

A Revised View of the LSU Gene Family: New Functions in Plant Stress Responses and Phytohormone Signaling


Abstract
1. Introduction
2. General Features and Evolutionary History of the LSU Gene Family
2.1. The Discovery of the LSU Gene Family
2.2. Evolution of LSU Gene Family
3. Functional Analyses of the LSU Family
3.1. Subcellular Localization of LSU Proteins
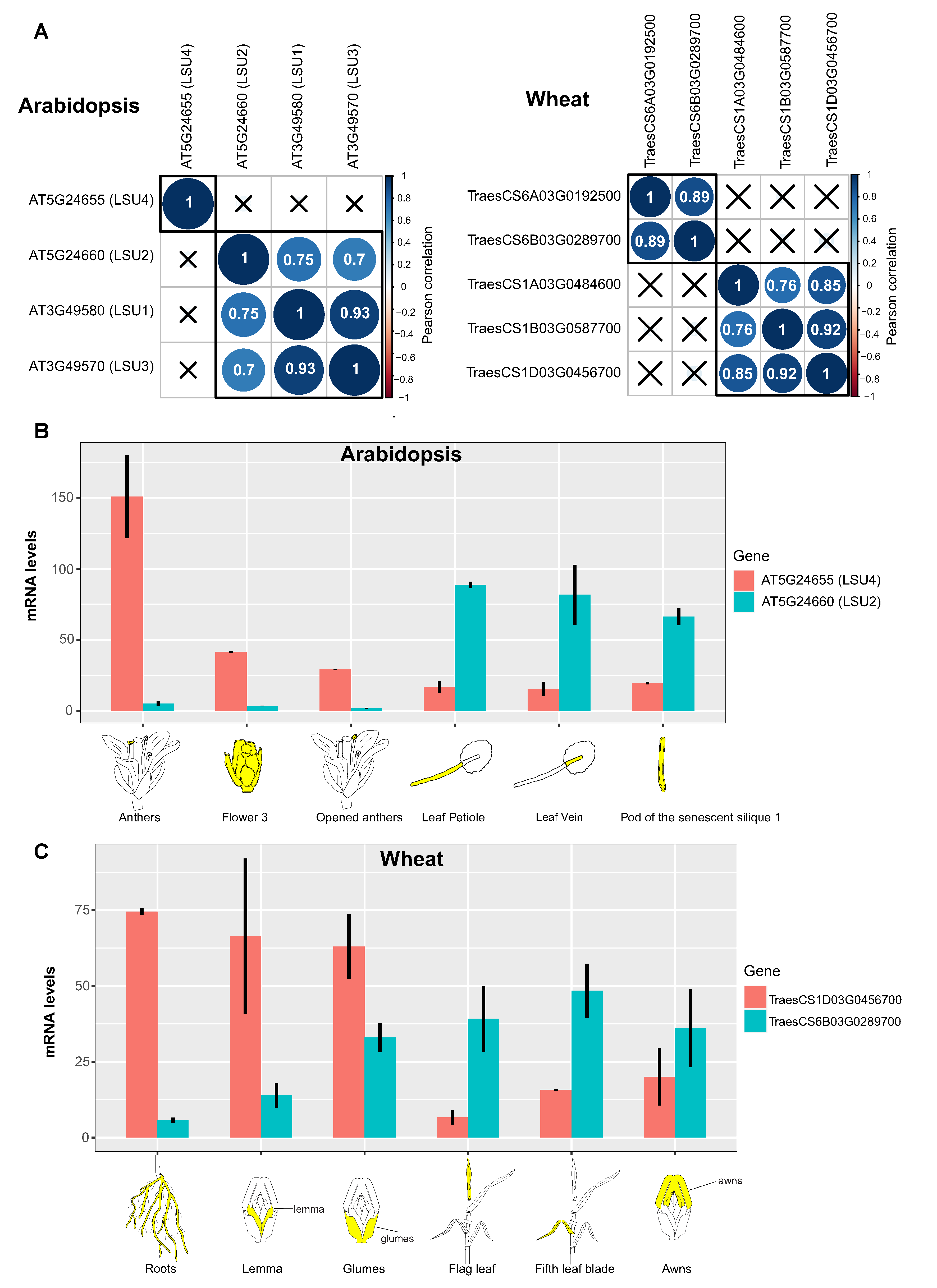
3.2. Different Members of the LSU Gene Family Showed Tissue-Specific Expression
3.3. Functional Analyses of LSUs in Arabidopsis
3.4. Functional Analyses of LSUs in Other Plants
4. LSU Protein Interactions and Phytohormone Signaling
4.1. LSU Protein Interactions Suggest Some Degree of Specialization within this Family in Arabidopsis
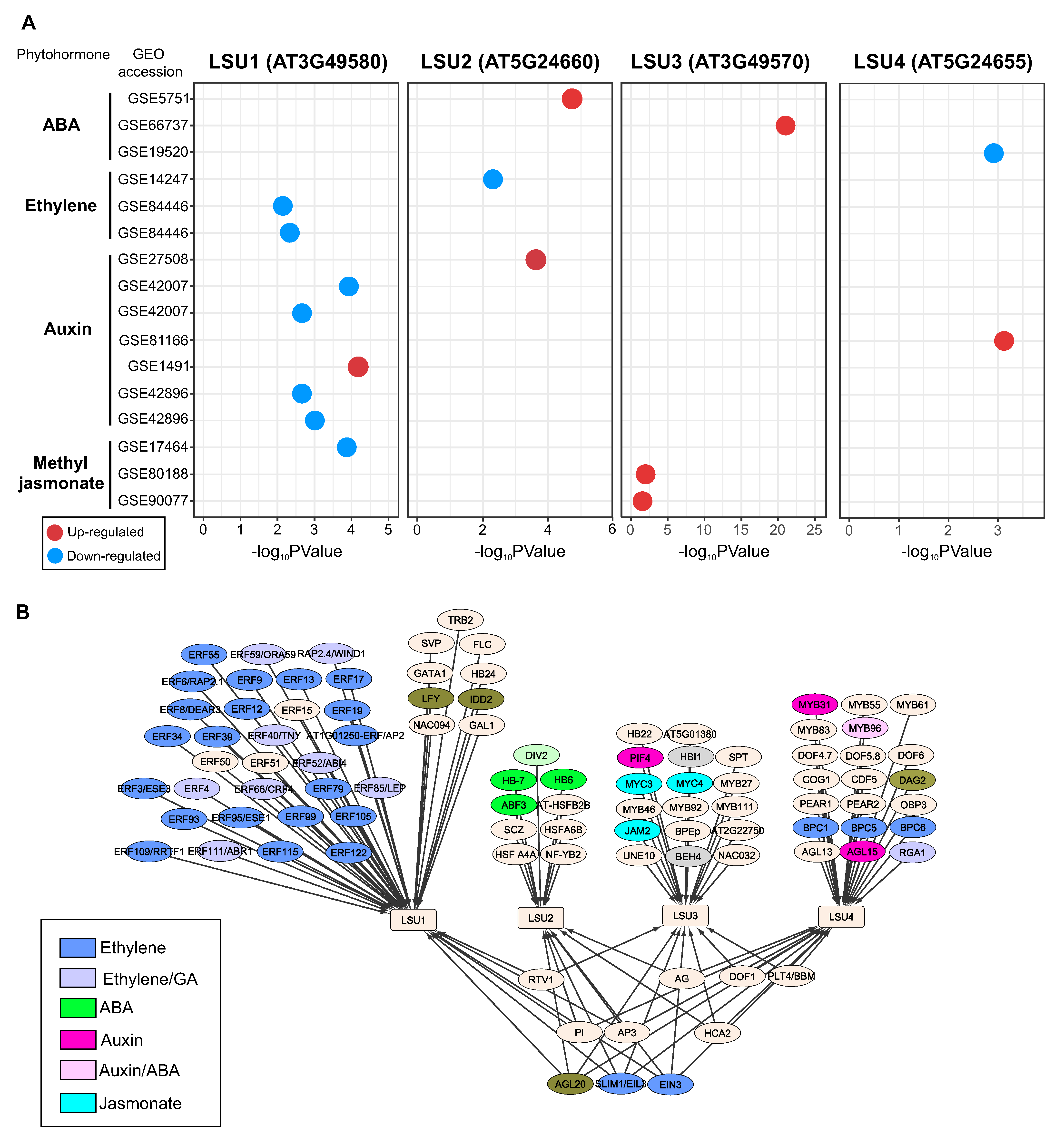
4.2. Crosstalk between LSUs and Phytohormones
4.3. Differential Expression of Arabidopsis LSU Genes in Response to Phytohormone Treatment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Pramanik, K.; Panda, D.; Dutta, D.; Karmakar, S.; Bose, B. Sulfur in Seeds: An Overview. Plants 2022, 11, 450. [Google Scholar] [CrossRef] [PubMed]
- Aarabi, F.; Naake, T.; Fernie, A.R.; Hoefgen, R. Coordinating Sulfur Pools under Sulfate Deprivation. Trends Plant Sci. 2020, 25, 1227–1239. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A. Metabolic changes sustain the plant life in low-sulfur environments. Curr. Opin. Plant Biol. 2017, 39, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Juhasz, A.; Islam, S.; Diepeveen, D.; Zhang, J.; Wang, P.; Ma, W. Impact of mid-season sulphur deficiency on wheat nitrogen metabolism and biosynthesis of grain protein. Sci. Rep. 2018, 8, 2499. [Google Scholar] [CrossRef]
- Watanabe, M.; Hoefgen, R. Sulphur systems biology—Making sense of omics data. J. Exp. Bot. 2019, 70, 4155–4170. [Google Scholar] [CrossRef]
