

DNA Methylation as a Diagnostic, Prognostic, and Predictive Biomarker in Head and Neck Cancer

 , , ,
, , ,
Abstract
1. Introduction
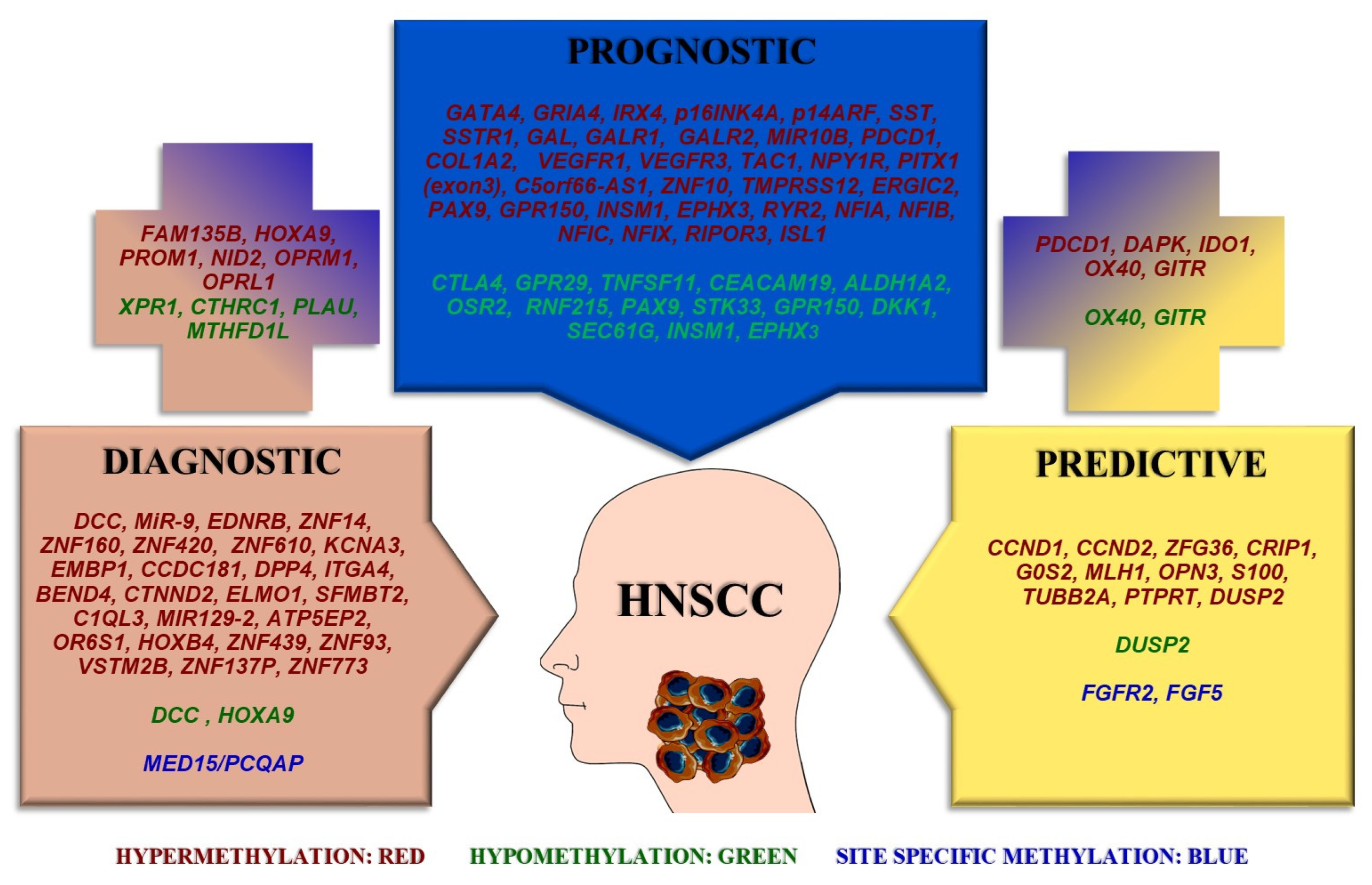
2. DNA Methylation Biomarkers with Diagnostic Potential
3. DNA Methylation Biomarkers with Prognostic Potential
4. DNA Methylation Biomarkers with Predictive Potential
4.1. DNA Methylation Evaluated Pre-Clinically (Cell Lines)
4.2. DNA Methylation Evaluated in Tissue Samples
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arantes, L.M.R.B.; De Carvalho, A.C.; Melendez, M.E.; Lopes Carvalho, A. Serum, Plasma and Saliva Biomarkers for Head and Neck Cancer. Expert Rev. Mol. Diagn. 2018, 18, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Galvis, M.; Loveless, R.; Kowalski, L.P.; Teng, Y. Impacts of Environmental Factors on Head and Neck Cancer Pathogenesis and Progression. Cells 2021, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The Molecular Landscape of Head and Neck Cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Mesia, R.; Iglesias, L.; Lambea, J.; Martínez-Trufero, J.; Soria, A.; Taberna, M.; Trigo, J.; Chaves, M.; García-Castaño, A.; Cruz, J. SEOM Clinical Guidelines for the Treatment of Head and Neck Cancer 2020. Clin. Transl. Oncol. 2021, 23, 913–921. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Network Comprehensive Genomic Characterization of Head and Neck Squamous Cell Carcinomas. Nature 2015, 517, 576–582. [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Amenábar, J.M.; Da Silva, B.M.; Punyadeera, C. Salivary Protein Biomarkers for Head and Neck Cancer. Expert Rev. Mol. Diagn. 2020, 20, 305–313. [Google Scholar] [CrossRef]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef]
- Kwok Fung Lo, A.; Wai Lo, K.; Tsao, S.W.; Wong, H.L.; Ying Hui, J.W.; To, K.F.; Hayward, S.D.; Chui, Y.L.; Lau, Y.L.; Takada, K.; et al. Epstein-Barr Virus Infection Alters Cellular Signal Cascades in Human Nasopharyngeal Epithelial Cells. Neoplasia 2006, 8, 173–180. [Google Scholar] [CrossRef]
- Hennessey, P.T.; Westra, W.H.; Califano, J.A. Human Papillomavirus and Head and Neck Squamous Cell Carcinoma: Recent Evidence and Clinical Implications. J. Dent. Res. 2009, 88, 300–306. [Google Scholar] [CrossRef]
- Schmidt, H.; Kulasinghe, A.; Perry, C.; Nelson, C.; Punyadeera, C. A Liquid Biopsy for Head and Neck Cancers. Expert Rev. Mol. Diagn. 2016, 16, 165–172. [Google Scholar] [CrossRef]
- Hattori, N.; Ushijima, T. Epigenetic Impact of Infection on Carcinogenesis: Mechanisms and Applications. Genome Med. 2016, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-H.; Hsia, S.-M.; Shih, Y.-H.; Shieh, T.-M. Association of Smoking, Alcohol Use, and Betel Quid Chewing with Epigenetic Aberrations in Cancers. IJMS 2017, 18, 1210. [Google Scholar] [CrossRef] [PubMed]
- Camuzi, D.; Simão, T.D.A.; Dias, F.; Ribeiro Pinto, L.F.; Soares-Lima, S.C. Head and Neck Cancers Are Not Alike When Tarred with the Same Brush: An Epigenetic Perspective from the Cancerization Field to Prognosis. Cancers 2021, 13, 5630. [Google Scholar] [CrossRef] [PubMed]
- Richards, K.L.; Zhang, B.; Baggerly, K.A.; Colella, S.; Lang, J.C.; Schuller, D.E.; Krahe, R. Genome-Wide Hypomethylation in Head and Neck Cancer is More Pronounced in HPV-Negative Tumors and is Associated with Genomic Instability. PLoS ONE 2009, 4, e4941. [Google Scholar] [CrossRef]
- Yong, W.-S.; Hsu, F.-M.; Chen, P.-Y. Profiling Genome-Wide DNA Methylation. Epigenet. Chromatin 2016, 9, 26. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA Methylation: Islands, Start Sites, Gene Bodies and Beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Guerrero-Preston, R.; Michailidi, C.; Marchionni, L.; Pickering, C.R.; Frederick, M.J.; Myers, J.N.; Yegnasubramanian, S.; Hadar, T.; Noordhuis, M.G.; Zizkova, V.; et al. Key Tumor Suppressor Genes Inactivated by “Greater Promoter” Methylation and Somatic Mutations in Head and Neck Cancer. Epigenetics 2014, 9, 1031–1046. [Google Scholar] [CrossRef]
- Aronson, J.K.; Ferner, R.E. Biomarkers—A General Review. Curr. Protoc. Pharmacol. 2017, 76, 9–23. [Google Scholar] [CrossRef]
- Califf, R.M. Biomarker Definitions and Their Applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef]
- Minor, J.; Wang, X.; Zhang, F.; Song, J.; Jimeno, A.; Wang, X.-J.; Lu, X.; Gross, N.; Kulesz-Martin, M.; Wang, D.; et al. Methylation of MicroRNA-9 Is a Specific and Sensitive Biomarker for Oral and Oropharyngeal Squamous Cell Carcinomas. Oral Oncol. 2012, 48, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Schussel, J.; Zhou, X.C.; Zhang, Z.; Pattani, K.; Bermudez, F.; Jean-Charles, G.; McCaffrey, T.; Padhya, T.; Phelan, J.; Spivakovsky, S.; et al. EDNRB and DCC Salivary Rinse Hypermethylation Has a Similar Performance as Expert Clinical Examination in Discrimination of Oral Cancer/Dysplasia versus Benign Lesions. Clin. Cancer Res. 2013, 19, 3268–3275. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, D.A.; Wan, Y.; Coman, W.B.; Pandit, P.; Cooper-White, J.J.; Herman, J.G.; Punyadeera, C. DNA Methylation at the Novel CpG Sites in the Promoter of MED15/PCQAP Gene as a Biomarker for Head and Neck Cancers. Biomark. Insights 2014, 9, BMI.S16199. [Google Scholar] [CrossRef]
- Beck, T.N.; Golemis, E.A. Genomic Insights into Head and Neck Cancer. Cancers Head Neck 2016, 1, 1. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, M.; Ni, S.; Li, Q.; Ye, D.; Li, J.; Shen, Z.; Deng, H. DNA Methylation Biomarkers for Head and Neck Squamous Cell Carcinoma. Epigenetics 2018, 13, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, J.; Li, Q.; Liu, H.; Ye, D.; Wu, Z.; Shen, Z.; Deng, H. The Clinical Significance of HOXA9 Promoter Hypermethylation in Head and Neck Squamous Cell Carcinoma. J. Clin. Lab. Anal. 2019, 33, e22873. [Google Scholar] [CrossRef]
- Ren, S.; Gaykalova, D.; Wang, J.; Guo, T.; Danilova, L.; Favorov, A.; Fertig, E.; Bishop, J.; Khan, Z.; Flam, E.; et al. Discovery and Development of Differentially Methylated Regions in Human Papillomavirus-Related Oropharyngeal Squamous Cell Carcinoma. Int. J. Cancer 2018, 143, 2425–2436. [Google Scholar] [CrossRef]
- Hu, Z.; Liu, H.; Zhang, X.; Hong, B.; Wu, Z.; Li, Q.; Zhou, C. Promoter Hypermethylation of CD133/PROM1 Is an Independent Poor Prognosis Factor for Head and Neck Squamous Cell Carcinoma. Medicine 2020, 99, e19491. [Google Scholar] [CrossRef]
