

Circadian Rhythms Disrupted by Light at Night and Mistimed Food Intake Alter Hormonal Rhythms and Metabolism


Abstract

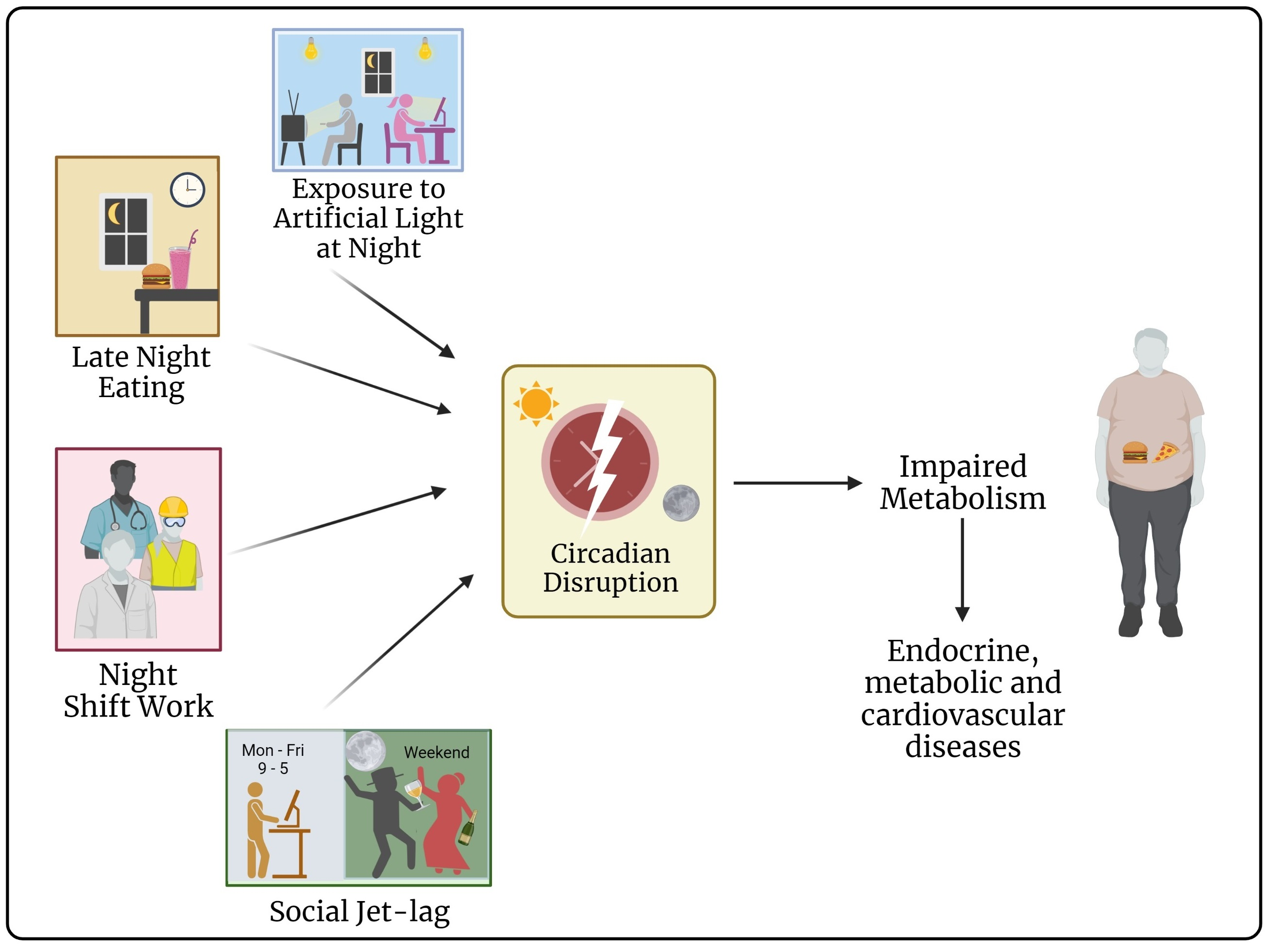{kind=link}
{kind=link}
1. Introduction
2. Circadian Rhythms and the Molecular Clock
2.1. Phototransduction to the SCN
2.2. SCN Signaling
3. Hypothalamic Control of Circadian Homeostasis and Hormone Regulation
3.1. Food Intake
3.2. Sleep
3.3. SCN-Mediated Hormonal Release and Function
3.3.1. Melatonin
3.3.2. Glucocorticoids
4. Changing Environment and Consequences on Hormonal Rhythms
4.1. Artificial Light at Night
4.2. Shift Work and Jet Lag
4.3. Mistimed Food Intake
5. Changing Environments, Misaligned Circadian Rhythms, and Resulting Disorders
Endocrine, Metabolic, and Cardiovascular Diseases
6. Strategies to Remediate Effects on Disrupted Rhythms in Humans
6.1. Dark Nights
6.2. Decrease Blue Light Exposure in the Evening
6.3. Early Morning Bright Light
6.4. Food Restriction to the Active Phase
6.5. Melatonin Supplementation
7. Recommendations for Night Shift Workers and Future Work
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Genet. 2005, 6, 544–556. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A. Common features of circadian timekeeping in diverse organisms. Curr. Opin. Physiol. 2018, 5, 58–67. [Google Scholar] [CrossRef]
- Wright, K.P., Jr.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [PubMed]
- Roenneberg, T.; Merrow, M. The Circadian Clock and Human Health. Curr. Biol. 2016, 26, R432–R443. [Google Scholar] [CrossRef] [PubMed]
- Bonnell, E.K.; Huggins, C.E.; Huggins, C.T.; McCaffrey, T.A.; Palermo, C.; Bonham, M.P. Influences on Dietary Choices during Day versus Night Shift in Shift Workers: A Mixed Methods Study. Nutrients 2017, 9, 193. [Google Scholar] [CrossRef]
- Gan, Y.; Yang, C.; Tong, X.; Sun, H.; Cong, Y.; Yin, X.; Li, L.; Cao, S.; Dong, X.; Gong, Y.; et al. Shift work and diabetes mellitus: A meta-analysis of observational studies. Occup. Environ. Med. 2014, 72, 72–78. [Google Scholar] [CrossRef]
- Rose, D.M.; Jung, D.; Parera, D.; Konietzko, J. Time zone shifts and jet lag after long-distance flights. Z Arztl Fortbild Qualitatssich. 1999, 93, 485–490. [Google Scholar]
- Sack, R.L. The pathophysiology of jet lag. Travel Med. Infect. Dis. 2009, 7, 102–110. [Google Scholar] [CrossRef]
- Nagano, M.; Adachi, A.; Nakahama, K.-I.; Nakamura, T.; Tamada, M.; Meyer-Bernstein, E.; Sehgal, A.; Shigeyoshi, Y. An Abrupt Shift in the Day/Night Cycle Causes Desynchrony in the Mammalian Circadian Center. J. Neurosci. 2003, 23, 6141–6151. [Google Scholar] [CrossRef] [PubMed]
- Mortola, J.P. Breathing around the clock: An overview of the circadian pattern of respiration. Eur. J. Appl. Physiol. 2004, 91, 119–129. [Google Scholar] [CrossRef]
- Kawano, Y. Diurnal blood pressure variation and related behavioral factors. Hypertens. Res. 2010, 34, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Vandewalle, G.; Middleton, B.; Rajaratnam, S.M.W.; Stone, B.M.; Thorleifsdottir, B.; Arendt, J.; Dijk, D. Robust CircadianRhythm in Heart Rate and Its Variability: Influence of Exogenous Melatonin and Photoperiod. J. Sleep Res. 2007, 16, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Fernández, O.; Walton, J.; DeVries, A.; Nelson, R. Clocks, Rhythms, Sex, and Hearts: How Disrupted Circadian Rhythms, Time-of-Day, and Sex Influence Cardiovascular Health. Biomolecules 2021, 11, 883. [Google Scholar] [CrossRef]
- Chellappa, S.; Vujovic, N.; Williams, J.S.; Scheer, F.A. Impact of Circadian Disruption on Cardiovascular Function and Disease. Trends Endocrinol. Metab. 2019, 30, 767–779. [Google Scholar] [CrossRef]
- Bedrosian, T.A.; Fonken, L.K.; Nelson, R.J. Endocrine Effects of Circadian Disruption. Annu. Rev. Physiol. 2016, 78, 109–131. [Google Scholar] [CrossRef]
- Walker, W.H.; Borniger, J.C.; Gaudier-Diaz, M.M.; Meléndez-Fernández, O.H.; Pascoe, J.L.; Devries, A.C.; Nelson, R.J. Acute exposure to low-level light at night is sufficient to induce neurological changes and depressive-like behavior. Mol. Psychiatry 2020, 25, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Nelson, R.J. The effects of light at night on circadian clocks and metabolism. Endocr. Rev. 2014, 35, 648–670. [Google Scholar] [CrossRef]
- Walker, W.H.; Bumgarner, J.R.; Walton, J.C.; Liu, J.A.; Meléndez-Fernández, O.H.; Nelson, R.J.; Devries, A.C. Light Pollution and Cancer. Int. J. Mol. Sci. 2020, 21, 9360. [Google Scholar] [CrossRef]
- Klepeis, N.E.; Nelson, W.C.; Ott, W.R.; Robinson, J.P.; Tsang, A.M.; Switzer, P.; Behar, J.V.; Hern, S.C.; Engelmann, W.H. The National Human Activity Pattern Survey (NHAPS): A resource for assessing exposure to environmental pollutants. J. Expo. Sci. Environ. Epidemiol. 2001, 11, 231–252. [Google Scholar] [CrossRef]
- Bhadra, U.; Thakkar, N.; Das, P.; Bhadra, M.P. Evolution of circadian rhythms: From bacteria to human. Sleep Med. 2017, 35, 49–61. [Google Scholar] [CrossRef]
- Vaze, K.M.; Sharma, V.K. On the Adaptive Significance of Circadian Clocks for Their Owners. Chrono Int. 2013, 30, 413–433. [Google Scholar] [CrossRef]
- Refinetti, R. Comparison of light, food, and temperature as environmental synchronizers of the circadian rhythm of activity in mice. J. Physiol. Sci. 2015, 65, 359–366. [Google Scholar] [CrossRef]
- Wever, R.A. Light Effects on Human Circadian Rhythms: A Review of Recent Andechs Experiments. J. Biol. Rhythm. 1989, 4, 49–73. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kumar, C.J.; Merrow, M. The human circadian clock entrains to sun time. Curr. Biol. 2007, 17, R44–R45. [Google Scholar] [CrossRef]
- Roenneberg, T.; Foster, R.G. Twilight Times: Light and the Circadian System. Photochem. Photobiol. 1997, 66, 549–561. [Google Scholar] [CrossRef]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164–179. [Google Scholar] [CrossRef]
- Fang, B.; Everett, L.J.; Jager, J.; Briggs, E.; Armour, S.M.; Feng, D.; Roy, A.; Gerhart-Hines, Z.; Sun, Z.; Lazar, M.A. Circadian Enhancers Coordinate Multiple Phases of Rhythmic Gene Transcription In Vivo. Cell 2014, 159, 1140–1152. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, W.; Li, P.; Zhang, Y.; Zhao, M.; Fan, Z.; Zhao, Z.; Yan, J. Long-Range Chromosome Interactions Mediated by Cohesin Shape Circadian Gene Expression. PLOS Genet. 2016, 12, e1005992. [Google Scholar] [CrossRef]
- Arnal, L.A.; Hakim, O.; Patel, V.R.; Baldi, P.; Hager, G.L.; Sassone-Corsi, P. Cycles in spatial and temporal chromosomal organization driven by the circadian clock. Nat. Struct. Mol. Biol. 2013, 20, 1206–1213. [Google Scholar] [CrossRef]
