
Emerging Roles of Salicylic Acid in Plant Saline Stress Tolerance

1
State Key Laboratory of Crop Biology, College of Life Sciences, Shandong Agricultural University, Taian 271018, China
2
State Key Laboratory of Hybrid Rice, College of Life Sciences, Wuhan University, Wuhan 430072, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(4), 3388; https://doi.org/10.3390/ijms24043388
Submission received: 15 December 2022
/
Revised: 18 January 2023
/
Accepted: 6 February 2023
/
Published: 8 February 2023
(This article belongs to the Special Issue Crop Biotic and Abiotic Stress Tolerance 2.0)
Abstract
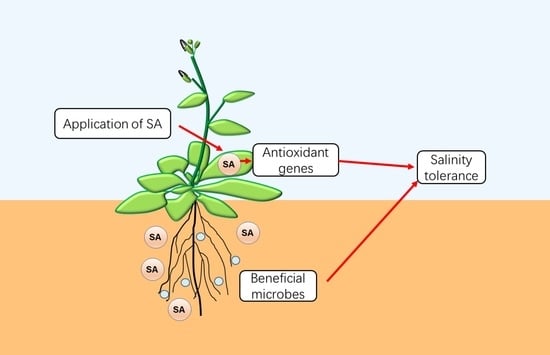
:One of the most important phytohormones is salicylic acid (SA), which is essential for the regulation of plant growth, development, ripening, and defense responses. The role of SA in plant–pathogen interactions has attracted a lot of attention. Aside from defense responses, SA is also important in responding to abiotic stimuli. It has been proposed to have great potential for improving the stress resistance of major agricultural crops. On the other hand, SA utilization is dependent on the dosage of the applied SA, the technique of application, and the status of the plants (e.g., developmental stage and acclimation). Here, we reviewed the impact of SA on saline stress responses and the associated molecular pathways, as well as recent studies toward understanding the hubs and crosstalk between SA-induced tolerances to biotic and saline stress. We propose that elucidating the mechanism of the SA-specific response to various stresses, as well as SA-induced rhizosphere-specific microbiome modeling, may provide more insights and support in coping with plant saline stress.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The global food supply is under severe strain, with overall food consumption expected to increase by 70–85 percent to feed 9 billion people for the increased world population by 2050 [1]. However, abiotic challenges, such as high temperature, salinity, drought, and cold, are important factors that restrict agricultural productivity. As a result of the effect of human activities and the intensification of climate change, crops are subjected to increasing abiotic stressors, which have become the primary cause of agricultural system degradation and grain production decline [2,3]. Furthermore, the increased demand has compelled more crops to be grown on virgin land that is frequently subjected to abiotic stress due to insufficient irrigation, salinity, metal toxicity, and a lack of nutrients. One of the most severe constraints is saline stress caused by soil salinization [4]. Salinization is the process of raising the concentration of total dissolved salts in soil and water, which can occur naturally (primary salinization) or as a result of anthropogenic actions (secondary salinization). Climate change enhances the amount of salinization that occurs, through either a rise in sea levels or higher evaporation during droughts, while the use of saline groundwater and low-quality wastewater for irrigation exacerbates the secondary salinization process [4,5]. Saline stress has a direct adverse effect on plant growth, development, and yield due to its impacts on plant biochemistry and physiology. It extends beyond all stages of development, from seed germination to maturity. Under moderate salinity environments, all the important glycophytic crops reduce their average yield by 50–80% [5]. As a consequence, the strategy of sustainable development will include planting salt tolerance varieties, as well as inducing cultivated plants to resist or tolerate abiotic stresses through previously unknown resistance mechanisms. For instance, salicylic acid (SA) has been proposed to play an important role in resistance and defense signaling in both saline and biotic stress [6,7].
SA is an endogenous small-molecule phenolic compound that acts as a signal sensor to regulate plant response. It protects plant cells from the toxicity of ion accumulation and cell death by managing processes such as antioxidant defense, nitrogen metabolism, photosynthesis, and water stress [8,9]. The variations in SA levels among plant species, as well as due to environmental challenges, are quite dramatic [10]. The effect of endogenous SA levels has been related to a plant’s developmental stage and exogenous stimulation levels [11]. Furthermore, plant tolerance to salt, drought, heat, cold, and heavy metals can be induced by exogenous SA treatment [12,13,14,15,16]. Abiotic stress may induce serial gene expressions in plants, some of which are related to SA-dependent activation. These genes have an impact on a variety of biological processes, including molecular chaperones, antioxidant biosynthesis, and secondary metabolite synthesis [17,18,19].
Significant progress have been made in signal transduction under saline stress in recent years, particularly in the early signaling induced by salt. The significant role of both calcium waves and reactive oxygen species (ROS) and their downstream targets has been demonstrated, while the cell wall has frequently been implicated as a modulator of cell expansion during salt stress [20]. On the other hand, significant progress has been achieved with the biosynthetic pathway of SA in plants. Despite many studies having been conducted on the role of SA in plant salt stress, information on relatively in-depth mechanisms is still limited. Here, we propose to integrate the biosynthesis of SA, the role of SA in various abiotic stresses, and the potential mechanism of SA-mediated plant tolerance to salt stresses in this review. We also discuss the similarities and differences in SA signal transmission modes when plants are subjected to salt or biotic stresses. This information is beneficial for a better understanding of the role of the SA signal system in a stressed plant. Our understanding of signal pathways and growth adaptation mainly comes from the model plant Arabidopsis, which has not been directly transferred to crops. Plant saline tolerance is a complex trait that can manifest itself in a variety of ways, including ion accumulation, tissue-specific growth rate, biomass production, and seed production. Different salt reactions must be optimized for different crops in order to increase yield. The interplay of the SA signaling pathway and saline tolerance signaling will allow for the development of novel salt-tolerant crop varieties [4].
2. SA Biosynthesis in Plants
SA is initially synthesized from chorismate in plants through two independent pathways: the phenylpropane (PAL) and isochorismate synthase (ICS) pathways. In Arabidopsis, the ICS pathway plays the predominant role in the production of SA during plant defense [21]. Isochorismate synthase 1 (ICS1, also known as SID2 or EDS16) converts chorismate into isochorismate in the chloroplast [22]. ics1 is sensitive to salinity [23], implying that SA biosynthesis through the ICS pathway is important for saline tolerance in plants. A chloroplast outer envelope protein named enhanced disease susceptibility 5 (EDS5) has also been characterized as a multidrug and toxin extrusion (MATE) transporter family protein; it transports the isochorismate from plastids to the cytosol [24,25]. In some bacteria, isochorismate is directly converted to SA by isochorismate pyruvate lyase (IPL), which has not been identified in any homologs in plants [26]. In plants, recent studies revealed that isochorismate could be converted to SA by AvrPphB Susceptible 3 (PBS3) in the cytosol via a completely different mechanism than that in bacteria [25,27]. PBS3 serves as a GH3 acyl-adenylate/thioester-forming enzyme, which catalyzes the conjugation of isochorismate to glutamate to produce a biosynthetic intermediate isochorismate-9-glutamate (IC-9-Glu). IC-9-Glu can spontaneously decay into SA [25] or be converted to SA by enhanced pseudomonas susceptibility 1 (EPS1) [27].
Using PAL defective mutants or treatment with specific inhibitors, the PAL pathway was also identified with SA production, independent of the ICS pathway, in Arabidopsis [28]. Isotope tracing experiments in tobacco were performed to identify the complete steps for SA production in the PAL pathway. Trans-cinnamic acid (t-CA) was considered to be first synthesized from phenylalanine (Phe) and then converted to SA via benzoic acid (BA) [29]. The conversion of Phe to t-CA by PAL is one of the rate-determining steps in SA biosynthesis. The means by which the PAL pathway promotes SA synthesis is still controversial. A recent study indicated that the PAL gene catalyzes phenylalanine to finally synthesize 4-hydroxybenzoic acid (4-HBA) rather than SA [30]. In any case, it is clear that BA is the precursor of SA. Abnormal Inflorescence Meristem 1 (AIM1), encoding a 3-Hydroxyacyl-CoA dehydrogenase involved in β-oxidation, is required for BA production in rice roots [31]. The means by which the enzymes determine the final conversion of BA to SA remains unclear; some studies suggest that benzoic acid 2-hydroxylase (BA2H) or the cytochrome 450 family protein may play a role [32]. Salinity increases rice endogenous SA levels by increasing BA2H activity, suggesting that the PAL pathway is involved in saline stress [33].
The SA synthesis pathway may vary in other crops. Rice accumulates high levels of SA, approximately 10-fold more than that of Arabidopsis leaves under normal growth conditions [10]. The OsPAL6-knockout mutant showed about a 60% decrease in the basal SA level and increased susceptibility to the fungal pathogen Magnaporthe oryzae [34]. The SA levels were also significantly reduced in osaim1 [31]. Interestingly, both the ICS and PAL pathways were shown to contribute to pathogen-induced SA accumulation in soybeans. Silencing of either the ICS or PAL genes led to a severe reduction in SA accumulation during infection by the bacterial and oomycete pathogen [35]. These findings indicate that significant variations exist in different plant species for the contributions of the two SA biosynthesis pathways. Thus far, no clear orthologs of PBS3 or EPS1 have been identified in most plant species besides the Brassicaceae family; the role of the ICS pathway for SA synthesis in the plant kingdom needs more evaluation [36]. Similarly, PAL genes are also involved in the production of many secondary metabolites that are unrelated to SA, such as flavonoids. In order to better understand the function of the PAL pathway, it is necessary to identify the BA2H enzymes involved in the conversion of BA to SA in the future.