- Sial, N.; Ahmad, M.; Hussain, M.S.; Iqbal, M.J.; Hameed, Y.; Khan, M.; Abbas, M.; Asif, R.; Rehman, J.U.; Atif, M.; et al. CTHRC1 Expression Is a Novel Shared Diagnostic and Prognostic Biomarker of Survival in Six Different Human Cancer Subtypes. Sci. Rep. 2021, 11, 19873. [Google Scholar] [CrossRef]
- Huo, Z.; Li, X.; Zhou, J.; Fan, Y.; Wang, Z.; Zhang, Z. Hypomethylation and Downregulation of MiR-23b-3p Are Associated with Upregulated PLAU: A Diagnostic and Prognostic Biomarker in Head and Neck Squamous Cell Carcinoma. Cancer Cell Int. 2021, 21, 564. [Google Scholar] [CrossRef]
- Sial, N.; Rehman, J.U.; Saeed, S.; Ahmad, M.; Hameed, Y.; Atif, M.; Rehman, A.; Asif, R.; Ahmed, H.; Hussain, M.S.; et al. Integrative Analysis Reveals Methylenetetrahydrofolate Dehydrogenase 1-like as an Independent Shared Diagnostic and Prognostic Biomarker in Five Different Human Cancers. Biosci. Rep. 2022, 42, BSR20211783. [Google Scholar] [CrossRef]
- Ishikawa, R.; Imai, A.; Mima, M.; Yamada, S.; Takeuchi, K.; Mochizuki, D.; Shinmura, D.; Kita, J.; Nakagawa, T.; Kurokawa, T.; et al. Novel Prognostic Value and Potential Utility of Opioid Receptor Gene Methylation in Liquid Biopsy for Oral Cavity Cancer. Curr. Probl. Cancer 2022, 46, 100834. [Google Scholar] [CrossRef]
- Wang, L. Bioinformatics Analyses Proposed Xenotropic and Polytropic Retrovirus Receptor 1 as a Potential Diagnostic and Prognostic Biomarker and Immunotherapeutic Target in Head and Neck Squamous Cell Carcinoma. Auris Nasus Larynx 2022, 50, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Al-Kaabi, A.; van Bockel, L.W.; Pothen, A.J.; Willems, S.M. P16 INK4A and P14 ARF Gene Promoter Hypermethylation as Prognostic Biomarker in Oral and Oropharyngeal Squamous Cell Carcinoma: A Review. Dis. Markers 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Misawa, K.; Misawa, Y.; Kondo, H.; Mochizuki, D.; Imai, A.; Fukushima, H.; Uehara, T.; Kanazawa, T.; Mineta, H. Aberrant Methylation Inactivates Somatostatin and Somatostatin Receptor Type 1 in Head and Neck Squamous Cell Carcinoma. PLoS ONE 2015, 10, e0118588. [Google Scholar] [CrossRef] [PubMed]
- Misawa, K.; Mochizuki, D.; Endo, S.; Mima, M.; Misawa, Y.; Imai, A.; Shinmura, K.; Kanazawa, T.; Carey, T.E.; Mineta, H. Site-Specific Methylation Patterns of the GAL and GALR1/2 Genes in Head and Neck Cancer: Potential Utility as Biomarkers for Prognosis. Mol. Carcinog. 2017, 56, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.M.; Dhas, K.; Nair, J.; Palve, V.; Bagwan, J.; Siddappa, G.; Suresh, A.; Kekatpure, V.D.; Kuriakose, M.A.; Panda, B. A Minimal DNA Methylation Signature in Oral Tongue Squamous Cell Carcinoma Links Altered Methylation with Tumor Attributes. Mol. Cancer Res. 2016, 14, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Goltz, D.; Gevensleben, H.; Dietrich, J.; Schroeck, F.; de Vos, L.; Droege, F.; Kristiansen, G.; Schroeck, A.; Landsberg, J.; Bootz, F.; et al. PDCD1 (PD-1) Promoter Methylation Predicts Outcome in Head and Neck Squamous Cell Carcinoma Patients. Oncotarget 2017, 8, 41011–41020. [Google Scholar] [CrossRef]
- Misawa, K.; Mochizuki, D.; Imai, A.; Mima, M.; Misawa, Y.; Mineta, H. Analysis of Site-Specific Methylation of Tumor-Related Genes in Head and Neck Cancer: Potential Utility as Biomarkers for Prognosis. Cancers 2018, 10, 27. [Google Scholar] [CrossRef]
- Sailer, V.; Charpentier, A.; Dietrich, J.; Vogt, T.J.; Franzen, A.; Bootz, F.; Dietrich, D.; Schroeck, A. Intragenic DNA Methylation of PITX1 and the Adjacent Long Non-Coding RNA C5orf66-AS1 Are Prognostic Biomarkers in Patients with Head and Neck Squamous Cell Carcinomas. PLoS ONE 2018, 13, e0192742. [Google Scholar] [CrossRef]
- Ma, J.; Li, R.; Wang, J. Characterization of a Prognostic Four-gene Methylation Signature Associated with Radiotherapy for Head and Neck Squamous Cell Carcinoma. Mol. Med. Rep. 2019, 20, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Sailer, V.; Sailer, U.; Bawden, E.G.; Zarbl, R.; Wiek, C.; Vogt, T.J.; Dietrich, J.; Loick, S.; Grünwald, I.; Toma, M.; et al. DNA Methylation of Indoleamine 2,3-Dioxygenase 1 (IDO1) in Head and Neck Squamous Cell Carcinomas Correlates with IDO1 Expression, HPV Status, Patients’ Survival, Immune Cell Infiltrates, Mutational Load, and Interferon γ Signature. EBioMedicine 2019, 48, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hei, N.; Zhao, J.; Peng, S.; Yang, K.; Chen, H.; Cui, Z.; Jin, L.; Sun, R.; Guo, J. A Two-CpG-Based Prognostic Signature for Oral Squamous Cell Carcinoma Overall Survival. J. Cell Biochem. 2019, 120, 9082–9090. [Google Scholar] [CrossRef]
- Bai, G.; Song, J.; Yuan, Y.; Chen, Z.; Tian, Y.; Yin, X.; Niu, Y.; Liu, J. Systematic Analysis of Differentially Methylated Expressed Genes and Site-Specific Methylation as Potential Prognostic Markers in Head and Neck Cancer. J. Cell Physiol. 2019, 234, 22687–22702. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, K.; Molfenter, B.; Laureano, N.K.; Tawk, B.; Bieg, M.; Hostench, X.P.; Weichenhan, D.; Ullrich, N.D.; Shang, V.; Richter, D.; et al. Somatic Mutations and Promotor Methylation of the Ryanodine Receptor 2 Is a Common Event in the Pathogenesis of Head and Neck Cancer. Int. J. Cancer 2019, 145, 3299–3310. [Google Scholar] [CrossRef]
- Li, Y.; Sun, C.; Tan, Y.; Li, L.; Zhang, H.; Liang, Y.; Zeng, J.; Zou, H. Transcription Levels and Prognostic Significance of the NFI Family Members in Human Cancers. PeerJ 2020, 8, e8816. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Rodrìguez, R.A.; Mullor, M.D.M.R.; Tan, Z.; Gui, Y.; Hu, J.; Zhu, T.; Huang, X.; Zhu, Y.; Xu, J. Analyzing the Prognostic Value of DKK1 Expression in Human Cancers Based on Bioinformatics. Ann. Transl. Med. 2020, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Chen, Y.; Gong, X.; Guo, Q.; Lin, C.; Luo, Q.; Tu, Z.; Pan, J.; Li, J. SEC61G Overexpression and DNA Amplification Correlates with Prognosis and Immune Cell Infiltration in Head and Neck Squamous Cell Carcinoma. Cancer Med. 2021, 10, 7847–7862. [Google Scholar] [CrossRef]
- Misawa, K.; Imai, A.; Matsui, H.; Kanai, A.; Misawa, Y.; Mochizuki, D.; Mima, M.; Yamada, S.; Kurokawa, T.; Nakagawa, T.; et al. Identification of Novel Methylation Markers in HPV-Associated Oropharyngeal Cancer: Genome-Wide Discovery, Tissue Verification and Validation Testing in CtDNA. Oncogene 2020, 39, 4741–4755. [Google Scholar] [CrossRef]
- Loick, S.M.; Fröhlich, A.; Gabrielpillai, J.; Franzen, A.; Vogt, T.J.; Dietrich, J.; Wiek, C.; Scheckenbach, K.; Strieth, S.; Landsberg, J.; et al. DNA Methylation and MRNA Expression of OX40 (TNFRSF4) and GITR (TNFRSF18, AITR) in Head and Neck Squamous Cell Carcinoma Correlates With HPV Status, Mutational Load, an Interferon-γ Signature, Signatures of Immune Infiltrates, and Survival. J. Immunother. 2022, 45, 194–206. [Google Scholar] [CrossRef]
