
Immunopathological Mechanisms of Bird-Related Hypersensitivity Pneumonitis

 ,
,  ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
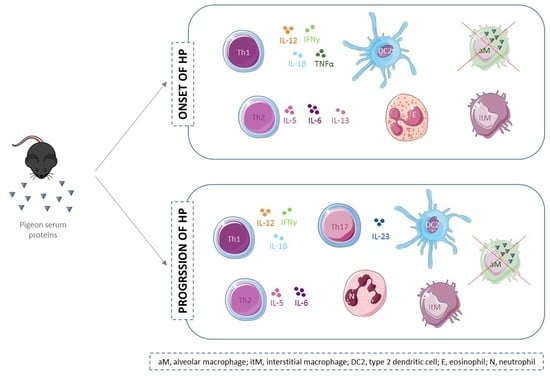
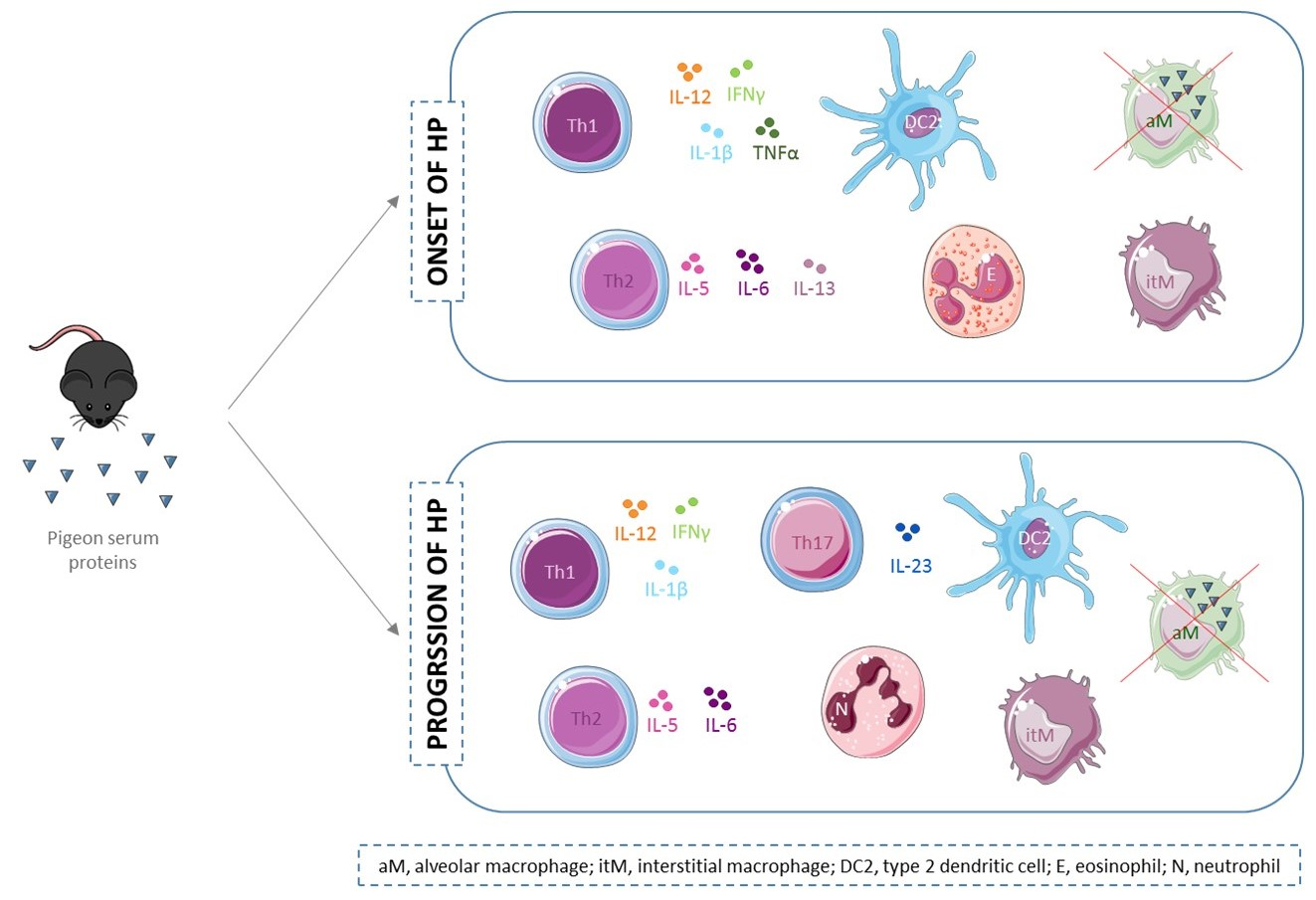
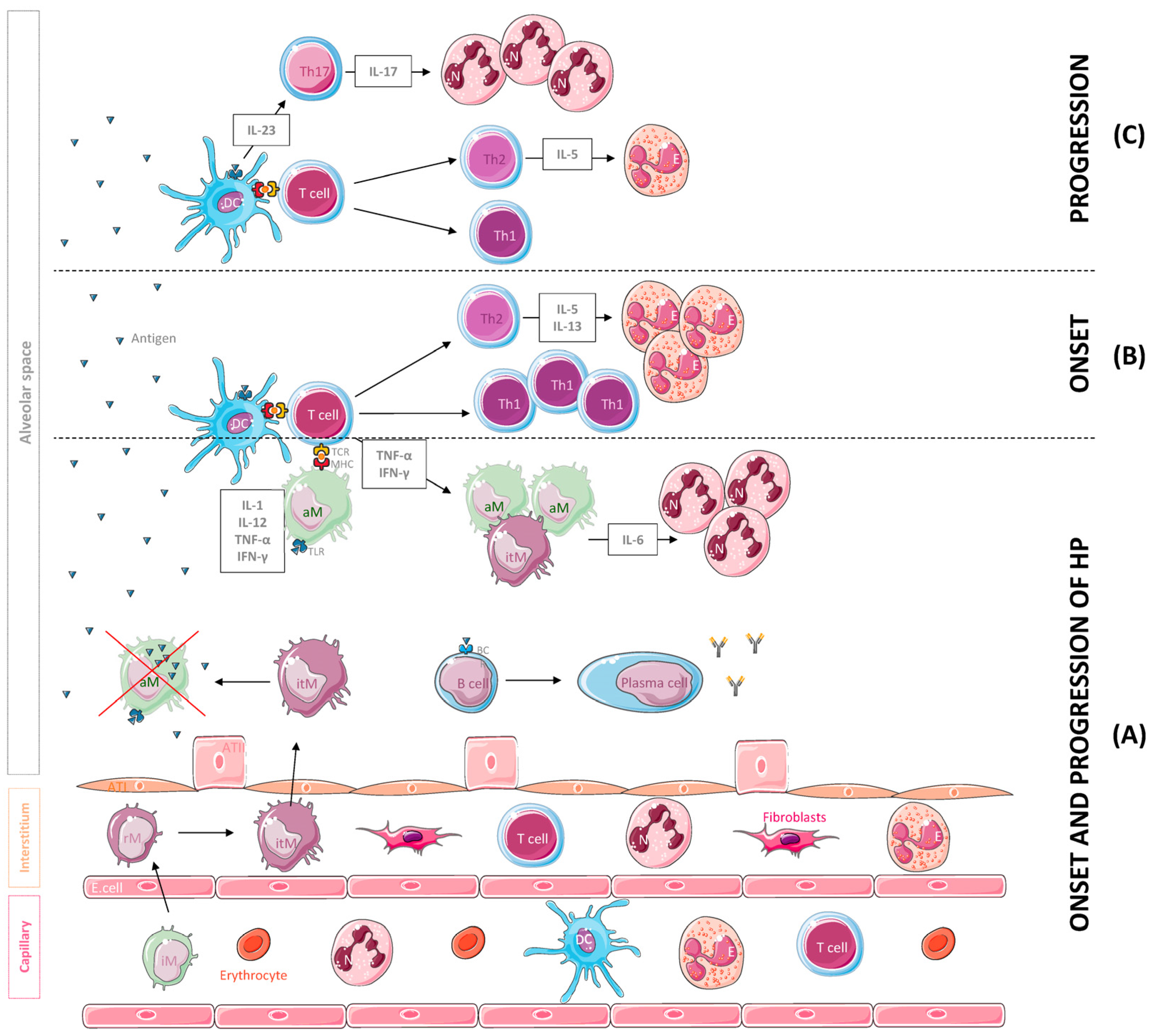
2. Results
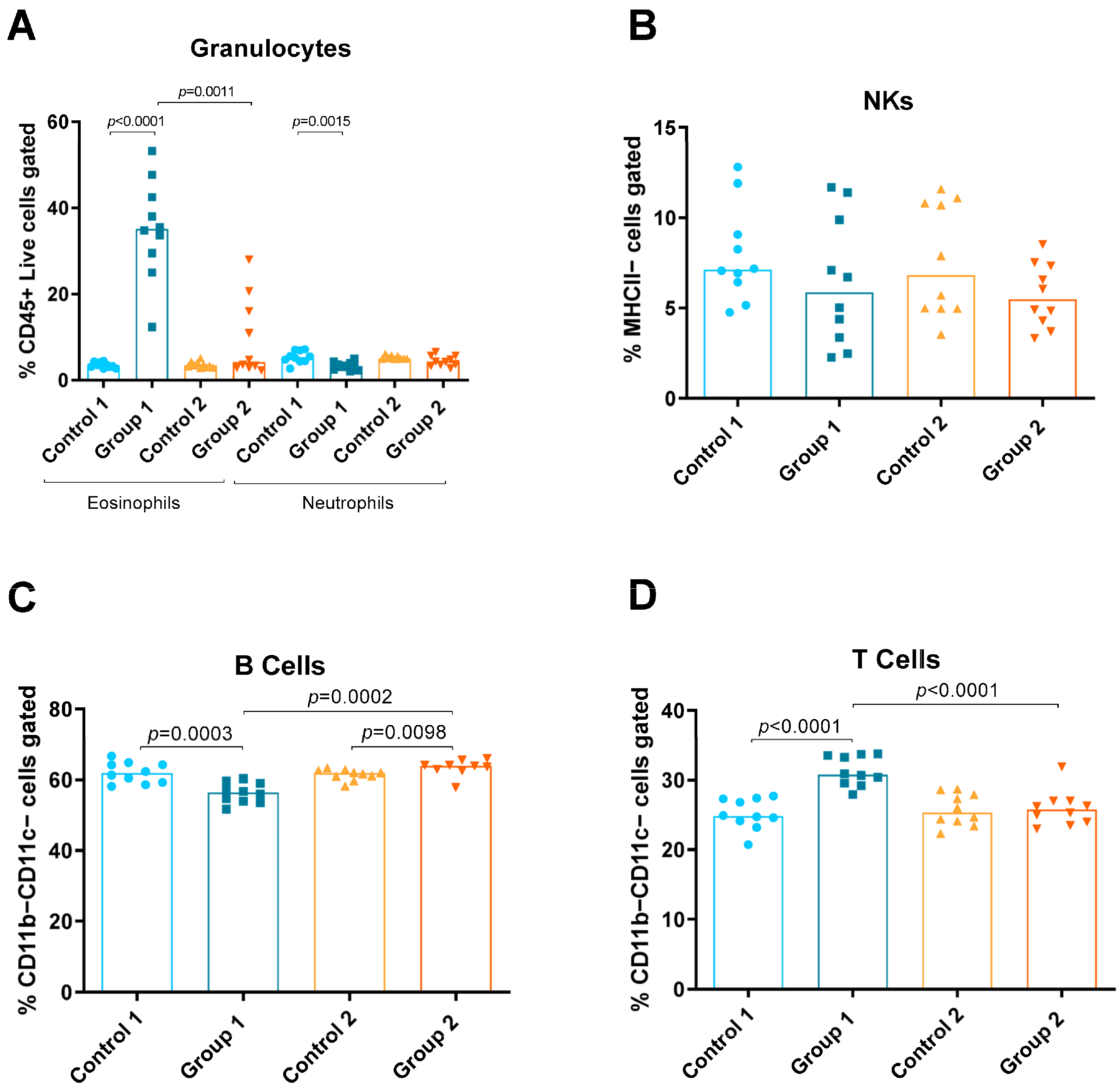
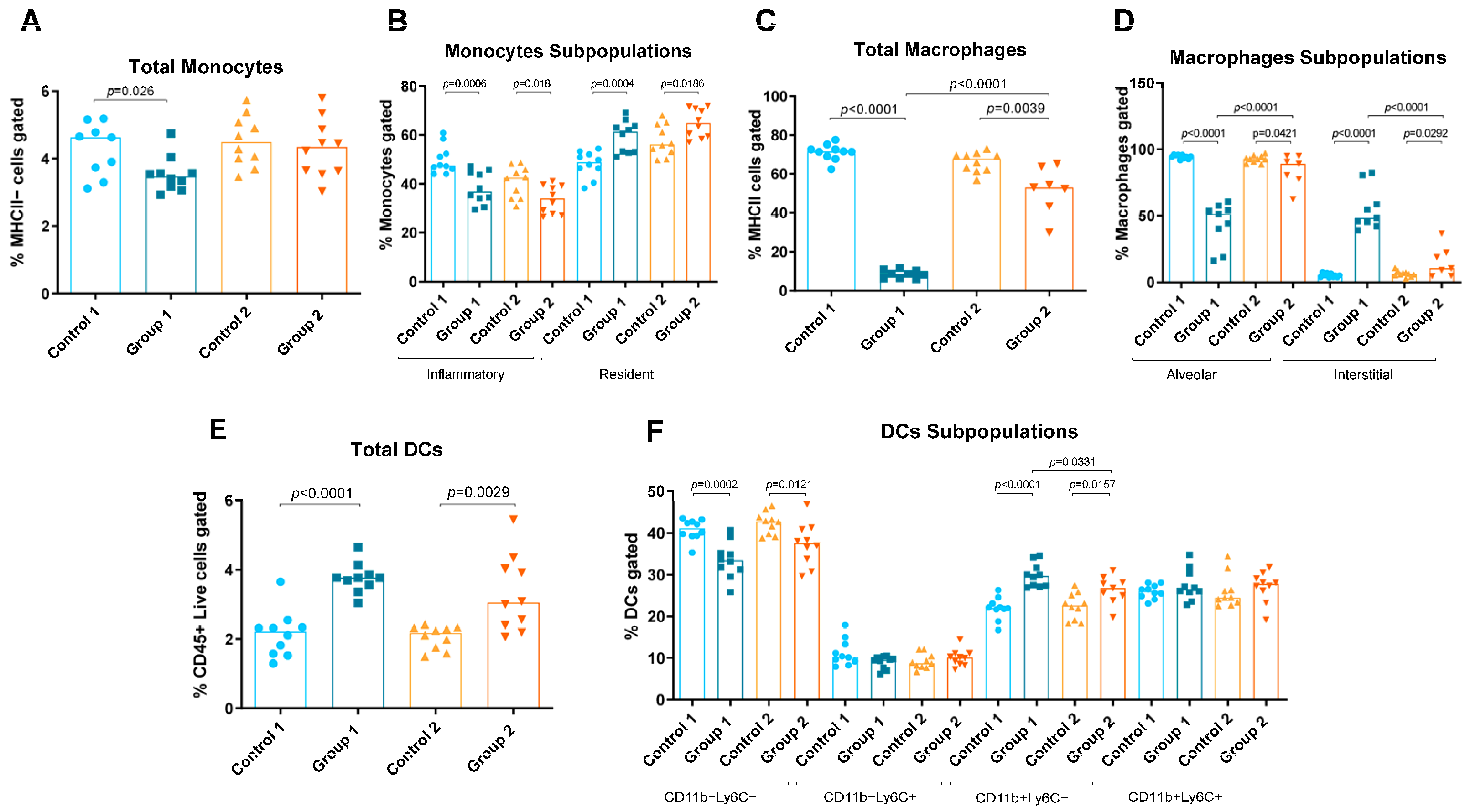
2.1. Flow Cytometry
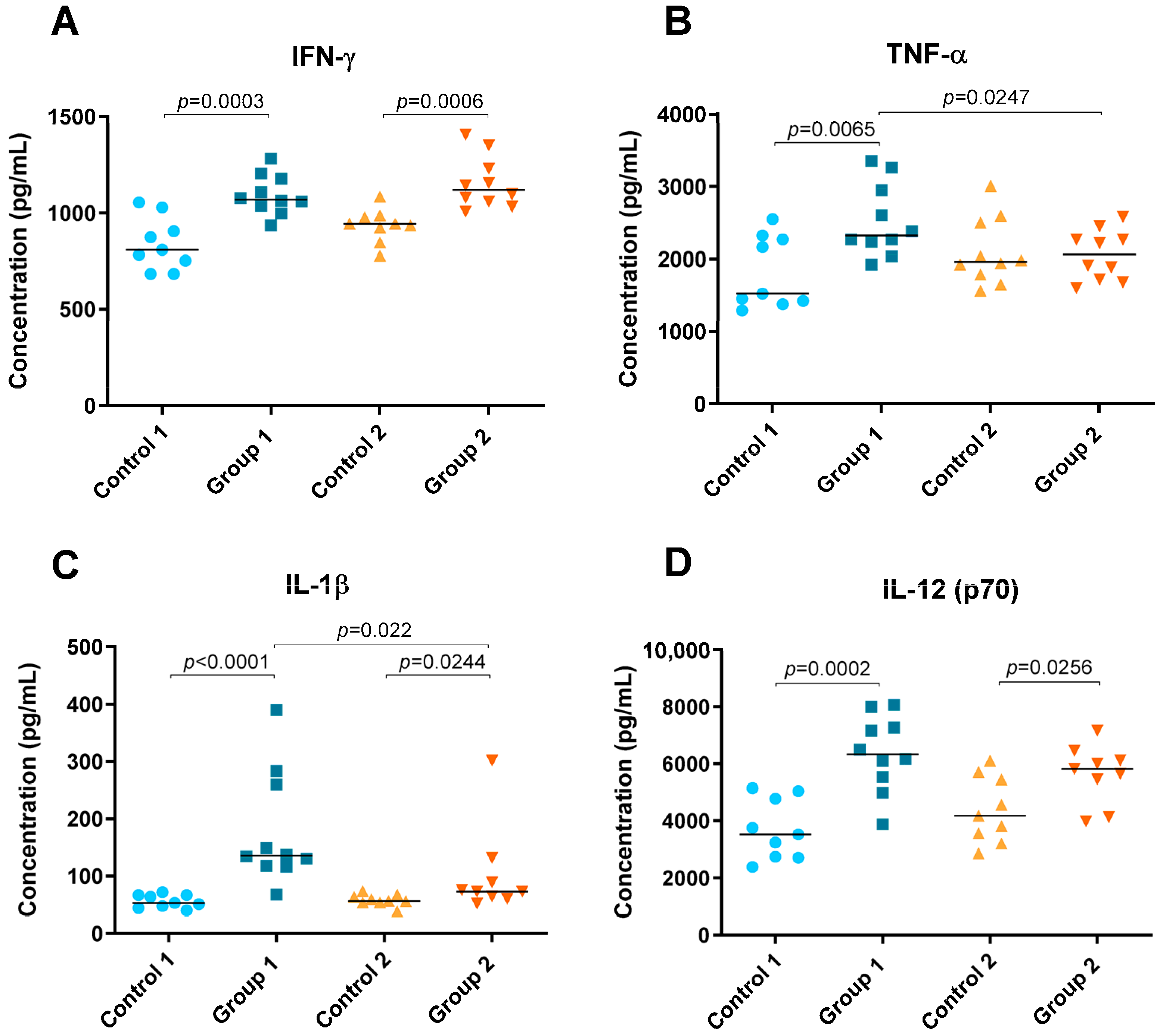
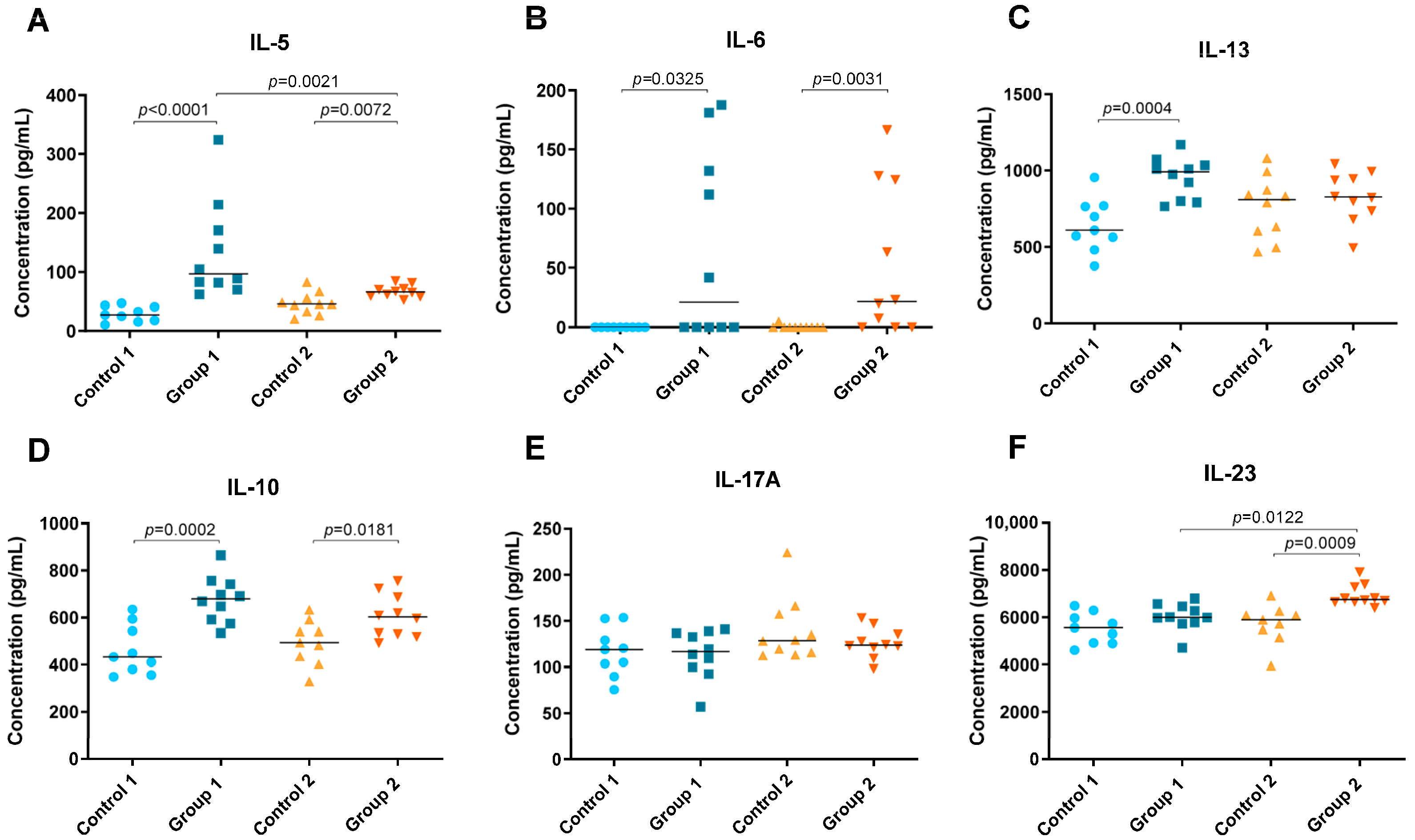
2.2. Cytokine Level Analysis
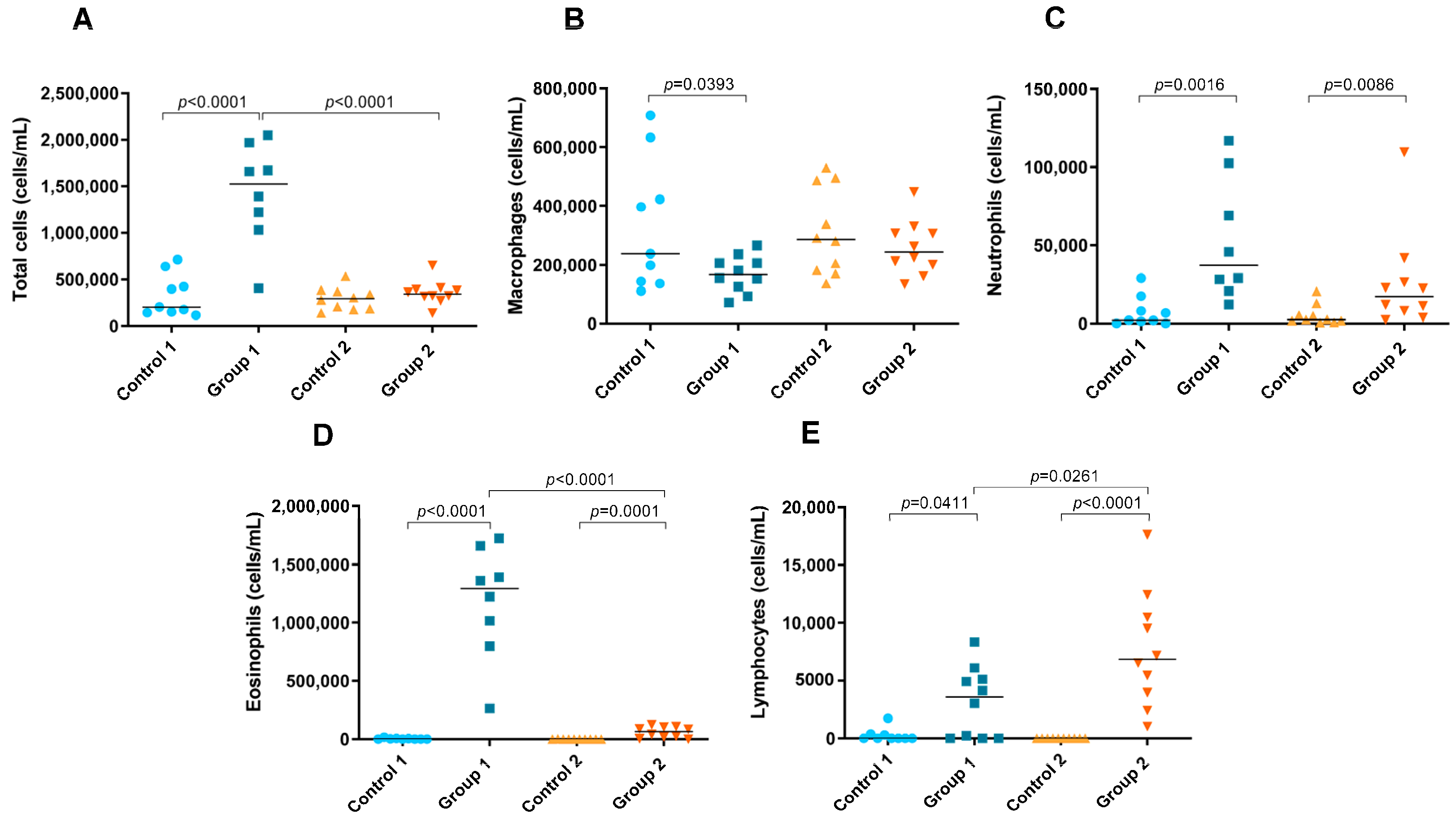
2.3. Total and Differential Cell Counts in BAL
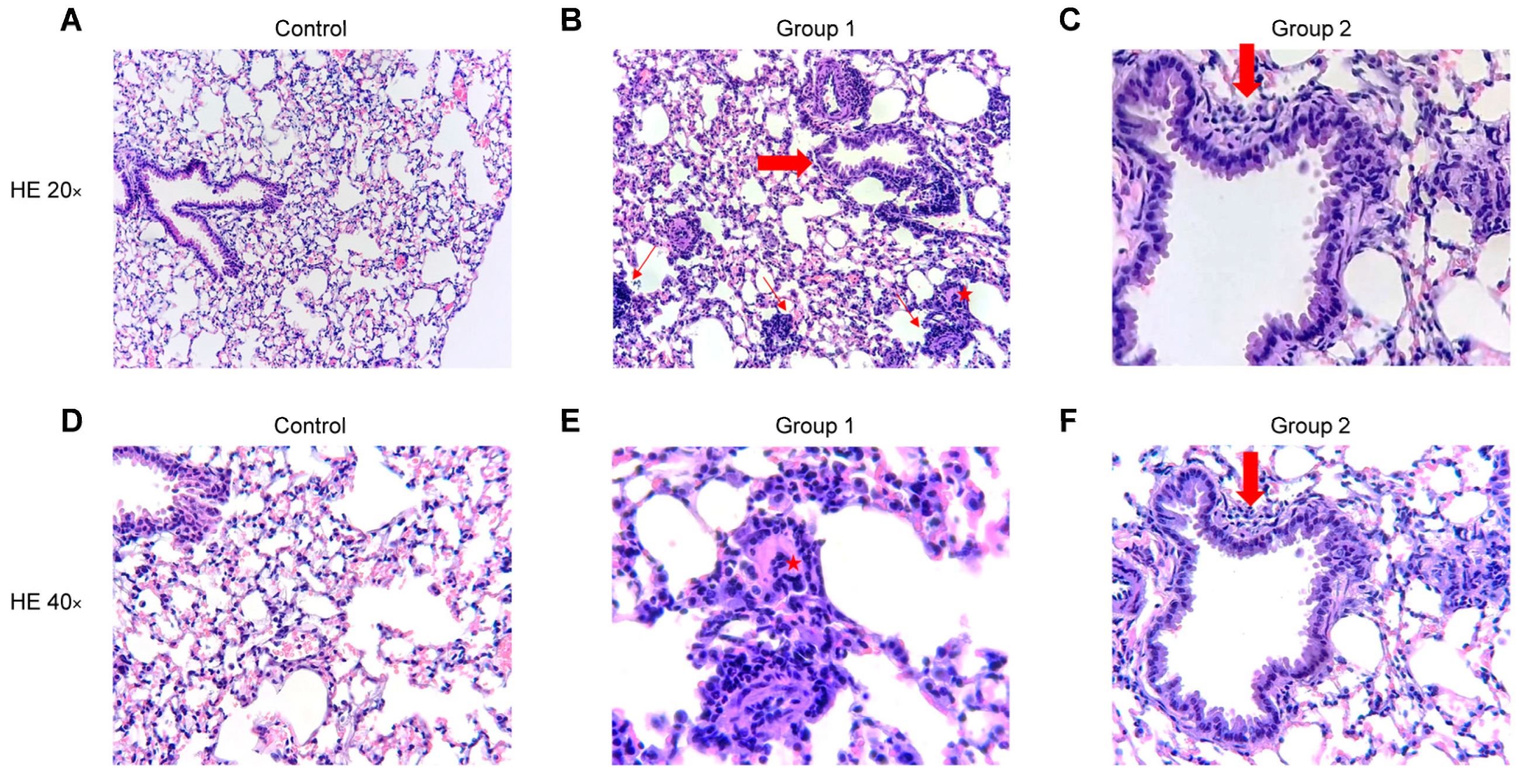
2.4. Histopathological Analysis
3. Discussion
4. Materials and Methods
4.1. Antigen Solution
4.2. Animals
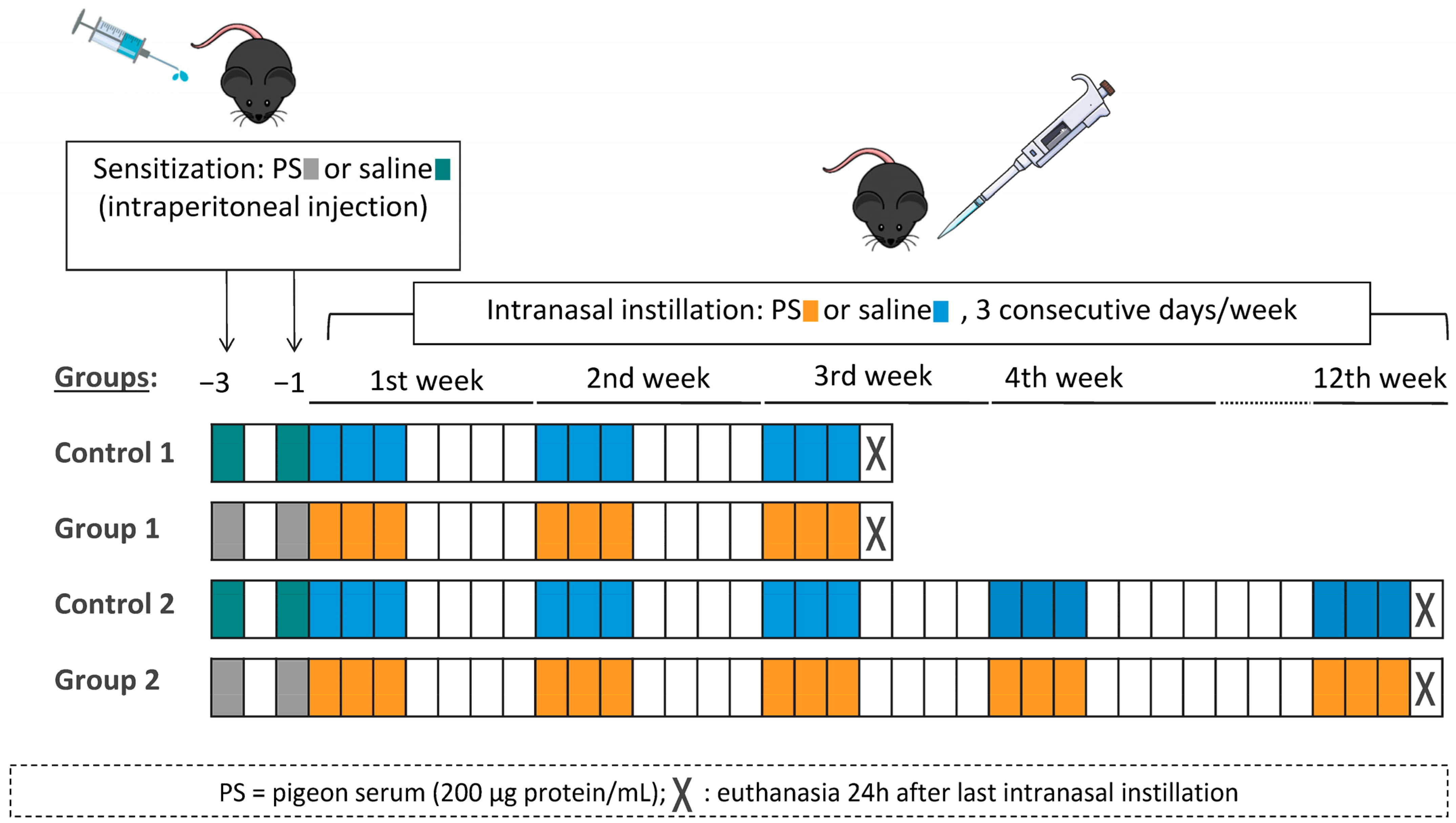
4.3. Experimental Design
4.4. Flow Cytometry
4.5. Lung Tissue Homogenate for Cytokine Level Analysis
4.6. Bronchoalveolar Lavage for Total and Differential Cell Counts
4.7. Histopathological Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quirce, S.; Vandenplas, O.; Campo, P.; Cruz, M.J.; de Blay, F.; Koschel, D.; Moscato, G.; Pala, G.; Raulf, M.; Sastre, J.; et al. Occupational hypersensitivity pneumonitis: An EAACI position paper. Allergy 2016, 71, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Vasakova, M.; Selman, M.; Morell, F.; Sterclova, M.; Molina-Molina, M.; Raghu, G. Hypersensitivity pneumonitis: Current concepts of pathogenesis and potential targets for