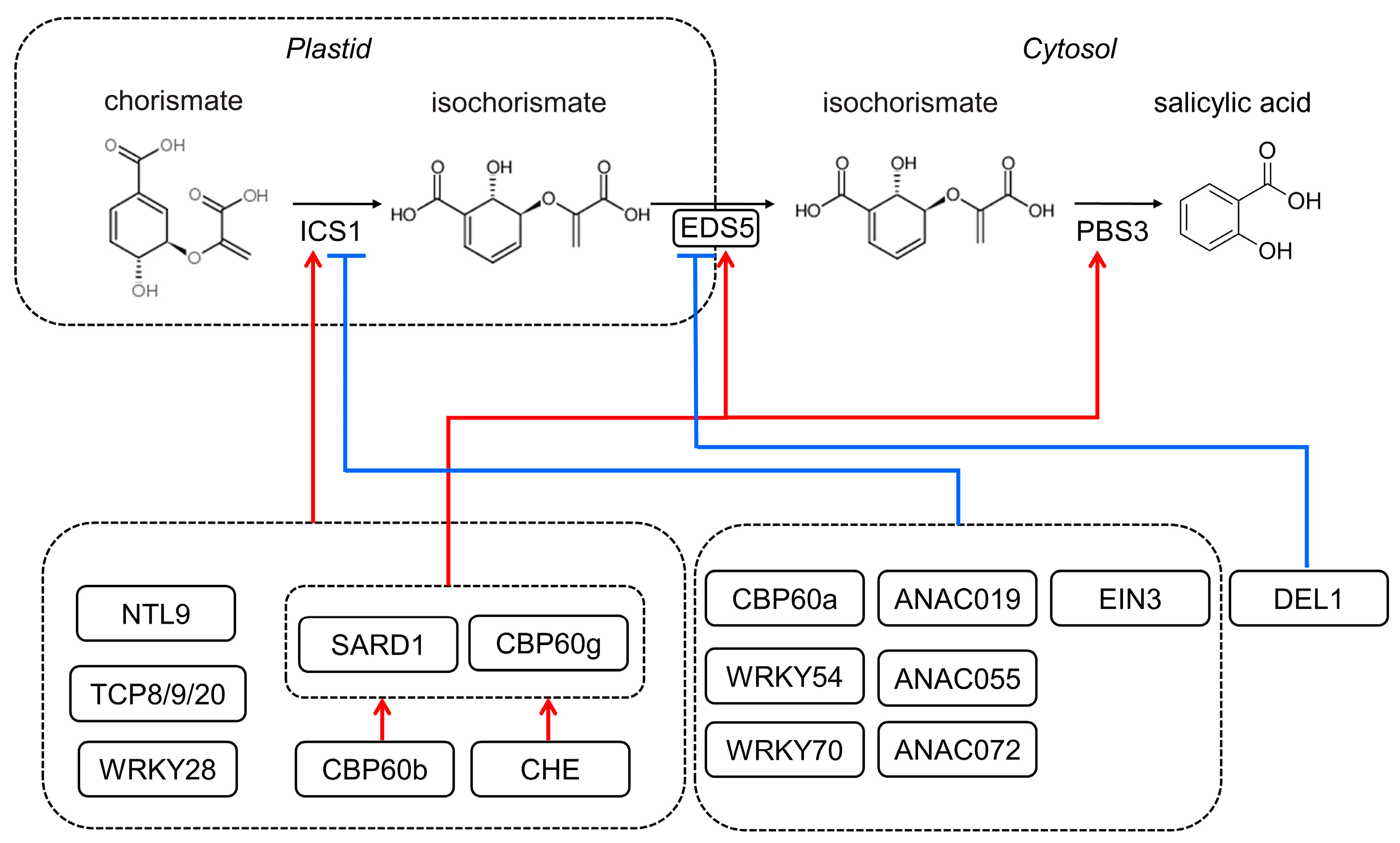
SA biosynthesis is strictly under regulation during biotic and abiotic stress. As summarized in Figure 1, TCP (Teosinte-like1/Cyclidea/PCF), WRKY (WRKY DNA-binding protein), and CBP60 (CAM Binding Protein 60) proteins have been identified as key transcription factors that control ICS1 expression [9,37]. Among these, the CBP60 proteins of SARD1 (systemic acquired resistance (SAR) defect 1) and CBP60g have been extensively studied [38]. Their expression was induced via TGA1 (TGACG-binding factor 1) and TGA4 by pathogen inoculation, which further binds to the promoter regions of ICS1, EDS5, and PBS3 to activate the SA synthesis-related gene expression [39,40]. CBP60b could influence the expression of SARD1 and CBP60g to modulate SA accumulation [41,42]. Coronatine stimulates the ANAC (abscisic acid-responsive NAC) transcription factors ANAC019, ANAC055, and ANAC072 via MYC2 to suppress the expression of ICS1 and lower the synthesis of SA [43]. Other transcription factors that suppress ICS1 expression include WRKY54, WRKY70, EIN3 (Ethylene Insensitive 3), and CBP60a [9,44]. Meanwhile, the atypical E2F transcriptional repressor DEL1 could suppress EDS5 expression [45].
3. Signal Transduction Pathway of SA
The produced SA in plants needs to be perceived by its receptors in order to trigger the downstream response. Several SA-binding proteins (SABPs) have been identified to perceive SA. NPR1 (non-expresser of PR genes 1), an SA-binding protein, has been well studied to play a crucial role in activating downstream disease resistance genes [46,47]. Pathogens promote phosphorylated NPR1, which is recruited to cullin 3 ubiquitin ligase for a series of physiological and biochemical reactions. The phosphorylated NPR1 is essential for SAR induction [36,48]. The monomer NPR1 is translocated into the nucleus for phosphorylation and interacts with bZIP (basic leucine zipper protein) transcription factor TGA family proteins to activate the expression of downstream resistance-related genes [9]. A recent study revealed that SA activates NPR1 in the nucleus to induce SAR and enhances the formation of NPR1 condensates during plant immunological responses [49]. Meanwhile, NPR1-dependent SA signaling is also central to saline and oxidative stress tolerance in Arabidopsis [50].
In addition, other NPRs operate as SA receptors and play a key regulatory role in the SA signal transduction cascade in Arabidopsis. NPR3 and NPR4 can function as E3 ubiquitin ligase linkers to mediate NPR1 degradation, during which SA sensing systems influence cell death and survival [51,52]. NPR1, NPR3, and NPR4 bind SA with varying affinities and play a role in binding to downstream TGA transcription factors in response to pathogen infections [53]. Moreover, the NPR3/4-mediated repression of CBP60g and SARD1 expression occurs under drought and bacterial combined stress in an SA-dependent manner, showing that NPR3/NPR4 is involved in plant resistance to abiotic stress [54]. NPR3/4 combine with TGA to inhibit downstream gene expression, whereas SA can relieve this inhibition [55]. As a result, two parallel signaling pathways may exist downstream of SA. On the one hand, when the concentration of SA in plants is very low, NPR3/4 block the expression of genes downstream of SA; the transcriptional repression effect of NPR3/4 on SA downstream genes is relieved based on elevated SA concentrations during pathogen infection. On the other hand, the accumulation of SA in plants activates the transcriptional activation activity of NPR1, which in turn induces the expression of downstream genes of SA [56].
For SA signal transduction, EDS1 (enhanced disease susceptibility 1) and PAD4 (phytoalexin deficiency 4) were regarded as essential upstream components of SA-dependent and SA-independent basal defense pathways [57]. They belong to the lipase-like enzymes family. The N-terminus of EDS1 and PAD4 interact with each other to form a binary complex. Broken interactions between EDS1 and PAD4 can hinder the induction and synthesis of SA and weaken the resistance to pathogenic bacteria [58]. SA accumulation will upregulate the expression of EDS1 and PAD4, thus forming a positive feedback signal loop [59]. EDS1 was protected by PBS3 from proteasome-mediated degradation, and the PBS3–EDS1 relationship may also boost EDS1 activity in SA and basal immunity [60].
4. Physiological Responses by SA during Saline Stress in Plants
The world’s saline soil area is close to 1 billion hm2, accounting for approximately 7% of the earth’s land area. It is extensively spread across Asia, the Americas, Australia, Africa, and other regions [61]. Furthermore, soil salinization threatens 77 million hm2 of land, and salinization has lowered productivity on roughly one-third of irrigated agriculture [62,63]. Meanwhile, as a consequence of global warming, the salinization area is dramatically increasing on a daily basis [4,64]. Salinization produces osmotic stress as well as ionic toxicity in cells. Plant production losses are frequently caused by an imbalance in cellular ionic and osmotic balances [4]. Furthermore, salt can cause plant stomatal closure and reduce intercellular CO2 concentration in the early stages of stress; in the latter stages, the chloroplasts become damaged and chlorophyll synthesis is blocked, affecting photosynthesis and resulting in decreased biomass, slowed plant growth, and even death [65,66,67]. SA can regulate important plant physiological processes, including photosynthesis, nitrogen metabolism, antioxidant defense system control, and water use efficiency improvement [8]. The pretreatment of rice with SA during germination during saline stress was shown to dramatically enhance the shoot and root lengths, resulting in greater salinity resistance [68]. SA was found to increase photosynthesis and the antioxidant response to boost yield components in the ASD16 and BR26 rice lines [69]. The participation of SA in ion transport, blooming, and photosynthesis may account for the contributions to the yield [70].