- Kojima, S.; Shingle, D.L.; Green, C.B. Post-transcriptional control of circadian rhythms. J. Cell Sci. 2011, 124, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Baker, C.L.; Loros, J.J.; Dunlap, J.C. Post-translational modifications in circadian rhythms. Trends Biochem. Sci. 2009, 34, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Erzberger, A.; Hampp, G.; Granada, A.; Albrecht, U.; Herzel, H. Genetic redundancy strengthens the circadian clock leading to a narrow entrainment range. J. R. Soc. Interface 2013, 10, 20130221. [Google Scholar] [CrossRef] [PubMed]
- Oster, H.; Baeriswyl, S.; van der Horst, G.T.; Albrecht, U. Loss of circadian rhythmicity in aging mPer1-/-mCry2-/- mutant mice. Genes Dev. 2003, 17, 1366–1379. [Google Scholar] [CrossRef]
- Oster, H.; Yasui, A.; van der Horst, G.T.; Albrecht, U. Disruption of mCry2 restores circadian rhythmicity in mPer2 mutant mice. Genes Dev. 2002, 16, 2633–2638. [Google Scholar] [CrossRef] [PubMed]
- Welsh, D.K.; Takahashi, J.S.; Kay, S.A. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2010, 72, 551–577. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.; Oster, H.; Korf, H.W.; Foster, R.G.; Erren, T.C. Food as a circadian time Cue-Evidence from human studies. Nat. Rev. Endocrinol. 2020, 16, 213–223. [Google Scholar] [PubMed]
- Lewis, P.; Korf, H.; Kuffer, L.; Groß, J.V.; Erren, T.C. Exercise time cues (zeitgebers) for human circadian systems can foster health and improve performance: A systematic review. BMJ Open Sport Exerc. Med. 2018, 4, e000443. [Google Scholar]
- Stephan, F.K. The "Other" Circadian System: Food as a Zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar]
- Yamazaki, S.; Goto, M.; Menaker, M. No Evidence for Extraocular Photoreceptors in the Circadian System of the Syrian Hamster. J. Biol. Rhythm. 1999, 14, 197–201. [Google Scholar] [CrossRef]
- Ibata, Y.; Takahashi, Y.; Okamura, H.; Kawakami, F.; Terubayashi, H.; Kubo, T.; Yanaihara, N. Vasoactive intestinal peptide (VIP)-like immunoreactive neurons located in the rat suprachiasmatic nucleus receive a direct retinal projection. Neurosci. Lett. 1989, 97, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y.; Speh, J.C.; Card, J.P. The retinohypothalamic tract originates from a distinct subset of retinal ganglion cells. J. Comp. Neurol. 1995, 352, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y.; Lenn, N.J. A retinohypothalamic projection in the rat. J. Comp. Neurol. 1972, 146, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Chen, S.-K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef]
- Schmidt, T.M.; Do, M.T.H.; Dacey, D.; Lucas, R.; Hattar, S.; Matynia, A.; Matynia, A. Melanopsin-Positive Intrinsically Photosensitive Retinal Ganglion Cells: From Form to Function. J. Neurosci. 2011, 31, 16094–16101. [Google Scholar] [CrossRef]
- Smith, M.R.; Revell, V.L.; Eastman, C.I. Phase advancing the human circadian clock with blue-enriched polychromatic light. Sleep Med. 2009, 10, 287–294. [Google Scholar] [CrossRef]
- Smith, M.R.; Eastman, C.I. Phase Delaying the Human Circadian Clock with Blue-Enriched Polychromatic Light. Chrono Int. 2009, 26, 709–725. [Google Scholar] [CrossRef]
- Wahl, S.; Engelhardt, M.; Schaupp, P.; Lappe, C.; Ivanov, I.V. The inner clock—Blue light sets the human rhythm. J. Biophotonics 2019, 12, e201900102. [Google Scholar]
- Tähkämö, L.; Partonen, T.; Pesonen, A.-K. Systematic review of light exposure impact on human circadian rhythm. Chronobiol. Int. 2019, 36, 151–170. [Google Scholar] [CrossRef]
- Lewy, A.J.; Wehr, T.A.; Goodwin, F.K.; Newsome, D.A.; Markey, S.P. Light Suppresses Melatonin Secretion in Humans. Science 1980, 210, 1267–1269. [Google Scholar] [CrossRef]
- Cajochen, C.; Münch, M.; Kobialka, S.; Kräuchi, K.; Steiner, R.; Oelhafen, P.; Orgül, S.; Wirz-Justice, A. High Sensitivity of Human Melatonin, Alertness, Thermoregulation, and Heart Rate to Short Wavelength Light. J. Clin. Endocrinol. Metab. 2005, 90, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Sato, T.K.; Castrucci, A.M.; Rollag, M.D.; DeGrip, W.J.; Hogenesch, J.B.; Provencio, I.; Kay, S.A. Melanopsin ( Opn4 ) Requirement for Normal Light-Induced Circadian Phase Shifting. Science 2002, 298, 2213–2216. [Google Scholar] [CrossRef] [PubMed]
- Ruby, N.F.; Brennan, T.J.; Xie, X.; Cao, V.; Franken, P.; Heller, H.C.; O’Hara, B.F. Role of Melanopsin in Circadian Responses to Light. Science 2002, 298, 2211–2213. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Provencio, I.; Tu, D.C.; Pires, S.S.; Rollag, M.D.; Castrucci, A.M.; Pletcher, M.T.; Sato, T.K.; Wiltshire, T.; Andahazy, M.; et al. Melanopsin Is Required for Non-Image-Forming Photic Responses in Blind Mice. Science 2003, 301, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Güler, A.D.; Ecker, J.L.; Lall, G.S.; Haq, S.; Altimus, C.M.; Liao, H.-W.; Barnard, A.R.; Cahill, H.; Badea, T.C.; Zhao, H.; et al. Melanopsin cells are the principal conduits for rod–cone input to non-image-forming vision. Nature 2008, 453, 102–105. [Google Scholar]
- Stopa, E.G.; Johnson, J.K.; I Friedman, D.; I Ryer, H.; Reidy, J.; Kuo-Leblanc, V.; E Albers, H. Neuropeptide Y receptor distribution and regulation in the suprachiasmatic nucleus of the Syrian hamster (Mesocricetus auratus). Pept. Res. 1995, 8, 95–100. [Google Scholar] [PubMed]
- Golombek, D.A.; Biello, S.M.; Rendon, R.A.; Harrington, M.E. Neuropeptide Y phase shifts the circadian clock in vitro via a Y2 receptor. Neuroreport 1996, 7, 1315–1319. [Google Scholar] [CrossRef]
- Huhman, K.; Gillespie, C.; Marvel, C.; Albers, H.E. Neuropeptide Y phase shifts circadian rhythms in vivo via a Y2 receptor. Neuroreport 1996, 7, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.A.; Dhandapani, K.M.; Goldman, B.D. The thalamic intergeniculate leaflet modulates photoperiod responsiveness in Siberian hamsters. Brain Res. 2004, 1028, 31–38. [Google Scholar] [CrossRef]
- Weber, E.; A Rea, M. Neuropeptide Y blocks light-induced phase advances but not delays of the circadian activity rhythm in hamsters. Neurosci. Lett. 1997, 231, 159–162. [Google Scholar] [CrossRef]
- Singh, C.; Rihel, J.; Prober, D.A. Neuropeptide Y Regulates Sleep by Modulating Noradrenergic Signaling. Curr. Biol. 2017, 27, 3796–3811. [Google Scholar] [CrossRef]
- Fu, L.-Y.; Acuna-Goycolea, C.; Pol, A.N.V.D. Neuropeptide Y Inhibits Hypocretin/Orexin Neurons by Multiple Presynaptic and Postsynaptic Mechanisms: Tonic Depression of the Hypothalamic Arousal System. J. Neurosci. 2004, 24, 8741–8751. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic nucleus in the mouse: Retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef]