treatment. Am. J. Respir. Crit. Care Med. 2019, 200, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Vasakova, M.; Morell, F.; Walsh, S.; Leslie, K.; Raghu, G. Hypersensitivity Pneumonitis: Perspectives in Diagnosis and Management. Am. J. Respir. Crit. Care Med. 2017, 196, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Morell, F.; Roger, À.; Reyes, L.; Cruz, M.J.; Murio, C.; Muñoz, X. Bird fancier’s lung: A series of 86 patients. Medicine 2008, 87, 110–130. [Google Scholar] [CrossRef]
- Raghu, G.; Wilson, K.C.; Bargagli, E.; Bendstrup, E.; Chami, H.A.; Chua, A.T.; Chung, J.H.; Collins, B.F.; Corte, T.J.; Dalphin, J.C.; et al. Diagnosis of hypersensitivity pneumonitis in adults: An official ATS/JRS/ALAT clinical practice guideline. Am. J. Respir. Crit. Care Med. 2020, 202, E36–E69. [Google Scholar] [CrossRef]
- Selman, M.; Pardo, A.; King, T.E. Hypersensitivity pneumonitis: Insights in diagnosis and pathobiology. Am. J. Respir. Crit. Care Med. 2012, 186, 314–324. [Google Scholar] [CrossRef]
- Woda, B.A. Hypersensitivity pneumonitis: An immunopathology review. Arch. Pathol. Lab. Med. 2008, 132, 204–205. [Google Scholar] [CrossRef]
- Hasan, S.A.; Eksteen, B.; Reid, D.; Paine, H.V.; Alansary, A.; Johannson, K.; Gwozd, C.; Goring, K.A.R.; Vo, T.; Proud, D.; et al. Role of IL-17A and neutrophils in fibrosis in experimental hypersensitivity pneumonitis. J. Allergy Clin. Immunol. 2013, 131, 1663–1673. [Google Scholar] [CrossRef]