- Zhang, K.; Da, J.; Liu, X.; Liu, X.; Wang, J.; Jin, H.; Li, Y.; Zhang, B. Downregulated Expression of RIPOR3 Correlated with Immune Infiltrates Predicts Poor Prognosis in Oral Tongue Cancer. Med. Sci. Monit. 2022, 28, e935055. [Google Scholar] [CrossRef]
- Wan, Z.; Xiong, H.; Tan, X.; Su, T.; Xia, K.; Wang, D. Integrative Multi-Omics Analysis Reveals Candidate Biomarkers for Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 794146. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.P.; Esteves, L.; Caramelo, F.; Carreira, I.M.; Melo, J.B. Integrated Multi-Omics Signature Predicts Survival in Head and Neck Cancer. Cells 2022, 11, 2536. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Adnan, A.; Rakha, A.; Nazir, S.; Tian, M.; Zhang, S.; Pang, H. Systematic Sequence Analysis of the FUT3 Gene Identifies 11 Novel Alleles in the Sindhi and Punjabi Populations from Pakistan. Sci. Rep. 2020, 10, 5543. [Google Scholar] [CrossRef] [PubMed]
- Mandell, M.A.; Jain, A.; Arko-Mensah, J.; Chauhan, S.; Kimura, T.; Dinkins, C.; Silvestri, G.; Münch, J.; Kirchhoff, F.; Simonsen, A.; et al. TRIM Proteins Regulate Autophagy and Can Target Autophagic Substrates by Direct Recognition. Dev. Cell. 2014, 30, 394–409. [Google Scholar] [CrossRef] [PubMed]
- Perot, B.P.; Ménager, M.M. Tetraspanin 7 and Its Closest Paralog Tetraspanin 6: Membrane Organizers with Key Functions in Brain Development, Viral Infection, Innate Immunity, Diabetes and Cancer. Med. Microbiol. Immunol. 2020, 209, 427–436. [Google Scholar] [CrossRef]
- Clark, A.M.; Reynolds, S.H.; Anderson, M.; Wiest, J.S. Mutational Activation of the MAP3K8 Protooncogene in Lung Cancer. Genes Chromosomes Cancer 2004, 41, 99–108. [Google Scholar] [CrossRef]
- Milosevic, N.; Kühnemuth, B.; Mühlberg, L.; Ripka, S.; Griesmann, H.; Lölkes, C.; Buchholz, M.; Aust, D.; Pilarsky, C.; Krug, S.; et al. Synthetic Lethality Screen Identifies RPS6KA2 as Modifier of Epidermal Growth Factor Receptor Activity in Pancreatic Cancer. Neoplasia 2013, 15, 1354–1362. [Google Scholar] [CrossRef]
- Patak, J.; Hess, J.L.; Zhang-James, Y.; Glatt, S.J.; Faraone, S.V. SLC9A9 Co-Expression Modules in Autism-Associated Brain Regions. Autism Res. 2017, 10, 414–429. [Google Scholar] [CrossRef]
- Luoma, L.M.; Berry, F.B. Molecular Analysis of NPAS3 Functional Domains and Variants. BMC Mol. Biol. 2018, 19, 14. [Google Scholar] [CrossRef]
- Neighbors, A.; Moss, T.; Holloway, L.; Yu, S.-H.; Annese, F.; Skinner, S.; Saneto, R.; Steet, R. Functional Analysis of a Novel Mutation in the TIMM8A Gene That Causes Deafness-Dystonia-Optic Neuronopathy Syndrome. Mol. Genet. Genom. Med. 2020, 8, e1121. [Google Scholar] [CrossRef] [PubMed]
- Brickner, J.R.; Soll, J.M.; Lombardi, P.M.; Vågbø, C.B.; Mudge, M.C.; Oyeniran, C.; Rabe, R.; Jackson, J.; Sullender, M.E.; Blazosky, E.; et al. A Ubiquitin-Dependent Signalling Axis Specific for ALKBH-Mediated DNA Dealkylation Repair. Nature 2017, 551, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Naemura, M.; Murasaki, C.; Inoue, Y.; Okamoto, H. Transcriptional Regulation of the P16 Tumor Suppressor Gene. Anticancer Res 2015, 35, 4397–4401. [Google Scholar] [PubMed]
- Mabeta, P.; Steenkamp, V. The VEGF/VEGFR Axis Revisited: Implications for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 15585. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, R.M.; Page, D.J.; Ostergaard, P.; Keavney, B.D. The Physiological and Pathological Functions of VEGFR3 in Cardiac and Lymphatic Development and Related Diseases. Cardiovasc. Res. 2021, 117, 1877–1890. [Google Scholar] [CrossRef]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Novel Functions of Death-Associated Protein Kinases through Mitogen-Activated Protein Kinase-Related Signals. Int. J. Mol. Sci. 2018, 19, 3031. [Google Scholar] [CrossRef]
- Sergeeva, O.A.; Mazur, K.; Kernder, A.; Haas, H.L.; De Luca, R. Tachykinins Amplify the Action of Capsaicin on Central Histaminergic Neurons. Peptides 2022, 150, 170729. [Google Scholar] [CrossRef]
- Barreda-Gómez, G.; Manuel, I.; Rodríguez-Puertas, R. Neuroanatomical Characterization of the G Protein-Coupled Receptor Activity Evoked by Galanin-Related Ligands. J. Chem. Neuroanat. 2022, 128, 102226. [Google Scholar] [CrossRef]
- Nelson, T.S.; Taylor, B.K. Targeting Spinal Neuropeptide Y1 Receptor-Expressing Interneurons to Alleviate Chronic Pain and Itch. Prog. Neurobiol. 2021, 196, 101894. [Google Scholar] [CrossRef]
- Lukomska, A.; Dobrzanski, G.; Liguz-Lecznar, M.; Kossut, M. Somatostatin Receptors (SSTR1-5) on Inhibitory Interneurons in the Barrel Cortex. Brain Struct. Funct. 2020, 225, 387–401. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, C.; Gu, R.; Zhao, R.; Li, W.; Ma, Z.; Jia, Y.; Yu, F.; Luo, J.; Fu, Y.; et al. Nidogen-2 is a Novel Endogenous Ligand of LGR4 to Inhibit Vascular Calcification. Circ. Res. 2022, 131, 1037–1054. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, X.; Wang, L.; Ma, X.; Gong, Z.; Zhang, S.; Li, Y. Targeting the IDO1 Pathway in Cancer: From Bench to Bedside. J. Hematol. Oncol. 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Zisi, Z.; Adamopoulos, P.G.; Kontos, C.K.; Scorilas, A. Identification and Expression Analysis of Novel Splice Variants of the Human Carcinoembryonic Antigen-Related Cell Adhesion Molecule 19 (CEACAM19) Gene Using a High-Throughput Sequencing Approach. Genomics 2020, 112, 4268–4276. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.T.; Coulombe, P.A.; Kwan, R.; Omary, M.B. Types I and II Keratin Intermediate Filaments. Cold Spring Harb. Perspect. Biol. 2018, 10, a018275. [Google Scholar] [CrossRef]
- Henry, C.; Close, A.-F.; Buteau, J. A Critical Role for the Neural Zinc Factor ST18 in Pancreatic β-Cell Apoptosis. J. Biol. Chem. 2014, 289, 8413–8419. [Google Scholar] [CrossRef]
- Ogawa, T.; Liggett, T.E.; Melnikov, A.A.; Monitto, C.L.; Kusuke, D.; Shiga, K.; Kobayashi, T.; Horii, A.; Chatterjee, A.; Levenson, V.V.; et al. Methylation of Death-Associated Protein Kinase Is Associated with Cetuximab and Erlotinib Resistance. Cell Cycle 2012, 11, 1656–1663. [Google Scholar] [CrossRef]
- Chen, X.; Liu, L.; Mims, J.; Punska, E.C.; Williams, K.E.; Zhao, W.; Arcaro, K.F.; Tsang, A.W.; Zhou, X.; Furdui, C.M. Analysis of DNA Methylation and Gene Expression in Radiation-Resistant Head and Neck Tumors. Epigenetics 2015, 10, 545–561. [Google Scholar] [CrossRef]