- Henríquez-Valencia, C.; Arenas-M, A.; Medina, J.; Canales, J. Integrative Transcriptomic Analysis Uncovers Novel Gene Modules That Underlie the Sulfate Response in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 470. [Google Scholar] [CrossRef]
- Arabidopsis Interactome Mapping Consortium; Dreze, M.; Carvunis, A.R.; Charloteaux, B.; Galli, M.; Pevzner, S.J.; Tasan, M.; Ahn, Y.Y.; Balumuri, P.; Barabási, A.L.; et al. Evidence for network evolution in an Arabidopsis interactome map. Science 2011, 333, 601–607. [Google Scholar] [CrossRef]
- Mukhtar, M.S.; Carvunis, A.R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently Evolved Virulence Effectors Converge onto Hubs in a Plant Immune System Network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef]
- Vandereyken, K.; Leene, J.V.; Coninck, B.D.; Cammue, B.P.A. Hub Protein Controversy: Taking a Closer Look at Plant Stress Response Hubs. Front. Plant Sci. 2018, 9, 694. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Niemiro, A.; Cysewski, D.; Brzywczy, J.; Wawrzyńska, A.; Sieńko, M.; Poznański, J.; Sirko, A. Similar but Not Identical—Binding Properties of LSU (Response to Low Sulfur) Proteins From Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1246. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko, G.; Skoneczny, M.; Zientara-Rytter, K.; Wawrzyńska, A.; Głów, D.; Cristescu, S.M.; Harren, F.J.M.; Sirko, A. Tobacco LSU-like protein couples sulphur-deficiency response with ethylene signalling pathway. J. Exp. Bot. 2013, 64, 5173–5182. [Google Scholar] [CrossRef] [PubMed]
- Sirko, A.; Wawrzynska, A.; Rodriguez, M.C.; Sęktas, P. The family of LSU-like proteins. Front. Plant Sci. 2015, 5, 774. [Google Scholar] [CrossRef] [PubMed]