- Girard, M.; Israël-Assayag, E.; Cormier, Y. Impaired function of regulatory T-cells in hypersensitivity pneumonitis. Eur. Respir. J. 2011, 37, 632–639. [Google Scholar] [CrossRef]
- Yu, B.; Yang, X.; Li, F.; Wu, C.; Wang, W.; Ding, W. Significance of Foxp3+CD4+ regulatory T cells in the peripheral blood of Uygur patients in the acute and chronic phases of pigeon breeder’s lung. Bosn. J. Basic Med. Sci. 2017, 17, 17–22. [Google Scholar] [CrossRef]
- Barrera, L.; Mendoza, F.; Zuñiga, J.; Estrada, A.; Zamora, A.C.; Melendro, E.I.; Ramírez, R.; Pardo, A.; Selman, M. Functional Diversity of T-Cell Subpopulations in Subacute and Chronic Hypersensitivity Pneumonitis. Am. J. Respir. Crit. Care Med. 2008, 177, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Mitaka, K.; Miyazaki, Y.; Yasui, M.; Furuie, M.; Miyake, S.; Inase, N.; Yoshizawa, Y. Th2-Biased Immune Responses Are Important in a Murine Model of Chronic Hypersensitivity Pneumonitis. Int. Arch. Allergy Immunol. 2011, 154, 264–274. [Google Scholar] [CrossRef]
- Hwang, S.J.; Kim, H.S.; Chung, D.H. Fas/Fas ligand-mediated apoptosis promotes hypersensitivity pneumonitis in mice by enhancing maturation of dendritic cells. Am. J. Respir. Crit. Care Med. 2010, 181, 1250–1261. [Google Scholar] [CrossRef] [PubMed]
- De García Alba, C.; Buendia-Roldán, I.; Salgado, A.; Becerril, C.; Ramírez, R.; González, Y.; Checa, M.; Navarro, C.; Ruiz, V.; Pardo, A.; et al. Fibrocytes contribute to inflammation and fibrosis in chronic hypersensitivity pneumonitis through paracrine effects. Am. J. Respir. Crit. Care Med. 2015, 191, 427–436. [Google Scholar] [CrossRef]