- Lee, W.; Kim, S.; An, J.; Kim, T.-K.; Cha, H.; Chang, H.; Kim, S.; Kim, S.; Han, M. Tristetraprolin Regulates Phagocytosis through Interaction with CD47 in Head and Neck Cancer. Exp. Ther. Med. 2022, 24, 541. [Google Scholar] [CrossRef]
- Viet, C.T.; Dang, D.; Achdjian, S.; Ye, Y.; Katz, S.G.; Schmidt, B.L. Decitabine Rescues Cisplatin Resistance in Head and Neck Squamous Cell Carcinoma. PLoS ONE 2014, 9, e112880. [Google Scholar] [CrossRef]
- Peyser, N.D.; Freilino, M.; Wang, L.; Zeng, Y.; Li, H.; Johnson, D.E.; Grandis, J.R. Frequent Promoter Hypermethylation of PTPRT Increases STAT3 Activation and Sensitivity to STAT3 Inhibition in Head and Neck Cancer. Oncogene 2016, 35, 1163–1169. [Google Scholar] [CrossRef]
- Starzer, A.M.; Heller, G.; Tomasich, E.; Melchardt, T.; Feldmann, K.; Hatziioannou, T.; Traint, S.; Minichsdorfer, C.; Schwarz-Nemec, U.; Nackenhorst, M.; et al. DNA Methylation Profiles Differ in Responders versus Non-Responders to Anti-PD-1 Immune Checkpoint Inhibitors in Patients with Advanced and Metastatic Head and Neck Squamous Cell Carcinoma. J. Immunother. Cancer 2022, 10, e003420. [Google Scholar] [CrossRef] [PubMed]
- Lo Nigro, C.; Vivenza, D.; Denaro, N.; Lattanzio, L.; Fortunato, M.; Crook, T.; Merlano, M.C. DUSP2 Methylation is a Candidate Biomarker of Outcome in Head and Neck Cancer. Ann. Transl. Med. 2018, 6, 271. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Gabrielpillai, J.; Dietrich, J.; Zarbl, R.; Strieth, S.; Schröck, F.; Dietrich, D. Fibroblast Growth Factor (FGF), FGF Receptor (FGFR), and Cyclin D1 (CCND1) DNA Methylation in Head and Neck Squamous Cell Carcinomas is Associated with Transcriptional Activity, Gene Amplification, Human Papillomavirus (HPV) Status, and Sensitivity to Tyrosine Kinase Inhibitors. Clin. Epigenet. 2021, 13, 228. [Google Scholar] [CrossRef]
- Wagner, W. How to Translate DNA Methylation Biomarkers into Clinical Practice. Front. Cell Dev. Biol. 2022, 10, 854797. [Google Scholar] [CrossRef]
- Taryma-Leśniak, O.; Sokolowska, K.E.; Wojdacz, T.K. Current Status of Development of Methylation Biomarkers for in Vitro Diagnostic IVD Applications. Clin. Epigenet. 2020, 12, 100. [Google Scholar] [CrossRef]
- Beltrán-García, J.; Osca-Verdegal, R.; Mena-Mollá, S.; García-Giménez, J.L. Epigenetic IVD Tests for Personalized Precision Medicine in Cancer. Front. Genet. 2019, 10, 621. [Google Scholar] [CrossRef]
- García-Giménez, J.L.; Seco-Cervera, M.; Tollefsbol, T.O.; Romá-Mateo, C.; Peiró-Chova, L.; Lapunzina, P.; Pallardó, F.V. Epigenetic Biomarkers: Current Strategies and Future Challenges for Their Use in the Clinical Laboratory. Crit. Rev. Clin. Lab. Sci. 2017, 54, 529–550. [Google Scholar] [CrossRef]
- Nadauld, L.D.; McDonnell, C.H.; Beer, T.M.; Liu, M.C.; Klein, E.A.; Hudnut, A.; Whittington, R.A.; Taylor, B.; Oxnard, G.R.; Lipson, J.; et al. The PATHFINDER Study: Assessment of the Implementation of an Investigational Multi-Cancer Early Detection Test into Clinical Practice. Cancers 2021, 13, 3501. [Google Scholar] [CrossRef]


{kind=link}
| References | Cancer Type | Biomarker Name-Gene | Function | Sample Type | Biomarker Behavior | Application |
|---|---|---|---|---|---|---|
| [21] | Oral and oropharyngeal | MiR-9 | Non-coding RNA implicated in the modulation of gene expression, tumor suppression through inhibition of cellular pro-proliferation and modulation of PTEN. | Tissue | Promoter hypermethylation/sites 1 and 3.Low expression. | Diagnosis |
| [22] | Oral | EDNRB | It encodes the B-type endothelin receptor (G protein-coupled receptor) that triggers a phosphatidylinositol-calcium second messenger system. | Saliva | Promoter hypermethylation in 40% of malignant samples. | Screening, Diagnosis |
| [22] | Oral | DCC | Tumor suppressor, encodes a transmembrane protein with structural homology to NCAM, which is associated with the differentiation of epithelial and neuronal cells. | Saliva | Promoter hypermethylation in 40% of malignant samples. | Screening, Diagnosis |
| [23] | HNSCC | MED15/ PCQAP | Encodes a cofactor with pleiotropic activity, important for the formation of the RNA polymerase II complex, which is involved in the expression of all protein-coding genes, attenuation of at least one of the signaling pathways (TGFβ/Activin signaling). | Tissue, saliva | Specific methylation patterns in the promoter. | Screening, Diagnosis |
| [24] | HNSCC | ZNF14, ZNF160, ZNF420 | Members of the zinc finger gene family. | Tissue, saliva | Differential methylation pattern. | Diagnosis |
| [25] | HNSCC | FAM135B | Involved in the maintenance of the nucleolus, cell proliferation, cell differentiation, and apoptosis. | Tissue | Hypermethylation. Low expression. | Diagnosis |
| [25] | HNSCC | ZNF610 | Involved in the maintenance of the nucleolus, cell proliferation, cell differentiation, and apoptosis. | Tissue | Hypermethylation. Low expression. | Diagnosis |
| [25] | HNSCC | HOXA9 | Involved in the maintenance of the nucleolus, cell proliferation, cell differentiation, and apoptosis. | Tissue | Hypomethylation. High expression. | Diagnosis |
| [25] | HNSCC | DCC | Involved in the maintenance of the nucleolus, cell proliferation, cell differentiation, and apoptosis. | Tissue | Hypomethylation. High expression. | Diagnosis |
| [26] | HNSCC | HOXA9 | Encodes a transcription factor. HOX genes regulate and specify different cell types during embryonic growth, and have important functions in the modulation of the sensitive balance between cell proliferation and differentiation during cancer development. | Tissue | Promoter hypermethylation. | Screening, Diagnosis |
| [27] | HPV+ OPSCC | KCNA3, EMBP1, CCDC181, DPP4, ITGA4, BEND4, CTNND2, ELMO1, SFMBT2, C1QL3, MIR129-2, ATP5EP2, OR6S1, NID2, HOXB4, ZNF439, ZNF93, VSTM2B, ZNF137P and ZNF773 | ITGA4: on the cell surface, promotes migration and adhesion to the microenvironment in chronic lymphocytic leukemia. NID2: component of the basement membrane that stabilizes the extracellular matrix (ECM) network. Suppresses migration and blocks metastasis by downregulating the EGFR/AKT and integrin/FAK/PLCγ pathways. HOXB4: a hematopoietic transcription factor, downregulates the expression of Prdm16, which is a proto-oncogene necessary for self-renewal and preservation of mouse hematopoietic stem cells. SFMBT2: tumor suppressor, negatively regulates migration and invasion by targeting MMP-9 and MMP-26. MIR129-2: tumor suppressor, inhibits migration and invasion by directly inhibiting HMGB1. DPP4 and CTNND2: act both as tumor suppressors and as markers of tumor invasiveness, depending on the type of tumor. KCNA3: its knockdown markedly suppresses cell proliferation and increases apoptosis. EMBP1: related to ER-positive breast cancer and lower grade breast tumors. ZNF93: implicated in the DNA repair pathway following DNA damage by chemotherapy. Little is known about six candidates (ATP5EP2, OR6S1, ZNF439, VSTM2B, ZNF137P, ZNF773). ZNF439, ZNF137P, ZNF773: belong to the zinc finger protein group, members of which have previously been shown to have tumor-suppressive activity. | Tissue | 20 differentially DMRs → hypermethylation. | Screening |
