
Group V Chitin Deacetylases Influence the Structure and Composition of the Midgut of Beet Armyworm, Spodoptera exigua

 ,
,
Abstract
1. Introduction
2. Results
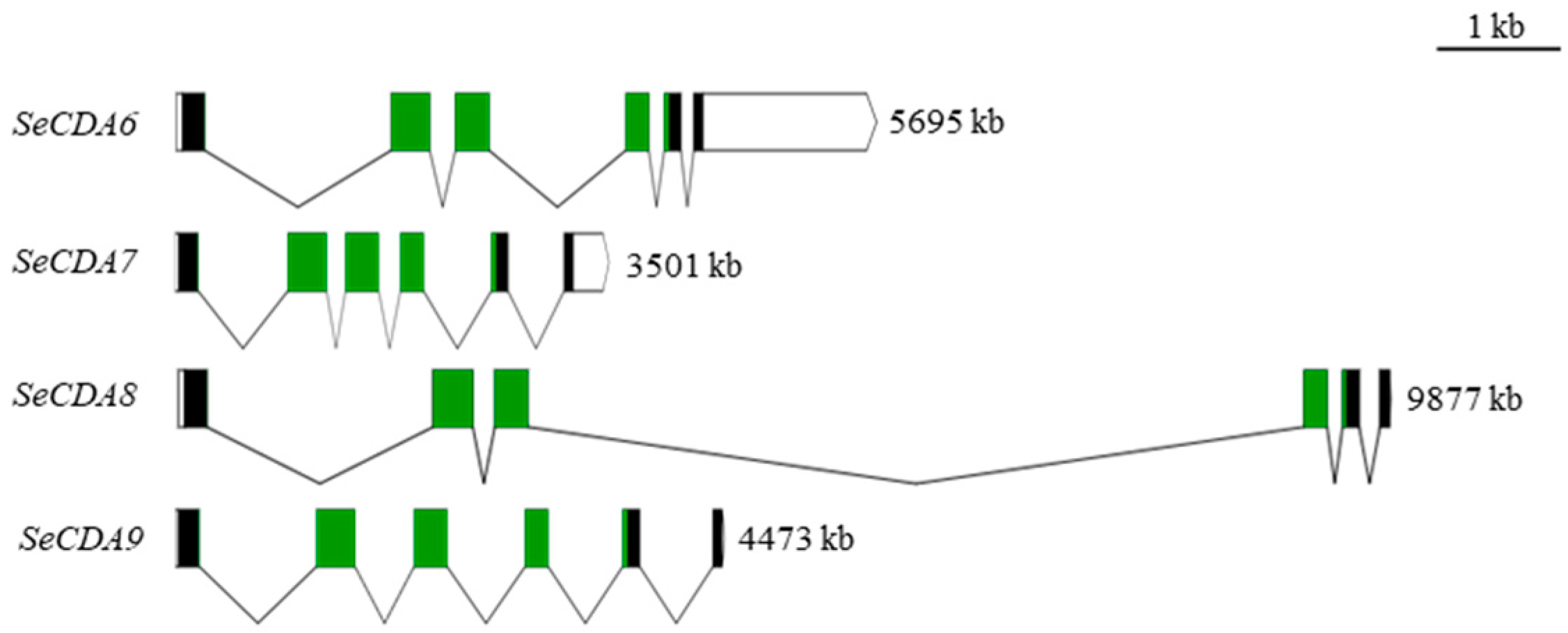
2.1. Group V SeCDAs Sequence Analysis
2.2. Group V SeCDAs Expression Analyses
2.3. Expression of Group V SeCDAs in Response to 20E and JHA Signaling
2.4. Functional Analysis of Group V SeCDAs
2.5. Immunohistochemistry
2.6. Scanning Electron Microscopy Analysis
3. Discussion
4. Materials and Methods
4.1. Insect Larvae
4.2. Sequence Analysis
4.3. Gene Expression Analyses
4.4. Preparation and Treatment with 20E and JHA
4.5. Functional Analysis of Group V CDA Genes
4.6. Immunohistochemistry
4.7. Scanning Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azadeh, K.M.; Yaghoub, F.; Ali, A.T.; Mohammad, B. Life Table Parameters and Survivorship of Spodoptera exigua (Lepidoptera: Noctuidae) at Constant Temperatures. Environ. Entomol. 2014, 43, 795–803. [Google Scholar]
- Zhang, W.J.; Dai, H.Q. A quantitative study on development, fecundity and mortality of beet armyworm, Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae), infected by SeMNPV. Arthropods 2018, 7, 26–30. [Google Scholar]
- Zheng, X.L.; Cong, X.P.; Wang, X.P.; Lei, C.L. A Review of Geographic Distribution, Overwintering and Migration in Spodoptera exigua Hübner (Lepidoptera: Noctuidae). J. Entomol. Res. Soc. 2011, 13, 39–48. [Google Scholar]
- Yonow, T.; Kriticos, D.J.; Kirichenko, N.; Ota, N. Considering biology when inferring range-limiting stress mechanisms for agricultural pests: A case study of the beet armyworm. J. Pest Sci. 2018, 91, 523–538. [Google Scholar] [CrossRef]
- Zuo, Y.Y.; Huang, J.L.; Wang, J.; Feng, Y.; Han, T.T.; Wu, Y.D.; Yang, Y.H. Knockout of a P-glycoprotein gene increases susceptibility to abamectin and emamectin benzoate in Spodoptera exigua. Insect Mol. Biol. 2018, 27, 36–45. [Google Scholar] [CrossRef]
- Sun, X.X.; Li, H.Y.; Jiang, Y.J.; Zhang, J.X.; Gu, H.L.; Gao, B.; Ma, J.J.; Wang, F.; Zhou, J.C.; Tian, X.R.; et al. Resistance Risk Evaluated by Metaflumizone Selection and the Effects on Toxicities Over Other Insecticides in Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol. 2019, 112, 2354–2361. [Google Scholar] [CrossRef]
- Wang, X.G.; Xiang, X.; Yu, H.L.; Liu, S.H.; Yin, Y.; Cui, P.; Wu, Y.Q.; Yang, J.; Jiang, C.X.; Yang, Q.F. Monitoring and biochemical characterization of beta-cypermethrin resistance in Spodoptera exigua (Lepidoptera: Noctuidae) in Sichuan Province, China. Pestic. Biochem. Phys. 2018, 146, 71–79. [Google Scholar] [CrossRef]
- Tsigos, I.; Martinou, A.; Kafetzopoulos, D.; Bouriotis, V. Chitin deacetylases: New, versatile tools in biotechnology. Trends Biotechnol. 2000, 18, 305–312. [Google Scholar] [CrossRef]