- Sánchez-Ortiz, M.; Cruz, M.J.; Sánchez-Díez, S.; Villar, A.; Ojanguren, I.; Muñoz, X. Immunomodulatory effect of pigeon serum in an acute and chronic murine model of bird fanciers lung. Environ. Res. 2020, 182, 108981. [Google Scholar] [CrossRef] [PubMed]
- de Homdedeu, M.; Cruz, M.; Sanchez-Díez, S.; Ojanguren, I.; Romero-Mesones, C.; Vanoirbeek, J.; Vande Velde, G.; Muñoz, X. The immunomodulatory effects of diesel exhaust particles in asthma. Environ. Pollut. 2020, 263 Pt A, 114600. [Google Scholar] [CrossRef]
- Ishiguro, T.; Yasui, M.; Nakade, Y.; Kimura, H.; Katayama, N.; Kasahara, K.; Fujimura, M. Extrinsic allergic alveolitis with eosinophil infiltration induced by 1,1,1,2-tetrafluoroethane (HFC-134a): A case report. Intern. Med. 2007, 46, 1455–1457. [Google Scholar] [CrossRef][Green Version]
- Drent, M.; Wagenaar, S.S.; Van Velzen-Blad, H.; Mulder, P.G.H.; Hoogsteden, H.C.; Van Den Bosch, J.M.M. Relationship between plasma cell levels and profile of bronchoalveolar lavage fluid in patients with subacute extrinsic allergic alveolitis. Thorax 1993, 48, 835–839. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Villar, A.; Muñoz, X.; Sanchez-Vidaurre, S.; Gómez-Ollés, S.; Morell, F.; Cruz, M.-J. Bronchial inflammation in hypersensitivity pneumonitis after antigen-specific inhalation challenge. Respirology 2014, 19, 891–899. [Google Scholar] [CrossRef]
