiogenesis and Cancer Progression. BMC Cancer 2018, 18, 718. [Google Scholar] [CrossRef] [PubMed]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the Fms-like Tyrosine Kinase 4 Gene Becomes Restricted to Lymphatic Endothelium during Development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3570. [Google Scholar] [CrossRef]
- Jussila, L.; Valtola, R.; Partanen, T.A.; Salven, P.; Heikkilä, P.; Matikainen, M.T.; Renkonen, R.; Kaipainen, A.; Detmar, M.; Tschachler, E.; et al. Lymphatic Endothelium and Kaposi’s Sarcoma Spindle Cells Detected by Antibodies against the Vascular Endothelial Growth Factor Receptor-3. Cancer Res. 1998, 58, 1599–1604. [Google Scholar] [PubMed]
- Lymboussaki, A.; Partanen, T.A.; Olofsson, B.; Thomas-Crusells, J.; Fletcher, C.D.M.; de Waal, R.M.W.; Kaipainen, A.; Alitalo, K. Expression of the Vascular Endothelial Growth Factor C Receptor VEGFR-3 in Lymphatic Endothelium of the Skin and in Vascular Tumors. Am. J. Pathol. 1998, 153, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Karroum, A.; Mirshahi, P.; Faussat, A.-M.; Therwath, A.; Mirshahi, M.; Hatmi, M. Tubular Network Formation by Adriamycin-Resistant MCF-7 Breast Cancer Cells Is Closely Linked to MMP-9 and VEGFR-2/VEGFR-3 over-Expressions. Eur. J. Pharmacol. 2012, 685, 1–7. [Google Scholar] [CrossRef]
- Zhang, P.; He, D.; Chen, Z.; Pan, Q.; Du, F.; Zang, X.; Wang, Y.; Tang, C.; Li, H.; Lu, H.; et al. Chemotherapy Enhances Tumor Vascularization via Notch Signaling-Mediated Formation of Tumor-Derived Endothelium in Breast Cancer. Biochem. Pharmacol. 2016, 118, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.A.A.; Green, A.; El-Shikh, S.; Paish, E.C.; Ellis, I.O.; Martin, S.G. Prognostic Significance of Vascular Endothelial Cell Growth Factors -A, -C and -D in Breast Cancer and Their Relationship with Angio- and Lymphangiogenesis. Br. J. Cancer 2007, 96, 1092–1100. [Google Scholar] [CrossRef]
- Van der Schaft, D.W.J.; Pauwels, P.; Hulsmans, S.; Zimmermann, M.; van de Poll-Franse, L.V.; Griffioen, A.W. Absence of Lymphangiogenesis in Ductal Breast Cancer at the Primary Tumor Site. Cancer Lett. 2007, 254, 128–136. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yasuoka, H.; Tsujimoto, M.; Yang, Q.; Imabun, S.; Nakahara, M.; Nakao, K.; Nakamura, M.; Mori, I.; Kakudo, K. Prognostic Significance of Vascular Endothelial Growth Factor D in Breast Carcinoma with Long-Term Follow-Up. Clin. Cancer Res. 2003, 9, 716–721. [Google Scholar]
- Dias, S.; Choy, M.; Alitalo, K.; Rafii, S. Vascular Endothelial Growth Factor (VEGF)–C Signaling through FLT-4 (VEGFR-3) Mediates Leukemic Cell Proliferation, Survival, and Resistance to Chemotherapy. Blood 2002, 99, 2179–2184. [Google Scholar] [CrossRef]
- Roberts, N.; Kloos, B.; Cassella, M.; Podgrabinska, S.; Persaud, K.; Wu, Y.; Pytowski, B.; Skobe, M. Inhibition of VEGFR-3 Activation with the Antagonistic Antibody More Potently Suppresses Lymph Node and Distant Metastases than Inactivation of VEGFR-2. Cancer Res. 2006, 66, 2650–2657. [Google Scholar] [CrossRef]
- Walsh, K.A.; Kastrappis, G.; Fifis, T.; Paolini, R.; Christophi, C.; Perini, M.V. SAR131675, a VEGRF3 Inhibitor, Modulates the Immune Response and Reduces the Growth of Colorectal Cancer Liver Metastasis. Cancers 2022, 14, 2715. [Google Scholar] [CrossRef]