- Dixit, R.; Arakane, Y.; Specht, C.A.; Richard, C.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Domain organization and phylogenetic analysis of proteins from the chitin deacetylase gene family of Tribolium castaneum and three other species of insects. Insect Biochem. Mol. Biol. 2008, 38, 440–451. [Google Scholar] [CrossRef]
- Tetreau, G.; Cao, X.L.; Chen, Y.R.; Muthukrishnan, S.; Jiang, H.B.; Blissard, G.W.; Kanost, M.R.; Wang, P. Overview of chitin metabolism enzymes in Manduca sexta: Identification, domain organization, phylogenetic analysis and gene expression. Insect Biochem. Mol. Biol. 2015, 62, 114–126. [Google Scholar] [CrossRef]
- Xi, Y.; Pan, P.L.; Ye, Y.X.; Yu, B.; Zhang, C.X. Chitin deacetylase family genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Mol. Biol. 2014, 23, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Dixit, R.; Begum, K.; Park, Y.; Specht, C.A.; Merzendorfer, H.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Analysis of functions of the chitin deacetylase gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2009, 39, 355–365. [Google Scholar] [CrossRef]
- Guo, W.; Li, G.; Pang, Y.; Wang, P. A novel chitin-binding protein identified from the peritrophic membrane of the cabbage looper, Trichoplusia ni. Insect Biochem. Mol. Biol. 2005, 35, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Wu, H.; Yan, X.P.; Guo, W.; Lu, X.J.; Guo, X.C.; Liu, Z.R.; Li, Y.Z. Molecular cloning and functional characterization of two insect chitin deacetylases from the American white moth, Hyphantria cunea. J. Appl. Entomol. 2021, 145, 697–706. [Google Scholar] [CrossRef]
- Yan, X.P.; Zhao, D.; Zhang, Y.K.; Guo, W.; Wang, W.; Zhao, K.L.; Gao, Y.J.; Wang, X.Y. Identification and characterization of chitin deacetylase 2 from the American white moth, Hyphantria cunea (Drury). Gene 2018, 670, 98–105. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Yan, J.M.; Liu, Q.; Zhang, Y.H.; Gong, J.; Hou, Y. Genome-Wide Analysis and Hormone Regulation of Chitin Deacetylases in Silkworm. Int. J. Mol. Sci. 2019, 20, 1679. [Google Scholar] [CrossRef] [PubMed]
- Han, G.Y.; Li, X.M.; Zhang, T.; Zhu, X.T.; Li, J.G. Cloning and Tissue-Specific Expression of a Chitin Deacetylase Gene from Helicoverpa armigera (Lepidoptera: Noctuidae) and Its Response to Bacillus thuringiensis. J. Insect Sci. 2015, 15, 95. [Google Scholar] [CrossRef]
- Quan, G.; Ladd, T.; Duan, J.; Wen, F.; Doucet, D.; Cusson, M.; Krell, P.J. Characterization of a spruce budworm chitin deacetylase gene: Stage- and tissue-specific expression, and inhibition using RNA interference. Insect Biochem. Mol. Biol. 2013, 43, 683–691. [Google Scholar] [CrossRef]