| [28] | HNSCC | PROM1/CD133 | Encodes a pentaspan membrane glycoprotein, which is frequently found in adult stem cells. | Tissue | Promoter hypermethylation. Low expression. | Diagnosis |
| [29] | HNSCC | CTHRC1 | Encodes an extracellular matrix protein, modulating tumor microenvironment | Tissue | Promoter hypomethylation. High expression. | Diagnosis |
| [30] | HNSCC | PLAU | Encodes a protease implicated in apoptosis, epithelial-mesenchymal transition (EMT), and the Ras/MAPK pathway. | Tissue | Hypomethylation. High expression. | Diagnosis |
| [31] | HNSCC | MTHFD1L | Encodes a cytoplasmic enzyme participating in the formation of tetrahydrofolic acid (THF) within mitochondria. | Tissue | Promoter hypomethylation. High expression. | Diagnosis |
| [32] | HNSCC | Opioid receptor mu 1(OPRM1) | Encodes an opioid receptor (G-protein-coupled receptor). | Tissue, plasma | Hypermethylation. | Screening |
| [32] | HNSCC | Opioid-related nociceptin receptor 1(OPRL1) | Encodes an opioid receptor (G-protein-coupled receptor). | Tissue, plasma | Hypermethylation. | Screening |
| [33] | HNSCC | XPR1 | Encodes a cell surface receptor for certain types of murine leukemia viruses, which plays an essential role in maintaining intracellular phosphate homeostasis by mediating phosphate export from the cell. | Tissue | Promoter hypomethylation. High expression. | Diagnosis |
| References | Cancer Type | Biomarker Name-Gene | Function | Sample Type | Biomarker Behavior | Application |
|---|---|---|---|---|---|---|
| [24] | HNSCC | Methylation signature of the promoter of 5 genes, GATA4, GRIA4, IRX4 ALDH1A2, OSR2 | GATA4: encodes GATA binding protein 4, a member of the GATA family of zinc-finger transcription factors. GRIA4: encodes a glutamate receptor. IRX4: encodes Iroquois homeobox 4. ALDH1A2: encodes aldehyde dehydrogenase 1 family member A2, an enzyme that catalyzes the synthesis of retinoic acid from retinaldehyde. OSR2: odd-skipped related transcription factor 2. | Tissue | GATA4, GRIA4, IRX4: promoter hypermethylation. ALDH1A2 and OSR2: promoter hypomethylation. | Prognosis |
| [25] | HNSCC | FAM135B | Involved in the maintenance of the nucleolus, cell proliferation, cell differentiation, and apoptosis. | Tissue | hypermethylation → favorable prognostic feature | Prognosis |
| [26] | HNSCC | HOXA9 | Encodes a transcription factor involved in the maintenance of the nucleolus, in cell proliferation and differentiation, and in apoptosis. | Tissue | Promoter hypermethylation → progression and metastasis. | Prognosis |
| [28] | HNSCC | PROM1/ CD133 | Encodes a pentaspan membrane glycoprotein, often expressed on adult stem cells. | Tissue | Promoter hypermethylation-low expression → poor overall survival and relapse-free survival. | Prognosis |
| [29] | HNSCC | CTHRC1 | Encodes an extracellular matrix protein, involved in the modulating of the tumor microenvironment. | Tissue | Overexpression → decreased overall survival (OS). | Prognosis |
| [30] | HNSCC | PLAU | Encodes a protease, involved in apoptosis, epithelial–mesenchymal transition (EMT), and Ras/MAPK pathway. | Tissue | Hypomethylation → higher clinical stage, more aggressive metastatic disease. Hypermethylation → increased survival time. | Prognosis |
| [31] | HNSCC | MTHFD1L | Encodes a cytoplasmic enzyme, involved in the formation of tetrahydrofolate (THF) within mitochondria. | Tissue | High expression → decreased OS. | Prognosis |
| [32] | HNSCC | opioid receptor mu 1(OPRM1) | Encodes an opioid receptor (G-protein-coupled receptor). | Tissue, plasma | Hypermethylation → aggressive clinical behavior. | Prognosis |
| [32] | HNSCC | opioid-related nociceptin receptor 1(OPRL1) | Encodes an opioid receptor (G-protein-coupled receptor). | Tissue, plasma | Hypermethylation → aggressive clinical behavior. | Prognosis |
| [33] | HNSCC | XPR1 | Encodes a cell surface receptor for certain types of murine leukemia viruses that is involved in the maintenance of the intracellular phosphate homeostasis by mediating phosphate export from the cell. | Tissue | High expression → lower OS, DSS and PFI. | Prognosis |
| [34] | Oral and oropharyngeal squamous cell carcinoma (OSCC) | p16INK4A p14ARF | Tumor suppressors, encode cell cycle regulatory proteins. | Tissue | Promoter hypermethylation-low expression. P16 promoter hypermethylation → progression of premalignant lesions to OSCC, advanced disease, local recurrence, and lower DSS (disease-specific survival). | Prognosis |
| [35] | HNSCC | SST | Tumor suppressor, encodes a growth hormone release-inhibitory factor. | Tissue | Promoter hypermethylation-low expression. Hypermethylation → decreased disease-free survival, tumor recurrence after resection. | Prognosis |
| [35] | HNSCC | SSTR1 | Tumor suppressor, encodes a somatostatin receptor type 1. | Tissue | Promoter hypermethylation-low expression. Hypermethylation → decreased disease-free survival, tumor recurrence after resection. | Prognosis |
| [36] | HNSCC | GAL | Tumor suppressor, encodes a 30-amino acid peptide in humans, that targets the galanin system via receptors GALR1, GALR2, and GALR3. | Tissue | Promoter hypermethylation-low expression. Hypermethylation → reduced disease-free survival. | Prognosis |
| [36] | HNSCC | GALR1/2 | Tumor suppressors, encode galanin receptors (G protein-coupled receptors). | Tissue | Promoter hypermethylation-low expression. Hypermethylation → reduced disease-free survival. | Prognosis |
| [37] | OTSCC | MIR10B | Encodes a miRNA that targets oncogenes NR4A3 and BCL2L11. | Tissue | Hypermethylation-low expression → downregulation of NR4A3 and BCL2L11 → better disease-free survival. | Prognosis |