- Kouro, T.; Takatsu, K. IL-5- and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef]
- Horie, S.; Okubo, Y.; Hossain, M.; Sato, E.; Nomura, H.; Koyama, S.; Suzuki, J.I.; Isobe, M.; Sekiguchi, M. Interleukin-13 but not interleukin-4 prolongs eosinophil survival and induces eosinophil chemotaxis. Intern. Med. 1997, 36, 179–185. [Google Scholar] [CrossRef]
- Luttmann, W.; Matthiesen, T.; Matthys, H.; Virchow, J.C. Synergistic effects of interleukin-4 or interleukin-13 and tumor necrosis factor-α on eosinophil activation in vitro. Am. J. Respir. Cell Mol. Biol. 1999, 20, 474–480. [Google Scholar] [CrossRef]
- Adderley, N.; Humphreys, C.J.; Barnes, H.; Ley, B.; Premji, Z.A.; Johannson, K.A. Bronchoalveolar lavage fluid lymphocytosis in chronic hypersensitivity pneumonitis: A systematic review and meta-analysis. Eur. Respir. J. 2020, 56, 2000206. [Google Scholar] [CrossRef] [PubMed]
- Costabel, U.; Bross, K.J.; Marxen, J.; Matthys, H. T-lymphocytes in bronchoalveolar lavage fluid of hypersensitivity pneumonitis. Changes in profile of T-cell subsets during the course of disease. Chest 1984, 85, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Ghilardi, N.; Xie, M.H.; De Sauvage, F.J.; Gurney, A.L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 2003, 278, 1910–1914. [Google Scholar] [CrossRef]