Saline stress not only affects the ability of plants’ roots to absorb water but also destroys leaf cells through transpiration, reducing plant growth. A Na+ overabundance in the soil will disrupt the ion dynamic equilibrium, causing a variety of stress responses, such as osmotic stress and ion stress in plants [18]. Excess Na+ ions also cause a K+ deficit and hinder a range of K+-dependent biological processes in plants. As a result, one of the primary symbols for measuring plant salt tolerance is the dynamic balance of Na+ and K+ in plant cells [65,70]. Saline stress also limits the absorption of other ions, such as Ca2+ and Mg2+, resulting in a nutritional shortfall [4]. SA may lower the Na+ and Cl− levels in the cell. Exogenous SA treatment was found to elevate internal SA levels and raise the K+ content, which decreased the Na+ levels in Arabidopsis and mung bean under saline stress, causing physiological changes in the plants [71].
A substantial number of reactive oxygen species (ROS) accumulate in plants in high-salt environments, reducing the fluidity of the cell membrane, increasing the permeability of the cell membrane, causing metabolic instability of the membrane system, and finally resulting in oxidative damage [72,73]. Some enzymes involved in oxidative stress, such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POX), and glutathione reductase (GR), elevated their enzymatic activities during saline stress [74]. However, these protein levels decreased within the cells and, therefore, restored the physiological mechanisms in plants after SA treatment [70,75]. Consistent with the physiological response, exogenously SA-treated plants showed increased expression in the transcripts of various antioxidant components, such as dehydroascorbate reductase (DHAR), glutathione peroxidase (GPX1, 2), glutathione synthetase (GS), and glutathione S-transferase (GST1, 2), under saline stress [76]. In the case of high saline stress, oxidative stress was minimized by the SA-mediated reduction of cellular malondialdehyde (MDA) and ROS production in Hordeum vulgare [8].
In addition, the buildup of osmotic regulators can improve plant saline tolerance. Plants resist salt-induced osmotic stress by maintaining the osmotic pressure equilibrium within and outside the cell, thereby managing the accumulation of organic or inorganic osmotic regulators in the cytoplasm [64,66,77]. Cl− and K+ are two major inorganic osmoregulators in plants; they do not require synthesis through metabolism [78]. Alanine, glutamate, asparagine, glycine, betaine, and sucrose are examples of organic osmoregulatory chemicals; the synthesis of these compounds increases considerably when exposed to saline stress. Betaine is an organic osmoregulation substance involved in the response to a variety of abiotic stressors. Plants can boost betaine synthesis in response to high salt, high temperature, and drought [64,79]. Treatment with 0.5 mM SA maximally increased the accumulation of glycine betaine (GB) (>40%), which acted against 50 mM NaCl-accrued impacts by minimizing the accumulation of Na+ and Cl− ions and, hence, oxidative stress [80]. Sugars and free amino acids are also key organic osmoregulators, and their buildup in the cytoplasm can improve saline tolerance in plants [81,82]. In particular, the concentration of proline has been utilized as an important physiological marker to quantify stress resistance in plants [83]. During exogenously applied SA, proline metabolism became significantly altered, leading to the maintenance of turgor by the accumulation of higher levels of free proline in lentils to enhance their saline tolerance [84].
5. Molecular Mechanisms Controlled by SA Regulation in Response to Saline Stress
The role of SA in strengthening salinity stress tolerance mechanisms has been reported in cereal plants such as rice, wheat, barley, and maize [85,86,87,88], and in other plant species including Cucumis sativus [89], Solanum lycopersicum [90], Vicia faba [91], and Medicago sativa [92]. Some SA synthesis genes, such as OsICS and OsPAL, show induced expression under saline stress [18,88]. Meanwhile, the introduction of the bacterial salicylate hydroxylase gene (NahG) into plants resulted in stronger tolerance to high saline stress than that of the wild type [93].
Under saline stress, the water potential of the soil solution is reduced, resulting in osmotic stress [94]. Plants have osmotic stress sensors to monitor osmotic stress and convert it into Ca2+ signals. Ca2+ signals are interpreted by Ca2+ binding proteins, which govern the plant’s adaptive responses to osmotic stress [70,95,96]. OSCA1 (Reduced hyperosmolality induced [Ca2+]i increase 1) is a Ca2+ channel that is regulated by high osmotic pressure in Arabidopsis. It selectively responds to exogenous osmotic stress as a receptor [95]. In addition, maintaining Na+ homeostasis at the cellular level is a crucial aspect of plant saline tolerance [77,97]. Na+ in soil mostly enters plant root cells through non-selective cation channels or high-affinity K+ transporters [97]. Significant progress has been made in the identification of MOCA1 (monocation-induced [Ca2+]i increases 1), which is expected to operate in extracellular salt sensing, including but not limited to Na+ ions [98]. The early reaction to Ca2+ waves that occurs in response to Na+, K+, or Li+ ions is absent in the moca1 mutant. In conclusion, Ca2+ should be the downstream signal molecule after saline stress perception. Ca2+ is also closely related to plant immunity and SA signaling. For example, a Ca2+/calmodulin-binding transcription factor AtSR1 directly interacted with the promoter and regulated the expression of EDS1 to modulate the SA content [99].
Several critical components that regulate Na+ homeostasis have been identified in plants. The function of the HKT1 (high-affinity K+ transporter) family of Na+ transporters is mainly to mediate the unloading of Na+ in xylem vessels, reduce the Na+ transport from the roots to shoots, lower the Na+ content in the shoots, and increase the saline tolerance [100]. NPR1-mediated SA signaling has been argued to be pivotal for controlling Na+ entry into the roots and the subsequent long-distance transport into the shoots, enhancing the H+-ATPase activity in the roots and increasing the K+ concentration in the shoots during saline stress [50]. The same study also reported that high endogenous-SA mutant nudt7 decreased the shoot Na+ concentration during prolonged saline stress [50].
The Na+/H+ reverse transporter NHXs (Na+/H+ exchanger 1), which are found on the vacuole membrane, store excess Na+ in the vacuole through ion regionalization, lowering the Na+ levels in the cytoplasm [101]. AtSOS1 is the principal Na+/H+ reverse transporter in Arabidopsis, and its transport activity is controlled by the proton transmembrane gradient maintained by plasma membrane H+-ATPase. Saline tolerance can be considerably increased by the overexpression of AtSOS1 [102]. In the tomato plant, NHX1 and SOS1 transcripts were found to be significantly altered in SA-treated plants; SA regulated the ion transporter transcription and maintained ion homeostasis [103].
When saline stress occurs, the intracellular free Ca2+ concentration increases instantly. The Ca2+ signal is detected by the Ca2+ binding protein SOS3, which stimulates the association of SOS3 with serine/threonine protein kinase SOS2. Then, the activated SOS2 can phosphorylate and activate SOS1 to promote the maintenance of Na+ homeostasis and saline tolerance [4]. The function of the SOS pathway has been described in numerous species, such as rice and barley, implying that the SOS system-mediated regulation of Na+ homeostasis is conserved [66]. These processes work together to improve plant saline tolerance by regulating Na+ homeostasis at the tissue and cellular levels [77].
Potassium (K) is an important mineral for plants. It is intertwined with a variety of vital daily activities. The content of K+ in the cytoplasm of Arabidopsis roots reduced quickly after they were exposed to salt. A plant’s saline tolerance can be considerably improved by increasing the intracellular K+ levels [103]. K+ uptake and efflux are mediated by K+ transporters (such as AKT1, Arabidopsis K+ transporter 1) or KORC (potassium outward rectifying channels) in plants. These genes are regulated by the transmembrane potential gradient and play a role in the steady-state maintenance of intracellular K+. Saline stress reduces the intracellular K+ concentration by destroying the transmembrane potential, causing membrane depolarization and inhibiting K+ transporter activity [104]. Ion transporters of the HAK (High-affinity K+)/KUP (K+ uptake permease)/KT family were initially discovered through high-affinity potassium transport abilities in fungi. They are involved in root K+ absorption and long-distance K+ transport from root to shoot in Arabidopsis and rice [105]. AtKUP, OsHAK1, OsHAK5, and OsAKT1 govern the long-distance transport of K+ by participating in K+ loading in the xylem, and then altering the K+ levels in shoots and the plant saline tolerance in rice or Arabidopsis [106]. Interestingly, they are also involved in the regulation of Na+ unloading in the xylem. For instance, ZmHAK4 may limit Na+ transport from the roots to shoots by moving Na+ in the xylem to the adjacent parenchyma cells, hence increasing salt resistance in maize [107]. SA can restore the membrane potential and limit salt-induced potassium efflux via the GORK (GATED OUTWARDLY-RECTIFYING K+ CHANNEL) channel, resulting in saline tolerance [23,71]. SA priming can be an important strategy for enhancing major GSH-based H2O2-metabolizing enzymes, such as GST. To this end, SA mitigated salinity stress injury in Solanum lycopersicum by causing characteristic changes in the expression pattern of GST- family members, such as SlGSTT2, SlGSTL3, and SlGSTF4 [108]. In another example, SA can regulate the generation of NaCl-induced oxygen and nitrogen reactive species in rice to induce the upregulation of OsGSNOR and increase the SNO content, resulting in considerable biomass accumulation under stress conditions [109,110].