- Maruyama-Nakashita, A.; Nakamura, Y.; Watanabe-Takahashi, A.; Inoue, E.; Yamaya, T.; Takahashi, H. Identification of a novel cis-acting element conferring sulfur deficiency response in Arabidopsis roots. Plant J. 2005, 42, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyńska, A.; Lewandowska, M.; Hawkesford, M.J.; Sirko, A. Using a suppression subtractive library-based approach to identify tobacco genes regulated in response to short-term sulphur deficit. J. Exp. Bot. 2005, 56, 1575–1590. [Google Scholar] [CrossRef]
- Uribe, F.; Henríquez-Valencia, C.; Arenas-M, A.; Medina, J.; Vidal, E.A.; Canales, J. Evolutionary and Gene Expression Analyses Reveal New Insights into the Role of LSU Gene-Family in Plant Responses to Sulfate-Deficiency. Plants 2022, 11, 1526. [Google Scholar] [CrossRef]
- Bel, M.V.; Silvestri, F.; Weitz, E.M.; Kreft, L.; Botzki, A.; Coppens, F.; Vandepoele, K. PLAZA 5.0: Extending the scope and power of comparative and functional genomics in plants. Nucleic Acids Res. 2021, 50, D1468–D1474. [Google Scholar] [CrossRef]
- Garcia-Molina, A.; Altmann, M.; Alkofer, A.; Epple, P.M.; Dangl, J.L.; Falter-Braun, P. LSU network hubs integrate abiotic and biotic stress responses via interaction with the superoxide dismutase FSD2. J. Exp. Bot. 2017, 68, 1185–1197. [Google Scholar] [CrossRef]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.K.; Aryamanesh, N.; Millar, A.H. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res. 2016, 45, D1064–D1074. [Google Scholar] [CrossRef]
- Waese, J.; Fan, J.; Pasha, A.; Yu, H.; Fucile, G.; Shi, R.; Cumming, M.; Kelley, L.A.; Sternberg, M.J.; Krishnakumar, V.; et al. ePlant: Visualizing and Exploring Multiple Levels of Data for Hypothesis Generation in Plant Biology. Plant Cell 2017, 29, 1806–1821. [Google Scholar] [CrossRef] [PubMed]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix. (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 15 December 2022).