| [37] | OTSCC | FUT3, TRIM5, TSPAN7, MAP3K8, RPS6KA2, SLC9A9, NPAS3 TIMM8A, RNF113A genes | FUT3: regulates the expression of Lewis antigens in the human Lewis blood group system [54]. TRIM5: acts as a selective autophagy receptor that targets autophagic substrates via direct recognition [55]. TSPAN7: cell surface glycoprotein and may have a role in the control of neurite outgrowth, synaptic transmission, and viral-induced inflammation [56]. MAP3K8: proto-oncogene that participates in the MAP kinase and JNK signaling pathways [57]. RPS6KA2: controls cell growth and differentiation [58]. SLC9A9: autism-risk gene localized in the endosomal system that is involved in the endocytic regulation of autophagy and cell survival [59]. NPAS3: encodes a transcription factor that is expressed in the developing central nervous system [61]. TIMM8A: localizes to the intermembrane space in mitochondria where it functions in the import of nuclear-encoded proteins into the mitochondrial inner membrane [60]. RNF113A: involved in gene regulation and DNA repair [62]. | Tissue | Methylation signature, differential methylation among the various categories of parameters. | Prognosis |
| [38] | HNSCC | PDCD1 (PD-1) | Programmed cell death protein 1, immune inhibitory receptor (also known as CD279 or PDCD1), a member of the extended CD28/CTLA-4 family, it is stably expressed only on T cells exposed to a chronic antigen. | Tissue | Hypermethylation → shorter OS after surgical resection. | Prognosis |
| [39] | HPV negative hypopharyngeal cancer | COL1A2 | Encodes a fibrillar collagen found in most connective tissues and is the main component of the organic part of bones. | Tissue | Hypermethylation-low expression → reduced survival and a higher likelihood of relapse. | Prognosis |
| [39] | HPV negative laryngeal cancer | COL1A2 | Encodes a fibrillar collagen found in most connective tissues and is the main component of the organic part of bones. | Tissue | Hypermethylation-low expression → worse survival. | Prognosis |
| [39] | HPV negative laryngeal cancer | p16, VEGFR1, VEGFR3, DAPK, TAC1, GALR1, NPY1R, SSTR1 | p16: tumor suppressor gene that prevents oncogenic transformation through the induction of cellular senescence [63]. VEGFR1: stimulates angiogenesis and vascular permeability [64]. VEGFR3: regulates the development and maintenance of the lymphatic system [65]. DAPK: involved in multiple cellular signaling pathways that trigger autophagy, apoptosis, and cell survival [66]. TAC1: functions as a neurotransmitter interacting with both nerve receptors and smooth muscle cells [67]. GALR1: acts as a neuropeptide that modulating various physiological functions, including nociception, cognition, and neuroendocrine regulation [68]. NPY1R: acts as a neuropeptide that regulates several physiological processes including food intake, emotional regulation, and cardiovascular function [69]. SSTR1: neuropeptide that is involved in neurotransmission, secretion, and cell proliferation [70]. | Tissue | Hypermethylation-low expression → worse survival, higher risk of recurrence. | Prognosis |
| [40] | HNSCC, especially in HPV-negative | PITX1 3 exon | Encodes a transcription factor in embryogenesis that is involved in mouth and hindlimb formation and pituitary development, it is an upstream inducer of RASAL1 and therefore an important mediator of the Ras signaling pathway. | Tissue | Hypermethylation-low expression → increased risk for death. | Prognosis |
| [40] | HNSCC, especially in HPV-negative | C5orf66-AS1 lincRNA | Encodes a long intergenic non-coding RNA, involved in tumorigenesis, likely by acting as (post)-transcriptional regulator of PITX1. | Tissue | Hypermethylation-low expression → increased risk for death. Hypomethylation → better overall survival. | Prognosis |
| [41] | HNSCC | Methylation signature of ZNF10 TMPRSS12 ERGIC2 RNF215 | ZNF10: encodes a transcription repressor, involved in development, differentiation, and metabolism. TMPRSS12: encodes a member of the serine protease family that participates in immune response and blood coagulation and production, it is associated with human infertility. ERGIC2 protein: encodes an endoplasmic reticulum (ER) resident protein implicated in protein trafficking between ER and Golgi bodies. RNF125 gene: encodes a novel E3 ubiquitin-protein ligase that may be involved in the T-cell receptor signaling pathway, plays a role in tumorigenesis and metastasis, strengthens p53 degradation, and suppresses p53 function. | Tissue | Hypermethylation of ZNF10 as well as TMPRSS12 and ERGIC2 → unfavorable prognosis. Hypomethylation-high expression of RNF125 → poorer overall survival. | Prognosis |
| [42] | HNSCC | IDO1 | Encodes an indoleamine 2,3-dioxygenase (IDO)—an enzyme that limits the rate of conversion of the crucial amino acid tryptophan to kynurenine, it is strongly expressed in many types of tumors and has been shown to play a role in immunosuppression through increased tryptophan metabolism in the tumor microenvironment (TME). Increased expression of IDO1 can result in suppression of anti-tumor T-cells, differentiation of CD4+ T-cells into immunosuppressive regulatory T-cells (Tregs), and polarization of anti-gene cells into a tolerogenic phenotype. | Tissue | Hypermethylation of the lateral promoter → poor overall survival. | Prognosis, |
| [43] | Oral squamous cell carcinoma (OSCC) | Methylation signature of cg17892178 and cg17378966 in NID2 and IDO1 | NID2: encodes for a cell adhesion protein involved in maintaining the structure of the vascular extracellular matrix [71]. IDO1: rate-limiting enzyme involved in tryptophan catabolism; also acts as an immune checkpoint molecule on the surface of dendritic cells [72]. | Tissue | AUC = 0.81 indicated that the signature prognostic risk score based on two CpGs was a good prognostic factor of survival in patients with OSCC. | Prognosis |
| [44] | HNSCC | five DMG model PAX9, STK33, GPR150, INSM1, and EPHX3 | Encode genes that are involved in extracellular matrix, cell adhesion, and immune responses. | Tissue | Hypomethylation of PAX9, STK33, GPR150, INSM1, and EPHX3 → lower survival time and increased tumor-related mortality PAX9, GPR150, INSM1, and EPHX3 with hypermethylation and weak expression and hypomethylation and elevated expression → strongly associated with OS. | Prognosis |
| [45] | HNSCC | RYR2 | Encodes an important component of the intracellular Ca2+ release pathway, it is associated with the sarcoplasmic or endoplasmic reticulum of various cell types, particularly in cardiomyocytes. | Tissue, cell lines | Variable methylation profile. Gradual promoter hypermethylation-low expression → upcoming cancerous transformation of dysplastic lesions. | Prognosis |