- Smith, E.; Zarbock, A.; Stark, M.A.; Burcin, T.L.; Bruce, A.C.; Foley, P.; Ley, K. IL-23 Is Required for Neutrophil Homeostasis in Normal and Neutrophilic Mice. J. Immunol. 2007, 179, 8274–8279. [Google Scholar] [CrossRef]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, M.; Miyazaki, Y.; Masuo, M.; Suhara, K.; Tateishi, T.; Yasui, M.; Inase, N. Interleukin-17A and Neutrophils in a Murine Model of Bird-Related Hypersensitivity Pneumonitis. PLoS ONE 2015, 10, e0137978. [Google Scholar] [CrossRef]
- Tsai, D.Y.; Hung, K.H.; Chang, C.W.; Lin, K.I. Regulatory mechanisms of B cell responses and the implication in B cell-related diseases. J. Biomed. Sci. 2019, 26, 64. [Google Scholar] [CrossRef]
- Fenoglio, C.M.; Reboux, G.; Sudre, B.; Mercier, M.; Roussel, S.; Cordier, J.F.; Piarroux, R.; Dalphin, J.C. Diagnostic value of serum precipitins to mould antigens in active hypersensitivity pneumonitis. Eur. Respir. J. 2007, 29, 706–712. [Google Scholar] [CrossRef]
- Terho, E.O.; Lindström, P.; Mäntyjärvi, R.; Tukiainen, H.; Wager, O. Circulating Immune Complexes and Rheumatoid Factors in Patients with Farmer’s Lung. Allergy 1983, 38, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Costabel, U.; Miyazaki, Y.; Pardo, A.; Koschel, D.; Bonella, F.; Spagnolo, P.; Guzman, J.; Ryerson, C.J.; Selman, M. Hypersensitivity pneumonitis. Nat. Rev. Dis. Prim. 2020, 6, 65. [Google Scholar] [CrossRef]