- Oblessuc, P.R.; Bisneta, M.V.; Melotto, M. Common and unique Arabidopsis proteins involved in stomatal susceptibility to Salmonella enterica Pseudomonas syringae. FEMS Microbiol. Lett. 2019, 366, fnz197. [Google Scholar] [CrossRef]
- Ruckle, M.E.; Burgoon, L.D.; Lawrence, L.A.; Sinkler, C.A.; Larkin, R.M. Plastids Are Major Regulators of Light Signaling in Arabidopsis. Plant Physiol. 2012, 159, 366–390. [Google Scholar] [CrossRef] [PubMed]
- Gläßer, C.; Haberer, G.; Finkemeier, I.; Pfannschmidt, T.; Kleine, T.; Leister, D.; Dietz, K.J.; Häusler, R.E.; Grimm, B.; Mayer, K.F.X. Meta-Analysis of Retrograde Signaling in Arabidopsis thaliana Reveals a Core Module of Genes Embedded in Complex Cellular Signaling Networks. Mol. Plant 2014, 7, 1167–1190. [Google Scholar] [CrossRef] [PubMed]
- Myakushina, Y.A.; Milyaeva, E.L.; Romanov, G.A.; Nikiforova, V.Y. Mutation in LSU4 gene affects flower development in Arabidopsis thaliana. Dokl. Biochem. Biophys. 2009, 428, 257–260. [Google Scholar] [CrossRef]
- Andrés-Barrao, C.; Alzubaidy, H.; Jalal, R.; Mariappan, K.G.; de Zélicourt, A.; Bokhari, A.; Artyukh, O.; Alwutayd, K.; Rawat, A.; Shekhawat, K.; et al. Coordinated bacterial and plant sulfur metabolism in Enterobacter sp. SA187–induced plant salt stress tolerance. Proc. Natl. Acad. Sci. USA 2021, 118, e2107417118. [Google Scholar] [CrossRef]
- Lewandowska, M.; Wawrzyńska, A.; Moniuszko, G.; Łukomska, J.; Zientara, K.; Piecho, M.; Hodurek, P.; Zhukov, I.; Liszewska, F.; Nikiforova, V.; et al. A Contribution to Identification of Novel Regulators of Plant Response to Sulfur Deficiency: Characteristics of a Tobacco Gene UP9C Its Protein Product and the Effects of UP9C Silencing. Mol. Plant 2010, 3, 347–360. [Google Scholar] [CrossRef]
- Canales, J.; Uribe, F.; Henríquez-Valencia, C.; Lovazzano, C.; Medina, J.; Vidal, E.A. Transcriptomic analysis at organ and time scale reveals gene regulatory networks controlling the sulfate starvation response of Solanum lycopersicum. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Wang, R.Y.; Liu, L.H.; Zhao, F.J.; Huang, X.Y. Local and Systemic Response to Heterogeneous Sulfate Resupply after Sulfur Deficiency in Rice. Int. J. Mol. Sci. 2022, 23, 6203. [Google Scholar] [CrossRef]
- Henriet, C.; Balliau, T.; Aimé, D.; Signor, C.L.; Kreplak, J.; Zivy, M.; Gallardo, K.; Vernoud, V. Proteomics of developing pea seeds reveals a complex antioxidant network underlying the response to sulfur deficiency and water stress. J. Exp. Bot. 2021, 72, 2611–2626. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Pan, Y.B.; Su, Y.; Zou, W.; Xu, F.; Sun, T.; Grisham, M.P.; Yang, S.; Xu, L.; Que, Y. WGCNA Identifies a Comprehensive and Dynamic Gene Co-Expression Network That Associates with Smut Resistance in Sugarcane. Int. J. Mol. Sci. 2022, 23, 10770. [Google Scholar] [CrossRef]
- Tandayu, E.; Borpatragohain, P.; Mauleon, R.; Kretzschmar, T. Genome-Wide Association Reveals Trait Loci for Seed Glucosinolate Accumulation in Indian Mustard (Brassica juncea L.). Plants 2022, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Martínez-Ballesta, M.; Moreno, D.; Carvajal, M. The Physiological Importance of Glucosinolates on Plant Response to Abiotic Stress in Brassica. Int. J. Mol. Sci. 2013, 14, 11607–11625. [Google Scholar] [CrossRef] [PubMed]