6. Mechanism Comparison of SA in Saline and Biotic Stress
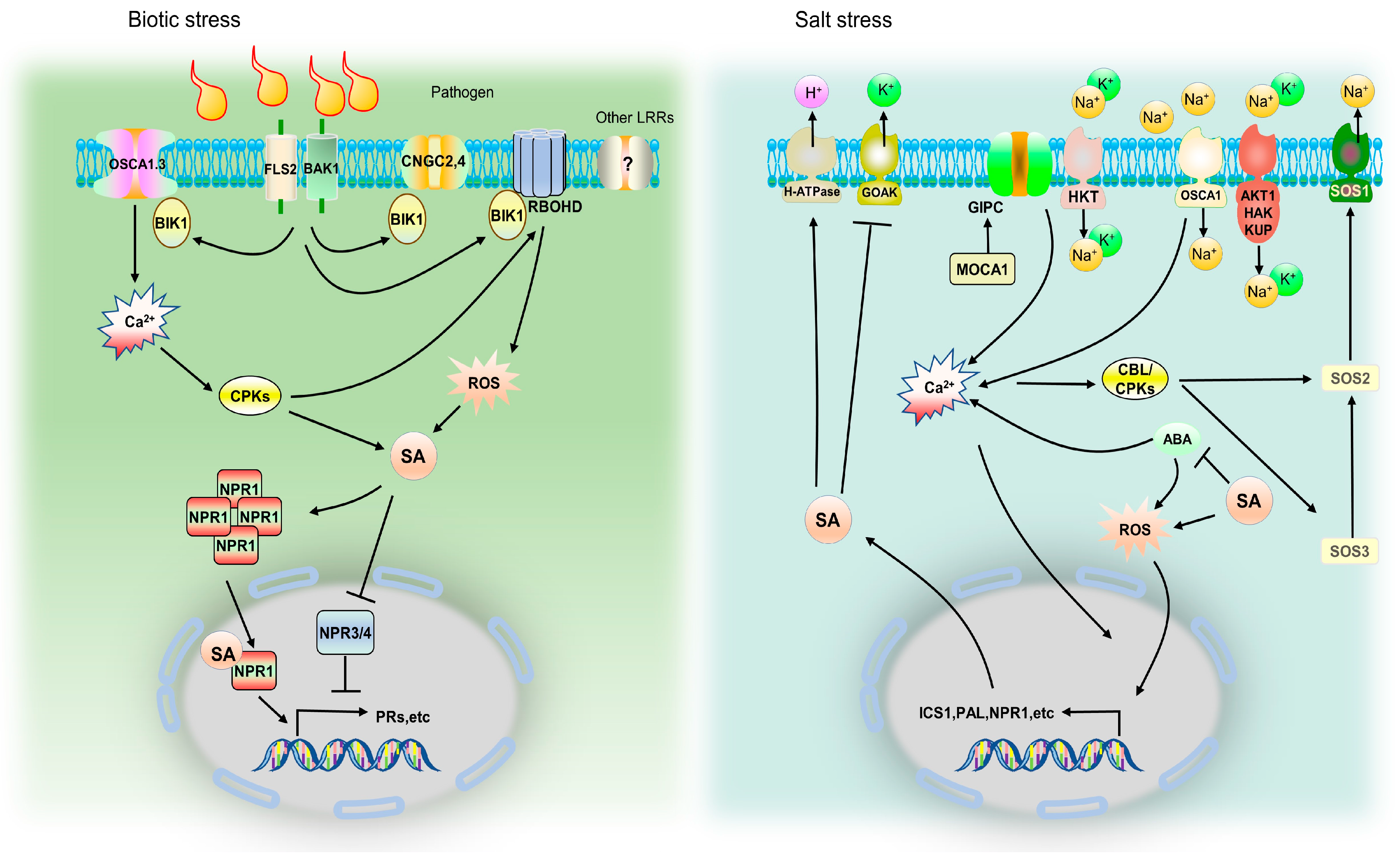
As a signal molecule, SA governs plant responses to both biotic and abiotic stress via a complex signal transduction network. In the molecular signaling pathway, plants share several key components, including Ca2+-driven and ROS-driven signals, among saline and pathogen infections. These two signals are regarded as the upstream of the SA pathway. However, there are differences in their specific mechanisms. For signal perception, studies have revealed that Ca2+ signal channels regulate the immune process, such as CNGC2, CNGC4, and OSCA1.3 [111,112], triggering a cascade of downstream immunological responses to connect with the SA pathway. Previously, glycosyl inositol phosphorylceramide (GIPC) sphingolipids generated by MOCA1 were shown to mix with monovalent cations and cause Ca2+ influx to respond to salinization; however, the Ca2+ channel that is involved has yet to be identified [98]. It is vital to note that the calcium channels that plants employ to detect biological stress and saline stress are distinctive.
Some of the described responses depend on cytosolic Ca2+ concentrations [9]. SOS3/CBL4 perhaps function as the signal hub of these two stresses. Ca2+ is detected by SOS3/CBL4 in conjunction with SOS2/CIPK24. This mechanism is supplemented by CBL10, which can form a complex with SOS2, which is involved in Na+ chelation in vacuoles [113]. In addition, the SOS3/CBL4-CIPK6 complex could regulate K+ allocation, and the cipk6 mutant was found to enhance disease resistance with increased SA accumulation [114]. Normally, several calcineurin B-like proteins (CBL) bind Ca2+ and promote protein phosphorylation via contact with CBL interacting protein kinase (CIPK) to regulate the output of Ca2+ flow, thereby adjusting the salt or immune response, respectively [115,116], such as CPK3, which phosphorylates the vacuolar K+ channel TPK1 (two pore K+ channel 1). In agreement with that, both cpk3 and tpk1 mutants were sensitive to salt [117]. The CBL2/CBL3-CIPK3/CIPK9/CIPK23/CIPK26 pathways enhance salt tolerance by regulating ion homeostasis, such as Ca2+ and K+ [114]. On the other hand, some CBL-CIPK modules act as regulator of pathogen-mediated programmed cell death (PCD) and SA transduction. SlCBL10-SlCIPK6 and TaCBL4-TaCIPK5 participate in plant immunity through ROS content alteration and downstream SA signals in tomato and wheat, respectively [118,119]. Enhanced resistance in Arabidopsis cipk14 mutants was also observed, which could be explained by a rise in SA accumulation and the expression of defense-related genes (PR1, EDS1, EDS5, ICS1) [120]. Parts of CBLs or CIPKs still need to have their biological functions clarified, particularly when compared functionally under saline or biological stress. This information could fill in the knowledge gaps in the SA signaling network.
Both saline stress and biotic stresses rapidly produce elevated levels of extracellular ROS molecules, such as hydrogen peroxide, singlet oxygen, superoxide, and hydroxyl radicals, which destroy redox homeostasis and cause oxidative damage to plant cells [121]. Both ROS and SA were found to be involved in defense signaling and the regulation of cell death. Some PAMP molecules trigger immunity (PTI) through phosphorylated RBOHD to induce reactive oxygen species (ROS) production [122], and H2O2-mediated SA accumulation could in part be explained by the catalytic activity of H2O2 on the BA2H enzyme involved in the conversion of benzoic acid to SA [31,123]. In Arabidopsis, the expression patterns of RBOH genes change dynamically. They produce ROS waves under saline stress within 24 h [124], indicating that the complex ROS production network is continuously active and plays a central role in the early salt response; the subsequently produced SA may be involved in the oxidative stress response of plants through its antioxidant activity (Figure 2).
The fundamental mechanism by which SA lessens salt damage may be its capacity to prevent K+ efflux through GORK [71]. To preserve the beneficial link between K+ retention in plant roots and salt tolerance, SA can reduce the effects of salinity and increase the K+ concentration in the roots [104]. The plasma membrane proton pump (H+-ATPase) activity, which plays a crucial role in regulating GORK function, is also lacking in the npr1 mutant, suggesting that SA-increased H+-ATPase activity is NPR1-dependent [50]. NPR1, the primary SA receptor protein, is important for many physiological processes as well as different types of plant disease resistance. It is interesting to note that saline stress causes rapid accumulation of the NPR1 protein in the chloroplast stroma, which prevents stress from causing a reduction in the photosynthetic capacity in a redox-dependent manner [125,126]. This suggests that the NPR1 protein performs an unidentified function in the response to saline stress.