- Yang, W.J.; Xu, K.K.; Yan, X.; Chen, C.X.; Cao, Y.; Meng, Y.L.; Li, C. Functional characterization of chitin deacetylase 1 gene disrupting larval–pupal transition in the drugstore beetle using RNA interference. Comp. Biochem. Physiol. B 2018, 219, 10–16. [Google Scholar] [CrossRef]
- Wu, J.J.; Chen, Z.C.; Wang, Y.W.; Fu, K.Y.; Guo, W.C.; Li, G.Q. Silencing chitin deacetylase 2 impairs larval–pupal and pupal-adult molts in Leptinotarsa decemlineata. Insect Mol. Biol. 2019, 28, 52–64. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, Z.R.; Wu, H.; Fu, C.R.; Li, Y.Z.; Lu, X.J.; Guo, W. RNA interference-mediated functional characterization of Group I chitin deacetylases in Holotrichia parallela Motschulsky. Pestic. Biochem. Physiol. 2021, 173, 104770. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.R.; Liu, W.M.; Zhao, X.M.; Zhang, M.; Li, D.Q.; Zuber, R.; Ma, E.B.; Zhu, K.Y.; Moussian, B.; Zhang, J.Z. LmCDA1 organizes the cuticle by chitin deacetylation in Locusta migratoria. Insect Mol. Biol. 2019, 28, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Li, H.F.; Yang, Y.; Yang, R.L.; Yang, W.J.; Jiang, H.B.; Dou, W.; Smagghe, G.; Wang, J.J. Genome-wide identification of chitinase and chitin deacetylase gene families in the oriental fruit fly, Bactrocera dorsalis (Hendel). Comp. Biochem. Physiol. D 2018, 27, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Luschnig, S.; Batz, T.; Armbruster, K.; Krasnow, M.A. Serpentine and vermiform encode matrix proteins with chitin binding and deacetylation domains that limit tracheal tube length in Drosophila. Curr. Biol. 2006, 16, 186–194. [Google Scholar] [CrossRef]
- Sandoval-Mojica, A.F.; Scharf, M.E. Gut genes associated with the peritrophic matrix in Reticulitermes flavipes (Blattodea: Rhinotermitidae): Identification and characterization. Arch. Insect Biochem. Physiol. 2016, 92, 127–142. [Google Scholar] [CrossRef]
- Toprak, U.; Baldwin, D.; Erlandson, M.; Gillott, C.; Hou, X.; Coutu, C.; Hegedus, D.D. A chitin deacetylase and putative insect intestinal lipases are components of the Mamestra configurata (Lepidoptera: Noctuidae) peritrophic matrix. Insect Mol. Biol. 2008, 17, 573–585. [Google Scholar] [CrossRef]
- Zhong, X.W.; Wang, X.H.; Tan, X.; Xia, Q.Y.; Xiang, Z.H.; Zhao, P. Identification and molecular characterization of a chitin deacetylase from Bombyx mori peritrophic membrane. Int. J. Mol. Sci. 2014, 15, 1946–1961. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.M.; Cao, A.T.; Hines, E.R.; East, P.D.; Gordon, K.H.J. Proteomic analysis of the peritrophic matrix from the gut of the caterpillar, Helicoverpa armigera. Insect Biochem. Mol. Biol. 2008, 38, 950–958. [Google Scholar] [CrossRef]