| [46] | HNSCC | nuclear factor I (NFI): NFIA, NFIB, NFIC and NFIX | Transcription factors | Tissue | Promoter hypermethylation-low expression. Decreased levels of NFIA, NFIB, and NFIC → shorter overall survival. | Prognosis |
| [47] | HNSCC | DKK1 | Encodes a protein with inhibitory activity on the Wnt signaling pathway. | Tissue | Promoter hypomethylation-high expression. Overexpression → worse OS-DFS. | Prognosis |
| [49] | HPV+ OPC | CALML5 | Encodes a skin-specific calcium-binding protein, which is strongly related to the differentiation of keratinocytes. It is a regulator of the final differentiation of epidermal cells and CALML5 high-density conditions promote the translocation of YAP1 into the cytoplasm. | Tissue, plasma | Methylation → shorter DFS. | Prognosis |
| [49] | HPV+ OPC | DNAJC5G | Encodes a protein that inhibits the replication efficiencies of adenovirus, vaccinia virus, and HIV-1. | Tissue, plasma | Methylation → shorter DFS. | Prognosis |
| [49] | HPV+ OPC | LY6D | Encodes a membrane-bound protein with a glycosylphosphatidylinositol anchor. It has an important role in the adhesion of head and neck cancer cells to endothelial cells, and is detected of micrometastases in lymph nodes of HNSCC patients. | Tissue, plasma | Methylation → shorter DFS. | Prognosis |
| [48] | HNSCC | SEC61G | Encodes a transmembrane heterotrimeric channel protein that transports nascent polypeptides and proteins to the ER, mediating membrane protein degradation. | Tissue | Hypomethylation-high expression. Overexpression → worse OS. | Prognosis |
| [50] | HNSCC | OX40 (TNFRSF4) | Encodes a tumor necrosis factor receptor, which is expressed mainly on the surface of activated T cells and is stimulated by the OX40L ligand, which is found on antigen-presenting cells, activated T cells, lymphoid tissue inductor cells, some endothelial cells, and mast cells. Its up-regulation promotes differentiation, proliferation, and prolonged survival of T cell activation through inhibition of activation-induced cell death as well as stimulation of cytokine production. | Tissue | Promoter hypermethylation → worse overall survival. Higher expression levels → longer overall survival. | Prognosis, |
| [50] | HNSCC | GITR (TNFRSF18, AITR) | Encodes a tumor necrosis factor receptor, expressed at high levels by regulatory T cells (Tregs) and at lower levels by naïve, effector and memory T cells. The binding of the GITR ligand in conjunction with T cell receptor stimulation induces activation of the MAPK/ERK and NFkB pathway, resulting in immunoregulation of the immune system with T cell proliferation, production of proinflammatory cytokines, enhanced anti-cancer effector function and resistance of CD4+ and CD8+ T cells. GITR promotes anti-cancer immunity by enhancing effector T cell function on the one hand and inhibiting Treg proliferation. | Tissue | Promoter hypermethylation → worse overall survival. Higher expression levels → longer overall survival. | Prognosis, |
| [51] | Oral Tongue Cancer | RIPOR3 | Encodes a member of the extracellular region, regulates cellular processes in response to stimuli that is associated with immune cell infiltrations. | Tissue | Hypermethylation-low expression. Hypermethylation-low expression → poor prognosis. High expression → higher OS, PFS. | Prognosis |
| [52] | OSCC | CTLA4 (Cytotoxic T-lymphocyte associated protein 4) | Encodes a protein that is expressed in activated CD4+ T cells and constitutively expressed in CD4+Foxp3+ Treg cells. It is a potent immune inhibitor by decreasing the onset of T-cell activation mediated by the interactions between antigen presenting cells (APCs) and T cells. | Tissue | Hypomethylation. High expression → higher long-term survival. | Prognosis |
| [52] | OSCC | GPR29 | Encodes a G protein-coupled receptor. It is a chemokine receptor that is primarily expressed by T cells, immature dendritic cells, and B cells. Its unique partner CCL20 is known to be stably expressed by Th17 cells and secreted by intestinal epithelial cells in response to local enteropathogenic infection. The CCL20-CCR6 axis has a critical role in mucosal immunity, while the increased expression of CCR6 has previously been demonstrated in several cancers, with complicated anti-cancer or pre-cancer potential. | Tissue | Hypomethylation. High expression → higher long-term survival. | Prognosis |
| [52] | OSCC | TNFSF11 | Encodes a receptor activator of nuclear factor receptor kappa-B ligand (RANKL). It binds to the RANK receptor to modulate differentiation, activation, and survival of osteoclast cells. It has also been implicated in pathways such as the innate immune response, cell proliferation, and apoptosis. | Tissue | Hypomethylation. High expression → higher long-term survival. | Prognosis |
| [52] | OSCC | ISL1(ISL LIM homeobox 1) | Encodes the LIM-homeodomain transcription factor, which has been involved in many significant biological pathways, such as carcinogenesis, cell invasion, apoptosis, and cancer immunity. | Tissue | Hypermethylation. High expression → higher long-term survival. | Prognosis |
| [53] | HNSCC | Nine-gene multi-omics signature (methylation status of CEACAM19, KRT17, and ST18) | CEACAM19: immunoglobulin gene encoding for a variety of glycoproteins that are implicated in embryonic development, immunity, and may also serve as receptors for viruses and bacteria [73]. KRT17: involved in the formation and maintenance of several types of skin appendages, specifically in the shape and orientation of hair [74]. ST18: neural zinc finger transcription factor that negatively regulates cell proliferation [75]. | Tissue | Absence of high methylation of CEACAM19 → poor survival. Low expression of RPL29 and FCGR2C → poor survival. A methylation status greater than 0.625 in CEACAM19 → greater probability of survival. | Prognosis |
| References | Cancer Type | Biomarker Name-Gene | Function | Sample Type | Biomarker Behavior | Application |
|---|---|---|---|---|---|---|
| [38] | HNSCC | PDCD1 (PD-1) | The PD1/PD-L1 axis is implicated in the downregulation of immune responses in tumors, affecting T-cell responses in secondary lymphoid tissues, moving the balance from T-cell activation to antigen tolerance. | Tissue | Mpdcd1 → response to immunotherapies targeting the PD-1/PD-L1 axis. | Prediction |