- Sema, M.; Miyazaki, Y.; Tsutsui, T.; Tomita, M.; Eishi, Y.; Inase, N. Environmental levels of avian antigen are relevant to the progression of chronic hypersensitivity pneumonitis during antigen avoidance. Immun. Inflamm. Dis. 2018, 6, 154–162. [Google Scholar] [CrossRef]
- Landsman, L.; Jung, S. Lung Macrophages Serve as Obligatory Intermediate between Blood Monocytes and Alveolar Macrophages. J. Immunol. 2007, 179, 3488–3494. [Google Scholar] [CrossRef] [PubMed]
- Fossum, E.; Grødeland, G.; Terhorst, D.; Tveita, A.A.; Vikse, E.; Mjaaland, S.; Henri, S.; Malissen, B.; Bogen, B. Vaccine molecules targeting Xcr1 on cross-presenting DCs induce protective CD8+ T-cell responses against influenza virus. Eur. J. Immunol. 2015, 45, 624–635. [Google Scholar] [CrossRef]
- Macri, C.; Pang, E.S.; Patton, T.; O’Keeffe, M. Dendritic cell subsets. Semin. Cell Dev. Biol. 2018, 84, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Mesnil, C.; Sabatel, C.M.; Marichal, T.; Toussaint, M.; Cataldo, D.; Drion, P.V.; Lekeux, P.; Bureau, F.; Desmet, C.J. Resident CD11b+Ly6C- Lung Dendritic Cells Are Responsible for Allergic Airway Sensitization to House Dust Mite in Mice. PLoS ONE 2012, 7, e53242. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, A.; McGovern, N.; Teo, P.; Zelante, T.; Atarashi, K.; Low, D.; Ho, A.W.S.; See, P.; Shin, A.; Wasan, P.S.; et al. IRF4 Transcription Factor-Dependent CD11b+ Dendritic Cells in Human and Mouse Control Mucosal IL-17 Cytokine Responses. Immunity 2013, 38, 970–983. [Google