- Liu, L.; Qu, M.B.; Liu, T.; Chen, Q.; Guo, X.M.; Yang, J.; Yang, Q. Biochemical characterization of three midgut chitin deacetylases of the Lepidopteran insect Bombyx mori. J. Insect Physiol. 2019, 113, 42–48. [Google Scholar] [CrossRef]
- Gilbert, L.I.; Rybczynski, R.; Warren, J.T. Control and Biochemical Nature of the Ecdysteroidogenic Pathway. Annu. Rev. Entomol. 2002, 47, 883–916. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.Z.; Zhang, K.; Li, C.Y.; Hu, X.; Kausar, S.; Gu, H.Y.; Yang, L.Q.; Cui, H.J. A hemocyte-specific cathepsin L-like cysteine protease is involved in response to 20-hydroxyecdysone and microbial pathogens stimulation in silkworm, Bombyx mori. Mol. Immunol. 2021, 131, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.L.; Wang, J.Y.; Wang, Y.Z.; Yang, J.J.; Wei, B.F.; Li, M.M.; Wang, B.; Li, X.Z.; Cao, Y.; Tian, L.; et al. Transcriptome Analysis Reveals Potential Function of Long Non-coding RNAs in 20-hydroxyecdysone Regulated Autophagy in Bombyx Mori. BMC Genomics 2021, 22, 374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Su, J.J.; Chen, S.Y.; Yu, S.; Tan, J.; Xu, M.; Liang, H.H.; Zhao, Y.Z.; Chao, H.J.; Yang, L.Q.; et al. Molecular cloning, characterization and expression analysis of cathepsin O in silkworm Bombyx mori related to bacterial response. Mol. Immunol. 2015, 66, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Zhang, D.W.; Tang, B.; Chen, J.; Chen, J.; Lu, L.; Zhang, W.Q. Identification of 20-hydroxyecdysone late-response genes in the chitin biosynthesis pathway. PLoS ONE 2010, 5, e14058. [Google Scholar] [CrossRef]
- Cai, R.; Tao, G.; Zhao, P.; Xia, Q.Y.; He, H.W.; Wang, Y.J. POU-M2 promotes juvenile hormone biosynthesis by directly activating the transcription of juvenile hormone synthetic enzyme genes in Bombyx mori. Open Biol. 2022, 12, 220031. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.W.; Yao, X.G.; Yang, Q.F.; Wang, X.G.; Zhang, Y.M.; Wang, Y.M.; Shen, L.T. Fitness Costs of Chlorantraniliprole Resistance Related to the SeNPF Overexpression in the Spodoptera exigua (Lepidoptera: Noctuidae). Int. J. Mol. Sci. 2021, 22, 5027. [Google Scholar] [CrossRef]
- Yang, W.J.; Xu, K.K.; Yan, Y.; Li, C.; Jin, D.C. Role of Chitin Deacetylase 1 in the molting and metamorphosis of the Cigarette Beetle Lasioderma serricorne. Int. J. Mol. Sci. 2020, 21, 2449. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, J.; Yao, L.; Li, S.Y.; Chen, Y.R.; Yang, H.J.; Fan, D. Chitin deacetylase: A potential target for Mythimna separata (Walker) control. Arch. Insect. Biochem. Physiol. 2020, 104, e21666. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Xue, B.; Li, Y.Y.; Li, F.C.; Ni, M.; Shen, W.D.; Gu, Z.Y.; Li, B. Construction of silkworm midgut cDNA library for screen and sequence analysis of peritrophic membrane protein genes. Arch. Insect. Biochem. Physiol. 2016, 91, 3–16. [Google Scholar] [CrossRef]