- Swanson, L.W.; Cowan, W.M. The efferent connections of the suprachiasmatic nucleus of the hypothalamus. J. Comp. Neurol. 1975, 160, 1–12. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Leak, R.K.; Yackulic, C.B.; LeSauter, J.; Silver, R. Organization of suprachiasmatic nucleus projections in Syrian hamsters (Mesocricetus auratus): An anterograde and retrograde analysis. J. Comp. Neurol. 2003, 468, 361–379. [Google Scholar] [CrossRef]
- Vujovic, N.; Gooley, J.J.; Jhou, T.C.; Saper, C.B. Projections from the subparaventricular zone define four channels of output from the circadian timing system. J. Comp. Neurol. 2015, 523, 2714–2737. [Google Scholar] [CrossRef]
- Leibowitz, S.F. Reciprocal Hunger-Regulating Circuits Involving Alpha-and Beta-Adrenergic Receptors Located, Respectively, in the Ventromedial and Lateral Hypothalamus. Proc. Natl. Acad. Sci. USA 1970, 67, 1063–1070. [Google Scholar] [CrossRef]
- Berthoud, H.-R.; Münzberg, H. The lateral hypothalamus as integrator of metabolic and environmental needs: From electrical self-stimulation to opto-genetics. Physiol. Behav. 2011, 104, 29–39. [Google Scholar] [CrossRef]
- Arrigoni, E.; Chee, M.J.; Fuller, P.M. To eat or to sleep: That is a lateral hypothalamic question. Neuropharmacology 2018, 154, 34–49. [Google Scholar]
- Lu, J.; Zhang, Y.-H.; Chou, T.C.; Gaus, S.E.; Elmquist, J.K.; Shiromani, P.; Saper, C.B. Contrasting Effects of Ibotenate Lesions of the Paraventricular Nucleus and Subparaventricular Zone on Sleep–Wake Cycle and Temperature Regulation. J. Neurosci. 2001, 21, 4864–4874. [Google Scholar] [CrossRef]
- Asala, S.A.; Okano, Y.; Honda, K.; Inoué, S. Effects of medial preoptic area lesions on sleep and wakefulness in unrestrained rats. Neurosci. Lett. 1990, 114, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Mondino, A.; Hambrecht-Wiedbusch, V.S.; Li, D.; York, A.K.; Pal, D.; González, J.; Torterolo, P.; Mashour, G.A.; Vanini, G. Glutamatergic Neurons in the Preoptic Hypothalamus Promote Wakefulness, Destabilize NREM Sleep, Suppress REM Sleep, and Regulate Cortical Dynamics. J Neurosci. 2021, 41, 3462–3478. [Google Scholar] [CrossRef] [PubMed]
- Hiller-Sturmhöfel, S.; Bartke, A. The Endocrine System: An Overview. Alcohol Health Res. Alcohol Health Res World 1998, 22, 153–164. [Google Scholar]
- Yousefvand, S.; Hamidi, F. The Role of Ventromedial Hypothalamus Receptors in the Central Regulation of Food Intake. Int. J. Pept. Res. Ther. 2020, 27, 689–702. [Google Scholar] [CrossRef]
- Sternson, S.; Shepherd, G.M.G.; Friedman, J.M. Topographic mapping of VMH → arcuate nucleus microcircuits and their reorganization by fasting. Nat. Neurosci. 2005, 8, 1356–1363. [Google Scholar]
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Hashiguchi, H.; Sheng, Z.; Routh, V.; Gerzanich, V.; Simard, J.M.; Bryan, J. Direct versus indirect actions of ghrelin on hypothalamic NPY neurons. PLoS ONE 2017, 12, e0184261. [Google Scholar] [CrossRef]
- Barson, J.R.; Leibowitz, S.F. Orexin/Hypocretin System: Role in Food and Drug Overconsumption. Int. Rev. Neurobiol. 2017, 136, 199–237. [Google Scholar] [CrossRef]
- Zielinski, M.R.; Krueger, J.M. Sleep and innate immunity. Front. Biosci. 2011, S3, 632–642. [Google Scholar] [CrossRef]
- Zielinski, M.R.; McKenna, J.T.; McCarley, R.W. Functions and Mechanisms of Sleep. AIMS Neurosci. 2016, 3, 67–104. [Google Scholar] [CrossRef]
- Szymusiak, R.; Gvilia, I.; McGinty, D. Hypothalamic control of sleep. Sleep Med. 2007, 8, 291–301. [Google Scholar] [CrossRef]
- Cedernaes, J.; Waldeck, N.; Bass, J. Neurogenetic basis for circadian regulation of metabolism by the hypothalamus. Genes Dev. 2019, 33, 1136–1158. [Google Scholar] [CrossRef]
- Brzezinski, A.; Vangel, M.G.; Wurtman, R.J.; Norrie, G.; Zhdanova, I.; Ben-Shushan, A.; Ford, I. Effects of exogenous melatonin on sleep: A meta-analysis. Sleep Med. Rev. 2005, 9, 41–50. [Google Scholar] [CrossRef]
- Zhdanova, I.V. Melatonin as a hypnotic: Pro. Sleep Med. Rev. 2005, 9, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.V.; Mosser, E.A.; Oikonomou, G.; Prober, D.A. Melatonin Is Required for the Circadian Regulation of Sleep. Neuron 2015, 85, 1193–1199. [Google Scholar] [CrossRef]
- Zhdanova, I.V.; Geiger, D.A.; Schwagerl, A.L.; Leclair, O.U.; Killiany, R.; Taylor, J.A.; Rosene, D.L.; Moss, M.B.; Madras, B.K. Melatonin promotes sleep in three species of diurnal nonhuman primates. Physiol. Behav. 2002, 75, 523–529. [Google Scholar] [CrossRef]
- Goldstein, R.; Pavel, S. REM sleep suppression in cats by melatonin. Brain Res. Bull. 1981, 7, 723–724. [Google Scholar] [CrossRef]
- Mintz, E.M.; Phillips, N.H.; Berger, R.J. Daytime melatonin infusions induce sleep in pigeons without altering subsequent amounts of nocturnal sleep. Neurosci. Lett. 1998, 258, 61–64. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Garidou, M.-L.; Palm, I.F.; Van Der Vliet, J.; Simonneaux, V.; Pévet, P.; Buijs, R.M. Melatonin sees the light: Blocking GABA-ergic transmission in the paraventricular nucleus induces daytime secretion of melatonin. Eur. J. Neurosci. 2000, 12, 3146–3154. [Google Scholar]
- Klein, D.C. Arylalkylamine N-Acetyltransferase: “The Timezyme”. J. Biol. Chem. 2007, 282, 4233–4237. [Google Scholar]
- Chattoraj, A.; Liu, T.; Zhang, L.S.; Huang, Z.; Borjigin, J. Melatonin formation in mammals: In vivo perspectives. Rev. Endocr. Metab. Disord. 2009, 10, 237–243. [Google Scholar] [CrossRef]
- Macchi, M.M.; Bruce, J.N. Human pineal physiology and functional significance of melatonin. Front. Neuroendocr. 2004, 25, 177–195. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron 1994, 13, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Godson, C.; Mahle, C.D.; Weaver, D.R.; A Slaugenhaupt, S.; Gusella, J.F. Molecular characterization of a second melatonin receptor expressed in human retina and brain: The Mel1b melatonin receptor. Proc. Natl. Acad. Sci. USA 1995, 92, 8734–8738. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Rivera-Bermudez, M.A.; Gerdin, M.J.; Masana, M.I. Molecular pharmacology regulation and function of mammalian melatonin receptors. Front. Biosci. 2003, 8, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Weaver, D.R.; Jin, X.; Shearman, L.P.; Pieschl, R.L.; Gribkoff, V.K.; Reppert, S.M. Molecular Dissection of Two Distinct Actions of Melatonin on the Suprachiasmatic Circadian Clock. Neuron 1997, 19, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.A.; Allan, J.S.; Strogatz, S.H.; Ronda, J.M.; Sánchez, R.; Ríos, C.D.; Freitag, W.O.; Richardson, G.S.; Kronauer, R.E. Bright Light Resets the Human Circadian Pacemaker Independent of the Timing of the Sleep-Wake Cycle. Science 1986, 233, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Lewy, A.J.; Menaker, M.; Fredrickson, R.H.; Miller, L.S.; Weleber, R.G.; Cassone, V.; Hudson, D. Dose-response relationship between light irradiance and the suppression of plasma melatonin in human volunteers. Brain Res. 1988, 454, 212–218. [Google Scholar]