Several transporters contribute to Na+ absorption and efflux during saline stress (HKT, AKT1, GOAK, OSCA1, etc.). The membrane depolarizes when Na+ enters the cell. Since many K+ and Ca2+ channels are voltage-regulated, this shift in membrane voltage affects ion transport. H+-ATPase action can repolarize the membrane. Na+ binding to Glycosyl-Inositol-Phosphoryl-Ceramides (GIPCs) in the plasma membrane is required for the formation of saline stress-induced Ca2+ signals. Extracellular Na+ directly binds to negatively charged GIPCs (catalytic production by MOCA1) in the plasma membrane’s outer leaflet under saline stress. Na+-bound GIPCs may directly activate/regulate Ca2+ channels. The Ca2+ sensor protein CBL4 (SOS3) decodes the saline stress-induced Ca2+ signal by activating CIPK24 (SOS2), which then phosphorylates and activates SOS1 to pump excess Na+ out of the cell. The osmotic and saline stress-sensing components, OSCA1 and GIPCs, have a direct influence on the formation of the major osmotic/salt stress-induced Ca2+ signal; however, this is questionable in the case of AKT1 (K+). For AKT1, a regulatory function for Ca2+-dependent phosphorylation via CBL1/9-CIPK23 has been identified. Meanwhile, for OSCA channels, kinase-dependent regulation has been characterized; however, no evidence of a direct relationship with Ca2+ has yet been revealed. The plant hormone ABA can similarly influence SA synthesis by promoting the production of Ca2+ and ROS. Simultaneously, SA feedback influences ABA. SA increases the activity of H+-ATPases via NPR1 and decreases K+ leakage via the GORK channel. SA also modulates other salt response responses via unknown mechanisms (probably through NPR1).
7. Summary and Future Prospects
SA is a hormone that plays a role in a plant’s reaction to biological stress. Plant response to saline stress has been linked to a variety of genes involved in protein synthesis and signal transmission, including SA-related genes such as ICS1, EDS5, PAL, and NPR1 [18,125]. However, it is unclear whether these reactions are triggered by the SA signal feedback control or the gene’s function. Saline stress response has also been linked to transcription factors downstream of SA, such as TGAs; however, the molecular mechanism remains unclear [127]. In fact, in trying to improve plant resistance to biological stress or saline stress, applying different amounts of SA to various plant species will result in contrasting outcomes. This could be due to the various treatment methods used, such as foliar spray or hydroponic culture. However, the response mechanism to SA may differ between plant species. In general, low concentrations or brief applications of SA can improve plant tolerance to abiotic stress, whereas high concentrations or the continuous application of SA can inhibit plant growth and reduce stress tolerance [128]. In any case, the application of SA holds a lot of promise.
The signaling mechanism of SA under biotic stress is generally obvious. It is unknown how SA plays a specialized function in saline stress and how each stress differentiates the accumulation of SA. If we can mechanistically analyze how plants discriminate the induction of SA by each stress and identify the promoters specifically induced by salt stress in the SA synthesis pathway, we will be able to better understand the specificity of plant responses to biotic or saline stress and apply SA more effectively.
Unlike drought, cold, heat, and other systemic stresses, a plant’s sensing of salt is entirely dependent on plant roots. With the rapid advancement of microbiome research, we should emphasize that plant saline tolerance is not only derived from the plant genome but also from symbiotic microbes, which may have a substantial impact. Rhizosphere bacteria can encourage plants to create systemic tolerance, regulate plant physiological responses, and ensure proper growth in high-salt environments [129]. Stress may also affect plant physiology and immunology, as well as soil parameters (PH, O2, and nutrient levels), by changing the content and concentration of root exudates and increasing the enrichment of certain microbes in the rhizosphere [130]. Many rhizosphere bacteria can produce SA [131] to control the microbial colonization of certain bacterial groups in the roots. In Arabidopsis root flora, there are considerable variations between SA synthesis and wild-type mutants; however, it is unknown how SA production attracts some bacteria while blocking others. Some selective microbes may employ SA as their carbon source [132]. Understanding the relevant mechanisms regarding how SA promotes the formation of certain microbial communities in the plant rhizosphere to promote saline stress tolerance is also a favorite topic for future study.
Author Contributions
W.Y. and Z.Z. collected the literature; W.Y. and Z.C. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of China (32272028) to W.Y., the Key Research and Development Program of Hubei Province (2022BFE003) to Z.C., and the Science and Technology Innovation Team of Hubei Province to Z.C. The APC was funded by the Shandong Modern Agricultural Technology and Industry system (SDAIT-17-06) to Z.C. and W.Y.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Haifeng Liu from Shandong Agricultural University for his comments on the earlier version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Fritschi, F.B.; Zandalinas, S.I.; Mittler, R. The impact of stress combination on reproductive processes in crops. Plant Sci. 2021, 311, 111007. [Google Scholar] [CrossRef] [PubMed]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Verma, K.; Agrawal, S.B. Salicylic acid-mediated defence signalling in respect to its perception, alteration and transduction. In Salicylic Acid: A Multifaceted Hormone; Springer: Singapore, 2017; pp. 97–122. [Google Scholar]
- Pokotylo, I.; Hodges, M.; Kravets, V.; Ruelland, E. A ménage à trois: Salicylic acid, growth inhibition, and immunity. Trends Plant Sci. 2021, 27, 460–471. [Google Scholar] [CrossRef]
- Khan, M.I.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic acid: Biosynthesis and signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Klessig, D.F.; Tian, M.; Choi, H.W. Multiple targets of salicylic acid and its derivatives in plants and animals. Front. Immunol. 2016, 7, 206. [Google Scholar] [CrossRef]
- Miura, K.; Tada, Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Iqbal, M.; Raza, S.H.; Iqbal, N. Efficacy of seed preconditioning with salicylic and ascorbic acid in increasing vigor of rice (Oryza sativa L.) seedling. Pak. J. Bot. 2013, 45, 157–162. [Google Scholar]
- Naser Alavi, S.M.; Arvin, M.J.; Manoochehri Kalantari, K. Salicylic acid and nitric oxide alleviate osmotic stress in wheat (Triticum aestivum L.) seedlings. J. Plant Interact. 2014, 9, 683–688. [Google Scholar] [CrossRef]
- Khan, M.I.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Effects of exogenously applied salicylic acid and putrescine alone and in combination with rhizobacteria on the phytoremediation of heavy metals and chickpea growth in sandy soil. Int. J. Phytoremediat. 2018, 20, 405–414. [Google Scholar] [CrossRef]
- Snyman, M.; Cronje, M.J. Modulation of heat shock factors accompanies salicylic acid-mediated potentiation of Hsp70 in tomato seedlings. J. Exp. Bot. 2008, 59, 2125–2132. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S.A. Effect of salicylic acid and sodium nitroprusside on growth parameters, photosynthetic pigments and secondary metabolites of safflower under drought stress. Sci. Hortic. 2020, 259, 108823. [Google Scholar] [CrossRef]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca2+ signaling. Curr. Biol. 2018, 28, 666–675. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Jones, J.D.G.; Ding, P. Plant immune networks. Trends Plant Sci. 2022, 27, 255–273. [Google Scholar] [CrossRef]
- Strawn, M.A.; Marr, S.K.; Inoue, K.; Inada, N.; Zubieta, C.; Wildermuth, M.C. Arabidopsis isochorismate synthase functional in pathogen-induced salicylate biosynthesis exhibits properties consistent with a role in diverse stress responses. J. Biol. Chem. 2007, 282, 5919–5933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Serrano, M.; Wang, B.; Aryal, B.; Garcion, C.; Abou-Mansour, E.; Heck, S.; Geisler, M.; Mauch, F.; Nawrath, C.; Metraux, J.P. Export of salicylic acid from the chloroplast requires the multidrug and toxin extrusion-like transporter EDS5. Plant Physiol. 2013, 162, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; van der Drift, K.M.; Olsson, P.E.; Thomas-Oates, J.E.; van Loon, L.C.; Bakker, P.A. Analysis of the pmsCEAB gene cluster involved in biosynthesis of salicylic acid and the siderophore pseudomonine in the biocontrol strain Pseudomonas fluorescens WCS374. J. Bacteriol. 2001, 183, 1909–1920. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Bobokalonova, A.; Carballo, V.; Glinkerman, C.M.; Pluskal, T.; Shen, A.; Weng, J.K. PBS3 and EPS1 complete salicylic acid biosynthesis from isochorismate in Arabidopsis. Mol. Plant 2019, 12, 1577–1586. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Yalpani, N.; León, J.; Lawton, M.A.; Raskin, I. Pathway of salicylic acid biosynthesis in healthy and virus-inoculated tobacco. Plant Physiol. 1993, 103, 315–321. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, W.; Zhao, Q. Salicylic acid biosynthesis is not from phenylalanine in Arabidopsis. J. Integr. Plant Biol. 2022, 1–7. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, H.; Ruan, W.; Deng, M.; Wang, F.; Peng, J.; Luo, J.; Chen, Z.; Yi, K. ABNORMAL INFLORESCENCE MERISTEM1 functions in salicylic acid biosynthesis to maintain proper reactive oxygen species levels for root meristem activity in rice. Plant Cell 2017, 29, 560–574. [Google Scholar] [CrossRef]