- Liu, L.; Qu, M.; Yang, J.; Yang, Q. The physiological differentiation along the midgut of Bombyx mori–inspirations from proteomics and gene expression patterns of the secreted proteins in the ectoperitrophic space. Insect Mol. Biol. 2018, 27, 247–259. [Google Scholar] [CrossRef]
- Harper, M.S.; Hopkins, T.L. Peritrophic membrane structure and secretion in European corn borer larvae (Ostrinia nubilalis). Tissue Cell 1997, 29, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Toprak, U.; Hegedus, D.D.; Baldwin, D.; Coutu, C.; Erlandson, M. Spatial and temporal synthesis of Mamestra configurata peritrophic matrix through a larval stadium. Insect Biochem. Mol. Biol. 2014, 54, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Jayaram, S.J.; Senti, K.; Tsarouhas, V.; Jin, H.; Samakovlis, C. Septate-junction-dependent luminal deposition of chitin deacetylases restricts tube elongation in the Drosophila trachea. Curr. Biol. 2006, 16, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.R.; Liu, W.M.; Li, D.Q.; Zhao, X.M.; Ding, G.W.; Zhang, M.; Ma, E.B.; Zhu, K.Y.; Li, S.; Moussian, B.; et al. Helicoidal Organization of Chitin in the Cuticle of the Migratory Locust Requires the Function of the Chitin Deacetylase2 Enzyme (LmCDA2). J. Biol. Chem. 2016, 291, 24352–24363. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska, A.K.; Caccia, S.; Gordon, K.H.; Ferre, J.; Herrero, S. Downregulation of a chitin deacetylase-like protein in response to baculovirus infection and its application for improving baculovirus infectivity. J. Virol. 2010, 84, 2547–2555. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Zhao, D.; Wu, H.; Ji, Y.J.; Liu, Z.R.; Guo, X.C.; Guo, W.; Bi, Y. Bt GS57 interaction with gut microbiota accelerates Spodoptera exigua mortality. Front. Microbiol. 2022, 13, 835227. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cDNA | GenBank Accession | cDNA Length (bp) | ORF (aa) | M.V (kDa) | pI | Chromosome Position |
|---|---|---|---|---|---|---|
| SeCDA6 | AYX80514.1 | 1164 | 387 | 44 | 4.82 | Chr24 |
| SeCDA7 | AYX80515.1 | 1137 | 378 | 43 | 5.29 | Chr24 |
| SeCDA8 | OL689577 | 1158 | 385 | 44 | 5.19 | Chr24 |
| SeCDA9 | AMY98414.1 | 1152 | 383 | 43 | 4.98 | Chr24 |
| Primers | Sequences | Purpose |
|---|---|---|
| SeActinF | TTCCCATCCATCGTAGGT | qRT-PCR |
| SeActinR | GGATACCTCTCTTGCTCTGG | |
| SeCDA6F-Q | GAGTGCTTTGCTATTAATTCGC | |