- Slotten, H.A.; Krekling, S.; Sicard, B.; Pévet, P. Daily infusion of melatonin entrains circadian activity rhythms in the diurnal rodent Arvicanthis ansorgei. Behav. Brain Res. 2001, 133, 11–19. [Google Scholar] [CrossRef]
- Pévet, P.; Bothorel, B.; Slotten, H.; Saboureau, M. The chronobiotic properties of melatonin. Cell Tissue Res. 2002, 309, 183–191. [Google Scholar] [CrossRef]
- Slotten, H.A.; Pitrosky, B.; Krekling, S.; Pévet, P. Entrainment of circadian activity rhythms in rats to melatonin administered at T cycles different from 24 hours. Neurosignals 2002, 11, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Mao, L.; Duplessis, T.; Yuan, L.; Dauchy, R.; Dauchy, E.; Blask, D.; Frasch, T.; Hill, S. Oscillation of Clock and Clock Controlled Genes Induced by Serum Shock in Human Breast Epithelial and Breast Cancer Cells: Regulation by Melatonin. Breast Cancer Basic Clin. Res. 2012, 6, 137–150. [Google Scholar] [CrossRef]
- Pechanova, O.; Paulis, L.; Simko, F. Peripheral and Central Effects of Melatonin on Blood Pressure Regulation. Int. J. Mol. Sci. 2014, 15, 17920–17937. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Dardente, H.; Menet, J.S.; Poirel, V.-J.; Streicher, D.; Gauer, F.; Vivien-Roels, B.; Klosen, P.; Pévet, P.; Masson-Pévet, M. Melatonin induces Cry1 expression in the pars tuberalis of the rat. Mol. Brain Res. 2003, 114, 101–106. [Google Scholar] [CrossRef] [PubMed]
- von Gall, C.; Weaver, D.R.; Moek, J.; Jilg, A.; Stehle, J.H.; Korf, H.W. Melatonin plays a crucial role in the regulation of rhythmic clock gene expression in the mouse pars tuberalis. Ann. New York Acad. Sci. 2005, 1040, 508–511. [Google Scholar]
- Messager, S.; Garabette, M.L.; Hastings, M.H.; Hazlerigg, D.G. Tissue-specific abolition of Per1 expression in the pars tuberalis by pinealectomy in the Syrian hamster. Neuroreport 2001, 12, 579–582. [Google Scholar] [CrossRef]
- Agez, L.; Laurent, V.; Guerrero, H.Y.; Pévet, P.; Masson-Pévet, M.; Gauer, F. Endogenous melatonin provides an effective circadian message to both the suprachiasmatic nuclei and the pars tuberalis of the rat. J. Pineal Res. 2009, 46, 95–105. [Google Scholar] [CrossRef]
- Lockley, S.W.; Skene, D.J.; James, K.; Thapan, K.; Wright, J.; Arendt, J. Melatonin administration can entrain the free-running circadian system of blind subjects. J. Endocrinol. 2000, 164, R1–R6. [Google Scholar] [CrossRef]
- Sack, R.L.; Brandes, R.W.; Kendall, A.R.; Lewy, A.J. Entrainment of Free-Running Circadian Rhythms by Melatonin in Blind People. N. Engl. J. Med. 2000, 343, 1070–1077. [Google Scholar] [CrossRef]
- Lewy, A.J.; Bauer, V.K.; Hasler, B.P.; Kendall, A.R.; Pires, M.N.; Sack, R.L. Capturing the circadian rhythms of free-running blind people with 0.5 mg melatonin. Brain Res. 2001, 918, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Kazimi, N.; Cahill, G.M. Development of a circadian melatonin rhythm in embryonic zebrafish. Dev. Brain Res. 1999, 117, 47–52. [Google Scholar] [CrossRef]
- Lewy, A.J.; Ahmed, S.; Jackson, J.M.L.; Sack, R.L. Melatonin Shifts Human Orcadian Rhythms According to a Phase-Response Curve. Chrono Int. 1992, 9, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Turek, F.; Gillette, M. Melatonin, sleep, and circadian rhythms: Rationale for development of specific melatonin agonists. Sleep Med. 2004, 5, 523–532. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef]
- Dijk, D.-J.; Cajochen, C. Melatonin and the Circadian Regulation of Sleep Initiation, Consolidation, Structure, and the Sleep EEG. J. Biol. Rhythm. 1997, 12, 627–635. [Google Scholar] [CrossRef]
- Lavie, P. Melatonin: Role in Gating Nocturnal Rise in Sleep Propensity. J. Biol. Rhythm. 1997, 12, 657–665. [Google Scholar] [CrossRef]
- Srinivasan, V.; Pandi-Perumal, S.R.; Trahkt, I.; Spence, D.W.; Poeggeler, B.; Hardeland, R.; Cardinali, D.P. Melatonin and Melatonergic Drugs on Sleep: Possible Mechanisms of Action. Int. J. Neurosci. 2009, 119, 821–846. [Google Scholar] [CrossRef]
- Williams, W.P.; McLin, D.E.; Dressman, M.A.; Neubauer, D.N. Comparative Review of Approved Melatonin Agonists for the Treatment of Circadian Rhythm Sleep-Wake Disorders. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2016, 36, 1028–1041. [Google Scholar] [CrossRef]
- Lusardi, P.; Piazza, E.; Fogari, R. Cardiovascular effects of melatonin in hypertensive patients well controlled by nifedipine: A 24-hour study. Br. J. Clin. Pharmacol. 2000, 49, 423–427. [Google Scholar] [CrossRef]
- Zhou, H.; Ma, Q.; Zhu, P.; Ren, J.; Reiter, R.J.; Chen, Y. Protective role of melatonin in cardiac ischemia-reperfusion injury: From pathogenesis to targeted therapy. J. Pineal Res. 2018, 64, e12471. [Google Scholar]
- Baker, J.; Kimpinski, K. Role of melatonin in blood pressure regulation: An adjunct anti-hypertensive agent. Clin. Exp. Pharmacol. Physiol. 2018, 45, 755–766. [Google Scholar] [PubMed]
- Ting, K.N.; A Blaylock, N.; Sugden, D.; Delagrange, P.; Scalbert, E.; Wilson, V.G. Molecular and pharmacological evidence for MT1 melatonin receptor subtype in the tail artery of juvenile Wistar rats. Br. J. Pharmacol. 1999, 127, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.N.; Barrios, V.E.; Duckles, S.P. Melatonin receptors mediate potentiation of contractile responses to adrenergic nerve stimulation in rat caudal artery. Eur. J. Pharmacol. 1995, 276, 207–213. [Google Scholar] [CrossRef]
- Geary, G.G.; Duckles, S.P.; Krause, D.N. Effect of melatonin in the rat tail artery: Role of K+ channels and endothelial factors. Br. J. Pharmacol. 1998, 123, 1533–1540. [Google Scholar] [CrossRef]
- Viswanathan, M.; Scalbert, E.; Delagrange, P.; Guardiola-Lemaître, B.; Saavedra, J.M. Melatonin receptors mediate contraction of a rat cerebral artery. Neuroreport 1997, 8, 3847–3849. [Google Scholar] [CrossRef]
- Geary, G.G.; Krause, D.N.; Duckles, S.P. Melatonin directly constricts rat cerebral arteries through modulation of potassium channels. Am. J. Physiol. Circ. Physiol. 1997, 273, H1530–H1536. [Google Scholar] [CrossRef]
- Doolen, S.; Krause, D.N.; Dubocovich, M.L.; Duckles, S.P. Melatonin mediates two distinct responses in vascular smooth muscle. Eur. J. Pharmacol. 1998, 345, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Duckles, S.P.; Dubocovich, M.L.; Krause, D.N. MT2 Melatonin Receptors Are Present and Functional in Rat Caudal Artery. J. Pharmacol. Exp. Ther. 2002. Available online: https://jpet.aspetjournals.org/content/302/3/1295.short?casa_token=zrUt7JhvzmMAAAAA:5a6J1SvtfYHjtWNg-i9tVHY8fNYCYg1XlulSvfwajsXjhUVI3dub9b2IKwOrqtN3JhdnG8OgWHdn (accessed on 13 January 2023).