- Leon, J.; Shulaev, V.; Yalpani, N.; Lawton, M.A.; Raskin, I. Benzoic acid 2-hydroxylase, a soluble oxygenase from tobacco, catalyzes salicylic acid biosynthesis. Proc. Natl. Acad. Sci. USA 1995, 92, 10413–10417. [Google Scholar] [CrossRef] [Green Version]
- Sawada, H.; Shim, I.S.; Usui, K. Induction of benzoic acid 2-hydroxylase and salicylic acid biosynthesis modulation by salt stress in rice seedlings. Plant Sci. 2006, 171, 263–270. [Google Scholar] [CrossRef]
- Duan, L.; Liu, H.; Li, X.; Xiao, J.; Wang, S. Multiple phytohormones and phytoalexins are involved in disease resistance to Magnaporthe oryzae invaded from roots in rice. Physiol. Plant 2014, 152, 486–500. [Google Scholar] [CrossRef]
- Shine, M.B.; Yang, J.W.; El-Habbak, M.; Nagyabhyru, P.; Fu, D.Q.; Navarre, D.; Ghabrial, S.; Kachroo, P.; Kachroo, A. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean. New Phytol. 2016, 212, 627–636. [Google Scholar] [CrossRef]
- Huang, W.; Wang, Y.; Li, X.; Zhang, Y. Biosynthesis and regulation of salicylic acid and N-hydroxypipecolic acid in plant immunity. Mol. Plant 2020, 13, 31–41. [Google Scholar] [CrossRef]
- Wang, X.; Gao, J.; Zhu, Z.; Dong, X.; Wang, X.; Ren, G.; Zhou, X.; Kuai, B. TCP transcription factors are critical for the coordinated regulation of isochorismate synthase 1 expression in Arabidopsis thaliana. Plant J. 2015, 82, 151–162. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z.; et al. Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18220–18225. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, Y.; Li, Y.; Zhang, Q.; Ding, Y.; Zhang, Y. ChIP-seq reveals broad roles of SARD1 and CBP60g in regulating plant immunity. Nat. Commun. 2015, 6, 10159. [Google Scholar] [CrossRef]
- Sun, T.; Busta, L.; Zhang, Q.; Ding, P.; Jetter, R.; Zhang, Y. TGACG-BINDING FACTOR 1 (TGA1) and TGA4 regulate salicylic acid and pipecolic acid biosynthesis by modulating the expression of SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1 (SARD1) and CALMODULIN-BINDING PROTEIN 60g (CBP60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Li, L.S.; Ying, J.; Li, E.; Ma, T.; Li, M.; Gong, L.M.; Wei, G.; Zhang, Y.; Li, S. Arabidopsis CBP60b is a central transcriptional activator of immunity. Plant Physiol. 2021, 186, 1645–1659. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wu, Z.; Tian, H.; Li, X.; Zhang, Y. Arabidopsis CALMODULIN-BINDING PROTEIN 60b plays dual roles in plant immunity. Plant Commun. 2021, 2, 100213. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Y.; Spivey, N.W.; Zeng, W.; Liu, P.P.; Fu, Z.Q.; Klessig, D.F.; He, S.; Dong, X. Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation. Cell Host Microbe 2012, 11, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Truman, W.; Sreekanta, S.; Lu, Y.; Bethke, G.; Tsuda, K.; Katagiri, F.; Glazebrook, J. The CALMODULIN-BINDING PROTEIN60 family includes both negative and positive regulators of plant immunity. Plant Physiol. 2013, 163, 1741–1751. [Google Scholar] [CrossRef]
- Chandran, D.; Rickert, J.; Huang, Y.; Steinwand, M.A.; Marr, S.K.; Wildermuth, M.C. Atypical E2F transcriptional repressor DEL1 acts at the intersection of plant growth and immunity by controlling the hormone salicylic acid. Cell Host Microbe 2014, 15, 506–513. [Google Scholar] [CrossRef]
- Lai, Y.S.; Renna, L.; Yarema, J.; Ruberti, C.; He, S.Y.; Brandizzi, F. Salicylic acid-independent role of NPR1 is required for protection from proteotoxic stress in the plant endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2018, 115, E5203–E5212. [Google Scholar] [CrossRef]
- Mhamdi, A. NPR1 has everything under control. Plant Physiol. 2019, 181, 6–7. [Google Scholar] [CrossRef]
- Zavaliev, R.; Mohan, R.; Chen, T.; Dong, X. Formation of NPR1 condensates promotes cell survival during the plant immune response. Cell 2020, 182, 1093–1108. [Google Scholar] [CrossRef]
- Liu, Q.; Ning, Y.; Zhang, Y.; Yu, N.; Zhao, C.; Zhan, X.; Wu, W.; Chen, D.; Wei, X.; Wang, G.L.; et al. OsCUL3a negatively regulates cell death and immunity by degrading OsNPR1 in rice. Plant Cell 2017, 29, 345–359. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Shabala, S.; Massart, A.; Poschenrieder, C.; Rengel, Z. The NPR1-dependent salicylic acid signalling pathway is pivotal for enhanced salt and oxidative stress tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 1865–1875. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, T.; Sun, Y.; Zhang, Y.; Radojicic, A.; Ding, Y.; Tian, H.; Huang, X.; Lan, J.; Chen, S.; et al. Diverse roles of the salicylic acid receptors NPR1 and NPR3/NPR4 in plant immunity. Plant Cell 2020, 32, 4002–4016. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Withers, J.; Li, H.; Zwack, P.J.; Rusnac, D.V.; Shi, H.; Liu, L.; Yan, S.; Hinds, T.R.; Guttman, M.; et al. Structural basis of salicylic acid perception by Arabidopsis NPR proteins. Nature 2020, 586, 311–316. [Google Scholar] [CrossRef]
- Choudhary, A.; Senthil-Kumar, M. Drought attenuates plant defence against bacterial pathogens by suppressing the expression of CBP60g/SARD1 during combined stress. Plant Cell Environ. 2022, 45, 1127–1145. [Google Scholar] [CrossRef]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Ding, P.; Ding, Y. Stories of salicylic acid: A plant defense hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Lapin, D.; Bhandari, D.D.; Parker, J.E. Origins and immunity networking functions of EDS1 family proteins. Annu. Rev. Phytopathol. 2020, 58, 253–276. [Google Scholar] [CrossRef]
- Wagner, S.; Stuttmann, J.; Rietz, S.; Guerois, R.; Brunstein, E.; Bautor, J.; Niefind, K.; Parker, J.E. Structural basis for signaling by exclusive EDS1 heteromeric complexes with SAG101 or PAD4 in plant innate immunity. Cell Host Microbe 2013, 14, 619–630. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- Chang, M.; Zhao, J.; Chen, H.; Li, G.; Chen, J.; Li, M.; Palmer, A.; Song, J.; Alfano, J.; Liu, F.; et al. PBS3 protects EDS1 from proteasome-mediated degradation in plant immunity. Mol. Plant 2019, 12, 678–688. [Google Scholar] [CrossRef]
- Zewdu, S.; Suryabhagavan, K.V.; Balakrishnan, M. Land-use/land-cover dynamics in Sego Irrigation Farm, southern Ethiopia: A comparison of temporal soil salinization using geospatial tools. J. Saudi Soc. Agric. Sci. 2016, 15, 91–97. [Google Scholar] [CrossRef]
- Abbas, A.; Khan, S.; Hussain, N.; Hanjra, M.A.; Akbar, S. Characterizing soil salinity in irrigated agriculture using a remote sensing approach. Phys. Chem. Earth Parts A/B/C 2013, 55–57, 43–52. [Google Scholar] [CrossRef]
- Li, P.; Wu, J.; Qian, H. Regulation of secondary soil salinization in semi-arid regions: A simulation research in the Nanshantaizi area along the Silk Road, northwest China. Environ. Earth Sci. 2016, 75, 698. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Chandra, A.; Anand, A.; Dubey, A. Effect of salicylic acid on morphological and biochemical attributes in cowpea. J. Environ. Biol. 2007, 28, 193–196. [Google Scholar]
- Jini, D.; Joseph, B. Physiological mechanism of salicylic acid for alleviation of salt stress in rice. Rice Sci. 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid improves salinity tolerance in Arabidopsis by restoring membrane potential and preventing salt-induced K+ loss via a GORK channel. J. Exp. Bot. 2013, 64, 2255–2268. [Google Scholar] [CrossRef]
- Duarte, B.; Santos, D.; Marques, J.C.; Cacador, I. Ecophysiological adaptations of two halophytes to salt stress: Photosynthesis, PS II photochemistry and anti-oxidant feedback-implications for resilience in climate change. Plant Physiol. Biochem. 2013, 67, 178–188. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef] [Green Version]
- Nadarajah, K.; Abdul Hamid, N.W.; Abdul Rahman, N.S.N. SA-mediated regulation and control of abiotic stress tolerance in rice. Int. J. Mol. Sci. 2021, 22, 5591. [Google Scholar] [CrossRef]
- Saeidnejad, A.H.; Mardani, H.; Naghibolghora, M. Protective effects of salicylic acid on physiological parameters and antioxidants response in maize seedlings under salinity stress. J. Appl. Environ. Biol. Sci. 2012, 2, 364–373. [Google Scholar]
- Li, G.; Peng, X.; Wei, L.; Kang, G. Salicylic acid increases the contents of glutathione and ascorbate and temporally regulates the related gene expression in salt-stressed wheat seedlings. Gene 2013, 529, 321–325. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a beneficial macronutrient in higher plants: New roles and regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef]
- Syeed, S.; Sehar, Z.; Masood, A.; Anjum, N.A.; Khan, N.A. Control of elevated ion accumulation, oxidative stress, and lipid peroxidation with salicylic acid-induced accumulation of glycine betaine in salinity-exposed Vigna radiata L. Appl. Biochem. Biotech. 2021, 193, 3301–3320. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Manuka, R.; Penna, S. Plant sugars: Homeostasis and transport under abiotic stress in plants. Physiol. Plant 2021, 171, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, W.F.; Soares, L.H.; Fagan, E.B.; da Costa Mello, S.; Reichardt, K.; Dourado-Neto, D. Amino acids as stress reducers in soybean plant growth under different water-deficit conditions. J. Plant Growth Regul. 2020, 39, 905–919. [Google Scholar] [CrossRef]