- Shibata, M.; Shibata, E.; Tanaka, Y.; Shiraoka, C.; Kondo, Y. Soluble Vegfr3 Gene Therapy Suppresses Multi-organ Metastasis in a Mouse Mammary Cancer Model. Cancer Sci. 2020, 111, 2837–2849. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F.; González-Navarrete, I.; Bragado, R.; Dalmases, A.; Menéndez, S.; Cortes-Sempere, M.; Suárez, C.; Oliva, C.; Servitja, S.; Rodriguez-Fanjul, V.; et al. Mitogen-Activated Protein Kinase Phosphatase-1 in Human Breast Cancer Independently Predicts Prognosis and Is Repressed by Doxorubicin. Clin. Cancer Res. 2009, 15, 3530–3539. [Google Scholar] [CrossRef] [PubMed]
- Tormo, E.; Ballester, S.; Adam-Artigues, A.; Burgués, O.; Alonso, E.; Bermejo, B.; Menéndez, S.; Zazo, S.; Madoz-Gúrpide, J.; Rovira, A.; et al. The MiRNA-449 Family Mediates Doxorubicin Resistance in Triple-Negative Breast Cancer by Regulating Cell Cycle Factors. Sci. Rep. 2019, 9, 5316. [Google Scholar] [CrossRef] [PubMed]
- Tapia, M.A.; González-Navarrete, I.; Dalmases, A.; Bosch, M.; Rodriguez-Fanjul, V.; Rolfe, M.; Ross, J.S.; Mezquita, J.; Mezquita, C.; Bachs, O.; et al. Inhibition of the Canonical IKK/NFΚB Pathway Sensitizes Human Cancer Cells to Doxorubicin. Cell Cycle 2007, 6, 2284–2292. [Google Scholar] [CrossRef] [PubMed]
- Dalmases, A.; González, I.; Menendez, S.; Arpí, O.; Corominas, J.M.; Servitja, S.; Tusquets, I.; Chamizo, C.; Rincón, R.; Espinosa, L.; et al. Deficiency in P53 Is Required for Doxorubicin Induced Transcriptional Activation of NF-ΚB Target Genes in Human Breast Cancer. Oncotarget 2014, 5, 196–210. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | Subtype | Diag. Age | Diagnosis | Grade | RCB | Chemotherapy |
|---|---|---|---|---|---|---|
| n1 | TN | 41 | ICD | II | 1 | Taxolx12+FEC-100x4 |
| n2 | TN | 70 | ICD | III | 1 | Taxolx12+FEC-100x4 |
| n3 | TN | 78 | ICD | III | 1 | Atx4+CMFx4 |
| n4 | TN | 68 | ICD | II | 1 | Atx4+CMFx4 |
| n5 | TN | 42 | ICD | II | 2 | Atx4+CMFx4 |
| n6 | TN | 64 | ICD | II | 2 | Taxolx12+FEC-100x4 |
| n7 | TN | 43 | ICD | III | 2 | Taxolx12+FEC-100x4 |
| n8 | TN | 45 | ICD | II | 2 | Atx4+CMFx4 |
| n9 | TN | 52 | ICD | II | 3 | Taxolx12+FEC-100x4 |
| n10 | TN | 46 | ICD | III | 3 | Taxolx12+FEC-100x4 |
| n11 | TN | 65 | ILC | II | 3 | Atx4+CMFx4 |
| n12 | TN | 52 | ICD | II | 3 | Taxolx12+FEC-100x4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Ruiz, S.; Tormo, E.; Garrido-Cano, I.; Lameirinhas, A.; Rojo, F.; Madoz-Gúrpide, J.; Burgués, O.; Hernando, C.; Bermejo, B.; Martínez, M.T.; et al. High VEGFR3 Expression Reduces Doxorubicin Efficacy in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2023, 24, 3601. https://doi.org/10.3390/ijms24043601
Torres-Ruiz S, Tormo E, Garrido-Cano I, Lameirinhas A, Rojo F, Madoz-Gúrpide J, Burgués O, Hernando C, Bermejo B, Martínez MT, et al. High VEGFR3 Expression Reduces Doxorubicin Efficacy in Triple-Negative Breast Cancer. International Journal of Molecular Sciences. 2023; 24(4):3601. https://doi.org/10.3390/ijms24043601
Chicago/Turabian StyleTorres-Ruiz, Sandra, Eduardo Tormo, Iris Garrido-Cano, Ana Lameirinhas, Federico Rojo, Juan Madoz-Gúrpide, Octavio Burgués, Cristina Hernando, Begoña Bermejo, María Teresa Martínez, and et al. 2023. "High VEGFR3 Expression Reduces Doxorubicin Efficacy in Triple-Negative Breast Cancer" International Journal of Molecular Sciences 24, no. 4: 3601. https://doi.org/10.3390/ijms24043601
APA StyleTorres-Ruiz, S., Tormo, E., Garrido-Cano, I., Lameirinhas, A., Rojo, F., Madoz-Gúrpide, J., Burgués, O., Hernando, C., Bermejo, B., Martínez, M. T., Lluch, A., Cejalvo, J. M., & Eroles, P. (2023). High VEGFR3 Expression Reduces Doxorubicin Efficacy in Triple-Negative Breast Cancer. International Journal of Molecular Sciences, 24(4), 3601. https://doi.org/10.3390/ijms24043601