- Frerigmann, H.; Berger, B.; Gigolashvili, T. bHLH05 Is an Interaction Partner of MYB51 and a Novel Regulator of Glucosinolate Biosynthesis in Arabidopsis. Plant Physiol. 2014, 166, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tian, Y.; Shi, W.; Yu, P.; Hu, Y.; Lv, J.; Fu, C.; Fan, M.; Bai, M.Y. The miR396-GRFs Module Mediates the Prevention of Photo-oxidative Damage by Brassinosteroids during Seedling De-Etiolation in Arabidopsis. Plant Cell 2020, 32, 2525–2542. [Google Scholar] [CrossRef] [PubMed]
- Fristedt, R.; Hu, C.; Wheatley, N.; Roy, L.; Wachter, R.; Savage, L.; Harbinson, J.; Kramer, D.; Merchant, S.; Yeates, T.; et al. RAF2 is a RuBisCO assembly factor in Arabidopsis thaliana. Plant J. 2018, 94, 146–156. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, F.; Wu, Y.; Lou, L.; Liu, L.; Tian, M.; Ning, Y.; Shu, K.; Tang, S.; Xie, Q. The RING finger ubiquitin E3 ligase SDIR1 targets SDIR1-INTERACTING PROTEIN1 for degradation to modulate the salt stress response and ABA signaling in Arabidopsis. Plant Cell 2015, 27, 214–227. [Google Scholar] [CrossRef]
- Kim, S.; Guo, L.; Wang, X. Nuclear moonlighting of cytosolic glyceraldehyde-3-phosphate dehydrogenase regulates Arabidopsis response to heat stress. Nat. Commun. 2020, 11, 3439. [Google Scholar] [CrossRef]
- Schneider, M.; Knuesting, J.; Birkholz, O.; Heinisch, J.; Scheibe, R. Cytosolic GAPDH as a redox-dependent regulator of energy metabolism. BMC Plant Biol. 2018, 18, 184. [Google Scholar] [CrossRef]
- Tarnowski, L.; Rodriguez, M.; Brzywczy, J.; Piecho-Kabacik, M.; Krčkova, Z.; Martinec, J.; Wawrzynska, A.; Sirko, A. A selective autophagy cargo receptor NBR1 modulates abscisic acid signalling in Arabidopsis thaliana. Sci. Rep. 2020, 10, 7778. [Google Scholar] [CrossRef] [PubMed]
- Zientara-Rytter, K.; Łukomska, J.; Moniuszko, G.; Gwozdecki, R.; Surowiecki, P.; Lewandowska, M.; Liszewska, F.; Wawrzyńska, A.; Sirko, A. Identification and functional analysis of Joka2 a tobacco member of the family of selective autophagy cargo receptors. Autophagy 2011, 7, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shang, J.X.; Chen, Q.X.; Oses-Prieto, J.A.; Bai, M.Y.; Yang, Y.; Yuan, M.; Zhang, Y.L.; Mu, C.C.; Deng, Z.; et al. Identification of BZR1-interacting Proteins as Potential Components of the Brassinosteroid Signaling Pathway in Arabidopsis through Tandem Affinity Purification. Mol. Cell. Proteom. 2013, 12, 3653–3665. [Google Scholar] [CrossRef]
- Joseph, M.; Papdi, C.; Kozma-Bognár, L.; Nagy, I.; López-Carbonell, M.; Rigó, G.; Koncz, C.; Szabados, L. The Arabidopsis ZINC FINGER PROTEIN3 Interferes with Abscisic Acid and Light Signaling in Seed Germination and Plant Development. Plant Physiol. 2014, 165, 1203–1220. [Google Scholar] [CrossRef]
- Liu, C.; Cheng, J.; Zhuang, Y.; Ye, L.; Li, Z.; Wang, Y.; Qi, M.; Xu, L.; Zhang, Y. Polycomb repressive complex 2 attenuates ABA-induced senescence in Arabidopsis. Plant J. 2019, 97, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Gobbato, E.; Kracher, B.; Qiu, J.; Bautor, J.; Parker, J. A core function of EDS1 with PAD4 is to protect the salicylic acid defense sector in Arabidopsis immunity. New Phytol. 2017, 213, 1802–1817. [Google Scholar] [CrossRef]