Scholar] [CrossRef]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Benavent, M.I.; Cruz, M.J.; Rosell, M.; Murio, C.; Pascual, C.; Morell, F. Detection of specific antibodies to pigeon serum and bloom antigens by enzyme linked immunosorbent assay in pigeon breeder’s disease. Occup. Environ. Med. 2000, 57, 159–164. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Díez, S.; Cruz, M.J.; de Homdedeu, M.; Ojanguren, I.; Romero-Mesones, C.; Sansano, I.; Muñoz, X. Immunopathological Mechanisms of Bird-Related Hypersensitivity Pneumonitis. Int. J. Mol. Sci. 2023, 24, 2884. https://doi.org/10.3390/ijms24032884
Sánchez-Díez S, Cruz MJ, de Homdedeu M, Ojanguren I, Romero-Mesones C, Sansano I, Muñoz X. Immunopathological Mechanisms of Bird-Related Hypersensitivity Pneumonitis. International Journal of Molecular Sciences. 2023; 24(3):2884. https://doi.org/10.3390/ijms24032884
Chicago/Turabian StyleSánchez-Díez, Silvia, María Jesús Cruz, Miquel de Homdedeu, Iñigo Ojanguren, Christian Romero-Mesones, Irene Sansano, and Xavier Muñoz. 2023. "Immunopathological Mechanisms of Bird-Related Hypersensitivity Pneumonitis" International Journal of Molecular Sciences 24, no. 3: 2884. https://doi.org/10.3390/ijms24032884
APA StyleSánchez-Díez, S., Cruz, M. J., de Homdedeu, M., Ojanguren, I., Romero-Mesones, C., Sansano, I., & Muñoz, X. (2023). Immunopathological Mechanisms of Bird-Related Hypersensitivity Pneumonitis. International Journal of Molecular Sciences, 24(3), 2884. https://doi.org/10.3390/ijms24032884