- Misra, N.; Saxena, P. Effect of salicylic acid on proline metabolism in lentil grown under salinity stress. Plant Sci. 2009, 177, 181–189. [Google Scholar] [CrossRef]
- El-Tayeb, M.A. Response of barley grains to the interactive e.ect of salinity and salicylic acid. Plant Growth Regul. 2005, 45, 215–224. [Google Scholar] [CrossRef]
- Tufail, A.; Arfan, M.; Gurmani, A.R.; Khan, A.; Bano, A. Salicylic acid induced salinity tolerance in maize (Zea mays). Pak. J. Bot. 2013, 45, 75–82. [Google Scholar]
- Al-Whaibi, M.H.; Siddiqui, M.H.; Basalah, M.O. Salicylic acid and calcium-induced protection of wheat against salinity. Protoplasma 2012, 249, 769–778. [Google Scholar] [CrossRef]
- Jini, D.; Joseph, B. Salicylic acid mediated salt tolerance at different growth stages of Oryza sativa L. and its effect on salicylic acid biosynthetic pathway genes. Biotechnol. Ind. J. 2017, 13, 134. [Google Scholar]
- Nie, W.; Gong, B.; Chen, Y.; Wang, J.; Wei, M.; Shi, Q. Photosynthetic capacity, ion homeostasis and reactive oxygen metabolism were involved in exogenous salicylic acid increasing cucumber seedlings tolerance to alkaline stress. Sci. Hortic. 2018, 235, 413–423. [Google Scholar] [CrossRef]
- Gharbi, E.; Lutts, S.; Dailly, H.; Quinet, M. Comparison between the impacts of two different modes of salicylic acid application on tomato (Solanum lycopersicum) responses to salinity. Plant Signal. Behav. 2018, 13, e1469361. [Google Scholar] [CrossRef]
- Souana, K.; Taïbi, K.; Ait Abderrahim, L.; Amirat, M.; Achir, M.; Boussaid, M.; Mulet, J.M. Salt-tolerance in Vicia faba L. is mitigated by the capacity of salicylic acid to improve photosynthesis and antioxidant response. Sci. Hortic. 2020, 273, 109641. [Google Scholar] [CrossRef]
- Palma, F.; Lopez-Gomez, M.; Tejera, N.A.; Lluch, C. Salicylic acid improves the salinity tolerance of Medicago sativa in symbiosis with Sinorhizobium meliloti by preventing nitrogen fixation inhibition. Plant Sci. 2013, 208, 75–82. [Google Scholar] [CrossRef]
- Yastreb, T.O.; Kolupaev, Y.E.; Lugovaya, A.A.; Dmitriev, A.P. Hydrogen peroxide-induced salt tolerance in the Arabidopsis salicylate-deficient transformants NahG. Appl. Biochem. Microbiol. 2017, 53, 719–724. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Dong, Q.; Wallrad, L.; Almutairi, B.O.; Kudla, J. Ca2+ signaling in plant responses to abiotic stresses. J. Integr. Plant Biol. 2022, 64, 287–300. [Google Scholar]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhou, X.; Tao, M.; Yuan, F.; Liu, L.; Wu, F.; Wu, X.; Xiang, Y.; Niu, Y.; Liu, F.; et al. Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx. Nature 2019, 572, 341–346. [Google Scholar] [CrossRef]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef]
- Møller, I.S.; Gilliham, M.; Jha, D.; Mayo, G.M.; Roy, S.J.; Coates, J.C.; Haseloff, J.; Tester, M. Shoot Na+ exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na+ transport in Arabidopsis. Plant Cell 2009, 21, 2163–2178. [Google Scholar] [CrossRef]
- Bassil, E.; Zhang, S.; Gong, H.; Tajima, H.; Blumwald, E. Cation specificity of vacuolar NHX-type cation/H+ antiporters. Plant Physiol. 2019, 179, 616–629. [Google Scholar] [CrossRef]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.R.; Ansari, M.W.; Sahoo, R.K.; Wattal, R.K.; Tuteja, N.; Kumar, V.R. Salicylic acid modulates ACS, NHX1, sos1 and HKT1; 2 expression to regulate ethylene overproduction and Na+ ions toxicity that leads to improved physiological status and enhanced salinity stress tolerance in tomato plants cv. Pusa Ruby. Plant Signal. Behav. 2021, 16, 1950888. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant 2014, 151, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ transporters: Function and regulation. Semin. Cell Dev. Biol. 2018, 74, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, X.; Wang, L.; Cao, Y.; Song, W.; Shi, J.; Lai, J.; Jiang, C. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat. Plants 2019, 5, 1297–1308. [Google Scholar] [CrossRef]
- Csiszár, J.; Horvath, E.; Vary, Z.; Galle, A.; Bela, K.; Brunner, S.; Tari, I. Glutathione transferase supergene family in tomato: Salt stress-regulated expression of representative genes from distinct GST classes in plants primed with salicylic acid. Plant Physiol. Biochem. 2014, 78, 15–26. [Google Scholar] [CrossRef]
- Kim, Y.; Mun, B.G.; Khan, A.L.; Waqas, M.; Kim, H.H.; Shahzad, R.; Imran, M.; Yun, B.W.; Lee, I.J. Regulation of reactive oxygen and nitrogen species by salicylic acid in rice plants under salinity stress conditions. PLoS ONE 2018, 13, e0192650. [Google Scholar] [CrossRef]
- Rasheed, F.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. The key roles of salicylic acid and sulfur in plant salinity stress tolerance. J. Plant Growth Regul. 2022, 41, 1891–1904. [Google Scholar] [CrossRef]
- Tian, W.; Hou, C.; Ren, Z.; Wang, C.; Zhao, F.; Dahlbeck, D.; Hu, S.; Zhang, L.; Ni, Q.; Li, L.; et al. A calmodulin-gated calcium channel links pathogen patterns to plant immunity. Nature 2019, 572, 131–135. [Google Scholar] [CrossRef]
- Thor, K.; Jiang, S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.; Dindas, J.; Derbyshire, P.; Leitão, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef]
- Quan, R.; Lin, H.; Mendoza, I.; Zhang, Y.; Cao, W.; Yang, Y.; Shang, M.; Chen, S.; Pardo, J.M.; Guo, Y. SCABP8/CBL10, a putative calcium sensor, interacts with the protein kinase SOS2 to protect Arabidopsis shoots from salt stress. Plant Cell 2007, 19, 1415–1431. [Google Scholar] [CrossRef]
- Mao, J.; Mo, Z.; Yuan, G.; Xiang, H.; Visser, R.G.; Bai, Y.; Liu, H.; Wang, Q.; van der Linden, C.G. The CBL-CIPK network is involved in the physiological crosstalk between plant growth and stress adaptation. Plant Cell Environ. 2022, 1–11. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Ma, X.; Li, Q.H.; Yu, Y.N.; Qiao, Y.M.; Haq, S.U.; Gong, Z.H. The CBL-CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Latz, A.; Mehlmer, N.; Zapf, S.; Mueller, T.D.; Wurzinger, B.; Pfister, B.; Csaszar, E.; Hedrich, R.; Teige, M.; Becker, D. Salt stress triggers phosphorylation of the Arabidopsis vacuolar K+ channel TPK1 by calcium-dependent protein kinases (CDPKs). Mol. Plant 2013, 6, 1274–1289. [Google Scholar] [CrossRef] [Green Version]
- De la Torre, F.; Gutiérrez-Beltrán, E.; Pareja-Jaime, Y.; Chakravarthy, S.; Martin, G.B.; del Pozo, O. The tomato calcium sensor CBL10 and its interacting protein kinase CIPK6 define a signaling pathway in plant immunity. Plant Cell 2013, 25, 2748–2764. [Google Scholar] [CrossRef]
- Liu, P.; Duan, Y.; Liu, C.; Xue, Q.; Guo, J.; Qi, T.; Kang, Z.; Guo, J. The calcium sensor TaCBL4 and its interacting protein TaCIPK5 are required for wheat resistance to stripe rust fungus. J. Exp. Bot. 2018, 69, 4443–4457. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, Q.; He, J.; Cao, J.; Liu, Z.; Wang, J.; Yang, Y. The kinase CIPK14 functions as a negative regulator of plant immune responses to Pseudomonas syringae in Arabidopsis. Plant Sci. 2021, 312, 111017. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Chang, M.; Chen, H.; Liu, F.; Fu, Z.Q. PTI and ETI: Convergent pathways with diverse elicitors. Trends Plant Sci. 2021, 27, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Tak, Y.; Asthir, B. Salicylic acid: A key signal molecule ameliorating plant stresses. Cereal Res. Commun. 2022, 50, 617–626. [Google Scholar] [CrossRef]
- Xie, Y.J.; Xu, S.; Han, B.; Wu, M.Z.; Yuan, X.X.; Han, Y.; Gu, Q.; Xu, D.K.; Yang, Q.; Shen, W.B. Evidence of Arabidopsis salt acclimation induced by up-regulation of HY1 and the regulatory role of RbohD-derived reactive oxygen species synthesis. Plant J. 2011, 66, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.Y.; Wi, S.J.; Park, K.Y. Functional switching of NPR1 between chloroplast and nucleus for adaptive response to salt stress. Sci. Rep. 2020, 10, 4339. [Google Scholar] [CrossRef]
- Seo, S.Y.; Park, K.Y. Translocation of chloroplast NPR1 to the nucleus in retrograde signaling for adaptive response to salt stress in tobacco. BioRxiv 2021. [Google Scholar] [CrossRef]
- Li, B.; Liu, Y.; Cui, X.Y.; Fu, J.D.; Zhou, Y.B.; Zheng, W.J.; Lan, J.H.; Jin, L.G.; Chen, M.; Ma, Y.Z.; et al. Genome-wide characterization and expression analysis of soybean TGA transcription factors identified a novel TGA gene involved in drought and salt tolerance. Front. Plant Sci. 2019, 10, 549. [Google Scholar] [CrossRef] [Green Version]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef]
- Keswani, C.; Prakash, O.; Bharti, N.; Vilchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Ran, L.; Mercado-Blanco, J. Rhizobacterial salicylate production provokes headaches! Plant Soil. 2014, 382, 1–16. [Google Scholar] [CrossRef]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Rio, T.G.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The simplified transcriptional regulatory network for SA biosynthesis in Arabidopsis. The enzymes required for SA synthesis via the isochorismate (IC) pathway are encoded by the genes ICS1, EDS5, and PBS3. Transcription factors, including NTL9, TCP8/9/20, WRKY28, SARD1, and CBP60g, may positively regulate ICS1 gene expression; SARD1 and CBP60g also activate EDS5 and PBS3 gene expression (red arrow); and CBP60b and CHE control downstream genes by modulating SARD1 and CBP60g. TCP8/9 interact with WRKY28 and SARD1, which may play a role in their transcriptional control. CBP60a, WRKY54/70, ANAC019/055/072, and EIN3 negatively regulate ICS1 gene expression (blue line), while DEL1 suppresses EDS5 gene expression.