- Kong, X.; Li, C.; Zhang, F.; Yu, Q.; Gao, S.; Zhang, M.; Tian, H.; Zhang, J.; Yuan, X.; Ding, Z. Ethylene promotes cadmium-induced root growth inhibition through EIN3 controlled XTH33 LSU1 expression in Arabidopsis. Plant Cell Environ. 2018, 41, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Zhao, F.; Wang, Y.; Liu, J.; Zhuang, Y.; Ye, L.; Qi, M.; Cheng, J.; Zhang, Y. Plant Regulomics: A data-driven interface for retrieving upstream regulators from plant multi-omics data. Plant J. 2019, 101, 237–248. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2019, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Yang, R.; Liu, J.; Lin, Z.; Sun, W.; Wu, Z.; Hu, H.; Zhang, Y. ERF transcription factors involved in salt response in tomato. Plant Growth Regul. 2018, 84, 573–582. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Crop Species | Gene ID | Biological Function | Tissue | Evidence | References |
|---|---|---|---|---|---|
| Solanum lycopersicum | Solyc03g096760.1 | S-deficiency | Roots and leaves | RNA-seq, qPCR | [17,30] |
| Solyc03g096770.1 | S-deficiency | Roots and leaves | RNA-seq, qPCR | [17,30] | |
| Solyc03g096780.1 | S-deficiency | Roots and leaves | RNA-seq, qPCR | [17,30] | |
| Solyc06g072990.1 | S-deficiency | Roots and leaves | RNA-seq, qPCR | [17,30] | |
| Oryza sativa | Os02g0129800 | S-deficiency | Roots and shoots | RNA-seq | [31] |
| Os10g0509401 | S-deficiency | Roots and shoots | RNA-seq | [31] | |
| Os10g0509600 | S-deficiency | Roots and shoots | RNA-seq | [31] | |
| Triticum aestivum | TraesCS1D03G0456700 | S-deficiency | Roots and leaves | qPCR | [17] |
| Pisum sativum | Psat5g070520 | S-deficiency | Seeds | mRNA-protein network analysis | [32] |
| Saccharum spontaneum | S.offnewGene71819 | Fungus infection | Stems | RNA-seq and qPCR | [33] |
| Sspon.01G0026550-1A | Fungus infection | Stems | RNA-seq | [33] | |
| Sspon.01G0026550-2C | Fungus infection | Stems | RNA-seq | [33] | |
| Brassica juncea | BjuA033112 | Glucosinolate content | Seeds | GWAS | [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canales, J.; Arenas-M, A.; Medina, J.; Vidal, E.A. A Revised View of the LSU Gene Family: New Functions in Plant Stress Responses and Phytohormone Signaling. Int. J. Mol. Sci. 2023, 24, 2819. https://doi.org/10.3390/ijms24032819
Canales J, Arenas-M A, Medina J, Vidal EA. A Revised View of the LSU Gene Family: New Functions in Plant Stress Responses and Phytohormone Signaling. International Journal of Molecular Sciences. 2023; 24(3):2819. https://doi.org/10.3390/ijms24032819
Chicago/Turabian StyleCanales, Javier, Anita Arenas-M, Joaquín Medina, and Elena A. Vidal. 2023. "A Revised View of the LSU Gene Family: New Functions in Plant Stress Responses and Phytohormone Signaling" International Journal of Molecular Sciences 24, no. 3: 2819. https://doi.org/10.3390/ijms24032819
APA StyleCanales, J., Arenas-M, A., Medina, J., & Vidal, E. A. (2023). A Revised View of the LSU Gene Family: New Functions in Plant Stress Responses and Phytohormone Signaling. International Journal of Molecular Sciences, 24(3), 2819. https://doi.org/10.3390/ijms24032819