- Pandi-Perumal, S.R.; Bahammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; Dhandapany, P.S.; Brown, G.M.; Cardinali, D.P. Melatonin and Human Cardiovascular Disease. J. Cardiovasc. Pharmacol. Ther. 2016, 22, 122–132. [Google Scholar] [CrossRef]
- Lochner, A.; Marais, E.; Huisamen, B. Melatonin and cardioprotection against ischaemia/reperfusion injury: What’s new? A review. J. Pineal Res. 2018, 65, e12490. [Google Scholar] [CrossRef]
- Tobeiha, M.; Jafari, A.; Fadaei, S.; Mirazimi, S.M.A.; Dashti, F.; Amiri, A.; Khan, H.; Asemi, Z.; Reiter, R.J.; Hamblin, M.R.; et al. Evidence for the Benefits of Melatonin in Cardiovascular Disease. Front. Cardiovasc. Med. 2022, 9, 888319. [Google Scholar] [CrossRef]
- Alonso-Vale, M.I.C.; Andreotti, S.; Mukai, P.Y.; Borges-Silva, C.D.N.; Peres, S.B.; Cipolla-Neto, J.; Lima, F.B. Melatonin and the circadian entrainment of metabolic and hormonal activities in primary isolated adipocytes. J. Pineal Res. 2008, 45, 422–429. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Owens, J.A.; Voultsios, A.; Wight, N. Adipokines and Adipocyte Function inClockMutant Mice That Retain Melatonin Rhythmicity. Obesity 2012, 20, 295–305. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Chen, S.; Yin, J.; Wang, J.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Li, T.; et al. Melatonin signaling in T cells: Functions and applications. J. Pineal Res. 2017, 62, e12394. [Google Scholar] [CrossRef]
- Bondy, S.C. Melatonin and Regulation of Immune Function: Impact on Numerous Diseases. Curr. Aging Sci. 2020, 13, 92–101. [Google Scholar] [CrossRef]
- Carrillo-Vico, A.; Lardone, P.J.; Álvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: Buffering the Immune System. Int. J. Mol. Sci. 2013, 14, 8638–8683. [Google Scholar] [CrossRef]
- Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Esquifino, A.I.; Pandi-Perumal, S.R.; Miller, S.C. Melatonin, immune function and aging. Immun. Ageing 2005, 2, 17. [Google Scholar] [CrossRef]
- Fenn, A.M.; Fonken, L.K.; Nelson, R.J. Sustained melatonin treatment blocks body mass, pelage, reproductive, and fever responses to short day lengths in female Siberian hamsters. J. Pineal Res. 2011, 51, 180–186. [Google Scholar] [CrossRef]
- Nelson, R.J.; Demas, G.E.; Klein, S.L.; Kriegsfeld, L.J. Minireview The influence of season, photoperiod, and pineal melatonin on immune function. J. Pineal Res. 1995, 19, 149–165. [Google Scholar]
- Demas, G.E.; Nelson, R.J. Exogenous Melatonin Enhances Cell-Mediated, but Not Humoral, Immune Function in Adult Male Deer Mice (Peromyscus maniculatus). J. Biol. Rhythm. 1998, 13, 245–252. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Drazen, D.L.; Nelson, R.J. In vitro melatonin treatment enhances cell-mediated immune function in male prairie voles (Microtus ochrogaster ). J. Pineal Res. 2001, 30, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Parisotto, E.B.; Vidal, V.; García-Cerro, S.; Lantigua, S.; Filho, D.W.; Sanchez-Barceló, E.J.; Martínez-Cué, C.; Rueda, N. Chronic Melatonin Administration Reduced Oxidative Damage and Cellular Senescence in the Hippocampus of a Mouse Model of Down Syndrome. Neurochem. Res. 2016, 41, 2904–2913. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.-X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Poeggeler, B.; Reiter, R.J.; Tan, D.-X.; Chen, L.-D.; Manchester, L.C. Melatonin, hydroxyl radical-mediated oxidative damage, and aging: A hypothesis. J. Pineal Res. 1993, 14, 151–168. [Google Scholar] [CrossRef]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef]
- Kang, J.-E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-β Dynamics Are Regulated by Orexin and the Sleep-Wake Cycle. Science 2009, 326, 1005–1007. [Google Scholar] [CrossRef]
- Beaupre, L.M.M.; Brown, G.M.; Gonçalves, V.F.; Kennedy, J.L. Melatonin’s neuroprotective role in mitochondria and its potential as a biomarker in aging, cognition and psychiatric disorders. Transl. Psychiatry 2021, 11, 339. [Google Scholar] [CrossRef]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2014, 22, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Topete, D.; Myers, P.H.; Foley, J.F.; Willis, M.; Cidlowski, J. Corticosteroids Are Essential for Maintaining Cardiovascular Function in Male Mice. Endocrinology 2016, 157, 2759–2771. [Google Scholar] [CrossRef] [PubMed]
- Burford, N.G.; Webster, N.A.; Cruz-Topete, D. Hypothalamic-Pituitary-Adrenal Axis Modulation of Glucocorticoids in the Cardiovascular System. Int. J. Mol. Sci. 2017, 18, 2150. [Google Scholar] [CrossRef]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids and Reproduction: Traffic Control on the Road to Reproduction. Trends Endocrinol. Metab. 2017, 28, 399–415. [Google Scholar] [CrossRef]
- Oster, H.; Damerow, S.; Kiessling, S.; Jakubcakova, V.; Abraham, D.; Tian, J.; Hoffmann, M.W.; Eichele, G. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006, 4, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Leon-Mercado, L.; Chao, D.H.M.; Basualdo, M.D.C.; Kawata, M.; Escobar, C.; Buijs, R.M. The Arcuate Nucleus: A Site of Fast Negative Feedback for Corticosterone Secretion in Male Rats. Eneuro 2017, 4, e0350. [Google Scholar]
- Weitzman, E.D.; Fukushima, D.; Nogeire, C.; Roffwarg, H.; Gallagher, T.F.; Hellman, L. Twenty-four Hour Pattern of the Episodic Secretion of Cortisol in Normal Subjects. J. Clin. Endocrinol. Metab. 1971, 33, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Selmaoui, B.; Touitou, Y. Reproducibility of the circadian rhythms of serum cortisol and melatonin in healthy subjects: A study of three different 24-h cycles over six weeks. Life Sci. 2003, 73, 3339–3349. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of Glucose Homeostasis by Glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 99–126. [Google Scholar] [CrossRef]
- Albers, H.E.; Yogev, L.; Todd, R.B.; Goldman, B.D. Adrenal corticoids in hamsters: Role in circadian timing. Am. J. Physiol. Integr. Comp. Physiol. 1985, 248, R434–R438. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How Do Glucocorticoids Influence Stress Responses? Integrating Permissive, Suppressive, Stimulatory, and Preparative Actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [PubMed]
- Dallman, M.F.; Strack, A.M.; Akana, S.F.; Bradbury, M.J.; Hanson, E.S.; Scribner, K.A.; Smith, M. Feast and Famine: Critical Role of Glucocorticoids with Insulin in Daily Energy Flow. Front. Neuroendocr. 1993, 14, 303–347. [Google Scholar] [CrossRef]
- McEwen, B.S.; Brinton, R.E.; Sapolsky, R.M. Glucocorticoid Receptors and Behavior: Implications for the Stress Response. Adv. Exp. Med. Biol. 1988, 245, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Urbanski, H.F. Role of Circadian Neuroendocrine Rhythms in the Control of Behavior and Physiology. Neuroendocrinology 2011, 93, 211–222. [Google Scholar] [CrossRef]
- Takahashi, J.S. Molecular Architecture of the Circadian Clock in Mammals. In A Time for Metabolism and Hormones; Sassone-Corsi, P., Christen, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Ohta, H.; Yamazaki, S.; McMahon, D.G. Constant light desynchronizes mammalian clock neurons. Nat. Neurosci. 2005, 8, 267–269. [Google Scholar] [CrossRef]
- Mazzoccoli, G.; Pazienza, V.; Vinciguerra, M. Clock Genes and Clock-Controlled Genes in the Regulation of Metabolic Rhythms. Chrono Int. 2012, 29, 227–251. [Google Scholar] [CrossRef]
- Coomans, C.P.; Berg, S.A.A.; Houben, T.; Klinken, J.; Berg, R.; Pronk, A.C.M.; Havekes, L.M.; Romijn, J.A.; Dijk, K.W.; Biermasz, N.R.; et al. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J. 2012, 27, 1721–1732. [Google Scholar]
- Fonken, L.K.; Workman, J.L.; Walton, J.C.; Weil, Z.M.; Morris, J.S.; Haim, A.; Nelson, R.J. Light at night increases body mass by shifting the time of food intake. Proc. Natl. Acad. Sci. USA 2010, 107, 18664–18669. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Finy, M.S.; Walton, J.; Weil, Z.; Workman, J.L.; Ross, J.; Nelson, R.J. Influence of light at night on murine anxiety- and depressive-like responses. Behav. Brain Res. 2009, 205, 349–354. [Google Scholar] [CrossRef]