| [42] | HNSCC | IDO1 | Encodes indoleamine 2,3-dioxygenase (IDO)—an enzyme that limits the rate of conversion of the crucial amino acid tryptophan to kynurenine, it is strongly expressed in many types of tumors and has been shown to play a role in the immunosuppression of naïve cells through increased tryptophan metabolism in the tumor microenvironment (TME). Increased expression of IDO1 can result in suppression of anti-tumor T-cells, differentiation of CD4+ T-cells into immunosuppressive regulatory T-cells (Tregs), and polarization of anti-gene cells into a tolerogenic phenotype. | Tissue | Response to IDO1 immune checkpoint inhibitors. | Prediction |
| [50] | HNSCC | OX40 (TNFRSF4) | Encodes tumor necrosis factor receptor, which is primarily expressed on the surface of activated T cells and is stimulated by its receptor OX40L, which is located on antigenic cells, activated T cells, lymphoid tissue inductor cells, certain endothelial cells, and mast cells. Its regulation induces differentiation, proliferation, and extended survival of T cell-activated cells through inhibition of activation-induced cell death, as well as stimulation of cytokine synthesis. | Tissue | Identifying HNSCC patients who would benefit from adjuvant immunotherapy. | Prediction |
| [50] | HNSCC | GITR (TNFRSF18, AITR) | Encodes the tumor necrosis factor receptor, is highly expressed by regulatory T cells (Tregs) and expressed at lower levels on naïve, effector, and memory T cells. Binding of the GITR ligand in combination with T cell receptor stimulation causes activation of the MAPK/ERK and NFkB pathway, resulting in immune system upregulation with T cell proliferation, production of proinflammatory cytokines, enhanced anti-cancer effector function, and resistance of CD4+ and CD8+ T cells. GITR promotes anti-cancer immunity by enhancing effector T cell function and by suppressing Treg proliferation. | Tissue | Identifying HNSCC patients who would benefit from adjuvant immunotherapy. | Prediction |
| [76] | HNSCC | DAPK | Encodes calcium/calmodulin (CaM)-regulated serine/threonine protein kinase, has pro-apoptotic function, mediates cell death triggered by a variety of death-inducers, including interferon-γ, 20 TGF, 21 TNFα, and Fas ligand. | Cell lines | Hypermethylation -low expression → resistance to cetuximab and erlotinib. | Prediction |
| [77] | HNSCC | DNA methylation | ILK signaling, glucocorticoid receptor signaling, fatty acid α-oxidation, cell cycle regulation. | Cell lines | Hypermethylation → radiation resistance. | Prediction |
| [78] | HNSCC | ZFG36 gene -TTP | Tumor-suppressor, encodes Arna-binding protein, enhances decay of AU-rich element (ARE)-containing transcripts, and plays an important role in cellular differentiation, proliferation, tumorigenesis, and immunity, modulates the posttranscriptional control of inflammatory mediators, and immune gene expression. | Cell lines | Hypermethylation-low expression → overexpression of CD47 in a radioresistant cell line (HN31R) → inhibition of phagocytosis. Prediction of the efficacy of CD47 antibody in recurrent HNSCC patients after radiotherapy. | Prediction |
| [79] | HNSCC | Gene panel (CRIP1, G0S2, MLH1, OPN3, S100 and TUBB2A) | CRIP1: Cysteine Rich Protein 1 G0S2: G0/G1 switch gene 2 Mlh1: MutL protein homolog 1 OPN3: Opsin 3 S100 TUBB2A: Tubulin Beta 2A Class Iia | Tissue | Hypermethylation → cisplatin-resistance. | Prediction |
| [80] | HNSCC | PTPRT | A member of the PTPR family, receptor of PTPs (enzymes that catalyze the removal of a phosphate group from specific signaling proteins), they cover the membrane once and contain one or two intracellular catalytic sites, as well as a modular extracellular region that typically contains several protein-protein interaction sites. | Tissue | Hypermethylation → sensitivity to STAT3 targeting agents. | Prediction |
| [81] | HNSCC | DNA methylation profile | Genes involved in different molecular pathways, namely Axon guidance, Hippo signaling, Pathways in cancer and MAPK signaling | Tissue | Both hypermethylation and hypomethylation → predict response to ICI. | Prediction |
| [82] | LA-HNSCC | DUSP2 | Encodes a nuclear phosphatase that is strongly expressed in activated immune cells and catalyzes the dephosphorylation of serine, threonine, and tyrosine residues on various types of mitogen-activated protein kinases inside MAPK TXY. | Tissue | Patients treated with CRT: Low EGFR + unmethylated DUSP2 → longer overall survival Compared to low EGFR + meth-DUSP2 Unmethylated DUSP2 + Mtp53 → longer survival compared to unmethylated DUSP2 + wtTP53. Methylated DUSP2 + high EGFR, + wild type TP53 → highest OS and HR. | Prediction |
| [83] | HNSCC | CCND1 | Encodes a regulator of the G1/S phase transition, it is degraded as the cell enters the S phase. | Tissue | Response to selective FGFR1/3 inhibitors PD 173074 and AZD4547. | Prediction |
| [83] | HNSCC | FGFR2 | Encodes a fibroblast growth factor receptor. | Tissue | Response to selective FGFR1/3 inhibitors PD 173074 and AZD4547. | Prediction |
| [83] | HNSCC | FGF5 | Encodes a cell signaling protein. | Tissue | Response to selective FGFR1/3 inhibitors PD 173074 and AZD4547. | Prediction |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liouta, G.; Adamaki, M.; Tsintarakis, A.; Zoumpourlis, P.; Liouta, A.; Agelaki, S.; Zoumpourlis, V. DNA Methylation as a Diagnostic, Prognostic, and Predictive Biomarker in Head and Neck Cancer. Int. J. Mol. Sci. 2023, 24, 2996. https://doi.org/10.3390/ijms24032996
Liouta G, Adamaki M, Tsintarakis A, Zoumpourlis P, Liouta A, Agelaki S, Zoumpourlis V. DNA Methylation as a Diagnostic, Prognostic, and Predictive Biomarker in Head and Neck Cancer. International Journal of Molecular Sciences. 2023; 24(3):2996. https://doi.org/10.3390/ijms24032996
Chicago/Turabian StyleLiouta, Galateia, Maria Adamaki, Antonis Tsintarakis, Panagiotis Zoumpourlis, Anastasia Liouta, Sofia Agelaki, and Vassilis Zoumpourlis. 2023. "DNA Methylation as a Diagnostic, Prognostic, and Predictive Biomarker in Head and Neck Cancer" International Journal of Molecular Sciences 24, no. 3: 2996. https://doi.org/10.3390/ijms24032996
APA StyleLiouta, G., Adamaki, M., Tsintarakis, A., Zoumpourlis, P., Liouta, A., Agelaki, S., & Zoumpourlis, V. (2023). DNA Methylation as a Diagnostic, Prognostic, and Predictive Biomarker in Head and Neck Cancer. International Journal of Molecular Sciences, 24(3), 2996. https://doi.org/10.3390/ijms24032996