| SeCDA6R-Q | GGTAGATAACGCCCTCAAGG | |
| SeCDA7F-Q | CGAAGAGTTGTGTAAGTTGCCT | |
| SeCDA7R-Q | CGAGTGTAAGGCGATTTCA | |
| SeCDA8F-Q | ATGTTACTCCTTGAAGCCGTTC | |
| SeCDA8R-Q | GGAACTGAAATTGCACTGGTG | |
| SeCDA9F-Q | CCTGGACTCTGGCCTTACACTC | |
| SeCDA9R-Q | GGAATGAAGAAGCAAGCATCC | |
| T7-dsSeCDA6F | TAATACGACTCACTATAGGACACGAGTGCTTTGCTATTA | dsRNA synthesis |
| dsSeCDA6R | CCGTCGTAGCTGGAGATG | |
| dsSeCDA6F | ACACGAGTGCTTTGCTATTA | |
| T7-dsSeCDA6R | TAATACGACTCACTATAGGCCGTCGTAGCTGGAGATG | |
| T7-dsSeCDA7F | TAATACGACTCACTATAGGCGAAGAGTTGTGTAAGTTGCCT | |
| dsSeCDA7R | TGTTGCCAGCCAGTTGTAG | |
| dsSeCDA7F | CGAAGAGTTGTGTAAGTTGCCT | |
| T7-dsSeCDA7R | TAATACGACTCACTATAGGTGTTGCCAGCCAGTTGTAG | |
| T7-dsSeCDA8F | TAATACGACTCACTATAGGGCTACTACGCGCCTGACTTC | |
| dsSeCDA8R | GGAACTGAAATTGCACTGGTG | |
| dsSeCDA8F | GCTACTACGCGCCTGACTTC | |
| T7-dsSeCDA8R | TAATACGACTCACTATAGGGGAACTGAAATTGCACTGGTG | |
| T7-dsSeCDA9F | TAATACGACTCACTATAGGTGGCTGATTATGGTTTGGAG | |
| dsSeCDA9R | AGGATTCTTCACCCAGTCAA | |
| dsSeCDA9F | TGGCTGATTATGGTTTGGAG | |
| T7-dsSeCDA9R | TAATACGACTCACTATAGGAGGATTCTTCACCCAGTCAA | |
| T7-dsSeCDAVF | TAATACGACTCACTATAGGGTGAACGAGCTCTACAACCG | |
| dsSeCDAVR | GTTTAGGATGAACTGGAACC | |
| dsSeCDAVF | GTGAACGAGCTCTACAACCG | |
| T7-dsSeCDAVR | TAATACGACTCACTATAGGGTTTAGGATGAACTGGAACC | |
| T7-dsGFPF | TAATACGACTCACTATAGGCCACAAGTTCAGCGTGTCCG | |
| dsGFPR | AGTTCACCTTGATGCCGTTCT | |
| dsGFPF | CCACAAGTTCAGCGTGTCCG | |
| T7-dsGFPR | TAATACGACTCACTATAGGAGTTCACCTTGATGCCGTTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Zhao, D.; Guo, X.-C.; Liu, Z.-R.; Li, R.-J.; Lu, X.-J.; Guo, W. Group V Chitin Deacetylases Influence the Structure and Composition of the Midgut of Beet Armyworm, Spodoptera exigua. Int. J. Mol. Sci. 2023, 24, 3076. https://doi.org/10.3390/ijms24043076
Wu H, Zhao D, Guo X-C, Liu Z-R, Li R-J, Lu X-J, Guo W. Group V Chitin Deacetylases Influence the Structure and Composition of the Midgut of Beet Armyworm, Spodoptera exigua. International Journal of Molecular Sciences. 2023; 24(4):3076. https://doi.org/10.3390/ijms24043076
Chicago/Turabian StyleWu, Han, Dan Zhao, Xiao-Chang Guo, Zhao-Rui Liu, Rui-Jun Li, Xiu-Jun Lu, and Wei Guo. 2023. "Group V Chitin Deacetylases Influence the Structure and Composition of the Midgut of Beet Armyworm, Spodoptera exigua" International Journal of Molecular Sciences 24, no. 4: 3076. https://doi.org/10.3390/ijms24043076
APA StyleWu, H., Zhao, D., Guo, X.-C., Liu, Z.-R., Li, R.-J., Lu, X.-J., & Guo, W. (2023). Group V Chitin Deacetylases Influence the Structure and Composition of the Midgut of Beet Armyworm, Spodoptera exigua. International Journal of Molecular Sciences, 24(4), 3076. https://doi.org/10.3390/ijms24043076