- Catalano, F.; De Vito, F.; Cassano, V.; Fiorentino, T.V.; Sciacqua, A.; Hribal, M.L. Circadian Clock Desynchronization and Insulin Resistance. Int. J. Environ. Res. Public Health 2022, 20, 29. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, Y.; Zhang, F.; Xia, Y.; Liu, J.; Huang, R.; Wang, Y.; Hu, Y.; Wu, J.; Dai, C.; et al. CLOCK/BMAL1 regulates circadian change of mouse hepatic insulin sensitivity by SIRT1. Hepatology 2014, 59, 2196–2206. [Google Scholar] [CrossRef]
- Zhong, L.-X.; Li, X.-N.; Yang, G.-Y.; Zhang, X.; Li, W.-X.; Zhang, Q.-Q.; Pan, H.-X.; Zhang, H.-H.; Zhou, M.-Y.; Wang, Y.-D.; et al. Circadian misalignment alters insulin sensitivity during the light phase and shifts glucose tolerance rhythms in female mice. PLoS ONE 2019, 14, e0225813. [Google Scholar] [CrossRef]
- Mason, I.C.; Grimaldi, D.; Reid, K.J.; Warlick, C.D.; Malkani, R.G.; Abbott, S.M.; Zee, P.C. Light exposure during sleep impairs cardiometabolic function. Proc. Natl. Acad. Sci. USA 2022, 119, e2113290119. [Google Scholar] [CrossRef]
- Kooijman, S.; Berg, R.V.D.; Ramkisoensing, A.; Boon, M.R.; Kuipers, E.N.; Loef, M.; Zonneveld, T.C.M.; Lucassen, E.A.; Sips, H.C.M.; Chatzispyrou, I.A.; et al. Prolonged daily light exposure increases body fat mass through attenuation of brown adipose tissue activity. Proc. Natl. Acad. Sci. USA 2015, 112, 6748–6753. [Google Scholar] [CrossRef]
- Song, X.; Rusak, B. Acute effects of light on body temperature and activity in Syrian hamsters: Influence of circadian phase. Am. J. Physiol. Integr. Comp. Physiol. 2000, 278, R1369–R1380. [Google Scholar] [CrossRef]
- Jung, C.M.; Khalsa, S.B.; Scheer, F.; Cajochen, C.; Lockley, S.W.; Czeisler, C.A.; Wright, K.P. Acute Effects of Bright Light Exposure on Cortisol Levels. J. Biol. Rhythm. 2010, 25, 208–216. [Google Scholar] [CrossRef]
- Jones, J.R.; Chaturvedi, S.; Granados-Fuentes, D.; Herzog, E.D. Circadian neurons in the paraventricular nucleus entrain and sustain daily rhythms in glucocorticoids. Nat. Commun. 2021, 12, 5763. [Google Scholar] [CrossRef]
- Wilson, A.-L.; Downs, C.T. Light interference and melatonin affects digestion and glucocorticoid metabolites in striped mouse. Biol. Rhythm. Res. 2015, 46, 929–939. [Google Scholar] [CrossRef]
- Okuliarova, M.; Dzirbikova, Z.; Rumanova, V.S.; Foppen, E.; Kalsbeek, A.; Zeman, M. Disrupted Circadian Control of Hormonal Rhythms and Anticipatory Thirst by Dim Light at Night. Neuroendocrinology 2022, 112, 1116–1128. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Wright, H. Melatonin and circadian rhythms. Curr. Top. Med. Chem. 2002, 2, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Emmer, K.M.; Russart, K.L.G.; Walker, W.H.; Nelson, R.J.; DeVries, A.C. Effects of light at night on laboratory animals and research outcomes. Behav. Neurosci. 2018, 132, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Dauchy, R.T.; E Blask, D.; A Sauer, L.; Brainard, G.C.; A Krause, J. Dim light during darkness stimulates tumor progression by enhancing tumor fatty acid uptake and metabolism. Cancer Lett. 1999, 144, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Echakir, I.; Dumont, S.; Pã©Vet, P.; Eouarour, A.; Echallet, E.; Evuillez, P. Pineal melatonin is a circadian time-giver for leptin rhythm in Syrian hamsters. Front. Neurosci. 2015, 9, 190. [Google Scholar] [CrossRef]
- Scheving, E.L.; Pauly, E.J. Effect of light on corticosterone levels in plasma of rats. Am. J. Physiol. Content 1966, 210, 1112–1117. [Google Scholar] [CrossRef]
- Snyder, S.H.; Zweig, M.; Axelrod, J. Control of the circadian rhythm in serotonin content of the rat pineal gland. Life Sci. 1964, 3, 1175–1179. [Google Scholar] [CrossRef]
- Khan, S.; Duan, P.; Yao, L.; Hou, H. Shiftwork-Mediated Disruptions of Circadian Rhythms and Sleep Homeostasis Cause Serious Health Problems. Int. J. Genom. 2018, 2018, 8576890. [Google Scholar]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Fultz, N.E.; Bonmassar, G.; Setsompop, K.; Stickgold, R.A.; Rosen, B.R.; Polimeni, J.R.; Lewis, L.D. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 2019, 366, 628–631. [Google Scholar] [CrossRef]
- Gamble, K.L.; Resuehr, D.; Johnson, C.H.S. Work and Circadian Dysregulation of Reproduction. Front. Endocrinol. 2013, 4, 92. [Google Scholar] [CrossRef]
- Yaw, A.M.; McLane-Svoboda, A.K.; Hoffmann, H.M. Shiftwork and Light at Night Negatively Impact Molecular and Endocrine Timekeeping in the Female Reproductive Axis in Humans and Rodents. Int. J. Mol. Sci. 2020, 22, 324. [Google Scholar] [CrossRef]
- Figueiro, M.G.; Steverson, B.; Heerwagen, J.; Kampschroer, K.; Hunter, C.M.; Gonzales, K.; Plitnick, B.; Rea, M.S. The impact of daytime light exposures on sleep and mood in office workers. Sleep Health 2017, 3, 204–215. [Google Scholar] [CrossRef]
- Oike, H.; Sakurai, M.; Ippoushi, K.; Kobori, M. Time-fixed feeding prevents obesity induced by chronic advances of light/dark cycles in mouse models of jet-lag/shift work. Biochem. Biophys. Res. Commun. 2015, 465, 556–561. [Google Scholar] [CrossRef]
- Espitia-Bautista, E.; Velasco-Ramos, M.; Osnaya-Ramírez, I.; Ángeles-Castellanos, M.; Buijs, R.M.; Escobar, C. Social jet-lag potentiates obesity and metabolic syndrome when combined with cafeteria diet in rats. Metabolism 2017, 72, 83–93. [Google Scholar] [CrossRef]
- Scheer, F.A.J.L.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Shi, J.; Duan, P.; Liu, B.; Li, T.; Wang, C.; Li, H.; Yang, T.; Gan, Y.; Wang, X.; et al. Is shift work associated with a higher risk of overweight or obesity? A systematic review of observational studies with meta-analysis. Int. J. Epidemiol. 2018, 47, 1956–1971. [Google Scholar] [CrossRef]
- Mota, M.C.; Silva, C.M.; Balieiro, L.C.T.; Gonçalves, B.F.; Fahmy, W.M.; Crispim, C.A. Association between social jetlag food consumption and meal times in patients with obesity-related chronic diseases. PLoS ONE 2019, 14, e0212126. [Google Scholar] [CrossRef]
- Yong, M.; Nasterlack, M.; Pluto, R.-P.; Elmerich, K.; Karl, D.; Knauth, P. Is health, measured by work ability index, affected by 12-hour rotating shift schedules? Chrono Int. 2010, 27, 1135–1148. [Google Scholar] [CrossRef]
- Yong, M.; Nasterlack, M.; Messerer, P.; Oberlinner, C.; Lang, S. A retrospective cohort study of shift work and risk of cancer-specific mortality in German male chemical workers. Int. Arch. Occup. Environ. Health 2013, 87, 175–183. [Google Scholar] [CrossRef]
- Bittner, N.; Korf, H.-W.; Stumme, J.; Jockwitz, C.; Moebus, S.; Schmidt, B.; Dragano, N.; Caspers, S. Multimodal investigation of the association between shift work and the brain in a population-based sample of older adults. Sci. Rep. 2022, 12, 2969. [Google Scholar] [CrossRef]
- Wehrens, S.M.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; Skene, D.J.; Johnston, J.D. Meal Timing Regulates the Human Circadian System. Curr. Biol. 2017, 27, 1768–1775.e3. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Stunkard, A.J.; Rogers, N.L.; Van Dongen, H.P.; Allison, K.C.; O’Reardon, J.P.; Ahima, R.S.; Cummings, D.E.; Heo, M.; Dinges, D.F. Circadian Rhythm Profiles in Women with Night Eating Syndrome. J. Biol. Rhythm. 2009, 24, 85–94. [Google Scholar] [CrossRef]
- Fleury, G.; Masís-Vargas, A.; Kalsbeek, A. Metabolic Implications of Exposure to Light at Night: Lessons from Animal and Human Studies. Obesity 2020, 28, S18–S28. [Google Scholar] [CrossRef] [PubMed]
- Borniger, J.C.; Weil, Z.M.; Zhang, N.; Nelson, R.J. Dim Light at Night Does Not Disrupt Timing or Quality of Sleep in Mice. Chrono Int. 2013, 30, 1016–1023. [Google Scholar] [CrossRef]
- Stenvers, D.J.; van Dorp, R.; Foppen, E.; Mendoza, J.; Opperhuizen, A.-L.; Fliers, E.; Bisschop, P.H.; Meijer, J.H.; Kalsbeek, A.; Deboer, T. Dim light at night disturbs the daily sleep-wake cycle in the rat. Sci. Rep. 2016, 6, 35662. [Google Scholar] [CrossRef]
- Figueiro, M.G.; Radetsky, L.; Plitnick, B.; Rea, M.S. Glucose tolerance in mice exposed to light–dark stimulus patterns mirroring dayshift and rotating shift schedules. Sci. Rep. 2017, 7, 40661. [Google Scholar]
- Rumanova, V.S.; Okuliarova, M.; Foppen, E.; Kalsbeek, A.; Zeman, M. Exposure to dim light at night alters daily rhythms of glucose and lipid metabolism in rats. Front. Physiol. 2022, 13, 973461. [Google Scholar] [CrossRef]
- Fonken, L.; Weil, Z.; Nelson, R. Dark nights reverse metabolic disruption caused by dim light at night. Obesity 2013, 21, 1159–1164. [Google Scholar] [CrossRef]