Figure 1.
The simplified transcriptional regulatory network for SA biosynthesis in Arabidopsis. The enzymes required for SA synthesis via the isochorismate (IC) pathway are encoded by the genes ICS1, EDS5, and PBS3. Transcription factors, including NTL9, TCP8/9/20, WRKY28, SARD1, and CBP60g, may positively regulate ICS1 gene expression; SARD1 and CBP60g also activate EDS5 and PBS3 gene expression (red arrow); and CBP60b and CHE control downstream genes by modulating SARD1 and CBP60g. TCP8/9 interact with WRKY28 and SARD1, which may play a role in their transcriptional control. CBP60a, WRKY54/70, ANAC019/055/072, and EIN3 negatively regulate ICS1 gene expression (blue line), while DEL1 suppresses EDS5 gene expression.
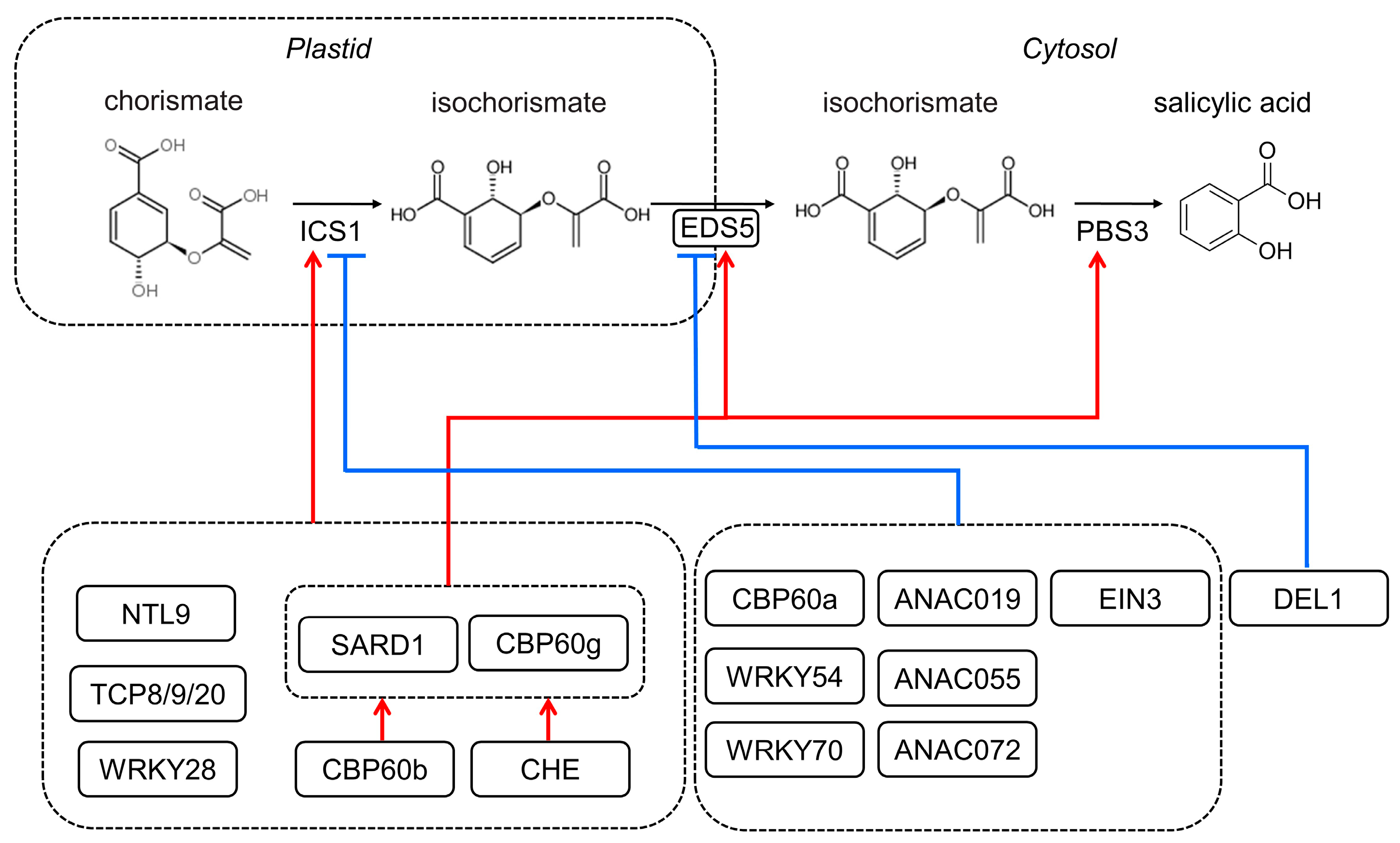
Figure 2.
The signaling pathway of salicylic acid mediated by pathogens and saline stress. Pathogen-associated molecular patterns (PAMPs) are recognized by some receptor-like kinases (RLKs, such as FLS2 and BAK1) and activate PAMP-triggered immunity (PTI). The binding of flg22 triggers the dissociation of BIK1 from the FLS2-BAK1 complex. Then, BIK1 phosphorylates RBOHD to induce reactive oxygen species (ROS) production. BIK1 also phosphorylates and activates the CaM-gated OSCA1.3 and CNGC2-CNGC4 channels, triggering Ca2+ to enter the cell. Ca2+ binding to EF-hand motifs in RBOHD and CDPKs activates RBOHD and boosts ROS production, hence propagating the defense signal and activating SA biosynthesis. In the SA signaling cascade, NONEXPRESSER OF PR GENES 1 (NPR1) monomerization requires SA binding to NPR1 as well as redox regulation and leads to the expression of pathogenesis-related (PR) genes via NPR1-TGA transcription factor interactions. NPR3 and NPR4 proteins (paralogs of NPR1) also bind SA while acting as transcriptional corepressors.
Figure 2.
The signaling pathway of salicylic acid mediated by pathogens and saline stress. Pathogen-associated molecular patterns (PAMPs) are recognized by some receptor-like kinases (RLKs, such as FLS2 and BAK1) and activate PAMP-triggered immunity (PTI). The binding of flg22 triggers the dissociation of BIK1 from the FLS2-BAK1 complex. Then, BIK1 phosphorylates RBOHD to induce reactive oxygen species (ROS) production. BIK1 also phosphorylates and activates the CaM-gated OSCA1.3 and CNGC2-CNGC4 channels, triggering Ca2+ to enter the cell. Ca2+ binding to EF-hand motifs in RBOHD and CDPKs activates RBOHD and boosts ROS production, hence propagating the defense signal and activating SA biosynthesis. In the SA signaling cascade, NONEXPRESSER OF PR GENES 1 (NPR1) monomerization requires SA binding to NPR1 as well as redox regulation and leads to the expression of pathogenesis-related (PR) genes via NPR1-TGA transcription factor interactions. NPR3 and NPR4 proteins (paralogs of NPR1) also bind SA while acting as transcriptional corepressors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, W.; Zhou, Z.; Chu, Z. Emerging Roles of Salicylic Acid in Plant Saline Stress Tolerance. Int. J. Mol. Sci. 2023, 24, 3388. https://doi.org/10.3390/ijms24043388
AMA Style
Yang W, Zhou Z, Chu Z. Emerging Roles of Salicylic Acid in Plant Saline Stress Tolerance. International Journal of Molecular Sciences. 2023; 24(4):3388. https://doi.org/10.3390/ijms24043388
Chicago/Turabian StyleYang, Wei, Zhou Zhou, and Zhaohui Chu. 2023. "Emerging Roles of Salicylic Acid in Plant Saline Stress Tolerance" International Journal of Molecular Sciences 24, no. 4: 3388. https://doi.org/10.3390/ijms24043388
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.