- Masís-Vargas, A.; Hicks, D.; Kalsbeek, A.; Mendoza, J. Blue light at night acutely impairs glucose tolerance and increases sugar intake in the diurnal rodent Arvicanthis ansorgei in a sex-dependent manner. Physiol. Rep. 2019, 7, e14257. [Google Scholar] [CrossRef]
- Smale, L.; Lee, T.; Nunez, A.A. Mammalian Diurnality: Some Facts and Gaps. J. Biol. Rhythm. 2003, 18, 356–366. [Google Scholar] [CrossRef]
- Mendoza, J. Nighttime Light Hurts Mammalian Physiology: What Diurnal Rodent Models Are Telling Us. Clocks Sleep 2021, 3, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.-Q.; Li, J.; Wang, Y.; Wang, J.; Xu, J.-Y.; Kaneko, T. The effects of nocturnal life on endocrine circadian patterns in healthy adults. Life Sci. 2003, 73, 2467–2475. [Google Scholar] [CrossRef] [PubMed]
- Leproult, R.; Holmbäck, U.; Van Cauter, E. Circadian Misalignment Augments Markers of Insulin Resistance and Inflammation, Independently of Sleep Loss. Diabetes 2014, 63, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- Wefers, J.; van Moorsel, D.; Hansen, J.; Connell, N.J.; Havekes, B.; Hoeks, J.; Lichtenbelt, W.D.V.M.; Duez, H.; Phielix, E.; Kalsbeek, A.; et al. Circadian misalignment induces fatty acid metabolism gene profiles and compromises insulin sensitivity in human skeletal muscle. Proc. Natl. Acad. Sci. USA 2018, 115, 7789–7794. [Google Scholar] [CrossRef]
- Dinneen, S.; Alzaid, A.; Miles, J.; Rizza, R. Metabolic effects of the nocturnal rise in cortisol on carbohydrate metabolism in normal humans. J. Clin. Investig. 1993, 92, 2283–2290. [Google Scholar] [CrossRef]
- Rizza, R.A.; Mandarino, L.J.; Gerich, J.E. Cortisol-Induced Insulin Resistance in Man: Impaired Suppression of Glucose Production and Stimulation of Glucose Utilization due to a Postreceptor Defect of Insulin Action. J. Clin. Endocrinol. Metab. 1982, 54, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Stunkard, A.J.; Allison, K.C. Two forms of disordered eating in obesity: Binge eating and night eating. Int. J. Obes. 2003, 27, 1–12. [Google Scholar] [CrossRef]
- Schiavo-Cardozo, D.; Lima, M.M.O.; Pareja, J.C.; Geloneze, B. Appetite-regulating hormones from the upper gut: Disrupted control of xenin and ghrelin in night workers. Clin. Endocrinol. 2013, 79, 807–811. [Google Scholar] [CrossRef]
- Taylor, R. Insulin Resistance and Type 2 Diabetes. Diabetes 2012, 61, 778–779. [Google Scholar] [CrossRef]
- Donin, A.S.; Nightingale, C.M.; Owen, C.G.; Rudnicka, A.R.; Jebb, S.A.; Ambrosini, G.L.; Stephen, A.M.; Cook, D.G.; Whincup, P.H. Dietary Energy Intake Is Associated With Type 2 Diabetes Risk Markers in Children. Diabetes Care 2013, 37, 116–123. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Y.; Wei, F.; Song, J.; Cao, Z.; Chen, C.; Zhang, K.; Feng, S.; Wang, Y.; Li, W.-D. Triglyceride is an independent predictor of type 2 diabetes among middle-aged and older adults: A prospective study with 8-year follow-ups in two cohorts. J. Transl. Med. 2019, 17, 403. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, D.J.; Punjabi, N.M.; Newman, A.B.; Resnick, H.E.; Redline, S.; Baldwin, C.; Nieto, F.J. Association of Sleep Time With Diabetes Mellitus and Impaired Glucose Tolerance. Arch. Intern. Med. 2005, 165, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Renier, G. The Connection Between C-Reactive Protein (CRP) and Diabetic Vasculopathy. Focus on Preclinical Findings. Curr. Diabetes Rev. 2010, 6, 27–34. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Tune, J.D.; Goodwill, A.G.; Sassoon, D.J.; Mather, K.J. Cardiovascular consequences of metabolic syndrome. Transl. Res. 2017, 183, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Guembe, M.J.; Fernandez-Lazaro, C.I.; Sayon-Orea, C.; Toledo, E.; Moreno-Iribas, C.; Cosials, J.B.; Reyero, J.B.; Martínez, J.D.; Diego, P.G.; Uche, A.M.G.; et al. Risk for cardiovascular disease associated with metabolic syndrome and its components: A 13-year prospective study in the RIVANA cohort. Cardiovasc. Diabetol. 2020, 19, 195. [Google Scholar] [CrossRef]
- Ridker, P.M.; Glynn, R.J.; Hennekens, C.H. C-Reactive Protein Adds to the Predictive Value of Total and HDL Cholesterol in Determining Risk of First Myocardial Infarction. Circulation 1998, 97, 2007–2011. [Google Scholar] [CrossRef]
- Emerging Risk Factors Collaboration; Kaptoge, S.; Di Angelantonio, E.; Pennells, L.; Wood, A.M.; White, I.R.; Gao, P.; Walker, M.; Thompson, A.; Sarwar, N.; et al. C-Reactive Protein, Fibrinogen, and Cardiovascular Disease Prediction. N. Engl. J. Med. 2012, 367, 1310–1320. [Google Scholar] [CrossRef]
- Verma, S.; Wang, C.-H.; Li, S.-H.; Dumont, A.S.; Fedak, P.; Badiwala, M.V.; Dhillon, B.; Weisel, R.D.; Li, R.-K.; Mickle, D.A.; et al. A Self-Fulfilling Prophecy. Circulation 2002, 106, 913–919. [Google Scholar] [CrossRef]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Bayon, V.; Berger, M.; Solelhac, G.; Haba-Rubio, J.; Marques-Vidal, P.; Strippoli, M.-P.; Preisig, M.; Leger, D.; Heinzer, R. Impact of night and shift work on metabolic syndrome and its components: A cross-sectional study in an active middle-to-older-aged population-based sample. BMJ Open 2022, 12, e053591. [Google Scholar] [CrossRef] [PubMed]
- Brum, M.C.B.; Filho, F.F.D.; Schnorr, C.C.; Bertoletti, O.A.; Bottega, G.B.; Rodrigues, T.D.C. Night shift work, short sleep and obesity. Diabetol. Metab. Syndr. 2020, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Feng, W.; Wang, F.; Li, P.; Li, Z.; Li, M.; Tse, G.; Vlaanderen, J.; Vermeulen, R.; Tse, L.A. Meta-analysis on shift work and risks of specific obesity types. Obes. Rev. 2017, 19, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Shin, S.-Y.; Kang, Y.; Rhie, J. Effect of night shift work on the control of hypertension and diabetes in workers taking medication. Ann. Occup. Environ. Med. 2019, 31, e27. [Google Scholar] [CrossRef] [PubMed]
- Vetter, C.; Devore, E.E.; Ramin, C.A.; Speizer, F.E.; Willett, W.C.; Schernhammer, E.S. Mismatch of Sleep and Work Timing and Risk of Type 2 Diabetes. Diabetes Care 2015, 38, 1707–1713. [Google Scholar] [CrossRef]
- Yeom, J.H.; Sim, C.S.; Lee, J.; Yun, S.H.; Park, S.J.; Yoo, C.-I.; Sung, J.H. Effect of shift work on hypertension: Cross sectional study. Ann. Occup. Environ. Med. 2017, 29, 11. [Google Scholar] [CrossRef]
- Manohar, S.; Thongprayoon, C.; Cheungpasitporn, W.; Mao, M.A.; Herrmann, S. Associations of rotational shift work and night shift status with hypertension. J. Hypertens. 2017, 35, 1929–1937. [Google Scholar] [CrossRef]
- Vyas, M.V.; Garg, A.X.; Iansavichus, A.V.; Costella, J.; Donner, A.; Laugsand, L.E.; Janszky, I.; Mrkobrada, M.; Parraga, G.; Hackam, D.G. Shift work and vascular events: Systematic review and meta-analysis. BMJ 2012, 345, e4800. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, X.; Wan, F.; Gao, L.; Lin, N.; He, J.; Wei, L.; Dong, J.; Qin, Z.; Zhong, F.; et al. Disruption of Circadian Rhythms by Shift Work Exacerbates Reperfusion Injury in Myocardial Infarction. J. Am. Coll. Cardiol. 2022, 79, 2097–2115. [Google Scholar] [CrossRef]
- Bae, M.-J.; Song, Y.-M.; Shin, J.; Choi, B.-Y.; Keum, J.-H.; Lee, E.-A. The Association Between Shift Work and Health Behavior: Findings from the Korean National Health and Nutrition Examination Survey. Korean J. Fam. Med. 2017, 38, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Knutsson, A.; Nilsson, T. Tobacco use and exposure to environmental tobacco smoke in relation to certain work characteristics. Scand. J. Soc. Med. 1998, 26, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Trinkoff, A.M.; Storr, C.L. Work schedule characteristics and substance use in nurses. Am. J. Ind. Med. 1998, 34, 266–271. [Google Scholar] [CrossRef]
- Kivimäki, M.; Kuisma, P.; Virtanen, M.; Elovainio, M. Does shift work lead to poorer health habits? A comparison between women who had always done shift work with those who had never done shift work. Work. Stress 2001, 15, 3–13. [Google Scholar] [CrossRef]
- Richter, K.; Peter, L.; Rodenbeck, A.; Weess, H.G.; Riedel-Heller, S.G.; Hillemacher, T. Shiftwork and Alcohol Consumption: A Systematic Review of the Literature. Eur. Addict. Res. 2020, 27, 9–15. [Google Scholar] [CrossRef]
- Brum, M.C.B.; Filho, F.F.D.; Schnorr, C.C.; Bottega, G.B.; Rodrigues, T.C. Shift work and its association with metabolic disorders. Diabetol. Metab. Syndr. 2015, 7, 45. [Google Scholar] [CrossRef]
- Monnaatsie, M.; Biddle, S.J.; Khan, S.; Kolbe-Alexander, T. Physical activity and sedentary behaviour in shift and non-shift workers: A systematic review and meta-analysis. Prev. Med. Rep. 2021, 24, 101597. [Google Scholar] [CrossRef]
- Knutsson, A.; Akerstedt, T.; Jonsson, B.G. Prevalence of risk factors for coronary artery disease among day and shift workers. Scand. J. Work. Environ. Heal. 1988, 14, 317–321. [Google Scholar] [CrossRef]
- Romon, M.; Nuttens, M.-C.; Fievet, C.; Pot, P.; Bard, J.M.; Furon, D.; Fruchart, J.C. Increased triglyceride levels in shift workers. Am. J. Med. 1992, 93, 259–262. [Google Scholar] [CrossRef]
- Dutheil, F.; Baker, J.S.; Mermillod, M.; De Cesare, M.; Vidal, A.; Moustafa, F.; Pereira, B.; Navel, V. Shift work, and particularly permanent night shifts, promote dyslipidaemia: A systematic review and meta-analysis. Atherosclerosis 2020, 313, 156–169. [Google Scholar] [CrossRef]
- Shechter, A.; Kim, E.W.; St-Onge, M.-P.; Westwood, A.J. Blocking nocturnal blue light for insomnia: A randomized controlled trial. J. Psychiatr. Res. 2017, 96, 196–202. [Google Scholar] [CrossRef]
- Chang, A.-M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2014, 112, 1232–1237. [Google Scholar] [CrossRef]
- Hester, L.; Dang, D.; Barker, C.J.; Heath, M.; Mesiya, S.; Tienabeso, T.; Watson, K. Evening wear of blue-blocking glasses for sleep and mood disorders: A systematic review. Chrono Int. 2021, 38, 1375–1383. [Google Scholar] [CrossRef]
- Vethe, D.; Scott, J.; Engstrøm, M.; Salvesen, Ø.; Sand, T.; Olsen, A.; Morken, G.; Heglum, H.S.; Kjørstad, K.; Faaland, P.M.; et al. The evening light environment in hospitals can be designed to produce less disruptive effects on the circadian system and improve sleep. Sleep 2020, 44, zsaa194. [Google Scholar] [CrossRef]
- Avery, D.H.; Eder, D.N.; Bolte, M.A.; Hellekson, C.J.; Dunner, D.L.; Vitiello, M.V.; Prinz, P.N. Dawn simulation and bright light in the treatment of SAD: A controlled study. Biol. Psychiatry 2001, 50, 205–216. [Google Scholar] [CrossRef]
- Campbell, P.D.; Miller, A.M.; Woesner, M.E. Bright Light Therapy: Seasonal Affective Disorder and Beyond. Einstein J. Biol. Med. 2017, 32, E13–E25. [Google Scholar]
- Rosenthal, N.E.; Sack, D.A.; Gillin, J.C.; Lewy, A.J.; Goodwin, F.K.; Davenport, Y.; Mueller, P.S.; Newsome, D.A.; Wehr, T.A. Seasonal Affective Disorder. Arch. Gen. Psychiatry 1984, 41, 72–80. [Google Scholar] [CrossRef]
- Terman, J.S.; Terman, M.; Lo, E.-S.; Cooper, T.B. Circadian Time of Morning Light Administration and Therapeutic Response in Winter Depression. Arch. Gen. Psychiatry 2001, 58, 69–75. [Google Scholar] [CrossRef]
- Cibeira, N.; Maseda, A.; Lorenzo-López, L.; González-Abraldes, I.; López-López, R.; Rodríguez-Villamil, J.L.; Millán-Calenti, J.C. Bright Light Therapy in Older Adults with Moderate to Very Severe Dementia: Immediate Effects on Behavior, Mood, and Physiological Parameters. Healthcare 2021, 9, 1065. [Google Scholar] [CrossRef]
- Al-Karawi, D.; Jubair, L. Bright light therapy for nonseasonal depression: Meta-analysis of clinical trials. J. Affect. Disord. 2016, 198, 64–71. [Google Scholar] [CrossRef]
- Elliott, J.E.; McBride, A.A.; Balba, N.M.; Thomas, S.V.; Pattinson, C.L.; Morasco, B.J.; Wilkerson, A.; Gill, J.M.; Lim, M.M. Feasibility and preliminary efficacy for morning bright light therapy to improve sleep and plasma biomarkers in US Veterans with TBI. A prospective, open-label, single-arm trial. PLoS ONE 2022, 17, e0262955. [Google Scholar] [CrossRef]
- Korman, M.; Palm, D.; Uzoni, A.; Faltraco, F.; Tucha, O.; Thome, J.; Coogan, A.N. ADHD 24/7: Circadian clock genes, chronotherapy and sleep/wake cycle insufficiencies in ADHD. World J. Biol. Psychiatry 2018, 21, 156–171. [Google Scholar] [CrossRef]
- van Andel, E.; Bijlenga, D.; Vogel, S.W.N.; Beekman, A.T.F.; Kooij, J.J.S. Effects of chronotherapy on circadian rhythm and ADHD symptoms in adults with attention-deficit/hyperactivity disorder and delayed sleep phase syndrome: A randomized clinical trial. Chrono Int. 2020, 38, 260–269. [Google Scholar] [CrossRef]
- Laposky, A.D.; Bass, J.; Kohsaka, A.; Turek, F.W. Sleep and circadian rhythms: Key components in the regulation of energy metabolism. FEBS Lett. 2007, 582, 142–151. [Google Scholar] [CrossRef]
- Rumanova, V.S.; Okuliarova, M.; Zeman, M. Differential Effects of Constant Light and Dim Light at Night on the Circadian Control of Metabolism and Behavior. Int. J. Mol. Sci. 2020, 21, 5478. [Google Scholar] [CrossRef]
- Froy, O. Metabolism and Circadian Rhythms—Implications for Obesity. Endocr. Rev. 2009, 31, 1–24. [Google Scholar]
- Asher, G.; Sassone-Corsi, P. Time for Food: The Intimate Interplay between Nutrition, Metabolism, and the Circadian Clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef]
- Rana, S.; Prabhu, S.D.; Young, M.E. Chronobiological Influence Over Cardiovascular Function. Circ. Res. 2020, 126, 258–279. [Google Scholar] [CrossRef]
- Bray, M.S.; Shaw, C.A.; Moore, M.W.S.; Garcia, R.A.P.; Zanquetta, M.M.; Durgan, D.J.; Jeong, W.J.; Tsai, J.-Y.; Bugger, H.; Zhang, D.; et al. Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. Am. J. Physiol. Circ. Physiol. 2008, 294, H1036–H1047. [Google Scholar] [CrossRef]
- Arble, D.; Bass, J.; Laposky, A.D.; Vitaterna, M.H.; Turek, F.W. Circadian Timing of Food Intake Contributes to Weight Gain. Obesity 2009, 17, 2100–2102. [Google Scholar] [CrossRef]
- Challet, E. The circadian regulation of food intake. Nat. Rev. Endocrinol. 2019, 15, 393–405. [Google Scholar] [CrossRef]
- Acosta-Rodríguez, V.; Rijo-Ferreira, F.; Izumo, M.; Xu, P.; Wight-Carter, M.; Green, C.B.; Takahashi, J.S. Circadian alignment of early onset caloric restriction promotes longevity in male C57BL/6J mice. Science 2022, 376, 1192–1202. [Google Scholar] [CrossRef]
- Grigg-Damberger, M.M.; Ianakieva, D. Poor Quality Control of Over-the-Counter Melatonin: What They Say Is Often Not What You Get. J. Clin. Sleep Med. 2017, 13, 163–165. [Google Scholar] [CrossRef]
- Erland, L.; Saxena, P.K. Melatonin Natural Health Products and Supplements: Presence of Serotonin and Significant Variability of Melatonin Content. J. Clin. Sleep Med. 2017, 13, 275–281. [Google Scholar] [CrossRef]
- Li, J.; Somers, V.K.; Xu, H.; Lopez-Jimenez, F.; Covassin, N. Trends in Use of Melatonin Supplements Among US Adults, 1999–2018. JAMA 2022, 327, 483–485. [Google Scholar] [CrossRef]
- National Institutes of Health (NIH). Use of Melatonin Supplements Rising Among Adults. 2022. Available online: https://www.nih.gov/news-events/nih-research-matters/use-melatonin-supplements-rising-among-adults (accessed on 13 January 2023).
- Garde, A.H.; Begtrup, L.; Bjorvatn, B.; Bonde, J.P.; Hansen, J.; Hansen, Å.M.; Härmä, M.; Jensen, M.A.; Kecklund, G.; A Kolstad, H.; et al. How to schedule night shift work in order to reduce health and safety risks. Scand. J. Work. Environ. Health 2020, 46, 557–569. [Google Scholar] [CrossRef]
- der Holst, H.L.-V.; Murphy, A.; Wise, J.; Duffy, J. Sleep tips for shift workers in the time of pandemic. Southwest J Pulm Crit Care. Southwest J. Pulm. Crit. Care 2020, 20, 128–130. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meléndez-Fernández, O.H.; Liu, J.A.; Nelson, R.J. Circadian Rhythms Disrupted by Light at Night and Mistimed Food Intake Alter Hormonal Rhythms and Metabolism. Int. J. Mol. Sci. 2023, 24, 3392. https://doi.org/10.3390/ijms24043392
Meléndez-Fernández OH, Liu JA, Nelson RJ. Circadian Rhythms Disrupted by Light at Night and Mistimed Food Intake Alter Hormonal Rhythms and Metabolism. International Journal of Molecular Sciences. 2023; 24(4):3392. https://doi.org/10.3390/ijms24043392
Chicago/Turabian StyleMeléndez-Fernández, O. Hecmarie, Jennifer A. Liu, and Randy J. Nelson. 2023. "Circadian Rhythms Disrupted by Light at Night and Mistimed Food Intake Alter Hormonal Rhythms and Metabolism" International Journal of Molecular Sciences 24, no. 4: 3392. https://doi.org/10.3390/ijms24043392
APA StyleMeléndez-Fernández, O. H., Liu, J. A., & Nelson, R. J. (2023). Circadian Rhythms Disrupted by Light at Night and Mistimed Food Intake Alter Hormonal Rhythms and Metabolism. International Journal of Molecular Sciences, 24(4), 3392. https://doi.org/10.3390/ijms24043392